Introduction
Renal cell carcinoma (RCC) represents the most
common malignant tumor of the kidney. The overall incidence of RCC
increases significantly from the age of 50 to 70 years (1,2). With
high degree of heterogeneity, RCC exhibits poor prognosis because
of increased rate of metastasis and resistance to conventional
chemotherapy and radiotherapy (3).
Therefore, alternative novel therapeutic agents are needed for
effective treatment of RCC. Since the evasion from apoptosis is one
of the hallmarks of cancer, it has long been attempted to develop
anticancer drugs that can selectively induce apoptosis in cancer
cells (4). A wide variety of
natural products, especially phytochemicals, have been shown to
suppress cell growth, modulate cell differentiation and induce
cancer cell death (5).
Apoptosis, a physiological process of forcing
unwanted cells to commit suicide, is characterized by cell
shrinkage, plasma membrane blebbing, DNA fragmentation and the
formation of apoptotic bodies (6).
Biochemical mechanisms of apoptosis involve two different pathways,
one is the intrinsic or the mitochondria-mediated pathway, and the
other is the extrinsic or death receptor (DR)-mediated pathway
(7). The intrinsic pathway involves
disruption of mitochondrial membrane potential and the release of
cytochrome c from the mitochondrial inter-membrane space into the
cytosol, resulting in the activation of caspase-9, -7 and -3
through the proteolytic cleavage of respective pro-caspases
(8). The mitochondrial membrane
potential is regulated by proteins of the B cell lymphoma-2 (Bcl-2)
family (9). These include
anti-apoptotic Bcl-2 proteins (Bcl-2 and Bcl-xL), pro-apoptotic
multidomain protein (Bak and Bax), and BH-3-only pro-apoptotic
proteins (Bad, Bid and Bim) (10).
Alteration in cellular redox status shifts the balance between the
expression of anti-apoptotic and pro-apoptotic proteins, thereby
leading to cell fate decision (11). A critical determinant of cell fate
decision is the intracellular accumulation of reactive oxygen
species (ROS), which are generated as by-products of cellular
metabolism. High levels of ROS may lead to cell death through
mitochondrial collapse (12).
The extrinsic pathway, on the contrary, is activated
by death-inducing signal molecules of tumor necrosis factor (TNF)
family, such as FasL, TNF and TNF-related apoptosis inducing ligand
(TRAIL) (13), which bind to
various plasma membrane-bound DRs (14). Upon stimulation of an appropriate
apoptotic signal, the activation of DRs leads to the formation of
the death-inducing signaling complex (DISC) and the activation of
caspase-8 and -10, which in turn activates the rest of downstream
caspases, such as caspase-3, thereby inducing apoptosis (15). A number of plant polyphenols have
been reported to induce apoptosis via the intrinsic or extrinsic
pathway in various cancer cells (16).
Signal transducer and activator of transcription-3
(STAT3) plays a decisive role in regulating cell growth, survival,
angiogenesis and immune escape of tumor cells (17). The blockade of STAT3 signaling
caused induction of apoptosis, inhibition of cell proliferation,
suppression of angiogenesis and stimulation of immune responses
(18–21). In cancer cells, STAT3 becomes
constitutively active through the phosphorylation by upstream Src
family kinases or Janus-activated kinases (Jaks) (19). Since the STAT3-regulated gene
products, such as Bcl-xL, Bcl-2, survivin, c-Myc and D-series of
cyclins are involved in enhanced cell proliferation, selective
inhibition of STAT3 signaling can suppress proliferation and induce
apoptosis in cancer cells (19).
Previous studies have shown that suppression of STAT3 signaling
inhibits the growth of various cancer cells including those of the
stomach, liver, head and neck, skin, and lung (22). STAT3, a signal mediator of various
pro-inflammatory cytokines and growth factors acting
consti-tutively in an inflammatory tumor microenvironment, is an
important molecular target of various anticancer drugs (23).
Rosmarinus officinalis L., generally known as
rosemary, has a long-standing repution for improving memory and has
been used as a symbol of reminiscence in Europe (24). In addition, leaves of the plants
have been used as a means to weight loss due to its ability to
inhibit lipid absorption activities in the digestive system
(25). Major bioactive components
of the plant include polyphenolics, such as carnosic acid (CA),
carnosol, rosmarinic acid and ursolic acid (26–29).
CA (Fig. 1A) has a wide range of
biological activities, including antioxidant, anticancer,
anti-inflammatory, anti-adipogenic and neuroprotective effects
(30–34). The present study investigated the
underlying molecular mechanisms of anticancer effects of CA in
human renal carcinoma (Caki) cells. We found that CA induced
anti-proliferative and apoptotic effects in Caki cells through
mitochondria-dependent caspase activation and the interference with
STAT3 signaling pathway via the generation of ROS.
Materials and methods
Materials
CA (purity 99%) and N-acetyl cysteine (NAC)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies
against cleaved caspase-9, -7, -3, poly(ADP-ribose) polymerase
(PARP), Bcl-2, Bcl-xL, Bax, cytochrome c, STAT3, p-STAT3 (Y705),
p-STAT3 (S727), Src, p-Src, cyclin D1, D2, and D3 and survivin were
from Cell Signaling Technology Inc. (Beverly, MA, USA). Antibody
against each of p53, murine double minute-2 (Mdm2), p27, and
horse-raddish peroxidase-conjugated secondary antibodies were from
Santa Cruz Biotechnology (Paso Robles, CA, USA). β-Actin antibody
was obtained from Sigma Chemical Co. (St. Louis, MO, USA). The
2′-7′ dichlorofluorescin diacetate (DCF-DA) was from Invitrogen
(Carlsbad, CA, USA). Hank's balanced salt solution (HBSS) was from
the Meditech (Herndon, VA, USA).
Cell culture and treatment
Caki cells were obtained from Dr T.K. Kwon (Keimyung
University, Korea) and maintained in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum and
antibiotics (100 U/ml penicillin G and 100 mg/ml streptomycin) at
37°C in a humidified incubator containing 5% CO2 and 95%
air. In all the experiments, cells were seeded at 1×105
cells/well and incubated with CA at 50–60% confluence. All
chemicals were dissolved in ethanol and the final ethanol
concentration was <0.1%.
Cell viability assay
The cell growth effect was measured by the MTT
assay. Cells (2×103) were incubated in triplicate in a
96-well plate in presence or absence of CA in a final volume of 100
µl for different time intervals at 37°C. Thereafter, 10
µl of MTT solution (5 mg/ml) was added to each well and
incubated for 4 h. Medium was removed, formazan was dissolved in
DMSO and absorbance at 550 nm was measured by using microplate
reader (Tecan Trading AG, Männedorf, Switzerland). Cell viability
was described as the relative percentage of control.
Annexin V staining
Annexin V staining was performed using fluorescein
isothiocyanate (FITC)-Annexin V staining kit (BD Biosciences, San
Jose, CA, USA) following the manufacturer's instructions. Briefly,
CA-treated cells were washed with PBS and resuspended in binding
buffer containing Annexin V and propidium iodide (PI). Flourescence
intensity was measured using flow cytometry (BD Biosciences).
Western blot analysis
Cells were harvested and lysed with RIPA buffer, and
the resulting protein samples were quantified by using
bicinchoninic acid protein assay kit (Pierce Biotechnology,
Rockford, IL, USA). Equal amount of protein extracts were denatured
by boiling at 100°C for 5 min in sample buffer. The proteins were
separated on 8–12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred to polyvinylidene
difluoride membrane. The membranes were blocked with 5% skim milk
in Tris-buffered saline with Tween-20 buffer (TBS-T) (10 mM Tris,
150 mM NaCl, pH 7.5 and 0.1% Tween-20) for 1 h at room temperature.
The membranes were washed 3 times for 10 min each with TBS-T buffer
and incubated for 1 h with horseradish peroxidase-conjugated
secondary antibodies. The membranes were washed 3 times for 10 min
each with TBS-T buffer. Immunoblot membranes were incubated with
Super-signal pico-chemiluminescent substrate or dura-luminol
substrate (Thermo Scientific, Waltham, MA, USA) according to
manufacturer's instruction and visualized with imagequant™ LAS 4000
(Fujifilm Life Science, Tokyo, Japan).
Measurement of reactive oxygen species
(ROS) accumulation
Cells were treated with CA in the presence or
absence of NAC for 24 h and then loaded with 25 µM of
2′7′-dichlorofluorescin diacetate (DCF-DA). After incubation for 30
min at 37°C in a 5% CO2 incubator, cells were washed
twice with HBSS solution, suspended in the complete media and were
examined under a fluorescence microscope to detect the
intracellular ROS. Fluorescence of oxidized DCF was also measured
at an excitation wave length of 480 nm and emission wavelength of
525 nm using flow cytometry.
Electrophoretic mobility gel shift assay
(EMSA)
The nuclear extract was prepared from cells
incubated with or without CA. The STAT3 oligonucleotide probe
5′-AGC TTC ATT TCC CGT AAA TCC CTA-3′ (Biomedic, Korea) was labeled
with [γ-32P]-ATP using T4 polynucleotide kinase. The
EMSA was performed according to the protocol described earlier
(35).
Statistical analysis
When necessary, data were expressed as mean ± SD of
at least three independent experiments, and statistical analysis
for single comparison was performed using the Student's t-test and
p-value <0.05 was considered to indicate a statistically
significant difference.
Results
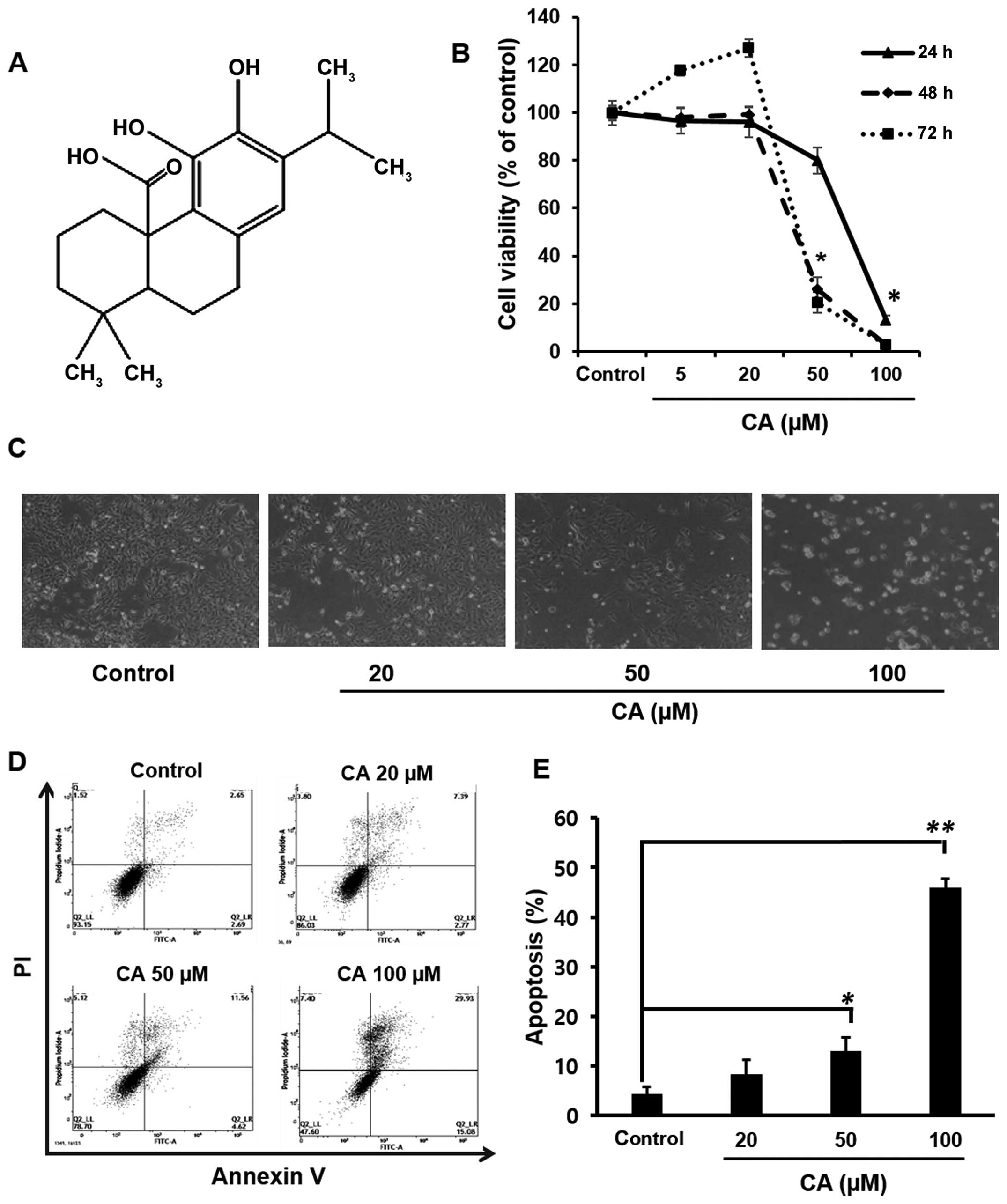
CA induces apoptosis in Caki cells
In the present study, the effect of CA on the
viability of Caki cells was first examined by the MTT assay. As
shown in Fig. 1B, treatment of Caki
cells with CA significantly reduced the cell viability in a
concentration- and time-dependent manner in comparison to the
untreated cells. Fig. 1C showed the
morphological changes of Caki cells upon incubation with varying
concentrations of CA. At the highest concentration, the cell
viability was inhibited approximately by 80% as compared to
untreated cells after 24 h. Based on this result, cells were
incubated with CA at a range of concentrations (20–100 µM)
for a period of 24 h in subsequent experiments. To elucidate
whether CA-mediated cytotoxicity could result from the induction of
apoptosis, FACS analysis was performed. Caki cells treated with CA
(20, 50 or 100 µM) for 24 h were analyzed by flow cytometry
using double staining with Annexin V and PI to quantify the
population of cells undergoing apoptosis (Annexin
V+/PI−). The results showed that the
treatment of cells with CA markedly increased the percentages of
apoptotic cells as compared with untreated control cells (Fig. 1D). Quantification of apoptotic cells
and statistical analysis of CA-induced apoptosis are presented in
Fig. 1E.
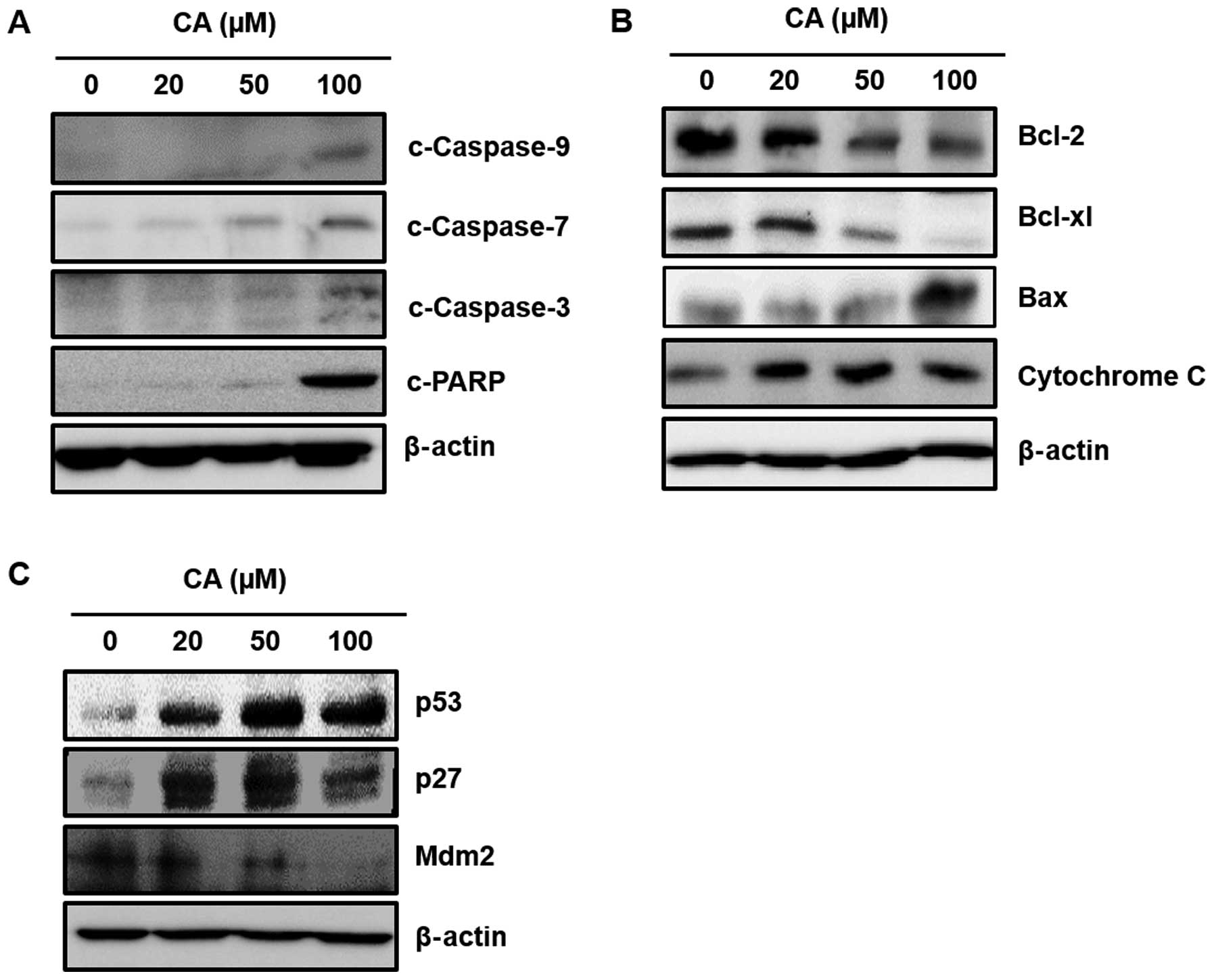
CA induces apoptosis through the
mitochondrial pathway in Caki cells
Caspases are crucial components of the apoptosis
pathway. The mitochondrial and caspase pathways are interconnected.
Release of cytochrome c into the cytosol activates the caspase
adaptor Apaf-1 and procaspase-9, which activates executioner
caspases (caspase-3, -6 and -7) (36). In the present study, incubation of
Caki cells with CA (20, 50 or 100 µM) induced activation of
caspase-9, -7 and -3 and the cleavage of PARP (Fig. 2A), indicating the involvement of
mitochondria in CA-induced apoptosis. Since Bcl-2 family proteins
regulate the mitochondrial membrane integrity, the effect of CA on
the expression of Bcl-2 family proteins was then examined. The
present study revealed that CA not only downregulated the
expression of anti-apoptotic protein Bcl-2 and Bcl-xL, but also
increased the expression of apoptosis inducing protein Bax, thereby
releasing cytochrome c into the cytosol (Fig. 2B). As shown in Fig. 2C, incubation of cells with CA
increased expression of p53 and diminished protein level of its
cytosolic repressor protein Mdm2 in a concentration-dependent
manner. p53 induces the cell cycle arrest through transcriptional
activation of its cell cycle regulatory gene product, p27. We
examined whether CA treatment has an effect on p27 expression in
Caki cells. As a result, expression of p27 is increased by CA
treatment in according to p53 protein levels.
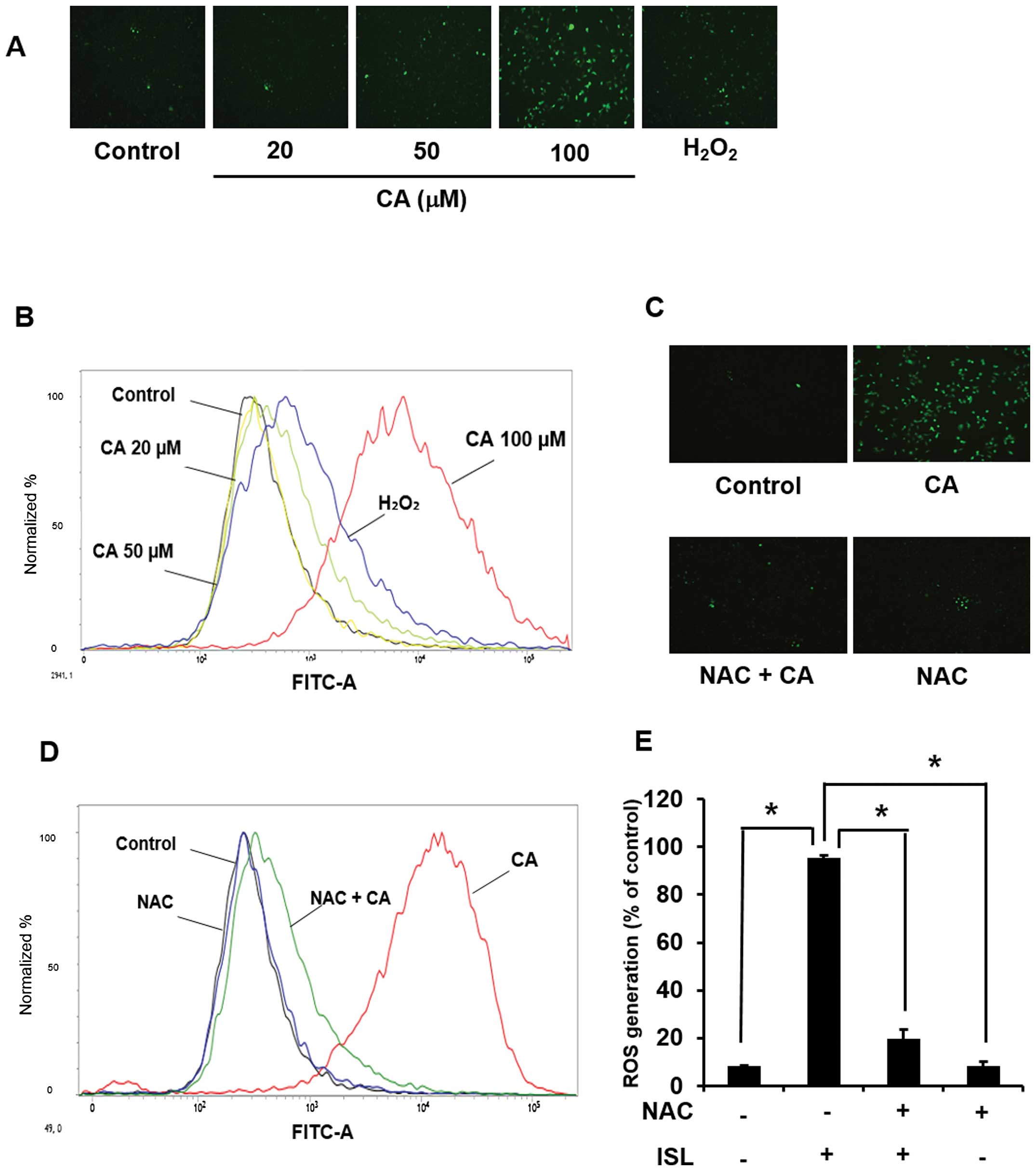
Involvement of ROS in CA-induced
apoptosis in Caki cells
Since the accumulation of intracellular ROS can
induce cell death, the effect of CA on ROS generation was examined.
Treatment of cells with CA (20, 50, 100 µM) for 24 h
generated ROS as revealed by the immunofluorescence analysis after
DCF-DA staining (Fig. 3A) as well
as by the FACS analysis (Fig. 3B).
Pretreatment of cells with NAC abolished CA-induced ROS
accumulation (Fig. 3C and D).
Quantification and statistical analysis of CA-induced ROS
generation are presented in Fig.
3E.
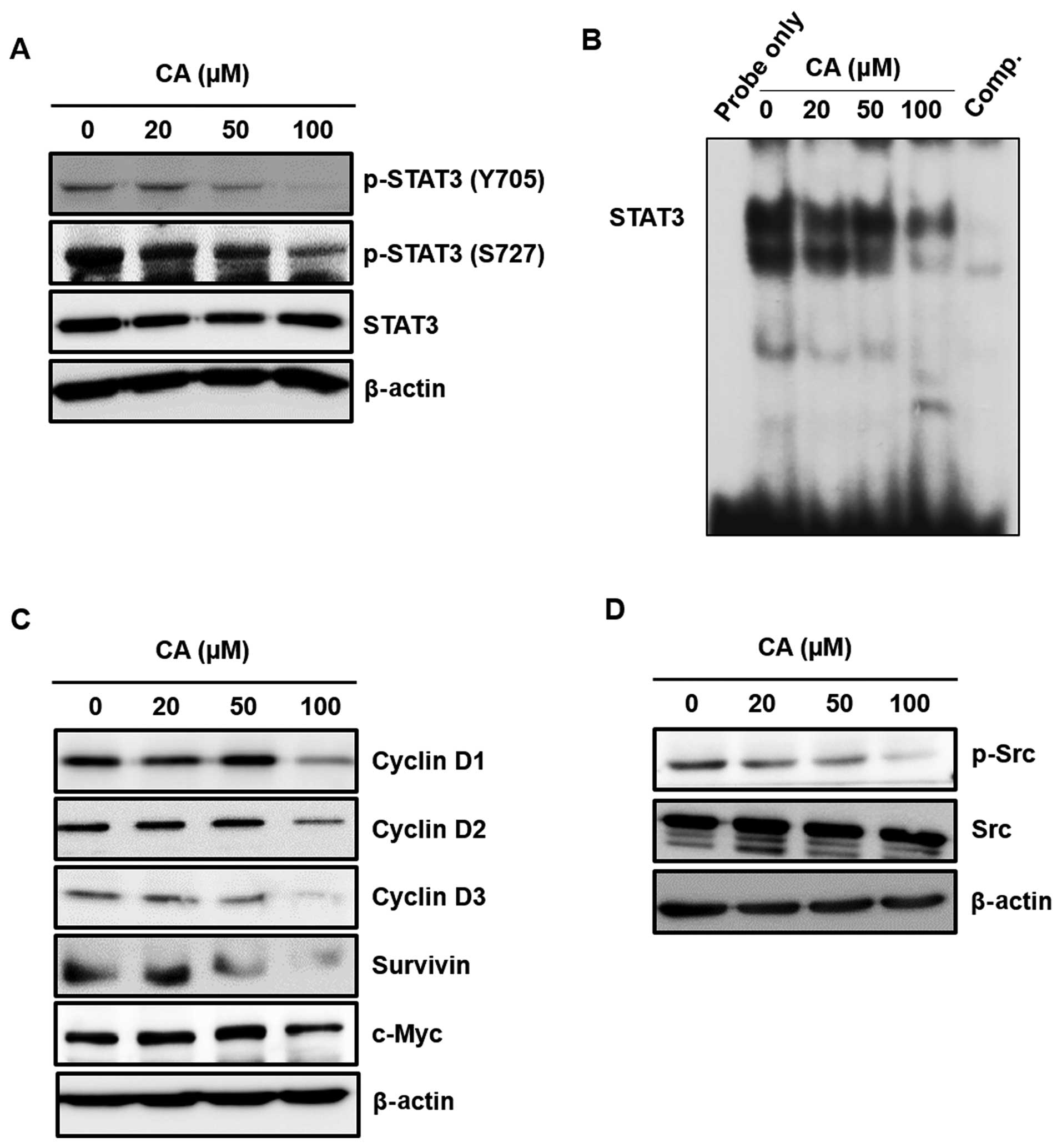
CA attenuates the activation of STAT3
signaling pathway
Since STAT3 plays a key role in cell proliferation
through transcriptional activation of pro-survival genes, we
examined the effect of CA on the expression of cell proliferation
markers, which are transcriptionally regulated by STAT3. The
present study showed that CA diminished the constitutive
phosphorylation at both Y705 and S727 residues of STAT3 (Fig. 4A). CA also reduced the constitutive
STAT3 DNA-binding activity (Fig.
4B). In addition, CA treatment suppressed the expression of
STAT3-regulated cell proliferative gene products, such as c-Myc,
survivin and D-series of cyclins (Fig.
4C). STAT3 is known to be phosphorylated by upstream kinase,
such as Src. Since CA was found to suppress STAT3 activation, the
effect of CA on the activation of upstream signaling proteins of
STAT3 were checked. As shown in Fig.
4D, CA treatment resulted in the suppression of Src
phosphorylation in concentration-dependent manner in Caki
cells.
Role of ROS in CA-induced inhibition of
STAT3 signaling and induction of apoptosis in Caki cells
We next examined the role of CA-induced ROS
generation in blocking STAT3 signaling and induction of apoptosis
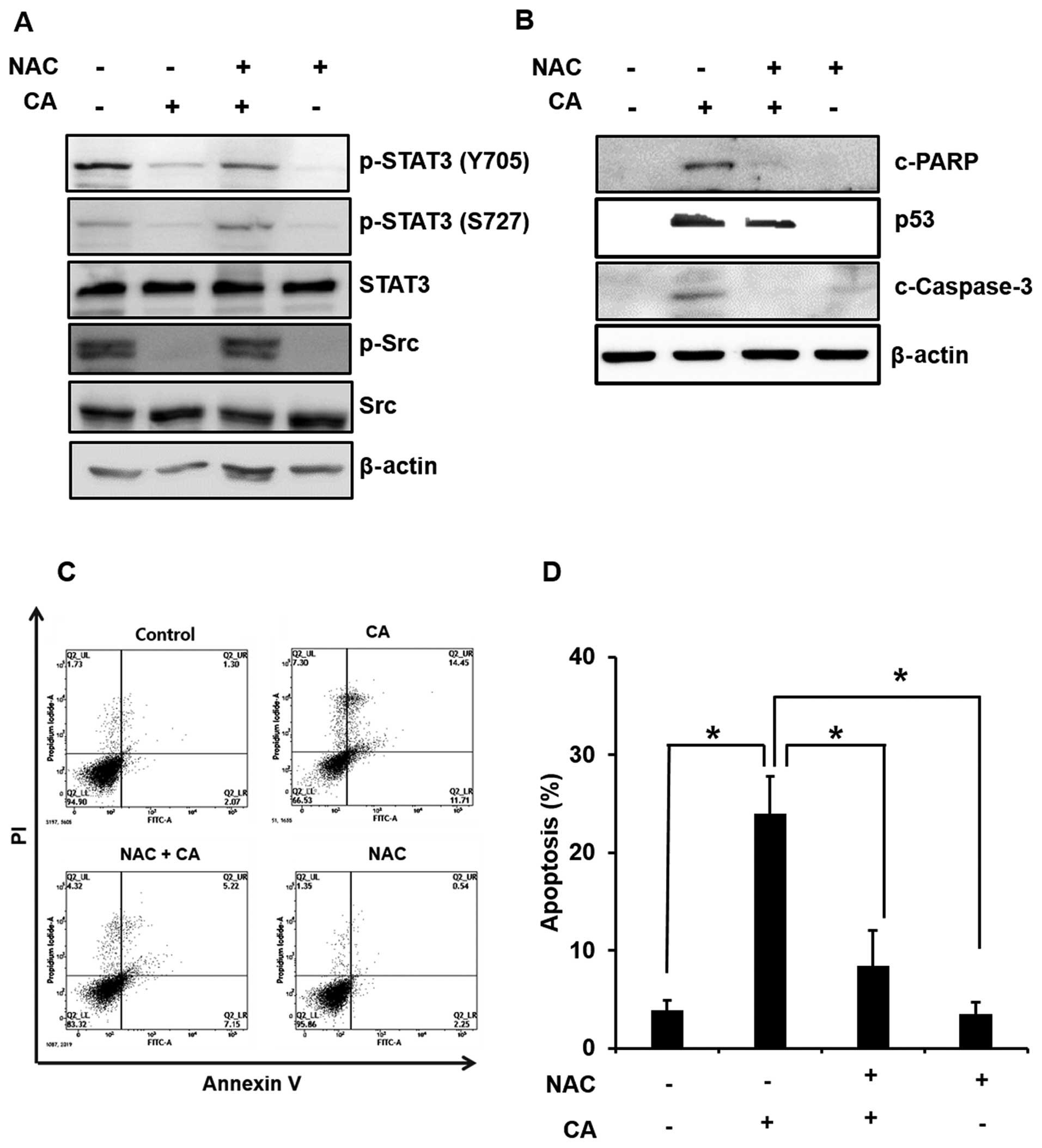
in Caki cells. Treatment with CA in the absence or presence of NAC
revealed that inhibition of ROS generation abrogated the inhibitory
effect of CA on the phosphorylation of STAT3 and Src (Fig. 5A). To investigate the possible
mechanism underlying the induction of apoptosis by CA via ROS
generation, we also examined the cleavage of caspase-3 and PARP and
expression of p53. As shown Fig. 5B
and 5C, activation of p53, the
cleavage of PARP and apoptosis induction through ROS generation was
abolished by pretreatment of NAC in CA-treated Caki cells. These
findings suggest that ROS play critical roles in CA-induced
apoptosis in Caki cells.
Discussion
Cancer is a leading cause of death worldwide
(37). Benign tumor is primarily
removed by surgery and the residual cancer cells are eradicated by
chemotherapy and/or radiotherapy. However, the chemotherapy
regimens frequently cause various side effects, such as
considerable death of healthy cells and drug tolerance. Thus,
agents that specifically target tumor cells are ideal for cancer
therapies. Based on this requirement, natural therapies, which take
advantage of the abundant and vast resources of natural compounds
derived from medicinal plants, are being developed to overcome the
side effects of chemotherapeutic agents. The present study
investigated the effect of CA, a major ingredient of medicinal herb
Rosmarinus officinalis L., against human renal cancer cells
and elucidated its mechanism of action.
Apoptosis is typically composed of two main
pathways: the extrinsic pathway that mediates signals via death
receptors, and the intrinsic pathway that involves mitochondrial
dysfunction (38). The extrinsic
pathway is provoked through the binding of cytokine ligands, such
as FasL, TNF and TRAIL to the plasma membrane receptors (14). The interaction between ligands and
receptors result in the formation of DISCs, which activate
caspase-8 and release the DISC into the cytoplasm (39). Caspase-8 can activate caspase-3
directly or via release of cytochrome c mediated by mitochondria.
The second pathway is the intrinsic pathway, which was triggered
with a variety of stress stimuli, such as DNA damage and the
actions of some oncoproteins (40).
Signaling of this pathway is accompanied by the alteration in the
expression of Bcl-2 family proteins, decreased mitochondrial
membrane potentials, and the release of cytochrome c from the
mitochondria to cytoplasm (41).
The release of cytochrome c and regulation of Bcl-2 family proteins
are critical processes in the mitochondria-mediated apoptosis
(41,42). In addition, the Bcl-2 family
proteins play a central function in inducing apoptosis and involve
members with either pro- or anti-apoptotic activity (43).
Our study demonstrated that cleaved PARP and
procas-pase-9, -7 and -3, accompanied by decrease in the expression
of anti-apoptotic proteins, such as Bcl-xL and Bcl-2, increase the
expression of pro-apoptotic protein Bax in CA-treated Caki cells.
These findings indicated that CA-induced apoptosis could partly be
mediated through mitochondria-mediated apoptotic pathway.
Cross-talk between the death-receptor and mitochondria-mediated
pathway is achieved by Bid protein, which is altered by caspase-8.
The death signal induced in the extrinsic pathway, for example, via
DR4 and DR5, can lead to the disruption of mitochondrial membrane
integrity (8,44). The signal cascade is associated with
the ability of caspase-8 to proteolytically cleave Bid into
truncated Bid (tBid). The pro-apoptotic protein tBid, a member of
the Bcl-2 family, transfers to the mitochondria where it allows
other proteins of the same family to combine with the outer
mitochondrial membrane to release cytochrome c. We found that
incubation of cells with CA upregulated the expression of DR4 and
DR5 and increased the proteolytic cleavage of caspase-8 in Caki
cells (data not shown). Taken together, the induction of apoptosis
by CA involves both the extrinsic and the intrinsic pathways.
Tumor suppressor p53 is another important factor
that affects the cellular response to drugs, while exert effects on
growth inhibition and apoptosis induction (45). The p53 is sequestered in cytoplasm
by binding with its inhibitory protein Mdm2, which possesses
ubiquitin ligase property and plays an important role in p53
turnover. Thus, a decrease in Mdm2 level leads to stabilization of
p53. In response to DNA damage, p53 is phosphorylated, dissociated
from Mdm2 and translo-cated to the nucleus. Within the nucleus,
phosphorylated p53 functions as a transcription factor that
regulates transcriptional activation of genes involved in cell
cycle regulation and apoptosis (46,47).
The p53-dependent apoptosis is associated with the caspase cleavage
and the expression of pro-apoptotic proteins, such as Bax (48). Moreover, p53 induces the cell cycle
arrest through transcriptional activation of its several cell cycle
regulatory gene products, such as p21 and p27. Thus, inhibiting
Mdm2 expression and concomitantly elevating expression of p53.
Taken together, these findings suggest that CA may cause apoptosis
in a p53-dependent manner.
Since the activation of p53 occurs in response to
oxidative stress (49), the effect
of CA on ROS generation in Caki cells was examined. ROS, including
free radicals such as super-oxide, hydroxyl radicals
(•OH), and the non-radical H2O2
are generated through multiple sources in the cells, such as the
electron transport chain in mitochondria, ionizing radiations and
enzymes (e.g., phagocytic and non-phagocytic NADPH oxidases,
lipoxygenases and cycloxygenases) producing superoxide anions.
Moreover, ROS is known to instigate the apoptotic cascade by
causing damage to many cellular components including proteins,
lipids, nucleic acid, and the mitochondria itself, which in turn
facilitates the release of apoptogenic factors (50,51).
The present study showed that incubation with CA significantly
increased the ROS level, which was scavenged by pretreatment of
cells with NAC. Furthermore, NAC treatment rescued cells from
CA-induced apoptosis by blocking the cleavage of PARP and the
expression of p53. Thus, CA-induced ROS generation may activate
p53, resulting in the downregulation of Bcl-2 and induction of Bax
expression, thereby leading to the activation of caspases and
induction of apoptosis.
Constitutive STAT3 activation has been related to a
variety of tumors including breast, prostate and colon cancer
(52). Recent studies have
indicated that the STAT3 signaling pathway is closely associated
with cell proliferation, differentiation and evasion of apoptosis
(53,54). The activation of STAT3 signaling
involves phosphorylation of Y705 and S727 residues followed by
dimerization and nuclear translocation, and subsequently DNA
binding for the transcriptional activation of its target genes
(55). Since CA abolished the
expression of Bcl-2 and Bcl-xL, which are regulated by STAT3, the
effect of CA on the activation of STAT3 signaling in Caki cells was
examined. The present study revealed that CA blunted the
constitutive phosphorylation of STAT3 and reduced the expression of
its target genes D-series of cyclins, survivin and c-Myc.
Intracellular kinases, such as Src, have been shown to
phosphorylate STAT3. Thus, the inhibitory effect of CA on the
phosphorylation of Src suggests that the compound interferes with
STAT3 signaling pathway.
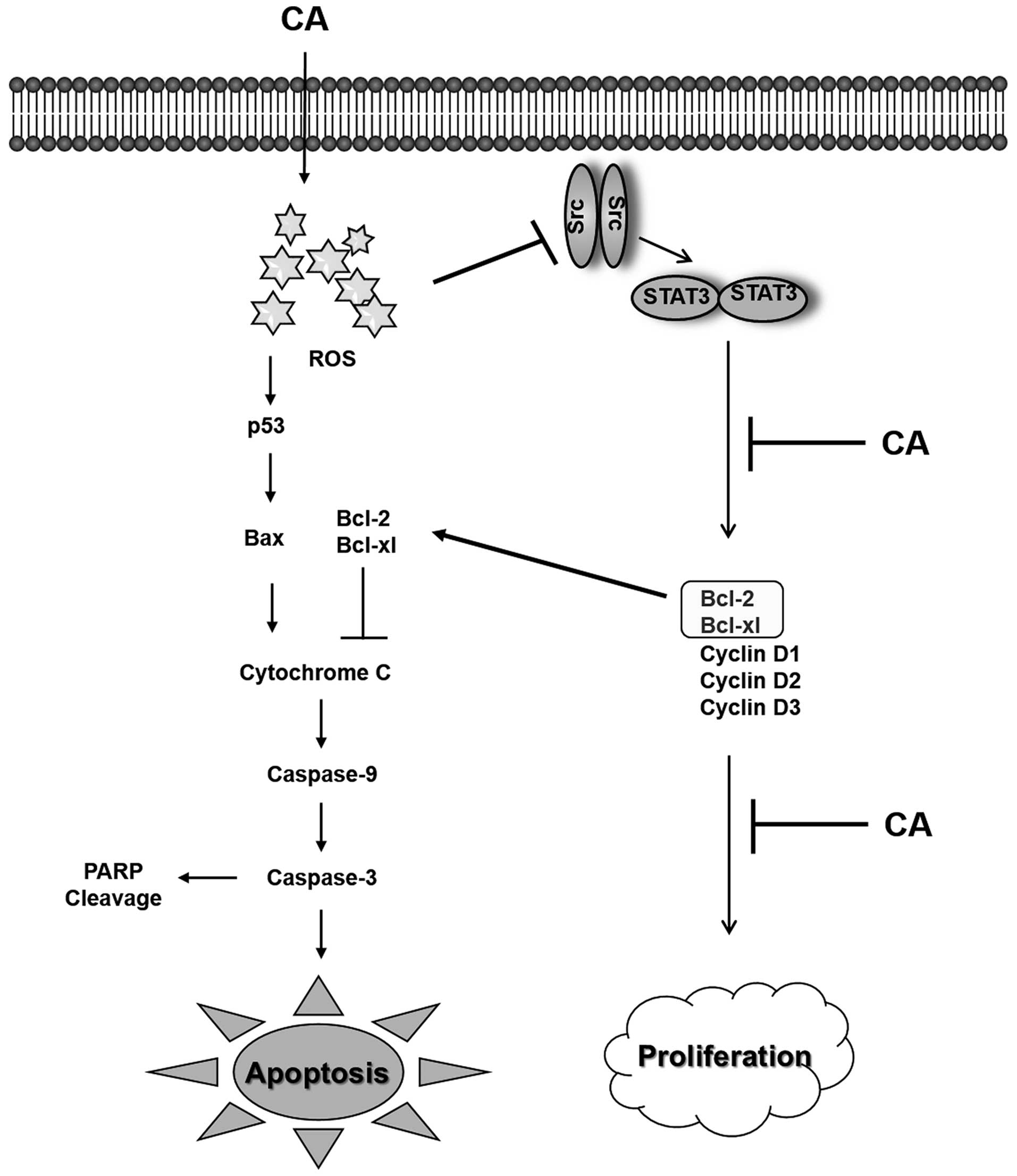
In conclusion, the present study demonstrates for
the first time that CA induces both the extrinsic and intrinsic
pathways of apoptosis in Caki cells through the generation of ROS
and inactivation of STAT3 signaling (Fig. 6).
Acknowledgments
This study was supported by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Science, ICT and Future Planning
(2014R1A2A2A01004698).
References
|
1
|
Motzer RJ, Bander NH and Nanus DM:
Renal-cell carcinoma. N Engl J Med. 335:865–875. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mulders P, Figlin R, deKernion JB,
Wiltrout R, Linehan M, Parkinson D, deWolf W and Belldegrun A:
Renal cell carcinoma: Recent progress and future directions. Cancer
Res. 57:5189–5195. 1997.PubMed/NCBI
|
|
3
|
Santoni M, De Tursi M, Felici A, Lo Re G,
Ricotta R, Ruggeri EM, Sabbatini R, Santini D, Vaccaro V and
Milella M: Management of metastatic renal cell carcinoma patients
with poor-risk features: Current status and future perspectives.
Expert Rev Anticancer Ther. 13:697–709. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang MT, Ho CT, Wang ZY, Ferraro T, Lou
YR, Stauber K, Ma W, Georgiadis C, Laskin JD and Conney AH:
Inhibition of skin tumorigenesis by rosemary and its constituents
carnosol and ursolic acid. Cancer Res. 54:701–708. 1994.PubMed/NCBI
|
|
5
|
Chen D and Dou QP: Tea polyphenols and
their roles in cancer prevention and chemotherapy. Int J Mol Sci.
9:1196–1206. 2008. View Article : Google Scholar
|
|
6
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Earnshaw WC, Martins LM and Kaufmann SH:
Mammalian caspases: Structure, activation, substrates, and
functions during apoptosis. Annu Rev Biochem. 68:383–424. 1999.
View Article : Google Scholar
|
|
8
|
Fulda S, Galluzzi L and Kroemer G:
Targeting mitochondria for cancer therapy. Nat Rev Drug Discov.
9:447–464. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Panaretakis T, Pokrovskaja K, Shoshan MC
and Grandér D: Interferon-alpha-induced apoptosis in U266 cells is
associated with activation of the proapoptotic Bcl-2 family members
Bak and Bax. Oncogene. 22:4543–4556. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar
|
|
11
|
Trachootham D, Lu W, Ogasawara MA, Nilsa
RD and Huang P: Redox regulation of cell survival. Antioxid Redox
Signal. 10:1343–1374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Akgul C, Moulding DA and Edwards SW:
Molecular control of neutrophil apoptosis. FEBS Lett. 487:318–322.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang YJ, Kim IY, Kim EH, Yoon MJ, Kim SU,
Kwon TK and Choi KS: Paxilline enhances TRAIL-mediated apoptosis of
glioma cells via modulation of c-FLIP, survivin and DR5. Exp Mol
Med. 43:24–34. 2011. View Article : Google Scholar :
|
|
14
|
Thorburn A: Death receptor-induced cell
killing. Cell Signal. 16:139–144. 2004. View Article : Google Scholar
|
|
15
|
Ashkenazi A: Targeting the extrinsic
apoptosis pathway in cancer. Cytokine Growth Factor Rev.
19:325–331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Neergheen VS, Bahorun T, Taylor EW, Jen LS
and Aruoma OI: Targeting specific cell signaling transduction
pathways by dietary and medicinal phytochemicals in cancer
chemoprevention. Toxicology. 278:229–241. 2010. View Article : Google Scholar
|
|
17
|
Huang S: Regulation of metastases by
signal transducer and activator of transcription 3 signaling
pathway: Clinical implications. Clin Cancer Res. 13:1362–1366.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang T, Niu G, Kortylewski M, Burdelya L,
Shain K, Zhang S, Bhattacharya R, Gabrilovich D, Heller R, Coppola
D, et al: Regulation of the innate and adaptive immune responses by
Stat-3 signaling in tumor cells. Nat Med. 10:48–54. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Ma X, Yan S, Shen S, Zhu H, Gu Y,
Wang H, Qin G and Yu Q: 17-hydroxy-jolkinolide B inhibits signal
transducers and activators of transcription 3 signaling by
covalently cross-linking Janus kinases and induces apoptosis of
human cancer cells. Cancer Res. 69:7302–7310. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu H and Jove R: The STATs of cancer - new
molecular targets come of age. Nat Rev Cancer. 4:97–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen YF, Yang JS, Chang WS, Tsai SC, Peng
SF and Zhou YR: Houttuynia cordata Thunb extract modulates G0/G1
arrest and Fas/CD95-mediated death receptor apoptotic cell death in
human lung cancer A549 cells. J Biomed Sci. 20:182013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim HG, Hwang YP and Jeong HG: Kahweol
blocks STAT3 phosphorylation and induces apoptosis in human lung
adenocarcinoma A549 cells. Toxicol Lett. 187:28–34. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jalali-Heravi M, Moazeni RS and Sereshti
H: Analysis of Iranian rosemary essential oil: Application of gas
chromatography-mass spectrometry combined with chemometrics. J
Chromatogr A. 1218:2569–2576. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ninomiya K, Matsuda H, Shimoda H, Nishida
N, Kasajima N, Yoshino T, Morikawa T and Yoshikawa M: Carnosic
acid, a new class of lipid absorption inhibitor from sage. Bioorg
Med Chem Lett. 14:1943–1946. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Almela L, Sánchez-Muñoz B, Fernández-López
JA, Roca MJ and Rabe V: Liquid chromatograpic-mass spectrometric
analysis of phenolics and free radical scavenging activity of
rosemary extract from different raw material. J Chromatogr A.
1120:221–229. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ramírez P, García-Risco MR, Santoyo S,
Señoráns FJ, Ibáñez E and Reglero G: Isolation of functional
ingredients from rosemary by preparative-supercritical fluid
chromatography (Prep-SFC). J Pharm Biomed Anal. 41:1606–1613. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
López-García S, Castañeda-Sanchez JI,
Jiménez-Arellanes A, Domínguez-López L, Castro-Mussot ME,
Hernández-Sanchéz J and Luna-Herrera J: Macrophage activation by
ursolic and oleanolic acids during mycobacterial infection.
Molecules. 20:14348–14364. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ortuño J, Serrano R and Bañón S:
Antioxidant and antimicrobial effects of dietary supplementation
with rosemary diterpenes (carnosic acid and carnosol) vs vitamin E
on lamb meat packed under protective atmosphere. Meat Sci.
110:62–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gaya M, Repetto V, Toneatto J, Anesini C,
Piwien-Pilipuk G and Moreno S: Antiadipogenic effect of carnosic
acid, a natural compound present in Rosmarinus officinalis, is
exerted through the C/EBPs and PPARγ pathways at the onset of the
differentiation program. Biochim Biophys Acta. 1830:3796–3806.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ibarra A, Cases J, Roller M, Chiralt-Boix
A, Coussaert A and Ripoll C: Carnosic acid-rich rosemary
(Rosmarinus officinalis L.) leaf extract limits weight gain and
improves cholesterol levels and glycaemia in mice on a high-fat
diet. Br J Nutr. 106:1182–1189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tsai CW, Liu KL, Lin YR and Kuo WC: The
mechanisms of carnosic acid attenuates tumor necrosis
factor-α-mediated inflammation and insulin resistance in 3T3-L1
adipocytes. Mol Nutr Food Res. 58:654–664. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang T, Takikawa Y, Satoh T, Yoshioka Y,
Kosaka K, Tatemichi Y and Suzuki K: Carnosic acid prevents obesity
and hepatic steatosis in ob/ob mice. Hepatol Res. 41:87–92. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiang Q, Liu Z, Wang Y, Xiao H, Wu W, Xiao
C and Liu X: Carnosic acid attenuates lipopolysaccharide-induced
liver injury in rats via fortifying cellular antioxidant defense
system. Food Chem Toxicol. 53:1–9. 2013. View Article : Google Scholar
|
|
35
|
Kundu JK, Shin YK, Kim SH and Surh YJ:
Resveratrol inhibits phorbol ester-induced expression of COX-2 and
activation of NF-kappaB in mouse skin by blocking IkappaB kinase
activity. Carcinogenesis. 27:1465–1474. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Herr I and Debatin KM: Cellular stress
response and apoptosis in cancer therapy. Blood. 98:2603–2614.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ouyang L, Shi Z, Zhao S, Wang FT, Zhou TT,
Liu B and Bao JK: Programmed cell death pathways in cancer: A
review of apoptosis, autophagy and programmed necrosis. Cell
Prolif. 45:487–498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Prasad S, Yadav VR, Kannappan R and
Aggarwal BB: Ursolic acid, a pentacyclin triterpene, potentiates
TRAIL-induced apoptosis through p53-independent up-regulation of
death receptors: Evidence for the role of reactive oxygen species
and JNK. J Biol Chem. 286:5546–5557. 2011. View Article : Google Scholar :
|
|
40
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang X: The expanding role of mitochondria
in apoptosis. Genes Dev. 15:2922–2933. 2001.PubMed/NCBI
|
|
42
|
Garrido C, Galluzzi L, Brunet M, Puig PE,
Didelot C and Kroemer G: Mechanisms of cytochrome c release from
mitochondria. Cell Death Differ. 13:1423–1433. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Antonsson B and Martinou JC: The Bcl-2
protein family. Exp Cell Res. 256:50–57. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Han J, Goldstein LA, Gastman BR and
Rabinowich H: Interrelated roles for Mcl-1 and BIM in regulation of
TRAIL-mediated mitochondrial apoptosis. J Biol Chem.
281:10153–10163. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
O'Connor PM, Jackman J, Bae I, Myers TG,
Fan S, Mutoh M, Scudiero DA, Monks A, Sausville EA, Weinstein JN,
et al: Characterization of the p53 tumor suppressor pathway in cell
lines of the National Cancer Institute anticancer drug screen and
correlations with the growth-inhibitory potency of 123 anticancer
agents. Cancer Res. 57:4285–4300. 1997.PubMed/NCBI
|
|
46
|
Giaccia AJ and Kastan MB: The complexity
of p53 modulation: Emerging patterns from divergent signals. Genes
Dev. 12:2973–2983. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shieh SY, Ikeda M, Taya Y and Prives C:
DNA damage-induced phosphorylation of p53 alleviates inhibition by
MDM2. Cell. 91:325–334. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gottlieb TM, Leal JF, Seger R, Taya Y and
Oren M: Cross-talk between Akt, p53 and Mdm2: Possible implications
for the regulation of apoptosis. Oncogene. 21:1299–1303. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lotem J, Peled-Kamar M, Groner Y and Sachs
L: Cellular oxidative stress and the control of apoptosis by
wild-type p53, cytotoxic compounds, and cytokines. Proc Natl Acad
Sci USA. 93:9166–9171. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Misra MK, Sarwat M, Bhakuni P, Tuteja R
and Tuteja N: Oxidative stress and ischemic myocardial syndromes.
Med Sci Monit. 15:RA209–RA219. 2009.PubMed/NCBI
|
|
51
|
Orrenius S, Gogvadze V and Zhivotovsky B:
Mitochondrial oxidative stress: Implications for cell death. Annu
Rev Pharmacol Toxicol. 47:143–183. 2007. View Article : Google Scholar
|
|
52
|
Kim DJ, Chan KS, Sano S and Digiovanni J:
Signal transducer and activator of transcription 3 (Stat3) in
epithelial carcinogenesis. Mol Carcinog. 46:725–731. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kusaba T, Nakayama T, Yamazumi K, Yakata
Y, Yoshizaki A, Inoue K, Nagayasu T and Sekine I: Activation of
STAT3 is a marker of poor prognosis in human colorectal cancer.
Oncol Rep. 15:1445–1451. 2006.PubMed/NCBI
|
|
54
|
Lin L, Liu A, Peng Z, Lin HJ, Li PK, Li C
and Lin J: STAT3 is necessary for proliferation and survival in
colon cancer-initiating cells. Cancer Res. 71:7226–7237. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Johnston PA and Grandis JR: STAT3
signaling: Anticancer strategies and challenges. Mol Interv.
11:18–26. 2011. View Article : Google Scholar : PubMed/NCBI
|