Introduction
Lung cancer is the leading cause of cancer-related
death worldwide. Despite advances in surgical techniques and
strategies of chemoradiotherapy and targeted therapy, the 5-year
survival rate of patients with lung cancer remains low (less than
15%). Therefore, it is important to investigate the molecular
mechanisms underlying lung cancer that would lead to new therapies
for improving patient survival and quality of life.
MicroRNAs (miRNAs) are a class of small endogenous
non-coding RNAs that regulate the expression of their target genes
at the post-transcriptional level. miRNAs play important roles in
various biological processes such as cell proliferation, migration,
development, differentiation and apoptosis (1,2).
Beyond the involvement in physiological processes, accumulating
studies also strongly suggest that the dysregulation of miRNAs may
contribute to the initiation and progression of cancer (3). miR-124 is the most abundant miRNA
expressed in neuronal cells and plays a role in neuronal
differentiation. In addition, emerging evidence implicates miR-124
in the pathogenesis of several human malignancies. It has been
reported that miR-124 can act as a putative tumor suppressor in
prostate (4), breast (5), colorectal (6), cervical (7) and gastric cancer (8), nasopharyngeal carcinoma (9), bladder (10) and ovarian cancer (11) and hepatocellular carcinoma (12). Several studies have also shown that
miR-124 is downregulated in lung cancer (13–15).
However, the role of miR-124 in lung adenocarcinoma (ADC) and the
underlying mechanisms through which miR-124 exerts its functions
remain unclear.
SOX9, which is one of the members of the SRY
box-containing (SOX) family, plays a key role in developmental
processes, including chondrogenesis, neurogenesis and male sex
determination (16,17). Subsequently, increasing evidence has
revealed that SOX9 is involved in the development of cancer. It has
been reported that SOX9 is upregulated in colorectal cancer
(18), lung ADC (19), prostate (20) and breast cancer (21), and pancreatic ductal ADC (22), whereas it is downregulated in
cervical carcinoma (23).
Therefore, SOX9 may function as an oncogene or tumor suppressor
depending on tumor origin. Intriguingly, several miRs, including
miR-145 (24), miR-32 (25) and miR-101 (26), have been found to target SOX9 in
different tissues. Real et al (27) showed that miR-124 can regulate the
expression of SOX9 in developing mouse ovarian cells. However, the
potential regulatory effect of miR-124 on SOX9 expression in lung
ADC has not been verified.
In the present study, our results showed that
miR-124 was substantially decreased in lung ADC tissues, and
upregulation of miR-124 inhibited the proliferation, migration and
invasion of A549 cells. Moreover, our data demonstrated that SOX9
is a direct target gene and functional mediator of miR-124 in lung
ADC cells.
Materials and methods
Patients and tissue specimens
Sixty-five paired ADC lung samples and adjacent
non-tumorous lung tissues were obtained from patients undergoing
surgical resection and histologically confirmed by two pathologists
at the Second Hospital of Shandong University. These tissue samples
were immediately frozen in liquid nitrogen and then stored at −80°C
until protein and RNA extraction. Written informed consent was
obtained from all patients, and the present study was approved by
the Institutional Research Ethics Committee of the Second Hospital
of Shandong University.
Cell culture and DNA construction
The human lung ADC cell line A549 was purchased from
the Cell Bank of the Chinese Academy of Medical Sciences (Beijing,
China), and was routinely cultured in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% heat-inactivated fetal bovine
serum (FBS) (both from Invitrogen, Carlsbad, CA, USA), 100 U/ml
penicillin and 100 µg/ml streptomycin (both from
Sigma-Aldrich, St. Louis, MO, USA). The cells were incubated at
37°C in a humidified atmosphere with 5% CO2. The SOX9
plasmid was constructed according to a previous study (19). Briefly, the coding sequences of SOX9
were PCR amplified with the forward primer,
5′-GGATCCCATGAATCTCCTGGACCCCT-3′ and the reverse primer,
5′-GAATTCTCAAGGTCGAGTGAGCTGTGTGT-3′; and then subcloned into the
pCMV-Tag2 expression vector (Stratagene, La Jolla, CA, USA).
Real-time RT-PCR
To determine the relative expression level of
miR-124, total RNA was extracted from the tissues and cultured
cells using the mirVana™ miRNA isolation kit (AM1560; Ambion,
Austin, TX, USA) in accordance with the manufacturer's
instructions. The RNA was treated with DNase I (AM1906; Ambion) to
eliminate genomic DNA contamination and then subjected to cDNA
synthesis using the miScript reverse transcription kit (218061;
Qiagen, Hilden, Germany). Subsequently, real-time PCR was carried
out with the miScript SYBR-Green PCR kit (218073; Qiagen) in a
LightCycler (Roche Diagnostics, Mannheim, Germany) according to the
manufacturer's instructions. Primers for mature miR-124 and U6
snRNA were purchased from Qiagen (MS00006622 and MS00007497,
Hilden, Germany). All reactions were run in triplicate. The
relative expression level of miR-124 was quantified by
normalization to endogenous U6 snRNA expression level using the
2−ΔΔCt method.
Dual-luciferase reporter assay
To determine whether miR-124 can bind to the 3′
untranslated region (3′UTR) of SOX9, we purchased the pEZX-SOX9
vector from GeneCopoeia (HmiT017635-MT06; Rockville, MD, USA). The
pEZX-SOX9 vector contains the firefly luciferase gene with the SOX9
3′UTR and the Renilla luciferase gene. The miR-124 mimic,
miR-124 mimic with mutant sequences (RiboBio, Guangzhou, China) or
the miR-124 mimic control was transiently co-transfected into A549
cells with the pEZX-SOX9 vector. Cells were harvested 48 h after
transfection and the luciferase activities were measured using the
Dual-Luciferase Reporter assay system (E1910; Promega, Madison, WI,
USA). Firefly luciferase activities were normalized to
Renilla luciferase activities to control for transfection
efficiency.
miR-124 overexpression in cultured
cells
The miR-124 expression vector was constructed using
BLOCK-iT™ Pol II miR RNAi expression vector kit with EmGFP
(K4936-00; Invitrogen) according to the manufacturer's protocol.
The negative control vector was provided by Invitrogen, which
contains an insert that can form a hairpin structure just as a
regular pre-miRNA, but is predicted not to target any known
vertebrate gene. The expression vector or control vector was
transfected into A549 cells with Lipofectamine 2000 reagent
(11668-019; Invitrogen). Twenty-four hours after transfection,
blasticidin (15205; Sigma, St. Louis, MO, USA) was added at a
concentration of 3 µg/ml for 10 days. Resistant cells were
analyzed by fluorescence microscopy (Nikon, Tokyo, Japan). For
transient transfection, miR-124 mimics (miR10000422) or miR-124
mimic negative control (miR01201) (both from RiboBio) were
transfected into A549 cells at a final concentration of 100 nM
following the provided instructions.
miR-124 knockdown
miR-124 inhibitor and miR-124 inhibitor control were
purchased from Exiqon (4102198 and 199006; Vedbaek, Denmark) and
transfected into A549 cells with Lipofectamine 2000 reagent, at a
final concentration of 50 nM. The cells were collected 48 h after
transfection, and the levels of miRNA-124 and SOX9 were determined
by real-time RT-PCR and western blotting.
Western blotting
Total proteins in the cells and tissues were
extracted using RIPA buffer (Beyotime Institute of Biotechnology,
Shanghai, China). The concentration of total proteins was measured
using the BCA™ protein assay kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA). A total of 40 µg proteins was separated
on 12% SDS-PAGE gels, and transferred onto nitrocellulose membranes
(Millipore, Billerica, MA, USA). The membranes were blocked with 5%
fat-free milk at room temperature for 2 h, followed by incubation
with the mouse anti-human primary monoclonal antibody against SOX9
(1:500; ab76997; Abcam, Cambridge, MA, USA) or GAPDH (1:5,000;
D190090; Sangon Biotech, Shanghai, China) at 4°C overnight. The
membranes were then washed in TBS for three times and incubated
with horseradish peroxidase (HRP)-conjugated goat anti-mouse
secondary antibody (1:5,000; D110087; Sangon Biotech) for 1 h at
room temperature. After another three times of washing in TBS,
signals from the HRP-conjugated secondary antibody were generated
using enhanced chemiluminescence solution (Amersham, Piscataway,
NJ, USA), and were detected by exposure of the membranes to X-ray
film (Kodak, Rochester, NY, USA). The relative signal intensity was
quantified by densitometry with UVIPhoto and UVISoft UVIB and
application V97.04 (UVItech, Cambridge, UK).
MTT assay
After transfection with the miR-124 mimic or miR-124
inhibitor, the A549 cells were harvested, plated into 96-well
plates (2.0×103 cells/well) and incubated at 37°C. At
different time points (4, 24, 48 or 72 h), 10 µl of MTT
reagent (5 mg/ml; Sigma-Aldrich) was added to each well and cells
were incubated for another 4 h. Then, the supernatant was
discarded, and 150 µl of dimethyl sulfoxide (DMSO) was added
to each well. Colorimetric analysis was performed at the wavelength
of 490 nm. Data were derived from three independent
experiments.
Migration and invasion assays
Cell migration and invasion capacities were measured
in vitro using CytoSelect™ 24-Well Cell Migration and
Invasion Assay Combo kit (Cell Biolabs, San Diego, CA, USA)
according to the manufacturer's instructions. Briefly,
1×105 transfected cells were seeded into the upper
chamber. For the invasion assays, the filter membrane was coated
with a uniform layer of dried basement membrane matrix solution.
DMEM containing 10% FBS was used as a chemoattractant, and added to
the lower chamber. Following a 24-h incubation, cells on the upper
side of the filters were carefully removed with cotton-tipped
swabs. Invaded cells on the lower membrane were stained and
colorimetric analysis was performed. The relative migratory and
invasive activities were determined by the measurement of the
optical density at 560 nm and using the value of the negative
control as 1.
Statistical analysis
All statistical analyses were performed using the
SPSS 17.0 software package (SPSS, Inc., Chicago, IL, USA). The data
are presented as the mean ± standard deviation (SD). Differences
between groups were analyzed using the Student's t-test. P-values
are two-sided, and P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-124 is downregulated in lung ADC
tissues and its correlation with the SOX9 protein level
The expression levels of miR-124 in ADC tissues and
corresponding adjacent non-cancerous tissues were detected by
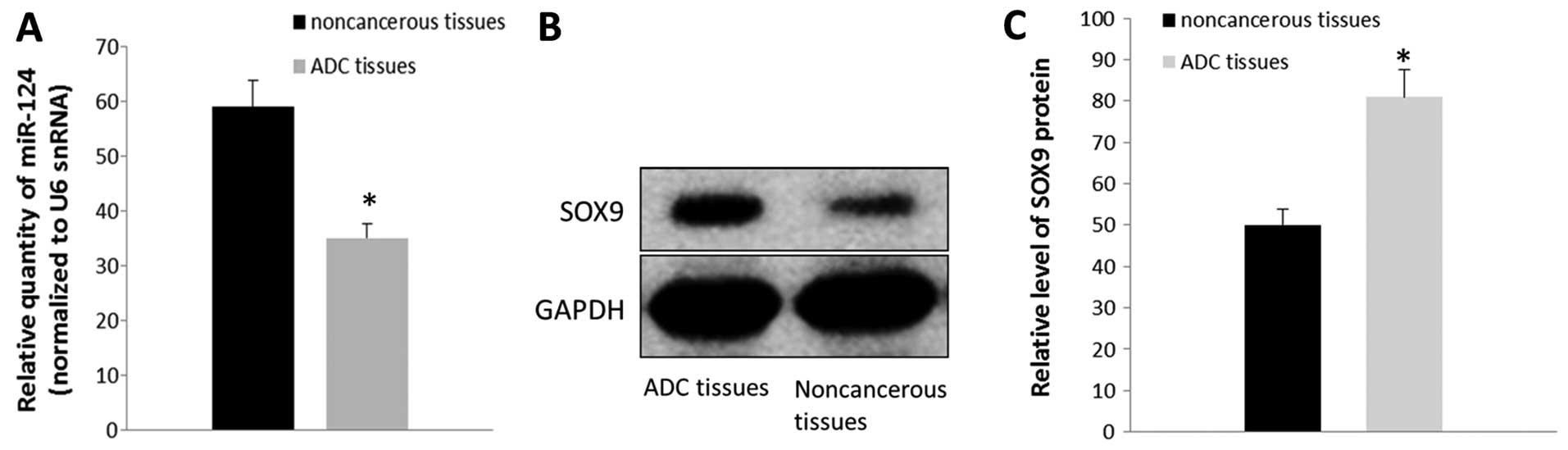
real-time RT-PCR. As shown in Fig.
1A, the results indicated that the relative level of miR-124
expression was significantly lower in the ADC specimens compared
with that noted in the controls (P<0.05) (Fig. 1A). SOX9 protein levels were also
detected by western blot analysis. The results showed that the SOX9
protein level was upregulated in the tumor samples when compared
with that noted in the adjacent non-cancerous tissues (P<0.05)
(Fig. 1B and C). Our data suggested
that the level of SOX9 protein is inversely associated with the
expression of miR-124.
miR-124 suppresses the proliferative,
migratory and invasive capacities of the A549 cells
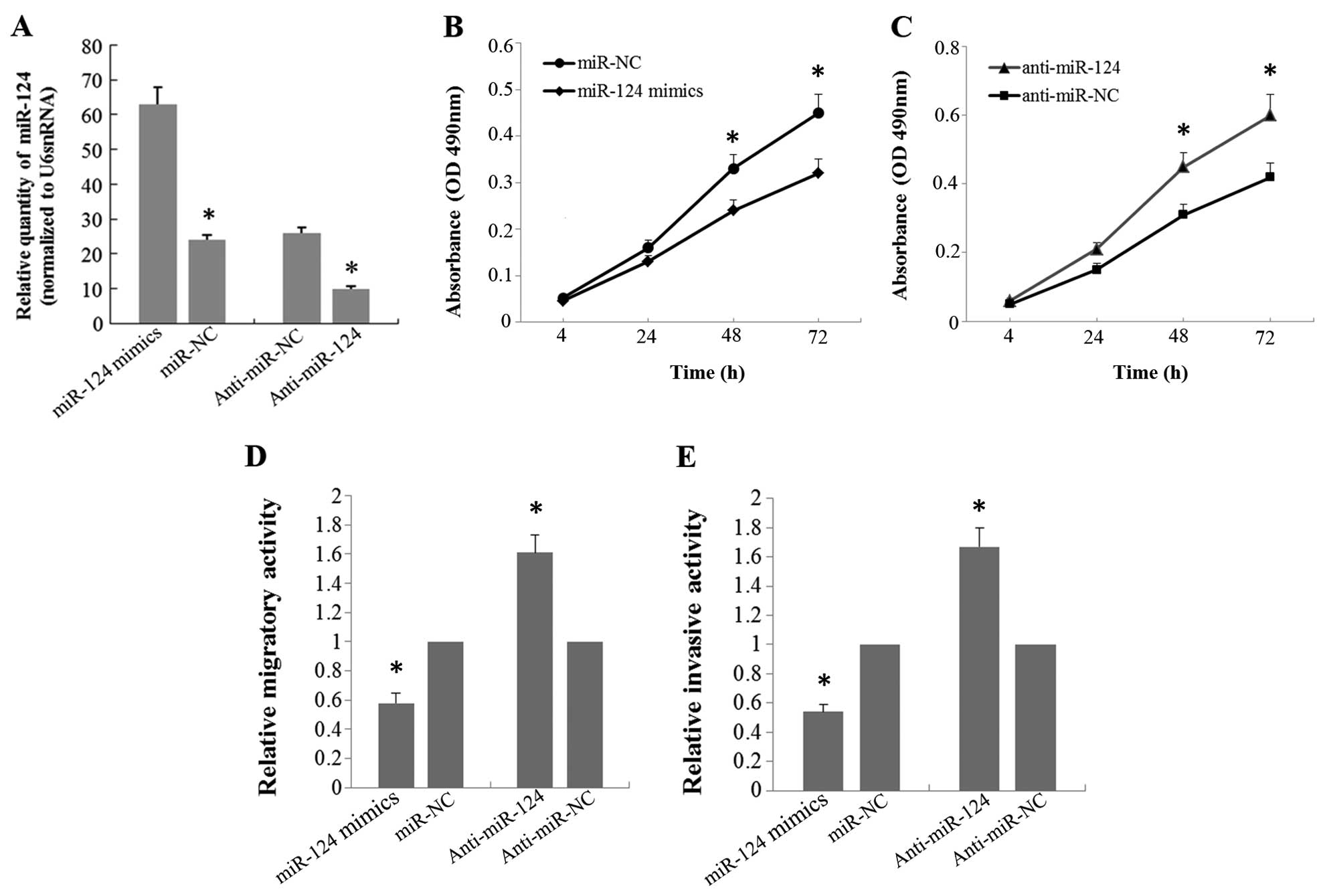
To investigate the effects of miR-124 on the
biological behaviors of the A549 cells, we transfected A549 cells
with the miR-124 mimic or miR-124 inhibitor. Real-time RT-PCR was
performed to confirm an increase or decrease in the miR-124 level
48 h after miR-124 mimic or miR-124 inhibitor transfection
(Fig. 2A). MTT assay was utilized
to evaluate cell proliferative capacity. The results showed that
overexpression of miR-124 significantly suppressed cell growth of
the A549 cells, whereas knockdown of miR-124 resulted in a
significant increase in proliferation of the A549 cells compared
with that noted in the controls (Fig.
2B and C). Transwell assay was performed to investigate the
effect of miR-124 on the migration and invasion of A549 cells. As
shown in Fig. 2D and E, the
migratory and invasive capabilities were significantly decreased in
the A549 cells transfected with the miR-124 mimic compared with
these capabilities noted in the control groups. Accordingly, when
A549 cells were transfected with the miR-124 inhibitor, the
migratory and invasive capabilities were markedly increased
compared with the controls.
SOX9 is a target gene of miR-124 in A549
cells
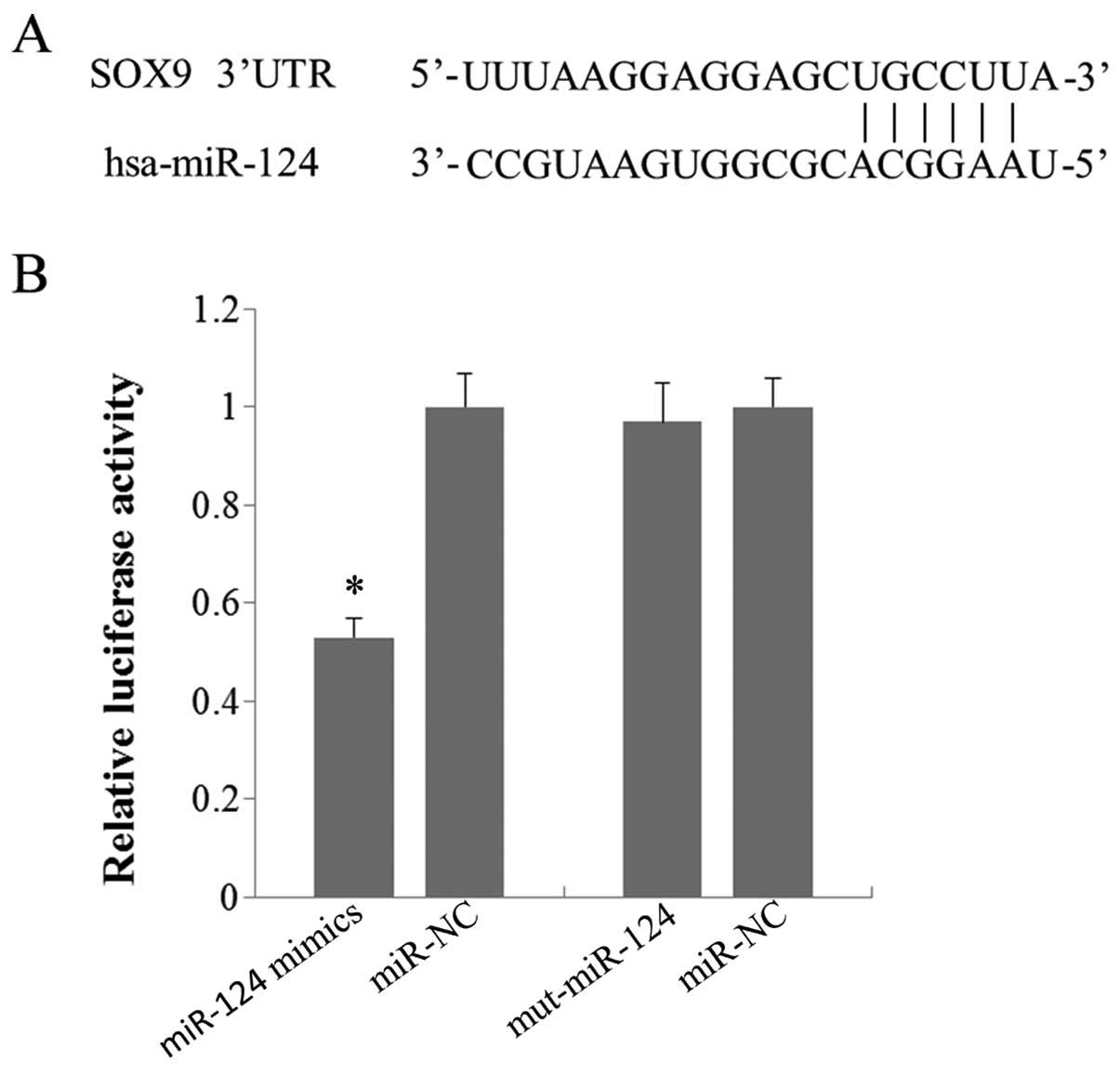
Using the bioinformatic tools at TargetScan database
(http://www.targetcan.org/), SOX9 was
identified as a candidate target of miR-124, since it contains a
putative miR-124 binding site in the 3′UTR (Fig. 3A). To determine whether miR-124 can
bind to its seed sequence present in the SOX9 3′UTR, we performed a
luciferase reporter assay using the pEXZ-SOX9 vector containing the
SOX9 3′UTR. The pEXZ-SOX9 vector was cotransfected into A549 cells
with the miR-124 mimic or miR-124 mimic control. Luciferase
activities were measured at 48 h after transfection. The results
showed that overexpression of miR-124 led to a 47% decrease in
luciferase expression, measured as relative luciferase activity,
compared to the controls (Fig. 3B).
However, when the pEXZ-SOX9 vector was cotransfected into the A549
cells with the miR-124 mimic with six mutated sequences
(UCCUUACCGCGGUGAAUGCC) or the miR-124 mimic control, only a very
slight effect on luciferase activity was observed (Fig. 3B). These results indicated that
miR-124 was able to bind to the predicted seed sequence in the
3′UTR of SOX9 mRNA.
miR-124 regulates SOX9 protein in A549
cells
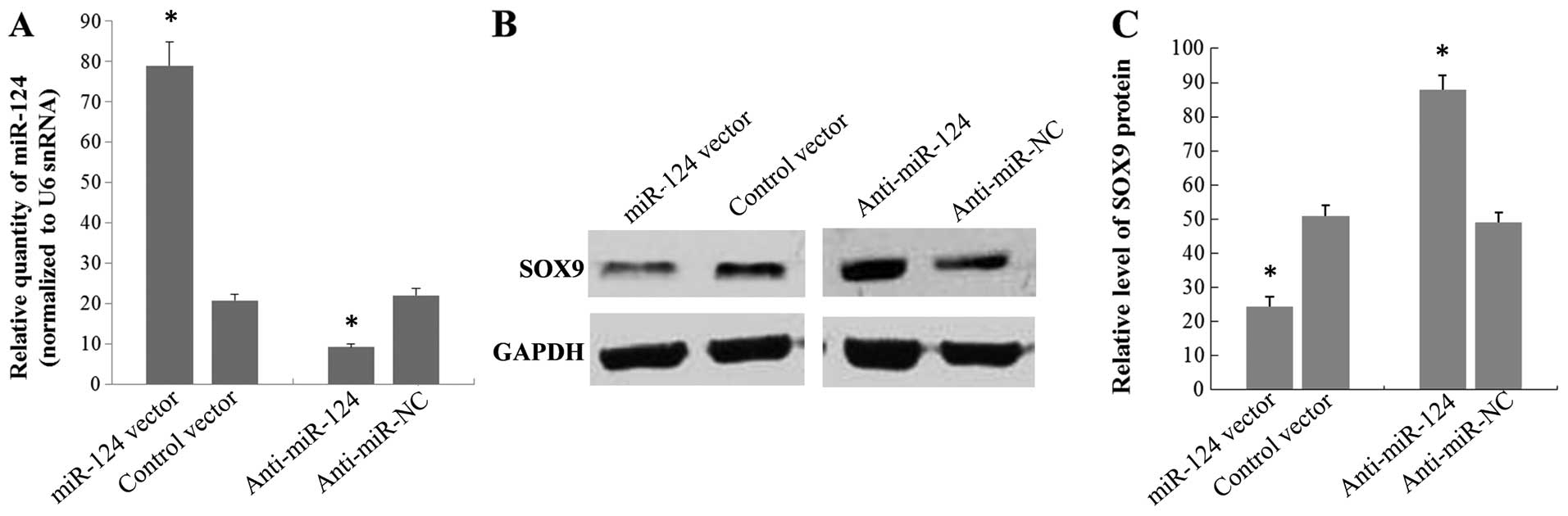
To investigate whether miR-124 affects SOX9
expression, we next examined the effect of this miRNA in a stable
transfectant cell line of miR-124. Results from real-time RT-PCR
showed that there was a 3.8-fold increase in miR-124 levels in the
stable transfectant cell line of miR-124, as compared to cells
stably transfected with the negative control vector (Fig. 4A). Western blot analysis performed
on the same cells showed that the SOX9 protein level was clearly
reduced (52%) in the stable transfection cell line of miR-124
compared with the controls (Fig. 4B and
C). We also knocked down miR-124 expression by transfecting
A549 cells with the miR-124 inhibitor and analyzed the effects on
SOX9 expression. A549 cells transfected with the miR-124 inhibitor
showed a 2.4-fold decrease in the miR-124 level compared with the
cells transfected with the miR-124 inhibitor control (Fig. 4A). As expected, the downregulation
of miR-124 led to an increase in the SOX9 protein level of 1.8-fold
compared to the negative controls (Fig.
4B and C). Taken together, our data suggested that the
expression of SOX9 is regulated by miR-124.
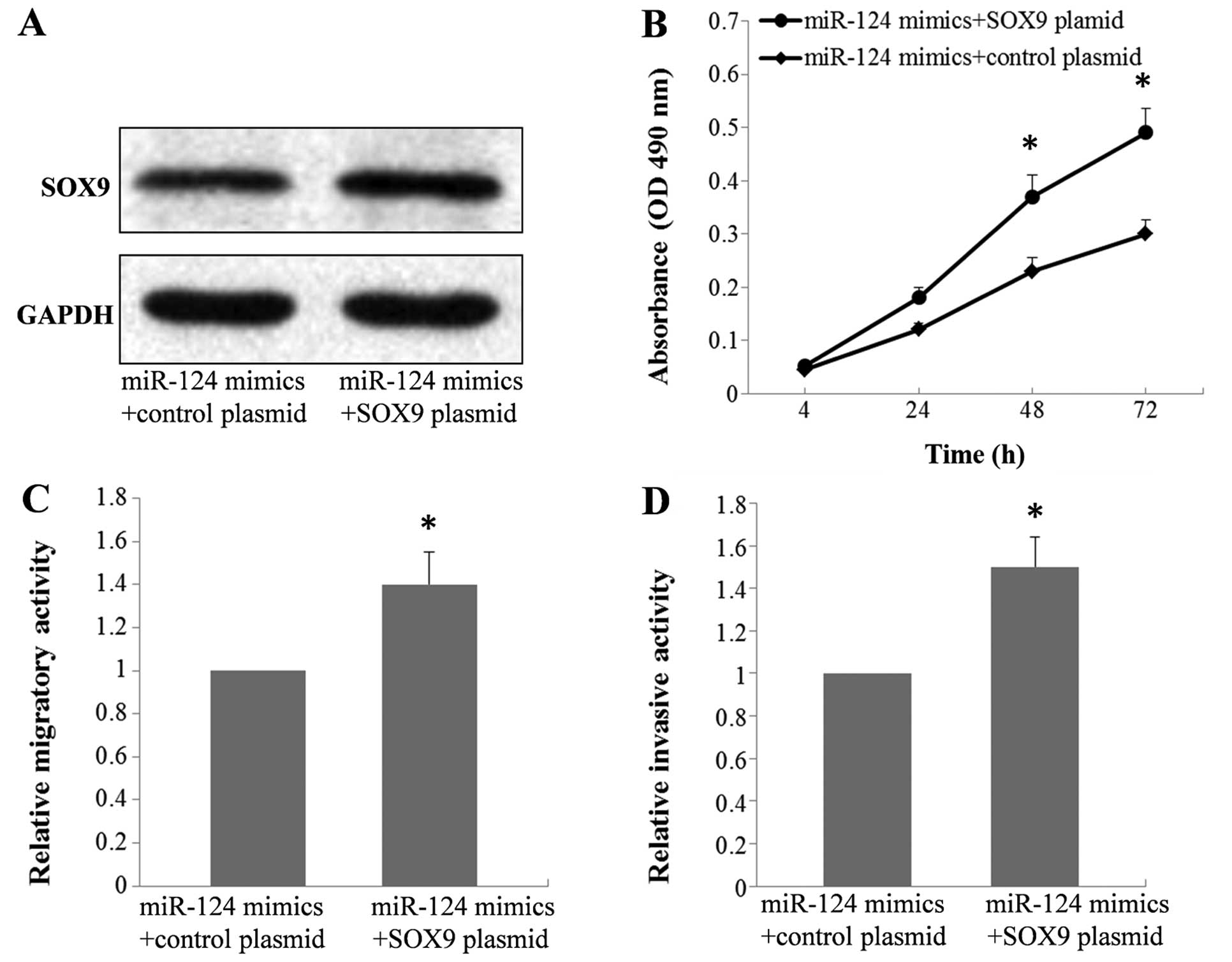
SOX9 is a functional mediator of miR-124
in A549 cells
In our previous study (28), we explored the functional role of
SOX9 in A549 cells and showed that upregulation of SOX9 promoted
cell proliferation, migration and invasion. To determine whether
the suppressive effect of miR-124 on the biological behaviors of
A549 cells is mediated by repression of SOX9, the miR-124 mimic was
cotransfected into the A549 cells with the SOX9 plasmid or control
plasmid. Western blot analysis was performed 48 h after
transfection and the results indicated that the protein level of
SOX9 was recovered after treatment with the SOX9 plasmid compared
to the controls (Fig. 5A).
Moreover, MTT, migration and invasion assays showed that exogenous
expression of SOX9 was able to restore the proliferative, migratory
and invasive activity compared with these cell behaviors noted in
the controls (Fig. 5B–D). This
indicated that the suppressive effect of miR-124 was mediated by
repression of SOX9, and SOX9 is a functional mediator of miR-124 in
A549 cells.
Discussion
The dysregulation of miRNAs is involved in the
initiation and progression of various types of tumors. However,
their potential roles in lung adenocarcinoma (ADC) remain unclear.
In the present study, we found that miR-124 was downregulated in
ADC tissues and the level of miR-124 was inversely correlated with
SOX9 expression. Moreover, the functional assays in human lung ADC
cell line A549 showed that enforced expression of miR-124
significantly inhibited cell proliferation, migration and invasion.
When A549 cells were transfected with the miR-124 expression vector
or miR-124 inhibitor, the SOX9 protein level was downregulated or
upregulated, which indicated that miR-124 could regulate SOX9
expression. The luciferase reporter assay confirmed that SOX9 is a
direct target of miR-124. Finally, we showed that exogenous
expression of SOX9 rescued the phenotype induced by overexpression
of miR-124 in the A549 cells. These results suggested that miR-494
affects cell proliferation, migration and invasion by directly
regulating SOX9 expression in lung ADC.
Previous studies have shown that miR-124 is
associated with several human malignancies and it is a putative
tumor suppressor. However, the function of miR-124 in lung ADC
remains unclear. In the present study, our results showed that
miR-124 was downregulated in the tissues of lung ADC patients using
real-time RT-PCR assays, and we focused on the functions of miR-124
in the proliferation, migration and invasion of lung ADC cells.
Using human lung ADC cell line A549 transfected with the miR-124
mimic or miR-124 inhibitor, we demonstrated that miR-124 was
functionally involved in suppression of cell proliferation,
migration and invasion.
SOX9, which belongs to a family of genes called SOX
[SRY (sex determining region Y)-boxes], is a key regulator of
embryonic development. Emerging evidence has provided a link
between SOX9 and cancer development. In some types of cancers, SOX9
is upregulated and functions as an oncogene, whereas in other types
of cancers, SOX9 is downregulated and functions as a tumor
suppressor. Our previous study found that SOX9 was upregulated in
lung ADC and promoted cell proliferation, migration and invasion.
In the present study, we found that SOX9 is a direct target gene of
miR-124. More importantly, exogenous expression of SOX9 restored
the proliferative, migratory and invasive activities induced by
overexpression of miR-124 in A549 cells. All these results suggest
that miR-124 can inhibit cell proliferation, migration and invasion
by directly targeting SOX9 and SOX9 is an important functional
mediator of miR-124 in lung ADC cells. As we know, there exists a
complicated connection between miRNAs and their target genes. One
miRNA can have many targets and several miRNAs can have the same
target. Some genes, such as talin 1, PDE4B and STAT3, have also
been found to be the target of miR-124 in several human
malignancies (4,15,29).
SOX9 may not be the only miR-124 target dysregulated in lung ADC.
Thus, further studies are needed to investigate the potential
functional targets of miR-124, and the actual mechanisms by which
miR-124 affects lung ADC progression require further
clarification.
In conclusion, the present study demonstrated that
miR-124 was downregulated in tissues of lung ADC patients.
Moreover, miR-124 inhibited cell proliferation, migration and
invasion of A549 cells. Finally, we confirmed that miR-124 directly
targets SOX9 by binding to the 3′UTR of SOX9 and SOX9 was found to
be an important functional mediator of miR-124 in A549 cells.
Overall, our findings indicate that miR-124 functions as a tumor
suppressor in lung ADC and may be a promising candidate for
miR-based therapy against lung ADC.
Acknowledgments
The present study was supported by grants from the
Shandong Provincial Natural Science Foundation of China (grant no.
ZR2015PH038), and the Seed Fund of the Second Hospital of Shandong
University (grant no. S2014010018).
References
|
1
|
Wang Y, Huang C, Chintagari NR, Xi D, Weng
T and Liu L: miR-124 regulates fetal pulmonary epithelial cell
maturation. Am J Physiol Lung Cell Mol Physiol. 309:L400–L413.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Osada H and Takahashi T: MicroRNAs in
biological processes and carcinogenesis. Carcinogenesis. 28:2–12.
2007. View Article : Google Scholar
|
|
3
|
Lovat F, Valeri N and Croce CM: MicroRNAs
in the pathogenesis of cancer. Semin Oncol. 38:724–733. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang W, Mao YQ, Wang H, Yin WJ, Zhu SX
and Wang WC: MiR-124 suppresses cell motility and adhesion by
targeting talin 1 in prostate cancer cells. Cancer Cell Int.
15:492015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dong LL, Chen LM, Wang WM and Zhang LM:
Decreased expression of microRNA-124 is an independent unfavorable
prognostic factor for patients with breast cancer. Diagn Pathol.
10:452015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xi ZW, Xin SY, Zhou LQ, Yuan HX, Wang Q
and Chen KX: Downregulation of rho-associated protein kinase 1 by
miR-124 in colorectal cancer. World J Gastroenterol. 21:5454–5464.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wan HY, Li QQ, Zhang Y, Tian W, Li YN, Liu
M, Li X and Tang H: MiR-124 represses vasculogenic mimicry and cell
motility by targeting amotL1 in cervical cancer cells. Cancer Lett.
355:148–158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu CB, Li QL, Hu JF, Zhang Q, Xie JP and
Deng L: miR-124 inhibits growth and invasion of gastric cancer by
targeting ROCK1. Asian Pac J Cancer Prev. 15:6543–6546. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng XH, Huang HR, Lu J, Liu X, Zhao FP,
Zhang B, Lin SX, Wang L, Chen HH, Xu X, et al: MiR-124 suppresses
tumor growth and metastasis by targeting Foxq1 in nasopharyngeal
carcinoma. Mol Cancer. 13:1862014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang T, Wang J, Zhai X, Li H, Li C and
Chang J: MiR-124 retards bladder cancer growth by directly
targeting CDK4. Acta Biochim Biophys Sin. 46:1072–1079. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang H, Wang Q, Zhao Q and Di W: MiR-124
inhibits the migration and invasion of ovarian cancer cells by
targeting SphK1. J Ovarian Res. 6:842013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu Y, Yue X, Cui Y, Zhang J and Wang K:
MicroRNA-124 suppresses growth of human hepatocellular carcinoma by
targeting STAT3. Biochem Biophys Res Commun. 441:873–879. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Y, Li H, Han J and Zhang Y:
Down-regulation of microRNA-124 is correlated with tumor metastasis
and poor prognosis in patients with lung cancer. Int J Clin Exp
Pathol. 8:1967–1972. 2015.PubMed/NCBI
|
|
14
|
Sun Y, Ai X, Shen S and Lu S:
NF-κB-mediated miR-124 suppresses metastasis of non-small-cell lung
cancer by targeting MYO10. Oncotarget. 6:8244–8254. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Yu Z, Li Y, Liu S, Gao C, Hou X, Yao
R and Cui L: The tumor suppressor miR-124 inhibits cell
proliferation by targeting STAT3 and functions as a prognostic
marker for postoperative NSCLC patients. Int J Oncol. 46:798–808.
2015.
|
|
16
|
Chaboissier MC, Kobayashi A, Vidal VI,
Lützkendorf S, van de Kant HJ, Wegner M, de Rooij DG, Behringer RR
and Schedl A: Functional analysis of Sox8 and Sox9 during sex
determination in the mouse. Development. 131:1891–1901. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akiyama H, Chaboissier MC, Martin JF,
Schedl A and de Crombrugghe B: The transcription factor Sox9 has
essential roles in successive steps of the chondrocyte
differentiation pathway and is required for expression of Sox5 and
Sox6. Genes Dev. 16:2813–2828. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bruun J, Kolberg M, Nesland JM, Svindland
A, Nesbakken A and Lothe RA: Prognostic significance of β-catenin,
E-cadherin, and SOX9 in colorectal cancer: Results from a large
population-representative series. Front Oncol. 4:1182014.
View Article : Google Scholar
|
|
19
|
Jiang SS, Fang WT, Hou YH, Huang SF, Yen
BL, Chang JL, Li SM, Liu HP, Liu YL, Huang CT, et al: Upregulation
of SOX9 in lung adenocarcinoma and its involvement in the
regulation of cell growth and tumorigenicity. Clin Cancer Res.
16:4363–4373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qin GQ, He HC, Han ZD, Liang YX, Yang SB,
Huang YQ, Zhou L, Fu H, Li JX, Jiang FN, et al: Combined
overexpression of HIVEP3 and SOX9 predicts unfavorable biochemical
recurrence-free survival in patients with prostate cancer. Onco
Targets Ther. 7:137–146. 2014.PubMed/NCBI
|
|
21
|
Müller P, Crofts JD, Newman BS,
Bridgewater LC, Lin CY, Gustafsson JA and Ström A: SOX9 mediates
the retinoic acid-induced HES-1 gene expression in human breast
cancer cells. Breast Cancer Res Treat. 120:317–326. 2010.
View Article : Google Scholar
|
|
22
|
Xia S, Feng Z, Qi X, Yin Y, Jin J, Wu Y,
Wu H, Feng Y and Tao M: Clinical implication of Sox9 and activated
Akt expression in pancreatic ductal adenocarcinoma. Med Oncol.
32:3582015. View Article : Google Scholar
|
|
23
|
Wang HY, Lian P and Zheng PS: SOX9, a
potential tumor suppressor in cervical cancer, transactivates
p21WAF1/CIP1 and suppresses cervical tumor growth.
Oncotarget. 6:20711–20722. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu D, Chen H, Yang X, Chen W, Wang L, Xu
J and Yu L: miR-32 functions as a tumor suppressor and directly
targets SOX9 in human non-small cell lung cancer. Onco Targets
Ther. 8:1773–1783. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li X, Wang ZX, Wang ZS and Li QF: Effect
of microRNA-101 on apoptosis of rabbit condylar cartilage cells by
inhibiting target gene SOX9. Asian Pac J Trop Med. 8:502–505. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mak IW, Singh S, Turcotte R and Ghert M:
The epigenetic regulation of SOX9 by miR-145 in human
chondrosarcoma. J Cell Biochem. 116:37–44. 2015. View Article : Google Scholar
|
|
27
|
Real FM, Sekido R, Lupiáñez DG,
Lovell-Badge R, Jiménez R and Burgos M: A microRNA (mmu-miR-124)
prevents Sox9 expression in developing mouse ovarian cells. Biol
Reprod. 89:782013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang X, Ju Y, Zhou MI, Liu X and Zhou C:
Upregulation of SOX9 promotes cell proliferation, migration and
invasion in lung adenocarcinoma. Oncol Lett. 10:990–994.
2015.PubMed/NCBI
|
|
29
|
Kim J, Jeong D, Nam J, Aung TN, Gim JA,
Park KU and Kim SW: MicroRNA-124 regulates glucocorticoid
sensitivity by targeting phosphodiesterase 4B in diffuse large B
cell lymphoma. Gene. 558:173–180. 2015. View Article : Google Scholar : PubMed/NCBI
|