Introduction
Gastric cancer is one of the most common malignant
tumors, with more than 1 million new cases diagnosed worldwide in
2012. Nearly two-fifths of these new cases were diagnosed in
patients in mainland China. Moreover, gastric cancer accounts for
~9% of all cancer-related deaths (~723,000/year) (1). Tumor metastasis is the direct cause of
death for the majority of patients with gastric cancer. Therefore,
therapies that inhibit the invasion and metastasis of tumors are a
primary focus of cancer researchers.
Ca2+ is a critical regulator of cell
migration in various cell types, including tumor cells (2), and controls a broad range of cellular
functions, such as gland secretion, neural excitation, muscle
contraction and apoptosis (2,3).
Store-operated calcium influx is the dominant mechanism through
which Ca2+ enters non-excitable cells, including tumor
cells (4,5). Recent studies have shown that stromal
interaction molecule 1 (STIM1) and Orai1 are involved in
store-operated Ca2+ entry. While Orai1 is an essential
channel protein, STIM1 is a 77-kDa, endoplasmic, single-pass
membrane protein that functions to maintain intracellular calcium
homeostasis and regulate intracellular calcium ion concentrations
(6). Indeed, the SOCE channel,
which consists of STIM1 and Orai1, is a major route of
Ca2+ entry in non-excitable cells (6,7). STIM1
has mostly been implicated in functions in the immune system
(8–10) and cardiovascular system (11,12).
Recent studies have also shown that STIM1 is expressed in a variety
of cancers, including breast (13),
cervical (14), liver (15) and colon cancer (16). Additionally, STIM1 knockdown has
been shown to accelerate motility in melanoma cells (17). In contrast, Yang et al
(13,15) demonstrated that silencing of STIM1
or Orai1 inhibited the migration and metastasis of breast cancer
and hepatocellular carcinoma. STIM1 has also been shown to mediate
cell proliferation and apoptosis in a variety of cell lines.
However, STIM1 does not affect apoptosis in breast cancer cells,
demonstrating that STIM1 may have cell type-specific effects on
cancer phenotypes. Additionally, the role of STIM1 in gastric
cancer has not been established.
Therefore, in the present study, we examined the
expression of STIM in gastric cancer tissues and evaluated the role
of STIM1 in promoting cancer-associated malignant behaviors in
gastric cancer cells.
Materials and methods
Cell culture and RNA interference
Human gastric cancer cells (MKN-45, SGC7-901, AGS
and NCI-N87 cells) were donated by the National Key Laboratory of
Biotherapy. Cells were maintained in Roswell Park Memorial
Institute (RPMI)-1640 medium supplemented with 10% fetal bovine
serum (FBS) (both from HyClone) in a humidified atmosphere with 5%
CO2 at 37°C for 24 h. Small interfering RNA (siRNA)
containing the target sequence for STIM1
(5′-GGUUUGCCUAUAUCCAGAACCGUUA-3′ and
5′-UAACGGUUCUGGAUAUAGGCAAACC-3′) or control siRNA (Invitrogen,
Carlsbad, CA, USA) was transfected into human gastric cancer cells.
At 24 or 48 h following transfection, the cells were treated and
prepared for cell migration, adhesion and invasion assays.
Collection of clinical specimens and
immunohistochemical analysis
Ninety gastric tumor tissues and 30 normal gastric
tissues were collected from the First Affiliated Hospital of
Chengdu Medical College (Chengdu, China). The protocol of the
present study was approved by the Institutional Ethics Committee of
Chengdu Medical College. Informed consent for the present study was
received from all patients prior to the commencement of the
experiments. The tissues were preserved in 4% polyoxymethylene
solution, embedded in paraffin and sectioned. Endogenous peroxidase
was blocked by incubation for 15 min in 0.3%
H2O2. Antigen retrieval was carried out in
0.01 M sodium citrate-hydrochloric acid buffer solution. Monoclonal
mouse anti-STIM1 antibodies (4 µg/ml; Abcam, Cambridge, UK)
were used for immunohistochemical analysis of STIM1 expression for
all samples. Monoclonal mouse anti-CD31 antibodies (1:1,000;
Epitomics, Burlingame, CA, USA) were used for analysis of the
expression of CD31 in vascular endothelial cells of the tissue
samples. Tissues were incubated with the primary antibody at 4°C
overnight. After washing with phosphate-buffered saline (PBS), the
cells were incubated with the appropriate secondary antibody (goat
anti-mouse IgG) for 30 min at 37°C. Peroxidase activity was
revealed by 3,3-diaminobenzidine and cells were counter-stained
with hematoxylin. Negative controls were processed with isotype
control antibody. Saturation and intensity of the immunostained
cells were evaluated over eight visual fields under a light
microscope (Olympus Optical, Tokyo, Japan). For statistical
analysis, total staining of the protein of interest was scored as
the product of the staining intensity (on a scale of 0–3: negative,
0; weak, 1; moderate, 2; and strong, 3) × the percentage of cells
stained (positively recorded on an ordered categorical scale: 0,
zero; 1, 1–25%; 2, 26–50%; and 3, 51–100%), resulting in a scale of
0–9. The evaluation was performed by two independent
investigators.
Western blot analysis
Total protein was extracted from the lysed cells or
tissues ground in liquid nitrogen. Protein samples were then
subjected to sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred to polyvinylidene difluoride
membranes. The membranes were blocked in 5% non-fat milk for 1 h
and then incubated overnight with primary mouse anti-STIM1
antibodies (2 µg/ml) at 4°C overnight. The dilution of the
antibodies against extracellular signal-regulated kinase (ERK),
mitogen-activated protein kinase (MEK), phospho-ERK1/2, p-MEK
(Abcam) and β-actin (Sigma-Aldrich, St. Louis, MO, USA) was
1:1,000. The membranes were then incubated with a 1:20,000 dilution
of peroxidase-linked anti-rabbit or anti-mouse IgG secondary
antibodies for 2 h at 37°C, and proteins were visualized using
BeyoECL Plus (both from Beyotime, Guangdong, China). The intensity
of the immunoreactive bands was analyzed using ImageJ and
normalized to the expression of β-actin. Western blotting was used
to explore the levels of ERK, MEK, p-ERK, p-MEK expressed in the
MKN-45 human gastric cancer cells.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from human stomach tissues
and cell lines with TRIzol reagent (Invitrogen). A NanoDrop 2000
was used to measure the concentration and quality of RNA. The total
RNA was then reverse transcribed into cDNA, followed by PCR using a
PrimeScript RT-PCR kit (Takara Bio, Shiga, Japan) according to the
manufacturer's instructions. Quantitative real-time PCR was
performed using SYBR Premix Ex Taq (Perfect Real-Time) (Takara
code: DRR041) according to the manufacturer's instructions. The
following primer sets were used for evaluation of STIM1 expression:
STIM1 forward, 5′-TGTGTCTCCCTTGTCCATGC-3′ and reverse,
5′-CATCTGAGGTTTGGGGG-3′; GAPDH forward, 5′-AGGGCTGCTTTTAACTCTGGT-3′
and reverse, 5′-CCCCACTTGATTTTGGAGGGA-3′. The relative differences
in expression of STIM1 mRNA were calculated using the comparative
cycle threshold method.
Flow cytometry
Gastric cancer cells were seeded into 24-well plates
and incubated at 37°C until reaching 60–70% confluency. The cells
were transfected with STIM1-siRNA for 24 or 48 h and then harvested
using trypsin. The cells were collected in 1.5-ml centrifuge tubes,
washed with PBS and resuspended in 1× binding buffer. Next, 2
µl of Annexin V was added, followed by addition of 2
µl of propidium iodide (PI). Cells were incubated at room
temperature for 30 min in the dark. Cell apoptosis was then
determined by flow cytometry (BD Biosciences, Franklin Lakes, NJ,
USA). Approximately 10,000 events (cells) were evaluated for each
sample.
Cell proliferation
Cell proliferation was determined by MTT assay.
Cells were transfected with STIM1-siRNA for 24 or 48 h. Ten
microliters of 10 mg/ml
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
was added to each well, and the plates were incubated at 37°C under
5% CO2 for 4 h. The supernatant was removed, and the
formed formazan crystals were dissolved in 100 µl dimethyl
sulphoxide. The absorbance at 450 nm was then measured with a BD
microplate reader. Growth inhibition was determined from the
differences in optical densities between the experimental and
control wells as a percentage of the control.
Wound healing assay
Cells (2×105 cells/ml) were seeded in
each well of a 24-well plate and incubated at 37°C until reaching
60–70% confluency. The cell monolayer was scratched with a
10-µl pipette tip, and the cells were then washed with PBS
and transfected with STIM1-siRNA. After 48 h, randomly chosen
fields were photographed under a light microscope with a 4×
objective.
Migration and invasion assays
Migration assays were carried out using Transwell
plates (24-well plates, 8-mm pore size; Corning, Shanghai, USA).
Cells (105/0.2 ml) were trypsinized, resuspended in 0.1%
BSA-1640, and plated in the upper chambers in duplicate. The lower
chamber was filled with RPMI-1640 medium containing 10% FBS. After
24 h, the non-migrating cells were removed from the upper membrane
surface with a cotton swab. Transwell membranes were then fixed
with 4% paraformaldehyde and stained with crystal violet. The
number of cells that migrated through to the lower side of the
membrane was counted under a light microscope with a 20 or 40×
objective. The invasion assay was performed using the ECM550
(Chemicon, Shanghai, USA) as an index of the invasive activity of
tumor cells. FBS (10%) was added to the lower chamber, as described
for the cell migration experiments, and cell counting was performed
as described for the cell migration experiments.
Adhesion assay
After transfection, cells (2×105
cells/ml) were seeded in 96-well plates pre-incubated with Matrigel
for 20 min at 37°C. Unattached cells were removed by washing with
PBS. Attached cells were fixed in 4% paraformaldehyde for 15 min
and stained with 0.02% crystal violet solution for 15 min. Randomly
chosen fields were photographed under a light microscope with a 20
or 40× objective.
Statistical analysis
All data are shown as the means ± standard
deviations (SDs). Statistical analyses were performed using the
Statistical Package for Social Science program (SPSS for Windows,
version 13.0; SPSS, Inc., Chicago, IL, USA) based on the results of
three independent experiments. One-way analysis of variance (ANOVA)
was used for comparing differences between groups. Differences with
P-values of <0.05 were considered to be statistically
significant.
Results
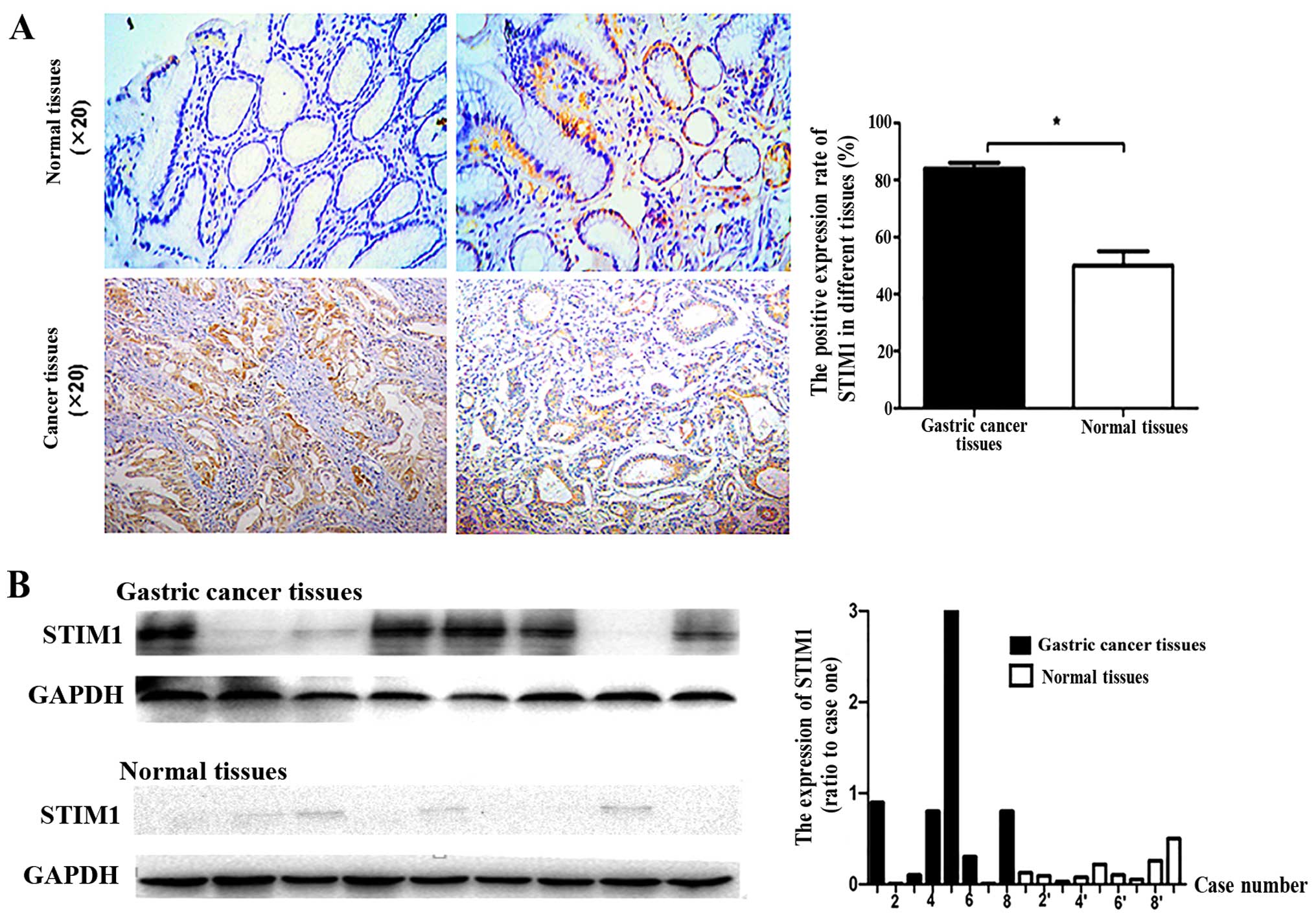
STIM1 is expressed at higher levels in
gastric tumor tissues than that in normal gastric tissues
To clarify the clinical relevance of STIM1
expression in gastric cancer progression, 90 gastric tumor and 30
normal gastric tissues were analyzed by immunohistochemistry.
Additionally, 8 fresh gastric tumor tissues and 9 fresh normal
gastric tissues were analyzed by western blotting. STIM1 was
expressed in both gastric cancer and normal gastric tissues
(Fig. 1). The rate of STIM1
positivity was higher in the gastric cancer tissues than the rate
in normal gastric tissues (Fig.
1A). Although STIM1 immunoreactivity was found in normal
tissues, enhanced immunoreactivity was detected in the cancer
tissues (Fig. 1B). These results
confirmed that STIM1 was more frequently expressed in gastric
cancer tissues than in normal tissues.
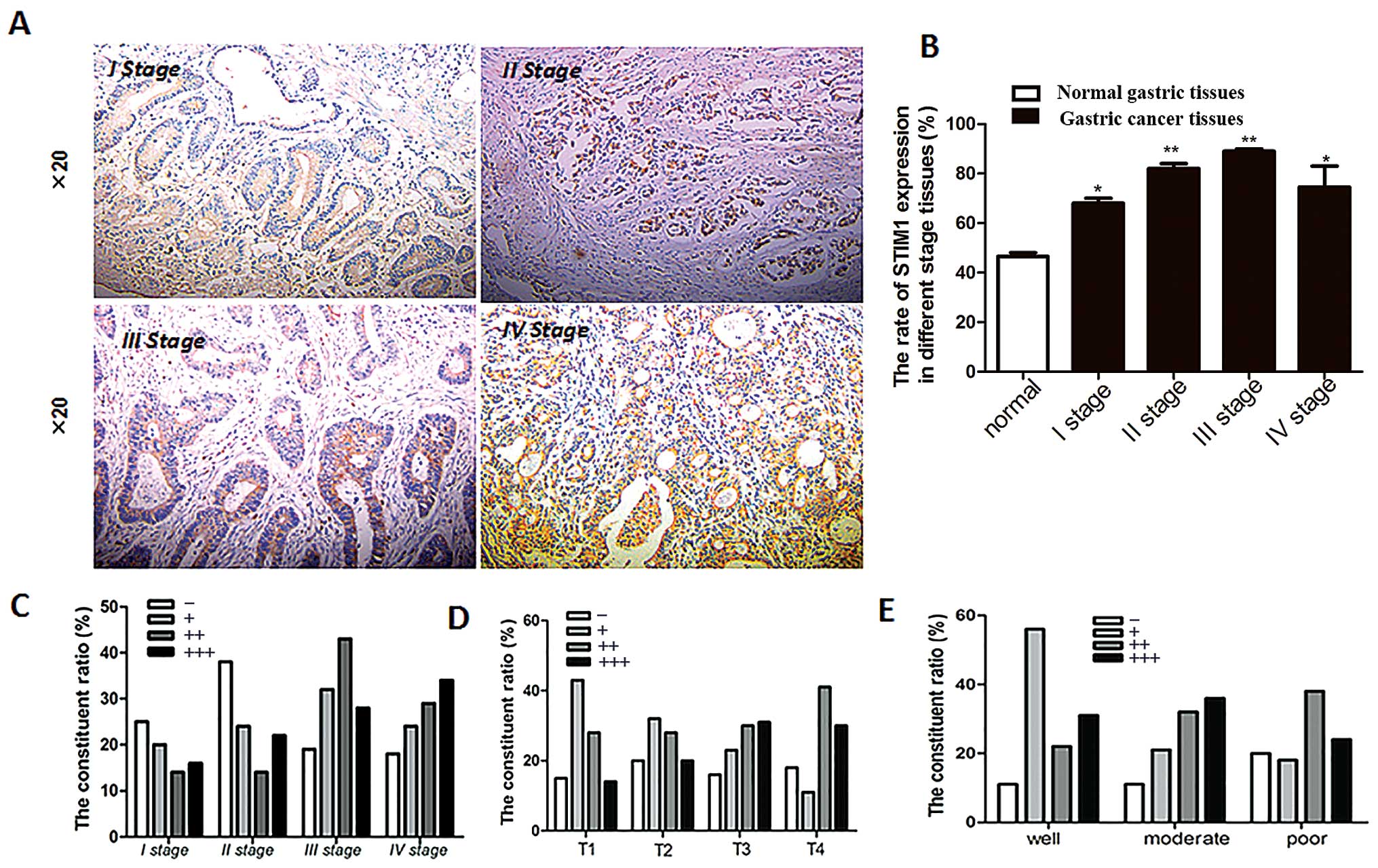
STIM1 and pathological factors
To analyze the relationship between STIM1 expression
and clinical stages of gastric cancer, gastric cancer tissue
samples were grouped according to patient age, gender, clinical and
T stages, and differentiation (Table
I). The rate of STIM1 positivity for gastric cancers of all
stages was higher than the rate in normal gastric tissues (Fig. 2A and B). However, differential
expression was not observed among the different clinical stages
(Fig. 2B). STIM1 expression in
patients with clinical stage I tended to be weaker than that in
patients with clinical stages II, III or IV (Fig. 2C), suggesting that STIM1 expression
may increase as the cancer becomes more advanced. Additionally,
STIM1 expression was increased with increasing T stage (Fig. 2D). Next, we measured changes in
STIM1 expression in tumor cells and normal cells with varying
degrees of differentiation, as a measure of the degree of
malignancy. STIM1 expression also increased as the degree of
differentiation decreased (Fig.
2E). Therefore, taken together, these data suggest that STIM1
expression is associated with the degree of malignancy in gastric
cancer.
 | Table IRelationships between STIM1 staining
intensity and pathological patient features. |
Table I
Relationships between STIM1 staining
intensity and pathological patient features.
| Feature | Immunoreactive score
| Total |
|---|
| − | + | ++ | +++ |
|---|
| Age (years) | | | | | 78 |
| <40 | 0 | 0 | 0 | 1 | 1 |
| 40–55 | 7 | 4 | 4 | 3 | 18 |
| 56–65 | 5 | 5 | 7 | 12 | 29 |
| >65 | 1 | 7 | 15 | 7 | 30 |
| Gender | | | | | 79 |
| Male | 7 | 11 | 20 | 19 | 57 |
| Female | 7 | 5 | 6 | 4 | 22 |
| Clinical stage | | | | | 87 |
| I | 4 | 6 | 3 | 3 | 16 |
| II | 5 | 6 | 8 | 6 | 25 |
| III | 4 | 4 | 12 | 8 | 28 |
| IV | 3 | 4 | 5 | 6 | 18 |
| T stage | | | | | 92 |
| T1 | 1 | 3 | 2 | 1 | 7 |
| T2 | 5 | 8 | 6 | 5 | 25 |
| T3 | 7 | 10 | 13 | 13 | 43 |
| T4 | 3 | 2 | 7 | 5 | 17 |
|
Differentiation | | | | | 62 |
| Well | 1 | 5 | 2 | 1 | 9 |
| Moderate | 2 | 4 | 6 | 7 | 19 |
| Poor | 7 | 6 | 13 | 8 | 34 |
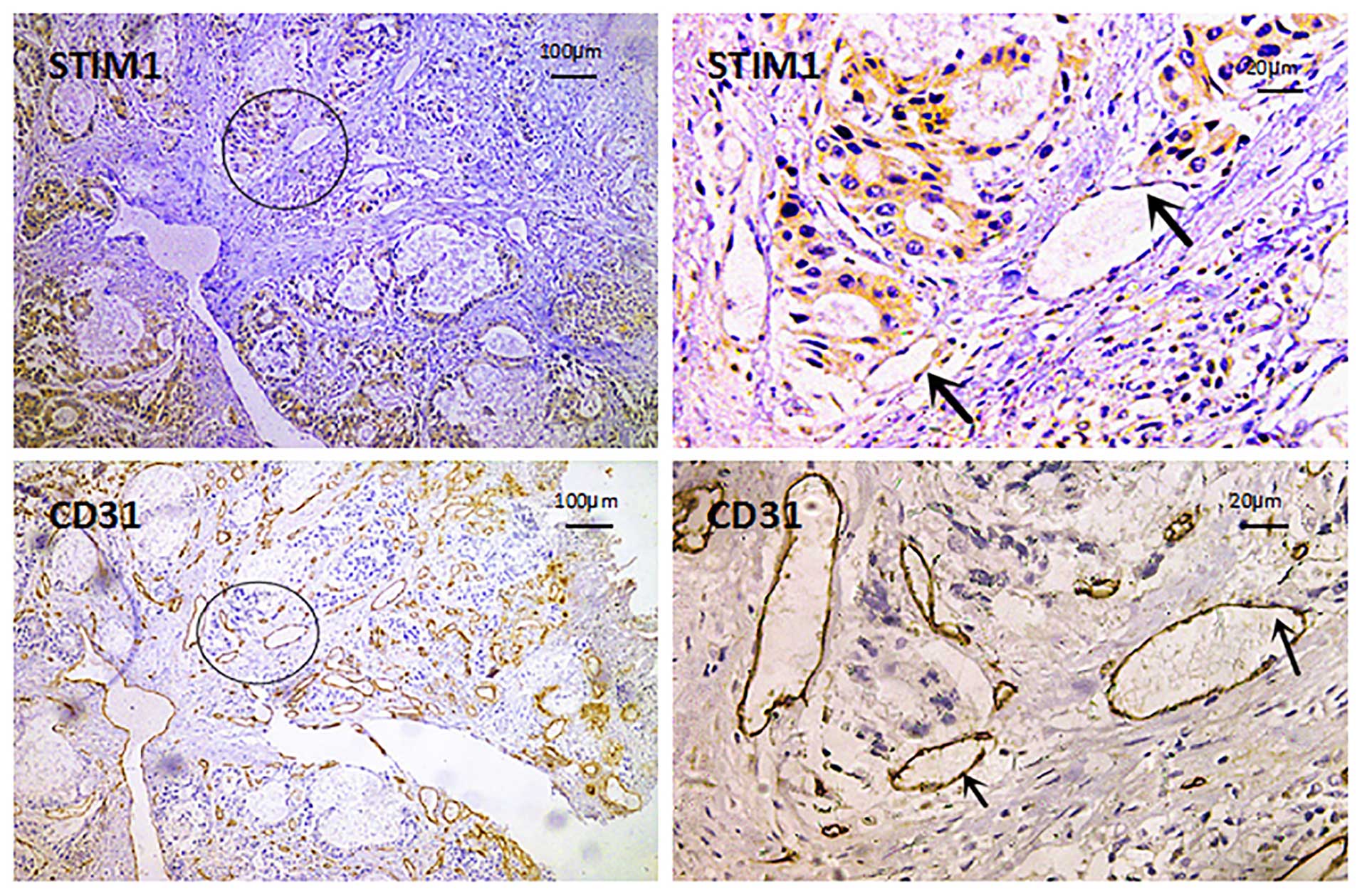
We then analyzed the expression of CD31 as a
molecular marker of endothelial cells using immunohistochemistry.
We found that STIM1-positive cells colocalized with CD31-positive
cells, suggesting that tumor vascular endothelial cells express
STIM1 (Fig. 3).
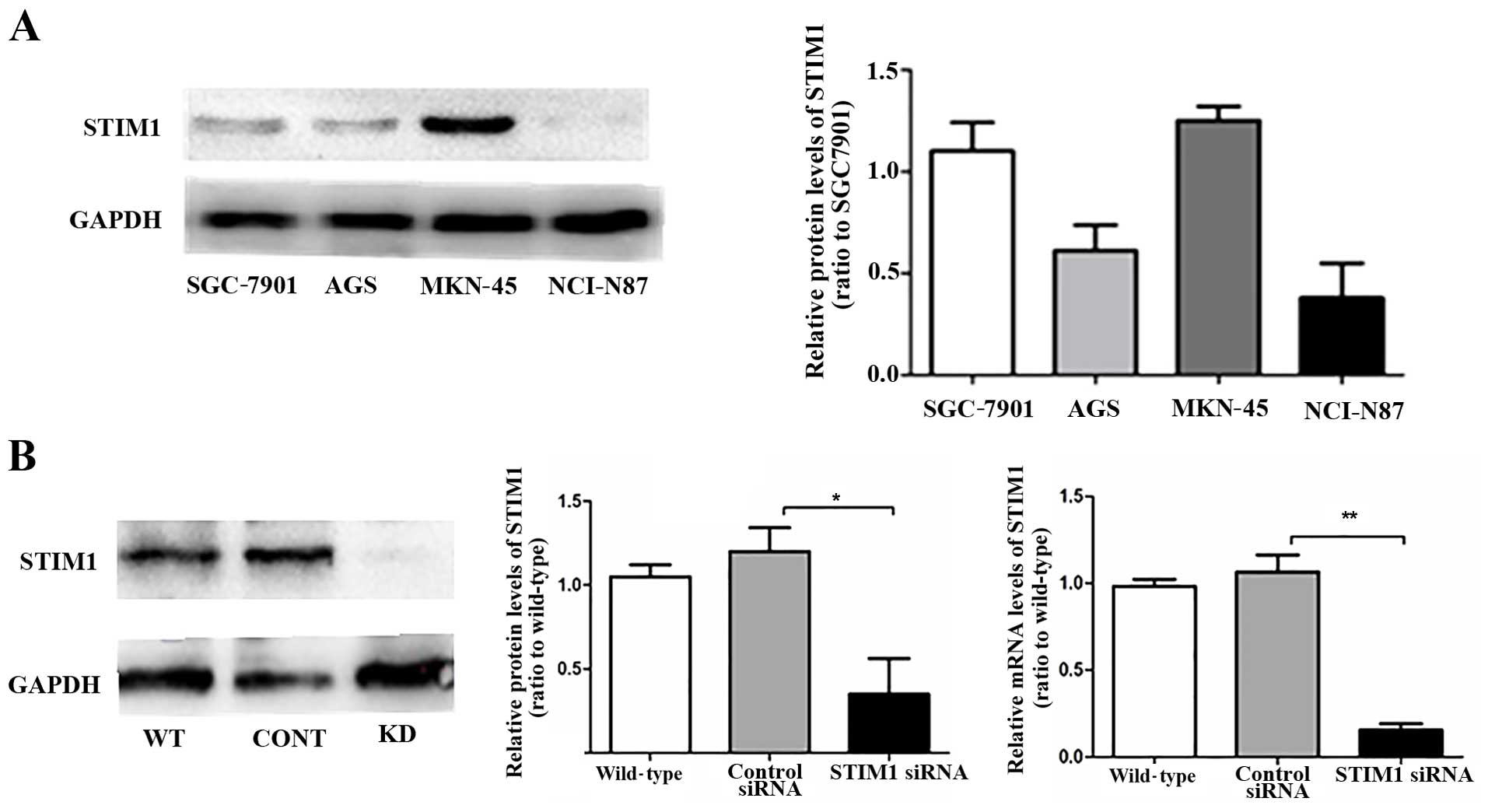
STIM1 is expressed in four gastric cancer
cell lines
We next analyzed the expression of STIM1 in MKN-45,
SGC-7901, AGS and NCI-N87 cells. MKN-45 and SGC-7901 cells
exhibited similar expression levels of STIM1, >2-fold higher
than that in the NCI-N87 cells (Fig.
4A). Notably, both of these cell lines expressing high levels
of STIM1 also showed high migration capacity. In preparation for
our next experiments, we then examined the effects of an siRNA
targeting STIM1 on STIM1 expression in MKN-45 cells. Western
blotting and RT-PCR showed that siRNA targeting STIM1 effectively
downregulated expression of STIM1 (Fig.
4B).
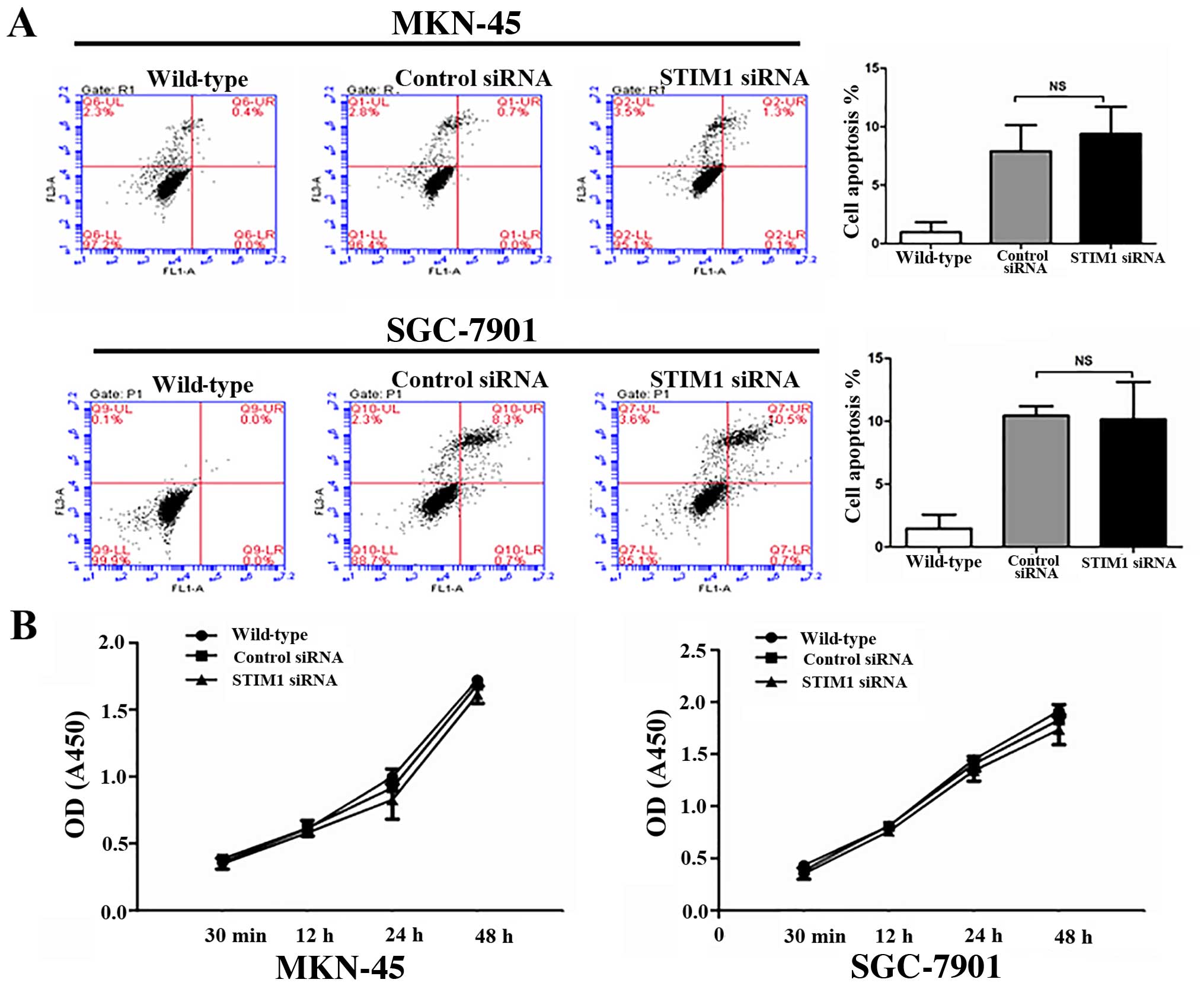
STIM1 does not affect proliferation or
apoptosis in gastric cancer cells
As shown, using MTT assays, transfection with STIM1
siRNA did not affect the proliferation of the MKN-45 and SGC-7901
cells (Fig. 5B). Additionally,
there were no statistically significant differences in the numbers
of apoptotic cells after transfection with control siRNA or siRNA
targeting STIM1, as measured using Annexin V-FITC/PI staining with
flow cytometry (Fig. 5A).
STIM1-knockdown affects the migration and
invasion of gastric cancer cells
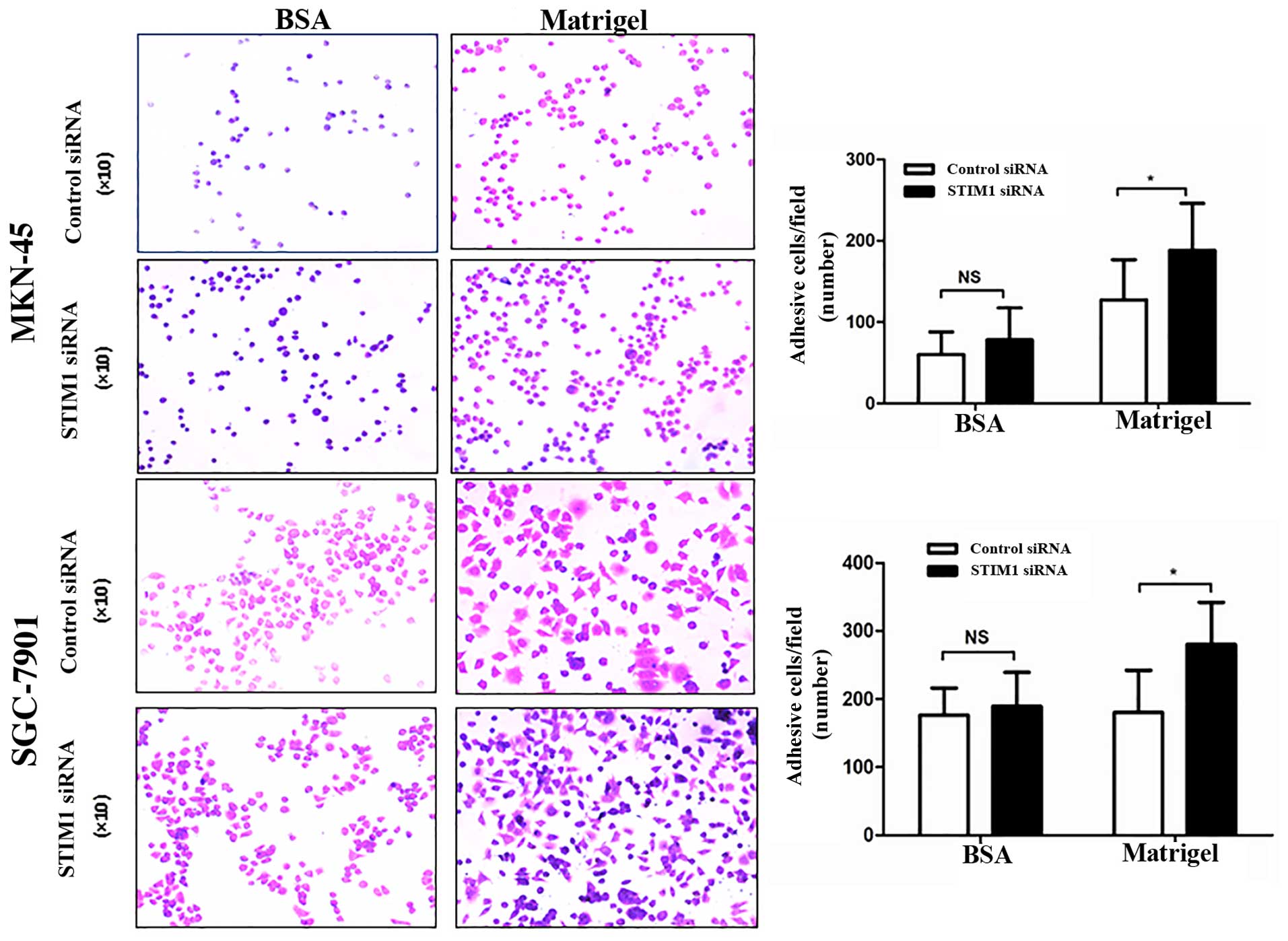
We then examined the adhesion capacity of gastric
cancer cells after transfection with STIM1 siRNA using
Matrigel-coated plates. Matrigel simulates extracellular matrix
(ECM) components and can be used to study adhesion and cell
motility. In our assay, transfection with STIM1 siRNA enhanced the
adhesion ability of the cells in the Matrigel-coated wells but did
not affect adhesion in the wells coated with 0.1% bovine serum
albumin (Fig. 6), indicating that
knockdown of STIM1 promoted the ability of gastric cancer cells to
detach from the ECM.
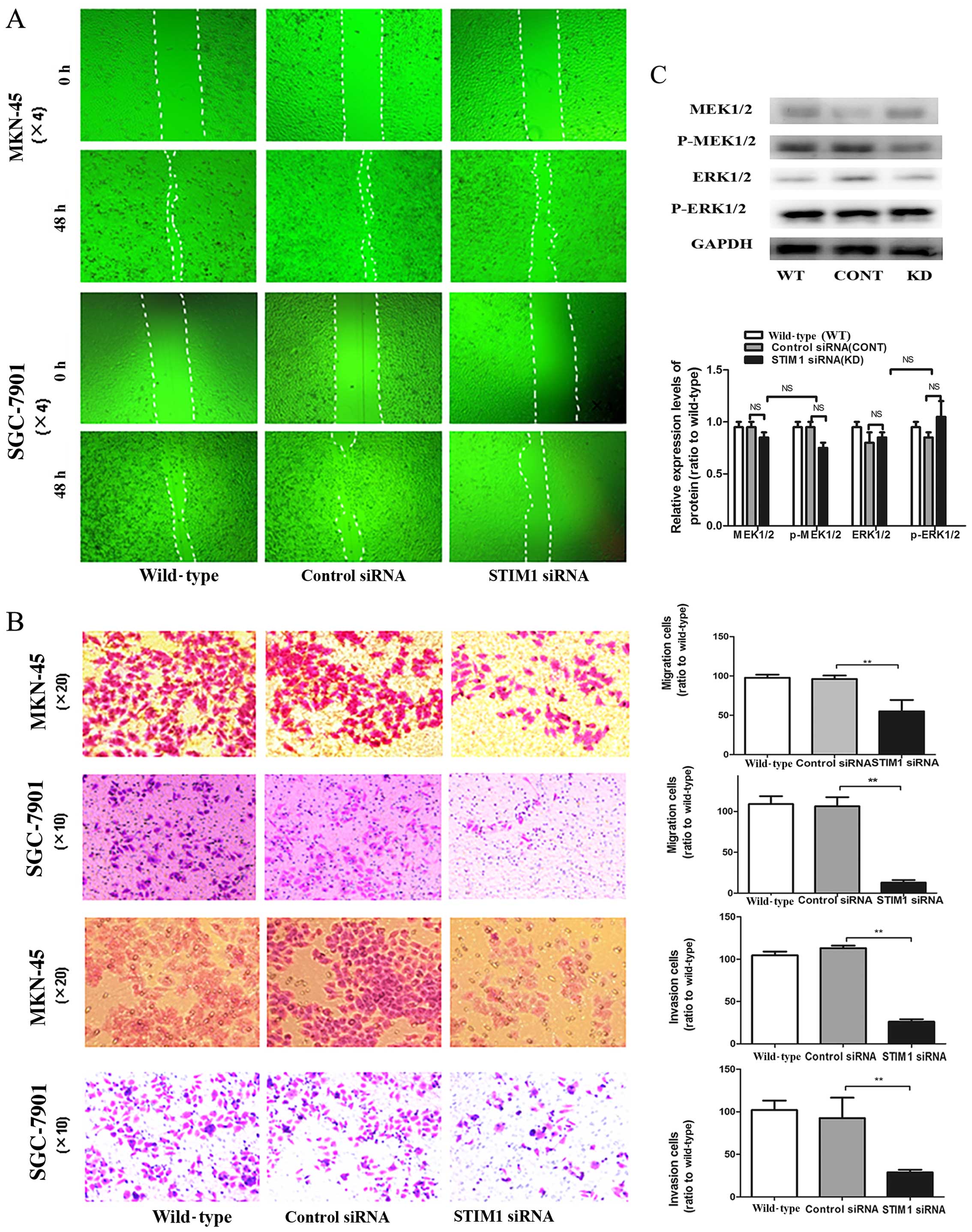
Next, we examined the effects of STIM1 knockdown on
cell migration and invasion, two process that are critical for
tumor metastasis. In the wound healing assay, the width of the
wound in the cells transfected with STIM1 siRNA was greater than
that in the control cells. As shown in Fig. 7B, knockdown of STIM1 by siRNA in the
MKN-45 and SGC-7901 cells reduced cell migration through membranes
compared with the migration assay of the control siRNA-transfected
cells. Furthermore, invasion assays using Transwell chambers
demonstrated that STIM1 knockdown also blocked cell invasion
(Fig. 7A). Taken together, these
results showed that STIM1 plays an important role in controlling
tumor cell motility.
Finally, we further examined the mechanisms through
which STIM1 mediated cell metastasis by evaluating changes in the
phosphorylation of MEK/ERK proteins, which are involved in
regulating cell division, differentiation, proliferation and
motility. Western blotting showed that STIM1 knockdown did not
alter the expression or phosphorylation of MEK or ERK (Fig. 7C). Therefore, STIM1 was involved in
regulation of motility in gastric cancer cells through a method
independent of the MEK/ERK pathway.
Discussion
In the present study, we examined the role of STIM1
in gastric cancer progression and metastasis. Our results showed
that STIM1 was highly expressed in gastric cancer cell lines and
tissues and that knockdown of STIM1 enhanced adhesion and reduced
metastasis-related cellular functions, including migration adn
invasion. Therefore, these data suggest that STIM1 may represent a
novel target in the treatment of gastric cancer.
Tumor growth and metastasis are closely related to
tumor angiogenesis. Several studies have shown that anti-angiogenic
agents can inhibit tumor growth and distant metastasis. Moreover,
STIM1 is expressed in human umbilical vein endothelial cells
(HUVECs) and pulmonary artery endothelial cells (HPAECs), promoting
endothelial cell proliferation and regulating the function of
endothelial tissue, which is involved in angiogenesis (18,19).
Our results also found that STIM1 was expressed in tumor vascular
endothelial cells. Therefore, the effects of STIM1 on the promotion
of gastric cancer metastasis may be associated with the regulation
of tumor angiogenesis.
Of the four gastric cancer cell lines examined in
the present study, the highest level of STIM1 expression was
observed in the MKN-45 cells, which were also the most highly
metastatic cells. Consistent with the importance of STIM1 in
cancer, STIM1 has been suggested to be a tumor suppressor, inducing
cell death in rhabdomyosarcoma and rhabdoid tumor cell lines, which
do not express endogenous STIM1 (20). In contrast, breast cancer cell lines
express easily detectable levels of STIM1 mRNA, and no
relationship between STIM1 expression and proliferation or
apoptosis was observed in breast cancer cell lines. However, in
other studies, STIM1 has been shown to mediate cell proliferation.
Indeed, STIM1 knockdown has been shown to decrease p21 and Cdc25C
protein levels in cervical cancer, whereas several cell cycle
regulators, including cyclin A and B1 and Cdk1, were not affected
by STIM1 knockdown (16). Numerous
studies have shown that STIM1 is involved in tumor cell apoptosis
in response to anticancer drugs (16,21).
However, our results did not support a direct role for STIM1 in
gastric cancer cell apoptosis, and knockdown of STIM1 did not
affect the proliferation of the MKN-45 or SGC-7901 cells.
Therefore, the role of STIM1 in cancer cell proliferation may vary
according to cell type.
In the present study, we also examined the effects
of STIM1 on cancer cell migration and invasion, which are related
to the metastatic potential of cancer cells and can be used to
distinguish between benign and malignant tumors. In previous
studies, Yang et al (13)
demonstrated that silencing of STIM1 or Orai1 inhibited the
migration and metastasis of breast cancer cells by suppressing
focal adhesion turnover. Moreover, reduction of STIM1 by RNA
interference or application of a pharmacological inhibitor of SOCE
promoted intercellular adhesion and weakened tumor cell invasion
and migration in hepatic carcinoma (15). In the present study, we found that
knockdown of STIM1 inhibited gastric cancer cell migration and
invasion and promoted adhesion. These effects may be mediated
through extracellular matrix (ECM) components. Cell migration is an
integrated process requiring both tumor cells and the surrounding
ECM, and we hypothesized that STIM1 may promote gastric cancer
metastasis through interaction with integrins as part of the ECM.
Alternatively, metastasis is a complex, highly regulated process
that also involves various intracellular signaling pathways,
including the MEK/ERK pathway, which regulates cell division,
differentiation, proliferation and motility (22–25).
For example, Ono et al (26)
and Neuzillet et al (27)
reported that silencing of PTK6 reduces ERK1/2 activation, whereas
PTK6 overexpression increased ERK1/2 activation, thereby promoting
cellular migration and invasion in pancreatic cancer. Additionally,
Nagini (28) found that vascular
endothelial growth factor (VEGF) expression was increased when the
ERK signaling pathway was inhibited. In the present study, we
showed that knockdown of STIM1 did not alter the expression or
activation of MEK or ERK, implying that STIM1 affected gastric
cancer cell migration through a pathway independent of the MEK/ERK
pathway.
In conclusion, our results showed that STIM1 was
expressed both in gastric cancer and normal gastric tissues, with
more frequent expression in cancerous tissues than in normal
tissues. STIM1 did not affect proliferation or apoptosis in gastric
cancer cells. However, knockdown of STIM1 blocked cell migration
and invasion and promoted cell adhesion. Further studies are
required to determine the mechanisms involved. We hypothesized that
STIM1, which promotes the proliferation of endothelial cells, may
affect cancer metastasis by regulating angiogenesis. Whether there
is a different level of STIM1 in tumor vascular endothelial and
normal vascular endothelial cells warrants further investigation.
This hypothesis will need to be tested in further in-depth studies.
In any case, our results provide a basis for the further analysis
of STIM1 as a specific molecular target in cancer therapy.
Acknowledgments
The present study was supported by the National
Clinical Medicine Research Foundation of China (grant no.
L2012055), and the National Natural Science Foundation of China
(grant no. 81302170).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Berridge MJ, Bootman MD and Roderick HL:
Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev
Mol Cell Biol. 4:517–529. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zheng L, Stathopulos PB, Li GY and Ikura
M: Biophysical characterization of the EF-hand and SAM domain
containing Ca2+ sensory region of STIM1 and STIM2.
Biochem Biophys Res Commun. 369:240–246. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oh-Hora M, Yamashita M, Hogan PG, Sharma
S, Lamperti E, Chung W, Prakriya M, Feske S and Rao A: Dual
functions for the endoplasmic reticulum calcium sensors STIM1 and
STIM2 in T cell activation and tolerance. Nat Immunol. 9:432–443.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hewavitharana T, Deng X, Soboloff J and
Gill DL: Role of STIM and Orai proteins in the store-operated
calcium signaling pathway. Cell Calcium. 42:173–182. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muik M, Schindl R, Fahrner M and Romanin
C: Ca2+ release-activated Ca2+ (CRAC)
current, structure, and function. Cell Mol Life Sci. 69:4163–4176.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stathopulos PB, Zheng L, Li GY, Plevin MJ
and Ikura M: Structural and mechanistic insights into
STIM1-mediated initiation of store-operated calcium entry. Cell.
135:110–122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Omilusik KD, Nohara LL, Stanwood S and
Jefferies WA: Weft, warp, and weave: The intricate tapestry of
calcium channels regulating T lymphocyte function. Front Immunol.
4:1642013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsumoto M and Baba Y: Role of
STIM-dependent Ca2+ influx in regulatory B cells.
Yakugaku Zasshi. 133:419–425. 2013.In Japanese. View Article : Google Scholar
|
|
10
|
Félix R, Crottès D, Delalande A,
Fauconnier J, Lebranchu Y, Le Guennec JY and Velge-Roussel F: The
Orai-1 and STIM-1 complex controls human dendritic cell maturation.
PLoS One. 8:e615952013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee KJ, Woo JS, Hwang JH, Hyun C, Cho CH,
Kim do H and Lee EH: STIM1 negatively regulates Ca2+
release from the sarcoplasmic reticulum in skeletal myotubes.
Biochem J. 453:187–200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fodor J, Matta C, Oláh T, Juhász T, Takács
R, Tóth A, Dienes B, Csernoch L and Zákány R: Store-operated
calcium entry and calcium influx via voltage-operated calcium
channels regulate intracellular calcium oscillations in
chondrogenic cells. Cell Calcium. 54:1–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang S, Zhang JJ and Huang XY: Orai1 and
STIM1 are critical for breast tumor cell migration and metastasis.
Cancer Cell. 15:124–134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen YF, Chiu WT, Chen YT, Lin PY, Huang
HJ, Chou CY, Chang HC, Tang MJ and Shen MR: Calcium store sensor
stromal-interaction molecule 1-dependent signaling plays an
important role in cervical cancer growth, migration, and
angiogenesis. Proc Natl Acad Sci USA. 108:15225–15230. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang N, Tang Y, Wang F, Zhang H, Xu D,
Shen Y, Sun S and Yang G: Blockade of store-operated
Ca2+ entry inhibits hepatocarcinoma cell migration and
invasion by regulating focal adhesion turnover. Cancer Lett.
330:163–169. 2013. View Article : Google Scholar
|
|
16
|
Sun S, Li W, Zhang H, Zha L, Xue Y, Wu X
and Zou F: Requirement for store-operated calcium entry in sodium
butyrate-induced apoptosis in human colon cancer cells. Biosci Rep.
32:83–90. 2012. View Article : Google Scholar
|
|
17
|
Suyama E, Wadhwa R, Kaur K, Miyagishi M,
Kaul SC, Kawasaki H and Taira K: Identification of
metastasis-related genes in a mouse model using a library of
randomized ribozymes. J Biol Chem. 279:38083–38086. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Abdullaev IF, Bisaillon JM, Potier M,
Gonzalez JC, Motiani RK and Trebak M: Stim1 and Orai1 mediate CRAC
currents and store-operated calcium entry important for endothelial
cell proliferation. Circ Res. 103:1289–1299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lodola F, Laforenza U, Bonetti E, Lim D,
Dragoni S, Bottino C, Ong HL, Guerra G, Ganini C, Massa M, et al:
Store-operated Ca2+ entry is remodelled and controls in
vitro angiogenesis in endothelial progenitor cells isolated from
tumoral patients. PLoS One. 7:e425412012. View Article : Google Scholar
|
|
20
|
Sabbioni S, Barbanti-Brodano G, Croce CM
and Negrini M: GOK: A gene at 11p15 involved in rhabdomyosarcoma
and rhabdoid tumor development. Cancer Res. 57:4493–4497.
1997.PubMed/NCBI
|
|
21
|
Li W, Zhang M, Xu L, Lin D, Cai S and Zou
F: The apoptosis of non-small cell lung cancer induced by cisplatin
through modulation of STIM1. Exp Toxicol Pathol. 65:1073–1081.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cerne JZ, Stegel V, Gersak K, Novakovic S
and Jasmina-Ziva: KRAS rs61764370 is associated with
HER2-overexpressed and poorly-differentiated breast cancer in
hormone replacement therapy users: A case control study. BMC
Cancer. 12:1052012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sui JQ, Xie KP, Zou W and Xie MJ: Emodin
inhibits breast cancer cell proliferation through the
ERα-MAPK/Akt-cyclin D1/Bcl-2 signaling pathway. Asian Pac J Cancer
Prev. 15:6247–6251. 2014. View Article : Google Scholar
|
|
24
|
Zhong LR, Chen X and Wei KM: Radix
tetrastigma hemsleyani flavone induces apoptosis in human lung
carcinoma a549 cells by modulating the MAPK pathway. Asian Pac J
Cancer Prev. 14:5983–5987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
de Mello RA, Marques DS, Medeiros R and
Araújo AM: Epidermal growth factor receptor and K-Ras in non-small
cell lung cancer-molecular pathways involved and targeted
therapies. World J Clin Oncol. 2:367–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ono H, Basson MD and Ito H: PTK6 promotes
cancer migration and invasion in pancreatic cancer cells dependent
on ERK signaling. PLoS One. 9:e960602014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Neuzillet C, Hammel P, Tijeras-Raballand
A, Couvelard A and Raymond E: Targeting the Ras-ERK pathway in
pancreatic adenocarcinoma. Cancer Metastasis Rev. 32:147–162. 2013.
View Article : Google Scholar
|
|
28
|
Nagini S: Carcinoma of the stomach: A
review of epidemiology, pathogenesis, molecular genetics and
chemoprevention. World J Gastrointest Oncol. 4:156–169. 2012.
View Article : Google Scholar : PubMed/NCBI
|