Introduction
Breast cancer is the most common cancer among women,
and its mortality rate ranks second only after lung cancer
(1). Metastasis is the main reason
for the mortality of breast cancer patients, and thus, it is
crucial to understand the molecular and cellular mechanisms that
cause primary tumors to metastasize.
RhoA is a prototypical member of the Rho GTPase
family, and plays an important role in the regulation of
cytoskeleton reorganization. RhoA regulates signal transduction
from cell surface receptors to intracellular target molecules and
is involved in a variety of biological processes, including cell
motility (2), morphogenesis
(3), cytokinesis (4,5) and
tumor progression (6,7). Rho proteins bind to and activate a
number of downstream effectors, leading to a cascade of biochemical
responses. Previous studies have found that RhoA is upregulated in
several types of human cancers, including colorectal (8,9) and
breast cancer (10,11), hepatocellular carcinoma (12,13),
gastric cancer (14,15), and alteration of the expression
levels of intracellular RhoA can directly affect the process of
invasion and metastasis in a variety of tumors (7,16,17).
MicroRNAs (miRNAs) are small, endogenous, non-coding
RNAs with 20–25 nucleotides that regulate the expression of
hundreds of target genes. miRNAs bind target mRNA sequences through
canonical base pairing between the seed sequence, which includes
nucleotides 2–8 from the 5′-end, and the complementary sequence
found in the 3′ untranslated region (3′UTR) of its target mRNA
(18–20). Recently, numerous studies have
demonstrated that miRNAs suppress the expression of cancer-related
genes and reduce tumorigenesis and metastasis in breast and several
other cancers (21–23).
miR-146a was found to act as a tumor suppressor by
directly regulating NF-κB expression and affecting tumor cell
signaling pathways (24). miR-146a
is involved in cell proliferation and influences cancer metastasis
in various types of tumors, including breast cancer (25–27).
However, the regulation of breast cancer metastasis by miR-146a is
a complex biological network. Therefore, the regulatory mechanisms
of breast cancer metastasis by miR-146a warrant further in-depth
study.
In the present study, among metastasis-related genes
as potential targets of miR-146a, an inverse correlation was noted
between RhoA and miR-146a expression in breast cancer cell lines.
Furthermore, we found that miR-146a specifically caused the
downregulation of RhoA expression. miR-146a was also found to
directly target the RhoA 3′UTR, thereby downregulating breast
cancer cell migration and invasion. The present study aimed to
elucidate the interrelation between miR-146a and RhoA, searching
for promising molecular targets to inhibit metastasis and potential
anti-metastatic agents for possible use in breast cancer
therapy.
Materials and methods
Cell culture
Human breast cancer cell lines, MCF-7 and
MDA-MB-231, were maintained in Dulbecco's modified Eagle's medium
(DMEM) supplemented with 10% fetal bovine serum (FBS) and 1%
antibiotics (Gibco, Carlsbad, CA, USA). The cells were incubated at
37°C in a 5% CO2 humidified atmosphere until reaching
75% confluency. To block the function of RhoA, the RhoA inhibitor
exoenzyme C3 transferase protein (C3) (Cytoskeleton, Denver, CO,
USA), was added to the culture medium at a final concentration of
2.0 µg/ml.
Transfection
miR-146a mimics, miR-146a inhibitor, and control
oligo were chemically synthesized by RiboBio (Guangzhou, Guangdong,
China). All transfections were carried out using Lipofectamine 2000
reagent (Invitrogen, Carlsbad, CA, USA), according to the
manufacturer's instructions. The MDA-MB-231 cells were harvested 24
h after transfection, and various analyses were performed. Three
independent experiments were carried out in duplicate.
Plasmid constructs
The 3′UTRs of the human RhoA gene (NM_001664) were
PCR amplified from human genomic DNA. The primers were digested and
cloned into the pmiR-RB-REPORT™ vector (RiboBio) at the XhoI
and NotI sites, and named pMIR-REPORT-RhoA-wt. The primer
sequences were: h-RhoA-3UTR-forward (Xhoi),
5′-CTTGACTCGAGAACCTTGCTGCAAGCACAG-3′ and h-RhoA-3UTR-reverse
(Noti), 5′-ATTGCGGCCGCTGCCTTTATTCTATTAGTAG-3′. Site-directed
mutagenesis with miR-146a target sites in the RhoA was carried out
using a site-directed mutagenesis kit (Takara, Dalian, Liaoning,
China), with pMIR-REPORT-RhoA-wt as a template, and named
pMIR-REPORT-RhoA-mut, respectively. The primer sequences were:
h-RhoA-3UTR-mut-forward, 5′-AGCCACGTCAAGAGATGGGACCCTCAGTCACAGAG-3′
and h-RhoA-3UTR-mut-reverse,
5′-TCCCATCTCTTGACGTGGCTCCTCTGGGAGGGAAC-3′.
Luciferase assay
MDA-MB-231 cells were seeded into a 24-well plate 24
h before transfection. They were transfected with 0.4 µg of
the pMiR-REPORT-RhoA-wt or pMiR-REPORT-RhoA-mut, 20 µM
miR-146a mimics or control oligo using Lipofectamine 2000. Firefly
and Renilla luciferase activities were measured
consecutively using a Dual-Luciferase assay (Promega, Madison, WI,
USA) according to the manufacturer's instructions. In addition,
0.02 µg of the Renilla luciferase vector pRL-TK
(Promega) was used for normalization. Three independent experiments
were performed in triplicate.
Quantitative real-time PCR analysis
(qRT-PCR)
Total RNA was prepared using TRIzol, and cDNA was
generated using the PrimerScript RT reagent kit and amplified using
RhoA primers with SYBR Premix Ex-Taq™ kit (all from Takara). The
primer sequences for RhoA were: 5′-TTTGGAGGTGGCATAGCCTT-3′
(forward) and 5′-ATGTTTAGTCAGCTGGAGAGAAGAG-3′ (reverse), and the
primer sequences for β-actin were: 5′-CCCTGGACTTCGAGCAAGAG-3′
(forward) and 5′-AATGCCAGGGTACATGGTGG-3′ (reverse). For analysis of
miRNA expression by qRT-PCR, the miRcute miRNA isolation kit,
miRcute miRNA First-Strand cDNA synthesis kit, miRcute miRNA qPCR
detection kit and miR-146a detection primer were purchased from
Tiangen Biotech (Beijing, China) to detect miR-146a and the control
miRNA (U6 snRNA). Amplification and detection were performed using
StepOne Plus QRT-PCR system (ABI, USA). β-actin was used to
normalize the expression of RhoA, whereas U6 was used to normalize
the expression of miR-146a; data were calculated based on the
following equation: RQ = 2−ΔΔCt.
Western blot analysis
Protein was extracted from breast cancer cell lines
using RIPA buffer [150 mM NaCl, 1% NP-40, 50 mM Tris-HCl (pH 7.4),
1 mM phenylmethysulfonyl fluoride, 1 µg/ml leupeptin, 1 mM
deoxycholic acid and 1 mM EDTA] including a protease inhibitor
cocktail and phosphatase inhibitors (Sigma, St. Louis, MO, USA),
according to the manufacturer's instructions. Equal amounts of
protein sample (50 µg) were separated by 12% SDS-PAGE and
transferred to polyvinylidene difluoride (PVDF) membranes using the
Bio-Rad semi-dry transfer system. Western blotting was performed
using anti-β-actin and anti-RhoA (CST, Denver, CO, USA). The
signals were detected with an ECL kit (Beyotime, Nantong, Jiangsu,
China) following the manufacturer's instructions.
Cell migration assay
Transwell invasion assays were performed using the
MDA-MB-231 cells, which were transfected with the miR-146a mimics,
miR-146a inhibitor and control oligo for 24 h. Then, the cells were
resuspended in serum-free DMEM, and placed in the top chamber with
a Matrigelcoated membrane. The lower portion of the chamber
contained 20% FBS as a chemoattractant. After the cells were
cultured for 24 h, we washed the chambers twice with
phosphate-buffered saline (PBS), and stained the chambers using
crystal violet. Five random fields were randomly selected, and
images were captured from each chamber. Then, the chambers were
decolorized by glacial acetic acid, and quantitative analysis was
carried out using the enzyme marker at 570 nm wavelength. Three
independent experiments were carried out in duplicate.
Wound healing assay
When cell confluency was <90% at 24 h after
transfection, wounds were created in the confluent cells using a
200-µl pipette tip, and any free-floating cells and debris
were removed by PBS. Medium was then added, and the culture plates
were incubated at 37°C. Wound healing within the scraped wound line
was observed at a 24-h time point, and representative scrape lines
for each cell line were photographed. The optical distance of the
wound was measured using ImageJ software. Three independent
experiments were carried out in duplicate.
Statistical analysis
All data are presented as mean ± SD of three
independent experiments. Assays for characterizing the phenotypes
of the cells were analyzed by Student's t-test or one-way ANOVA.
P-value of <0.05 was considered to indicate a statistically
significant result. All statistical analyses were performed using
the SPSS 22.0 software package (SPSS, Inc., Chicago, IL, USA).
Results
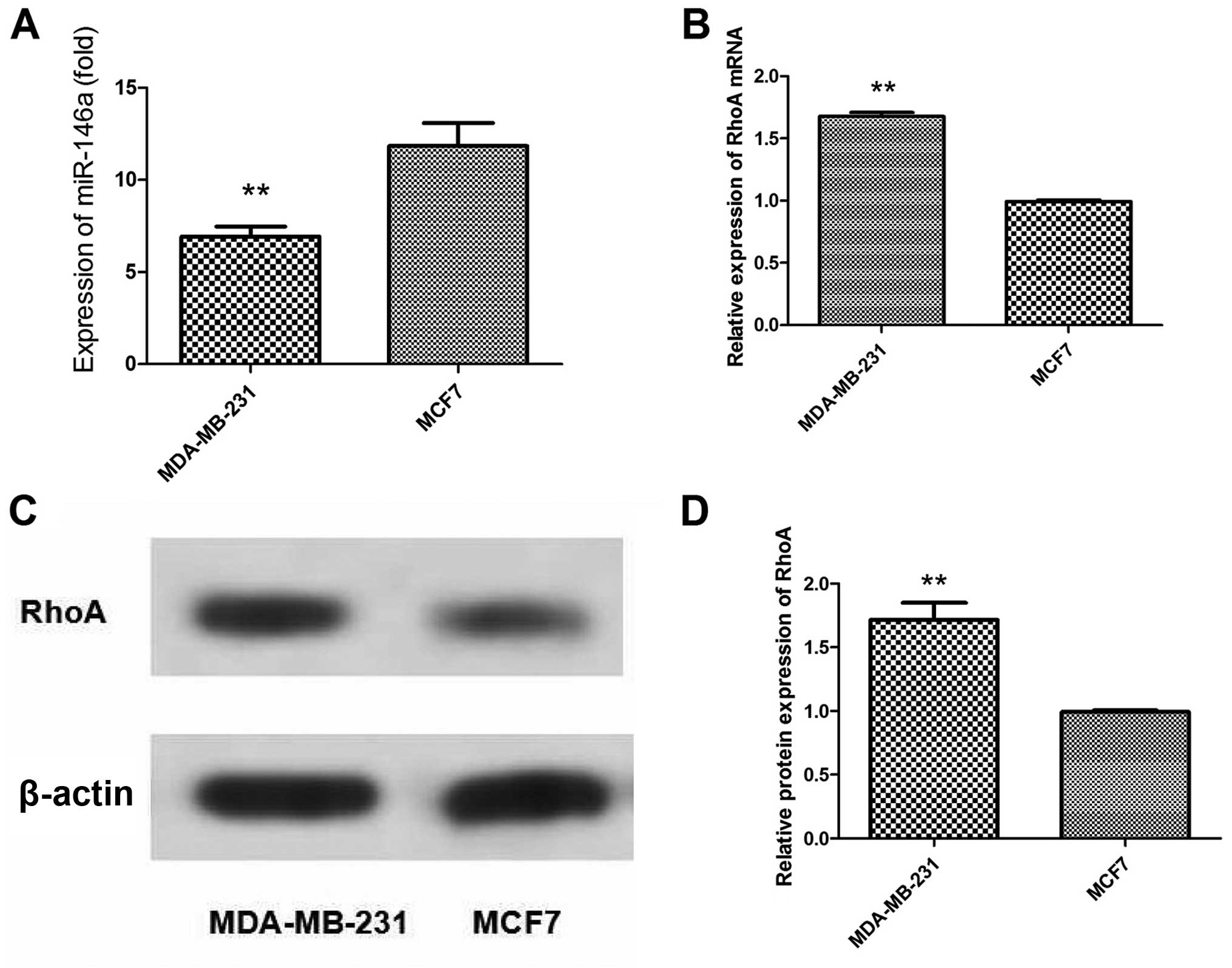
Differential expression of miR-146a and
RhoA in breast cancer cells with different metastatic activity
To assess whether miR-146a levels were altered in
breast cancer cell lines with different metastatic potential, two
breast-derived cell lines were used to assess the relative levels
of miR-146a. The basal expression level of miR-146a was generally
increased in the tumorigenic but weakly metastatic cell line MCF7,
compared with this level in the highly metastatic cell line
MDA-MB-231 (Fig. 1A). In contrast,
the expression levels of RhoA mRNA and protein in MCF7 cells were
obviously decreased when compared with these levels in the
MDA-MB-231 cells (Fig. 1B–D).
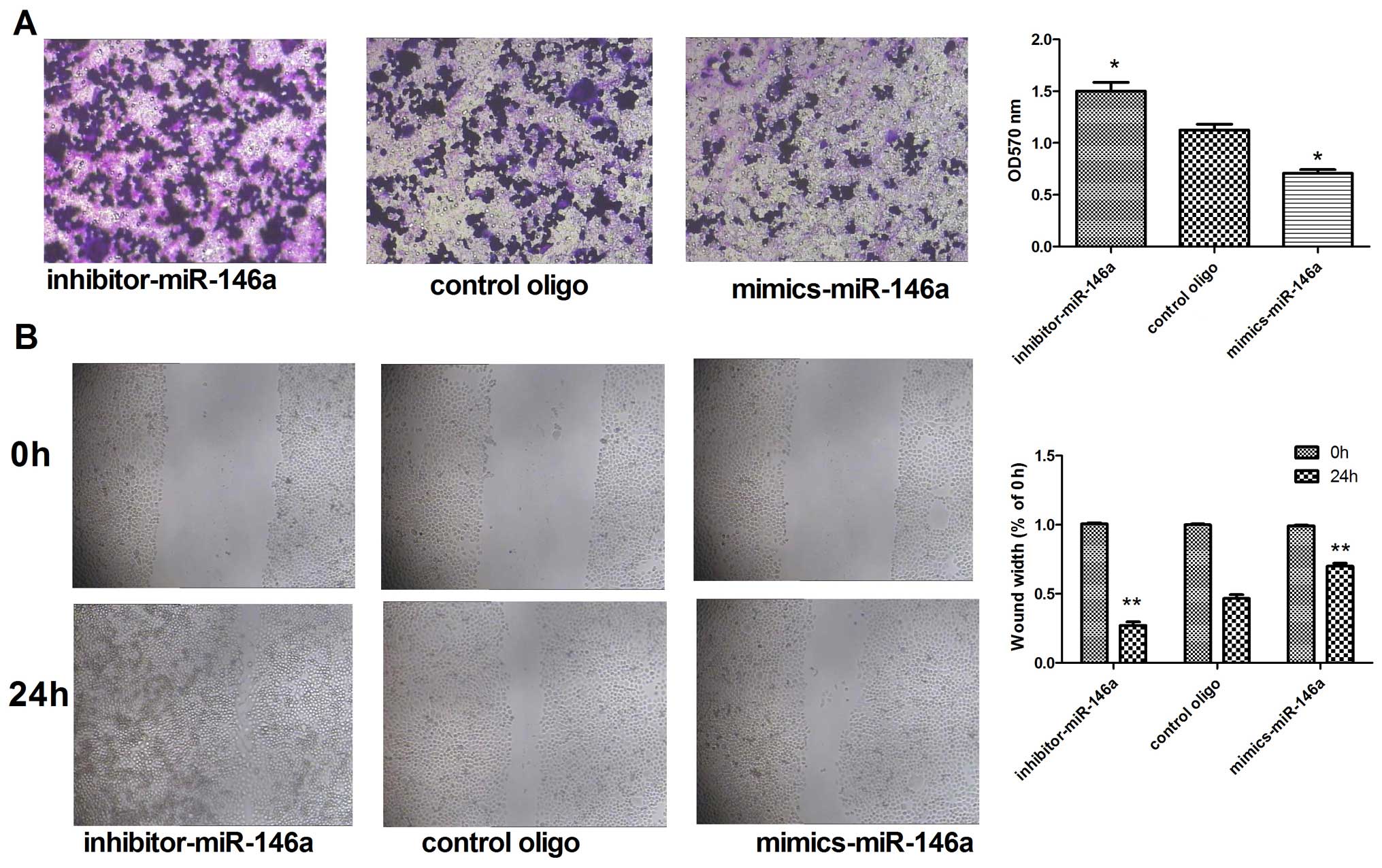
miR-146a suppresses metastatic activity
in the MDA-MB-231 cells
To confirm whether change in the expression of
miR-146a can change the invasion and metastasis capabilities of the
MDA-MB-231 cells, we upregulated miR-146a by transfecting the
MDA-MB-231 cells with miR-146a mimics, and downregulated miR-146a
by transfecting the MDA-MB-231 cells with the miR-146a inhibitor.
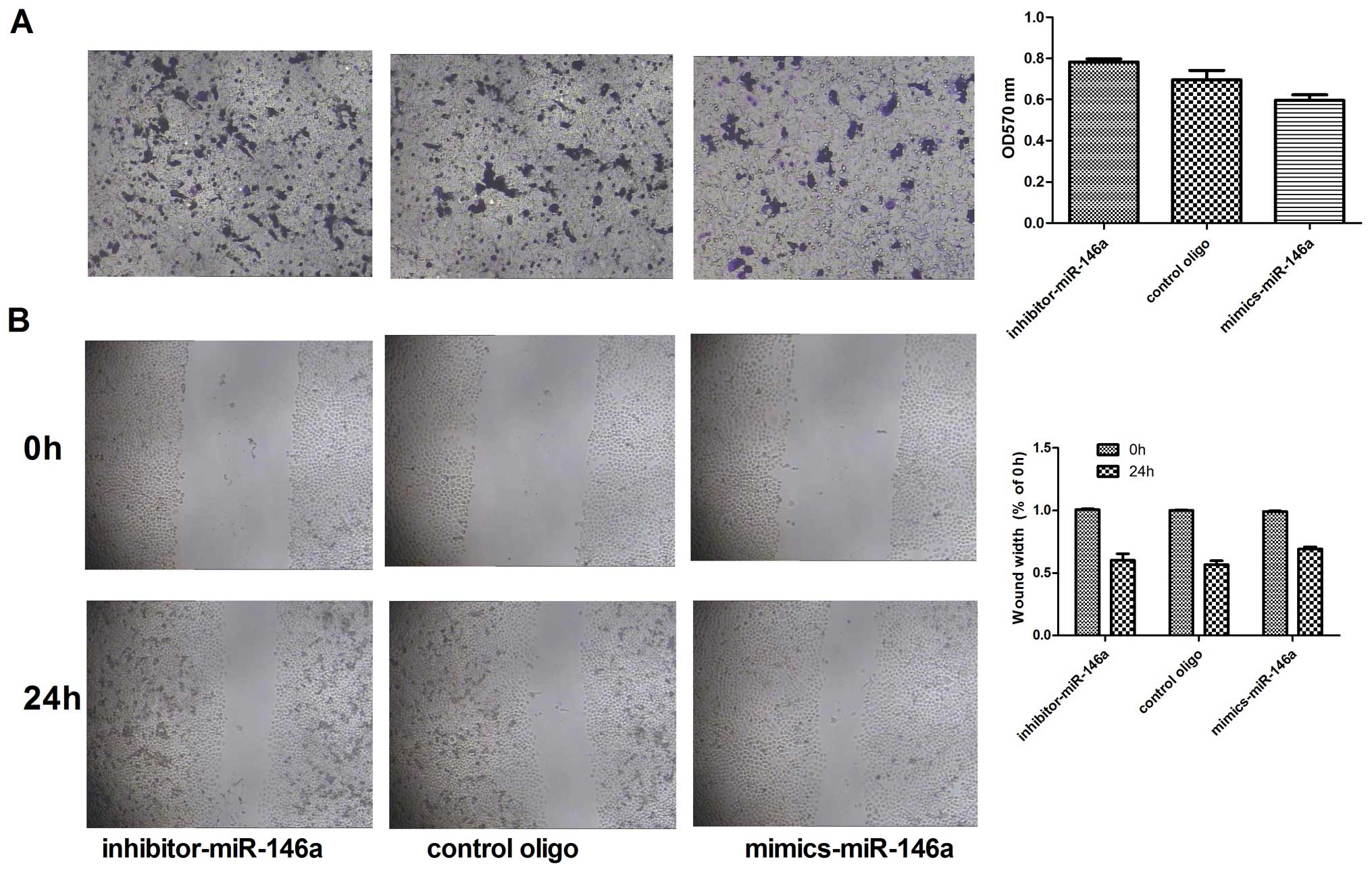
The results of the Transwell experiments showed that the number of
migrating cells was decreased after transfection with the miR-146a
mimics. In contrast, the number of migrating cells was increased
after transfection with the miR-146a inhibitor (Fig. 2A). A similar trend was observed in
the wound healing assays (Fig.
2B).
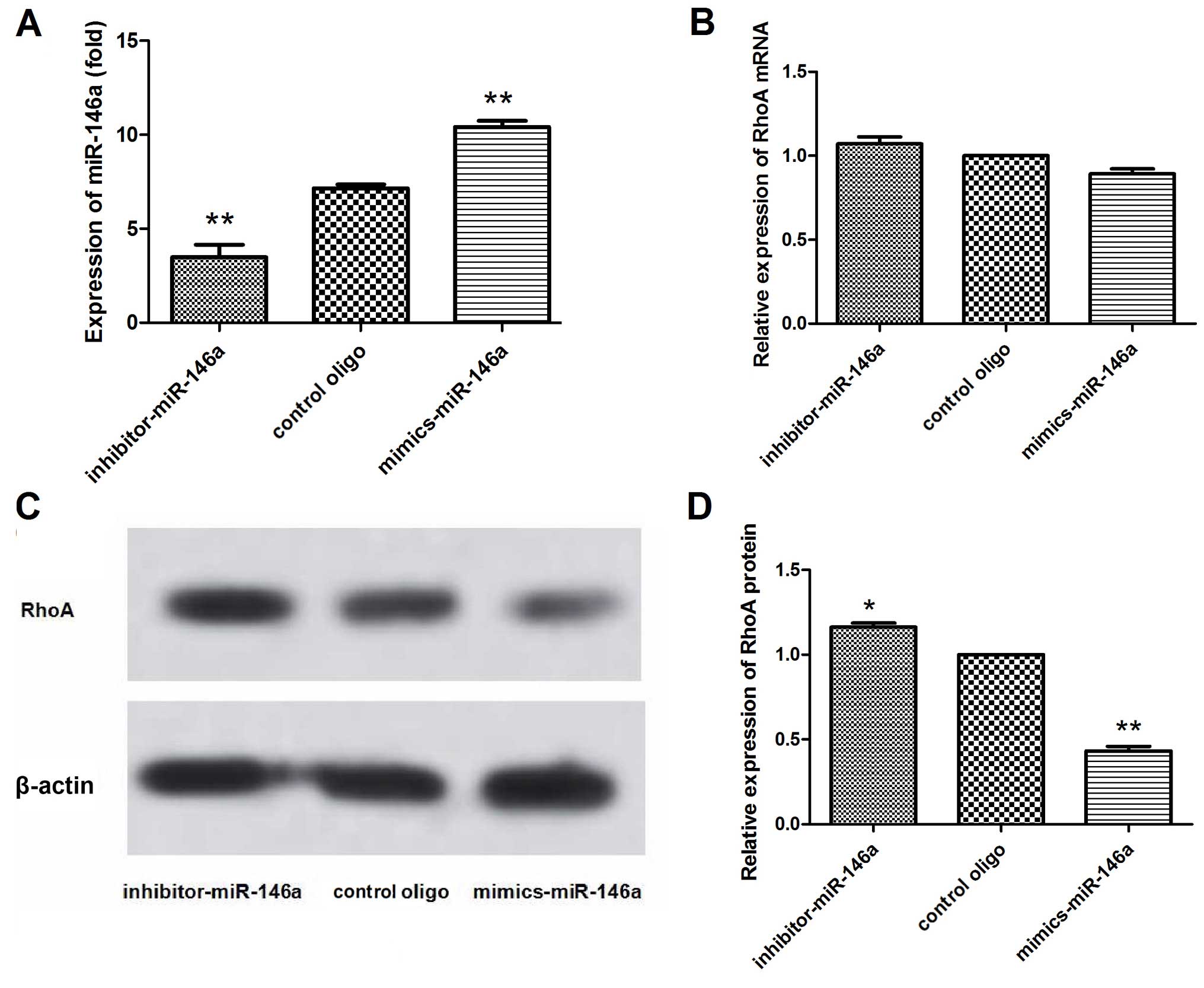
Expression of RhoA varies with the amount
of miR-146a expression in MDA-MB-231 cells
To confirm whether suppression of migration by
miR-146a is associated with changes in RhoA expression, we
transfected miR-146a mimics and the inhibitor into the MDA-MB-231
cells and detected the expression of RhoA at the mRNA and protein
levels by qRT-PCR and western blotting, respectively. The qRT-PCR
results showed that the expression of miR-146a was significantly
altered after transfection with the miR-146a mimics and inhibitor
in the MDA-MB-231 cells (Fig. 3A).
In addition, the qRT-PCR results showed that the expression of RhoA
mRNA was not significantly altered after transfection with the
miR-146a mimics and inhibitor in the MDA-MB-231 cells (Fig. 3B). However, in the western blot
analysis (Fig. 3C and D), the RhoA
protein concentration was decreased (P<0.01), when miR-146a was
upregulated. In contrast, the RhoA protein concentration was
increased (P<0.05), while miR-146a was downregulated. These
results indicated that miR-146a post-transcriptionally regulated
RhoA expression.
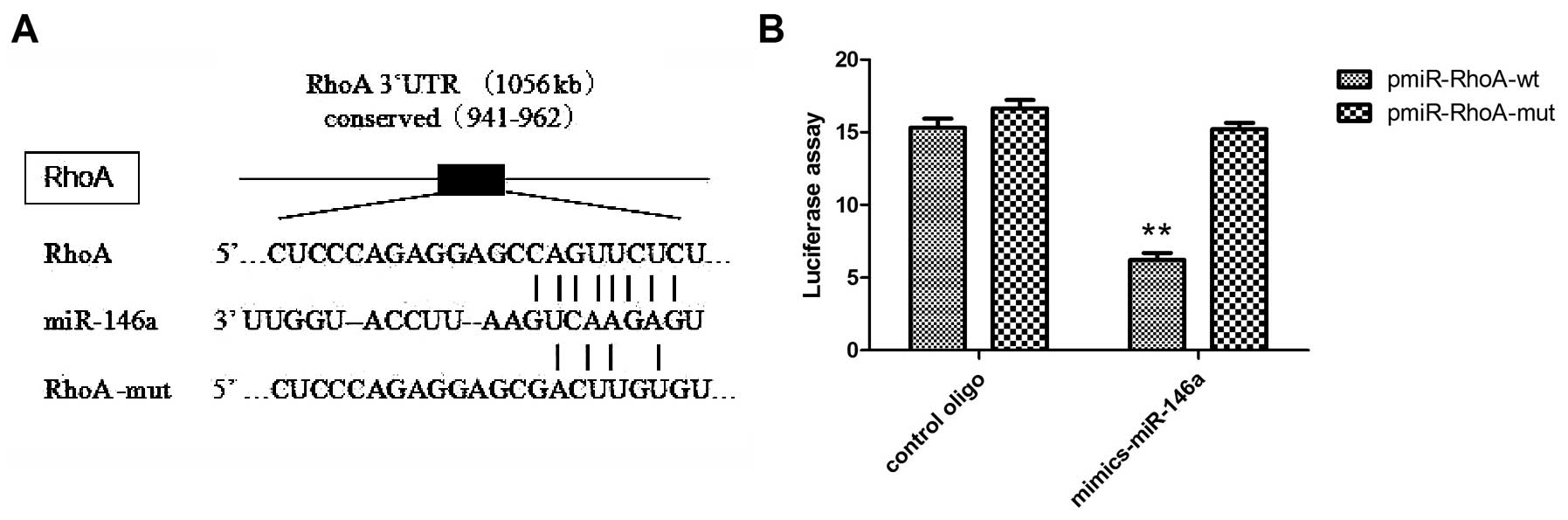
RhoA 3′UTR is a potential miR-146a target
gene
To study how miR-146a may regulate migration, we
proceeded to identify potential targets known to play a role in
cell mobility using miRBase databases. Among the candidates
surveyed, we found that the 3′UTR of the RhoA gene, which plays an
important role in cell junction formation and stabilization
(28,29), contains highly conserved regions
that may serve as a binding site for miR-146a as determined at
microrna.org (Fig.
4A).
To further demonstrate that RhoA is a potential
target of miR-146a, we generated luciferase reporters that
contained the 3′UTR of the RhoA gene. To determine whether the
seeding site responds to miR-146a, the reporter plasmids were
introduced into the MDA-MB-231 cells along with the miR-146a mimics
or the control oligo. The results showed that the reporter activity
was reduced by the ectopic expression of miR-146a (P<0.001;
Fig. 4B). We also generated
luciferase reporters that contained a mutated sequence within the
predicted target sites of the 3′UTR of the RhoA gene to further
demonstrate the interaction between miR-146a and the 3′UTR of RhoA.
Data showed that the reporter activity was not reduced by the
ectopic expression of miR-146a (Fig.
4B). Taken together, these results indicated that in MDA-MB-231
cells, the RhoA gene was a functional target of miR-146a.
Migratory ability exhibited no change
with the levels of miR-146a expression in RhoA inhibitor C3-treated
MDA-MB-231 cells
To further validate whether miR-146a inhibits
migration in a Rho-dependent manner, we blocked RhoA activity using
the Rho inhibitor C3 at a final concentration of 2.0 µg/ml.
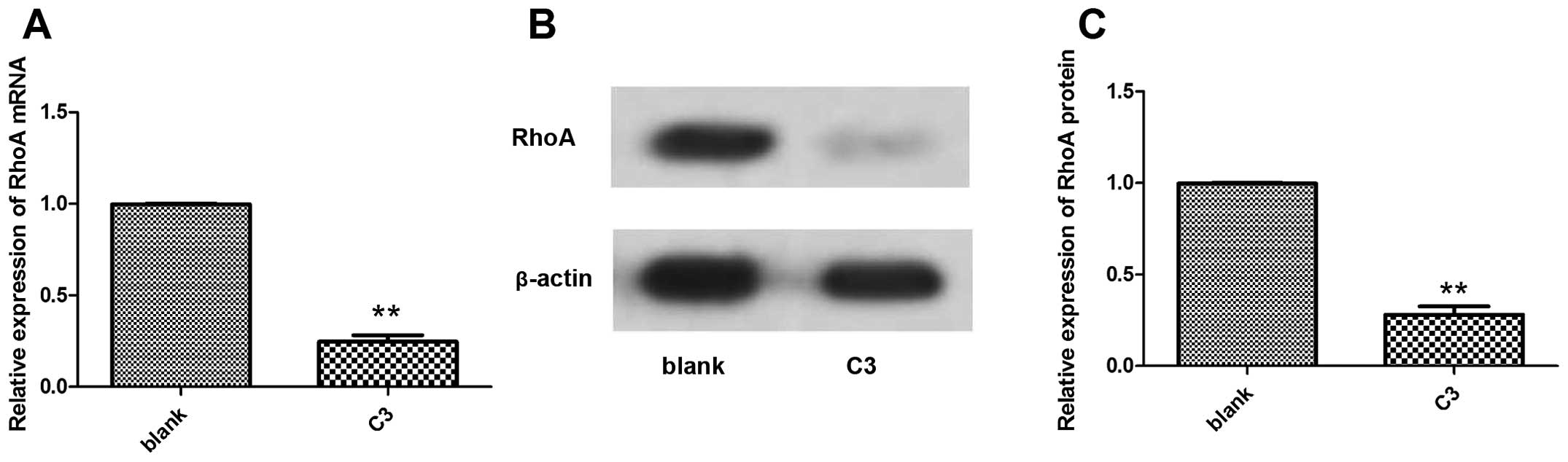
We found that the expression of RhoA mRNA and protein decreased
after treatment with C3 compared with the untreated cells (Fig. 5).
Moreover, after treatment with C3, the migratory
ability of the MDA-MB-231 cells transfected with the miR-146a
mimics or the miR-146a inhibitor was neither decreased nor
increased completely in the control oligo-transfected cells
(P<0.05; Fig. 6A and B). These
results suggest that miR-146a does not regulate RhoA when the RhoA
pathway is blocked.
Discussion
MicroRNAs (miRNAs) are a class of naturally existing
small non-coding RNAs generated from large transcripts, termed
pri-miRNAs and pre-miRNAs. In multicellular organisms, miRNA and
the 3′ untranslated region (3′UTR) of its target genes are
incompletely complementary, regulating the gene expression at a
post-transcriptional level and affecting almost all signaling
pathways, which are involved in a variety of physiological and
pathological processes and also play an important regulatory role
in the occurrence and progression of tumors. Research shows that
miRNAs have a role in the invasion and metastasis of tumor cells
(21–23). miRNAs are primarily negative gene
regulators of post-transcriptional repression (30), and their targets include tumor
suppressors and oncogenes (18–20).
Present studies have shown that miR-146a can inhibit
the occurrence, development and metastasis in gastric and
pancreatic cancer, and other tumors, while its specific mechanism
remains unclear (25–27,31–33).
At present, scholars have already conduct research on the
regulatory mechanism of miR-146a in breast cancer metastasis.
miR-146a was found to act as a tumor suppressor directly regulating
NF-κB expression, and affecting the tumor cell signaling pathway
(24). miR-146a is involved in cell
proliferation and influences cancer metastasis in various tumor
types, including breast cancer (25–27).
However, the regulation of breast cancer metastasis by miR-146a is
a complex biological network, and the regulatory mechanism of
breast cancer metastasis by miR-146a warrants further in-depth
study. The results of the present study led us to make conclusions
similar to these previous studies. We confirmed that the expression
of miR-146a differs in human breast cancer cell lines with
different metastatic potential, and miR-146a can reduce breast
cancer cell migration and invasion.
Rho/ROCK signaling channel has been researched in
regares to its involvement in the molecular mechanism of tumor
metastasis (34). Rho-subfamily
protein is one of the members of the Ras superfamily with GTP
enzyme activity, and mediates a variety of cellular effects, such
as cytoskeletal reorganization, membrane trafficking,
proliferation, apoptosis/survival, cell polarity, cell adhesion,
cell cycle and gene transcription (35–38).
As a key member of the Rho GTPase family, RhoA has been found to be
upregulated in several types of human cancers, including colorectal
(8,9) and breast cancer (10,11),
hepatocellular carcinoma (12,13)
and gastric cancer (14,15). Previous studies have found that
alteration of the expression levels of intracellular RhoA can
directly affect the process of invasion and metastasis in a variety
of tumors (7,16,17,20).
The results of our study suggest that miR-146a directly targets
RhoA, and may be involved in the migration of MDA-MB-231 cells.
Based on our data, we demonstrated that in the
breast cancer cell line MDA-MB-231, miR-146a directly regulates the
target gene RhoA by incomplete complementarity with the 3′UTR,
ultimately inhibiting the expression levels of the RhoA protein,
and thereby controling the migration of cells. We used
bioinformatics to determine that RhoA is closely associated with
cell migration and is a target gene regulated by miR-146a. In
addition, miR-146a targeting of RhoA 3′UTR was confirmed using
luciferase reporter gene containing RhoA 3′UTR or specific mutated
sites RhoA 3′UTR in the MDA-MB-231 cells.
The results confirmed our hypothesized that
upregulated expression of miR-146a negatively inhibited the
expression of RhoA protein, due to incomplete complementarity to
nucleotides within the 3′UTR of RhoA. Downregulation of miR-146a
expression led to increased expression of RhoA protein. However,
the level of RhoA mRNA had no significant changes. The
inconsistency of the degree of RhoA transcription and levels of
protein expression suggest that RhoA may be subject to
post-transcriptional regulation by miR-146a. According to the
experimental results, we can infer that miR-146a may act as an
anti-oncogene by suppressing MDA-MB-231 cell migration via the
post-transcriptional repression of RhoA. After we blocked RhoA
activity with the Rho inhibitor C3, we found that miR-146a was
unable to regulate RhoA. These results further validated the
possibility that miR-146a inhibited migration via a RhoA-dependent
pathway.
Collectively, we demonstrated that miR-146a
post-transcriptionally regulates the 3′UTR of RhoA mRNA and
inhibits expression of RhoA protein, ultimately leading to suppress
the migratory capacity of MDA-MB-231 cells. It is likely that
miR-146a is involved in the RhoA-associated pathway, which affects
the migration of the breast cancer cell line MDA-MB-231.
Acknowledgments
The present study was supported by grants from the
Zhejiang Provincial Natural Science Foundation (no. LQ13H160023),
the Zhejiang Provincial Natural Science Foundation (no. Y12H290042)
and the Population and Family Planning Scientific Research Project
of Zhejiang Province (no. 2014KYA237). We are grateful to Dr Cai
Zhijian (Immunology Research Institute of Zhejiang University,
Hangzhou, China) for providing the MDA-MB-231 and MCF-7 cell lines.
We are grateful to Dr Xiongfa Yang (Key Laboratory of Organosilicon
Chemistry and Material Technology of Education Ministry, Hangzhou
Normal University, Hangzhou, China) for the statistical analysis.
We also thank the members of our laboratory for the technical
assistance and useful suggestions.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zegers MM and Friedl P: Rho GTPases in
collective cell migration. Small GTPases. 5:e289972014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gonzalez-Billault C, Muñoz-Llancao P,
Henriquez DR, Wojnacki J, Conde C and Caceres A: The role of small
GTPases in neuronal morphogenesis and polarity. Cytoskeleton
Hoboken. 69:464–485. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chircop M: Rho GTPases as regulators of
mitosis and cytokinesis in mammalian cells. Small GTPases.
5(5)2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jordan SN and Canman JC: Rho GTPases in
animal cell cytokinesis: An occupation by the one percent.
Cytoskeleton Hoboken. 69:919–930. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
O'Connor K and Chen M: Dynamic functions
of RhoA in tumor cell migration and invasion. Small GTPases.
4:141–147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Orgaz JL, Herraiz C and Sanz-Moreno V: Rho
GTPases modulate malignant transformation of tumor cells. Small
GTPases. 5:e290192014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ariake K, Ohtsuka H, Motoi F, Douchi D,
Oikawa M, Rikiyama T, Fukase K, Katayose Y, Egawa S and Unno M:
GCF2/LRRFIP1 promotes colorectal cancer metastasis and liver
invasion through integrin-dependent RhoA activation. Cancer Lett.
325:99–107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai K, Mulatz K, Ard R, Nguyen T and Gee
SH: increased diacylglycerol kinase ζ expression in human
metastatic colon cancer cells augments Rho GTPase activity and
contributes to enhanced invasion. BMC Cancer. 14:2082014.
View Article : Google Scholar
|
|
10
|
Alho I, Costa L, Bicho M and Coelho C: Low
molecular weight protein tyrosine phosphatase isoforms regulate
breast cancer cells migration through a RhoA dependent mechanism.
PLoS One. 8:e763072013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dulong C, Fang YJ, Gest C, Zhou MH,
Patte-Mensah C, Mensah-Nyagan AG, Vannier JP, Lu H, Soria C, Cazin
L, et al: The small GTPase RhoA regulates the expression and
function of the sodium channel Nav1.5 in breast cancer cells. Int J
Oncol. 44:539–547. 2014.
|
|
12
|
Hu T, Guo H, Wang W, Yu S, Han L, Jiang L,
Ma J, Yang C, Guo Q and Nan K: Loss of p57 expression and RhoA
overexpression are associated with poor survival of patients with
hepatocellular carcinoma. Oncol Rep. 30:1707–1714. 2013.PubMed/NCBI
|
|
13
|
Wang SC, Lin XL, Li J, Zhang TT, Wang HY,
Shi JW, Yang S, Zhao WT, Xie RY, Wei F, et al: MicroRNA-122
triggers mesenchymal-epithelial transition and suppresses
hepatocellular carcinoma cell motility and invasion by targeting
RhoA. PLoS One. 9:e1013302014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Duan JT, Wang XM, Zhang SQ and Zhao GJ:
Effect of RhoA gene silencing on proliferation and migration of
gastric MGC-803 cells. Int J Clin Exp Med. 8:14410–14415.
2015.PubMed/NCBI
|
|
15
|
Zhou J, Hayakawa Y, Wang TC and Bass AJ:
RhoA mutations identified in diffuse gastric cancer. Cancer Cell.
26:9–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guan R, Xu X, Chen M, Hu H, Ge H, Wen S,
Zhou S and Pi R: Advances in the studies of roles of Rho/Rho-kinase
in diseases and the development of its inhibitors. Eur J Med Chem.
70:613–622. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu OM and Brown JH: G Protein-coupled
receptor and RhoA-stimulated transcriptional responses: Links to
inflammation, differentiation, and cell proliferation. Mol
Pharmacol. 88:171–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
De Ruyck K, Duprez F, Ferdinande L, Mbah
C, Rios-Velazquez E, Hoebers F, Praet M, Deron P, Bonte K, Speel
EJ, et al: A let-7 microRNA polymorphism in the KRAS3′-UTR is
prognostic in oropharyngeal cancer. Cancer Epidemiol. 38:591–598.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kong W, Yang H, He L, Zhao JJ, Coppola D,
Dalton WS and Cheng JQ: MicroRNA-155 is regulated by the
transforming growth factor beta/Smad pathway and contributes to
epithelial cell plasticity by targeting RhoA. Mol Cell Biol.
28:6773–6784. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu M, Lang N, Chen X, Tang Q, Liu S,
Huang J, Zheng Y and Bi F: miR-185 targets RhoA and Cdc42
expression and inhibits the proliferation potential of human
colorectal cells. Cancer Lett. 301:151–160. 2011. View Article : Google Scholar
|
|
21
|
Liu C and Tang DG: MicroRNA regulation of
cancer stem cells. Cancer Res. 71:5950–5954. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nair VS, Maeda LS and Ioannidis JP:
Clinical outcome prediction by microRNAs in human cancer: A
systematic review. J Natl Cancer Inst. 104:528–540. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang J, Ahmad A and Sarkar FH: The role of
microRNAs in breast cancer migration, invasion and metastasis. Int
J Mol Sci. 13:13414–13437. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bhaumik D, Scott GK, Schokrpur S, Patil
CK, Campisi J and Benz CC: Expression of microRNA-146 suppresses
NF-kappaB activity with reduction of metastatic potential in breast
cancer cells. Oncogene. 27:5643–5647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen G, Umelo IA, Lv S, Teugels E, Fostier
K, Kronenberger P, Dewaele A, Sadones J, Geers C and De Grève J:
miR-146a inhibits cell growth, cell migration and induces apoptosis
in non-small cell lung cancer cells. PLoS One. 8:e603172013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun Q, Zhao X, Liu X, Wang Y, Huang J,
Jiang B, Chen Q and Yu J: miR-146a functions as a tumor suppressor
in prostate cancer by targeting Rac1. Prostate. 74:1613–1621. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng T, Chou J, Zhang F, Liu Y, Ni H, Li
X, Zheng L, Tang T, Jin L and Xi T: CXCR4 3′UTR functions as a
ceRNA in promoting metastasis, proliferation and survival of MCF-7
cells by regulating miR-146a activity. Eur J Cell Biol. 94:458–469.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Citi S, Spadaro D, Schneider Y, Stutz J
and Pulimeno P: Regulation of small GTPases at epithelial cell-cell
junctions. Mol Membr Biol. 28:427–444. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wojnacki J, Quassollo G, Marzolo MP and
Cáceres A: Rho GTPases at the crossroad of signaling networks in
mammals: Impact of Rho-GTPases on microtubule organization and
dynamics. Small GTPases. 5:e284302014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lovat F, Valeri N and Croce CM: MicroRNAs
in the pathogenesis of cancer. Semin Oncol. 38:724–733. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hou Z, Xie L, Yu L, Qian X and Liu B:
MicroRNA-146a is downregulated in gastric cancer and regulates cell
proliferation and apoptosis. Med Oncol. 29:886–892. 2012.
View Article : Google Scholar
|
|
32
|
Li H, Xie S, Liu M, Chen Z, Liu X, Wang L,
Li D and Zhou Y: The clinical significance of downregulation of
mir-124-3p, mir-146a-5p, mir-155-5p and mir-335-5p in gastric
cancer tumorigenesis. Int J Oncol. 45:197–208. 2014.PubMed/NCBI
|
|
33
|
Li Y, Vandenboom TG II, Wang Z, Kong D,
Ali S, Philip PA and Sarkar FH: miR-146a suppresses invasion of
pancreatic cancer cells. Cancer Res. 70:1486–1495. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Matsuoka T and Yashiro M: Rho/ROCK
signaling in motility and metastasis of gastric cancer. World J
Gastroenterol. 20:13756–13766. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baranwal S and Alahari SK: Rho GTPase
effector functions in tumor cell invasion and metastasis. Curr Drug
Targets. 12:1194–1201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Duluc L and Wojciak-Stothard B: Rho
GTPases in the regulation of pulmonary vascular barrier function.
Cell Tissue Res. 355:675–685. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ellenbroek SI and Collard JG: Rho GTPases:
Functions and association with cancer. Clin Exp Metastasis.
24:657–672. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Street CA and Bryan BA: Rho kinase
proteins - pleiotropic modulators of cell survival and apoptosis.
Anticancer Res. 31:3645–3657. 2011.PubMed/NCBI
|