Introduction
Brachyury is the embryonic transcription factor of
the T-box family (1), which has
been reported as a prognostic factor in various types of carcinomas
such as gastric, colon, breast, prostate, esophageal, ovarian, and
particularly in non-small cell lung cancer (NSCLC), which is one of
the leading causes of cancer-related death worldwide (2–4).
Brachyury protein expression has been reported in 41% of primary
lung carcinomas, including 48% of adenocarcinomas and 25% of
squamous cell carcinomas (5).
Fernando et al (6) reported
that overexpression of brachyury in human carcinoma cells induced
changes characteristic of epithelial-mesenchymal transition (EMT),
including upregulation of mesenchymal markers, downregulation of
epithelial markers and an increase in cell migration and invasion.
Haro et al (2) examined 104
primary lung carcinoma samples and found that expression of
brachyury mRNA was significantly correlated with reduced 5-year
disease-free and overall survival rates and was significantly
correlated with vascular invasiveness, lymphatic permeation and
lymph node metastasis. Xu et al (7) showed that expression of brachyury was
significantly higher in 115 NSCLC tissue samples than that in
adjacent normal lung tissues.
Matrix metalloproteinases (MMPs) are zinc-dependent
endopeptidases and 23 members of the MMP family have been reported
to date. Although MMPs exhibit a wide range of biological functions
in cancer, their primary function is to degrade proteins in the
extracellular matrix (ECM) and mediate cell-cell adhesion, which
are implicated in EMT (8,9). Previous studies have reported that
MMP2 (10,11), MMP7 (12,13),
MMP9 (10,14), MMP10 (15), MMP12 (16,17),
MMP14 (18,19), MMP19 (20) and MMP26 (21) are significantly overexpressed in
NSCLC tissues and are correlated with poor patient prognosis.
MMPs and brachyury play an important role in lung
cancer cell invasion and metastasis. However, the correlation
between brachyury expression and MMP family genes has not been
investigated in non-small cell lung carcinomas. In the present
study, we aimed to explore the correlation between brachyury and
MMP members, and also further explore the mechanism involved in the
promotion of lung cancer metastasis by brachyury.
Materials and methods
Tumor cell lines and reagents
H1299, H460 and HEK293 cells in the present study
were purchased from the American Type Culture Collection (ATCC;
Gaithersburg, MD, USA) and propagated in RPMI-1640 medium or
Dulbecco's modified Eagle's medium (DMEM) (Gibco) supplemented with
10% fetal bovine serum (FBS) (Invitrogen) and 1% penicillin and
streptomycin in a cell incubator (37°C, 5% CO2).
The antibodies for MMP12 (cat. #ab52897;
Abcam®), brachyury (cat. #sc-17743 and cat. #sc-20109),
GAPDH (cat. #sc-32233), Hsp90 (cat. #sc-7949) (all from Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) were purchased and used
in the present study. HRP-goat anti-rabbit IgG and HRP-goat
anti-mouse IgG were purchased from Jackson ImmunoResearch
Laboratories. Transfection of DNA and RNAi was performed using
Lipofectamine 2000 from Life Technologies (cat. #11668019).
Plasmids and shRNAs
A lentivirus vector expressing human brachyury and
lentivirus particles were obtained from Shanghai R&S
Biotechnology (Shanghai, China). In brief, human brachyury cDNA
amplified from lung-derived cDNA samples was inserted into the
Pac1/Nhe1 restriction enzyme sites within the
multiple cloning site of the lentivirus vector
pLenti6.3-IRES2-EGFP/V5 DEST, resulting in the vector
pLenti-Brachyury-EGFP. Three siRNAs used to selectively silence the
expression of brachyury with the given sequences
(CCUCGAAUCCACAGUGATT, GCAAAUCCUCAUCCUCAGUTT and
CUCCAACCUAUUCUGACAATT) were synthesized by Shanghai GenePharma Co.,
Ltd., while the sequence for the negative control was
(UUCUCCGAACGUGUCACGUTT), as previously reported (22). The most efficient siRNA special for
Brachyury in the H460 cells line was selected for the following
shRNA construction (23), which
targets ORF of Brachyury (+1021/+1041 bp). Plenti6.3-IRES-EGFP/V5
DEST was used to construct Bra-shRNA and con-shRNA according to
shRNA construction protocol from Shanghai R&S Biotechnology
(Shanghai, China).
Transfection and culture of the stable
cells
Transfection of the PGL3-Basic and
pcDNA3.1-Flag-Brachyury plasmids was performed using Lipofectamine
2000 (cat. #11668019) following the manufacturer's instructions.
Basically, 5×105 H1299 cells were seeded in a 6-cm
culture dish overnight before transfection. Plasmid DNA (2
µg) and 6 µl Lipofectamine 2000 reagent were gently
mixed in 500 µl RPMI-1640 medium without serum separately at
room temperature for 5 min. The diluted DNA was combined with the
diluted Lipofectamine 2000 and incubated for 20 min. The mixture
was added into a culture dish drop by drop. Medium was replaced
after 24 h.
The constructed plenti 6.3-Brachyury/shRNA plasmids
and packaging mix (pLP1-Gag-pol, pLP-VSVG and pLP2-Rev) were
co-transfected into H293T cells using POLOdeliverer™ 3000
transfection reagent (cat. #CT001-1; Shanghai R&S
Biotechnology). The virus supernatant medium was collected after
H293T cell infection for 48 h. The mixture was centrifuged at 3,000
rpm × 15 min at 4°C. Then, the liquid was filtered using a
0.45-µm filter membrane. The acquired virus was stored at
−80°C until use. H1299-pBra (brachyury-overexpressing) and
H1299-pcon (negative control), H460-pshT (Brachyury-knockdown) and
H460-pshN stable cells were infected, and were then selected after
the processes of fluorescence activated cell sorting (GFP
selection) and Blasticidin treatment (cat. #60218ES50;
InvivoGen).
Semi-quantitative PCR and real-time
PCR
Reverse-transcription PCR was performed using the
SuperScript III One-Step RT-PCR system from Life Technologies (cat.
#1257418) following the protocol provided in the manual. Total RNA
(2 µg) was used for each reverse transcript reaction. The
mRNA expression of 23 MMP members were semi-quantitatively measured
via 2% agarose gel for H1299-pCon, H1299-pBra, H460-pshN and
H460-pshT cells. Each experiment was performed in duplicate and was
repeated 3 times. The MMP members were subjected to real-time PCR
using SYBR-Green Master Mix (Bio-Rad) and the Mx3005P-quantitative
RT-PCR system (Stratagene) according to the semi-quantitative PCR
results. Mean Ct values for the target genes were normalized to the
mean Ct values for the endogenous control 18s [−ΔCt = Ct (18s) − Ct
(target gene)]. The ratio of mRNA expression of the target gene vs.
18s was defined as 2−ΔCt.
Western blot analysis
Total protein extracts of each group were resolved
by 11% SDS-PAGE and transferred onto polyvinylidene difluoride
(PVDF) (Millipore) membranes. After blocking with 5% not-fat milk
for 1 h at room temperature, the PVDF membranes were washed 3 times
for 10 min with phosphate-buffered saline (PBS) followed by
incubation with the primary antibodies [goat anti-human brachyury
(1:8,000; 3% BSA dilution), rabbit anti-human MMP12 (1:250; 3%
no-fat milk dilution) and mouse anti-human HSP90 (1:4,000; 3% BSA
dilution)] overnight at 4°C. Following extensive washing, the
membranes were incubated with secondary antibodies for 2 h. After
washing 3 times for 10 min with Tris-buffered saline/Tween-20
(TBST) at room temperature once more, the immunoreactivity was
visualized using an ECL kit (cat. #SK6668-100; Sangon Biotech) and
the membranes were exposed to Kodak XAR-5 film (F5263;
Sigma-Aldrich, USA).
Matrigel invasion assay
Blind well chambers (Neuro Probe Inc.) with
8-µm pore polycarbonate filters coated with Matrigel (BD
Biosciences) were used. RPMI-1640 medium supplemented with 10% FBS
was added to the lower chambers; cells (1×105) were
added in serum-free medium to the upper chambers. After a 24-h
incubation at 37°C, the filters were fixed and stained with 0.1%
solution of crystal violet. Cells on the bottom side of the filters
were counted in five random ×100 objective microscopic fields.
Experiments were conducted in triplicate for each cell line. Two
MMP12 siRNAs were synthesized (Shanghai GenePharma, Co., Ltd.) as
previously reported (24). The
sequence for MMP12-siRNA1 was 5′-GCUGUUUUU AACCCACGUUTT-3′; and the
sequence for MMP12-siRNA2 was 5′-CCGUGAGGAUGUUGACUACTT-3′. Negative
control siRNA was also used.
Immunohistochemical staining
Immunohistochemical staining (IHC) was carried out
following a previously published protocol (25). Specimens were deparaffinized with
xylene, subjected to antigen repair, and were blocked for 15 min
with 5% goat serum. The slides were incubated with the
anti-brachyury (1:400 dilution) and anti-MMP12 (1;200 dilution)
antibodies at 4°C overnight, followed by incubation with
anti-rabbit second antibodies at room temperature for 30 min.
Positive expression of IHC was reflected as brown staining
estimated by averaging the numbers of positively stained cells
under ×200 high power vision fields. The demographic information of
the NSCLC cases used for IHC is provided in Table I. The present study was approved by
the Institutional Review Board of Shanghai Changzheng Hospital.
Informed consent was obtained before tissue collection.
Additionally, the Ethics Committee of Shanghai Changzheng Hospital
approved the use of the obtained patient specimens for this
study.
 | Table IDemographic data of the cases used
for immunohistochemistry. |
Table I
Demographic data of the cases used
for immunohistochemistry.
| Primary NSCLC | Metastatic
NSCLC |
|---|
| Mean age, in
years | 61.9 | 60.8 |
| Minimum age | 46 | 41 |
| Maximum age | 82 | 73 |
| Gender, n |
| Male | 25 | 17 |
| Female | 5 | 5 |
| Histological type,
n |
|
Adenocarcinoma | 14 | 16 |
| Squamous
carcinoma | 13 | 5 |
| Large cell
carcinoma | 3 | 1 |
ChIP assay
Brachyury transcription factor cDNA was
directionally cloned between XhoI and HindIII sites
of the pCDNA vector which contained an N-terminal flag tag.
Sequencing analysis was performed to verify the construct. H1299
cells were transfected with constructed pcDNA3.1-flag-Brachyury,
followed by chromatin immunoprecipitation assay (ChIP). Briefly,
nuclear proteins of the transfected H1299 cells were cross-linked
and sonicated to obtain DNA fragments of 200–1000 bp. The lysates
were diluted in ChIP dilution buffer and then incubated with salmon
sperm DNA/protein A agarose slurry (Invitrogen). The chromatin
solutions were incubated with anti-flag tag (Sigma) and RNA pol2
(cat. #sc-899; Santa Cruz Biotechnology) antibodies overnight at
4°C with rotation. Beads were pelleted by centrifugation. The IgG
was used as a negative control. PCR amplification of the genomic
fragments was performed with 10 specific primers flanking putative
binding sites on the MMP12 promoter. The PCR products were
separated by 2.0% agarose gel electrophoresis.
MMP12 luciferase reporter constructs and
luciferase assay
The brachyury promoter sequence (−1/−1800 bp) was
directionally cloned between the XhoI and HindIII
sites of the pGL3-basic vector with the following sense primer:
5′-CCGCTCGAGTGTGGCTTTGGATAAGTTAAGTTCA-3′ and antisense primer,
5′-CCCAAGCTTTGTAAACTTCTAAACGGATCAATTC-3′. Sequencing was performed
to verify the construct. Three subcloning vectors of point mutation
were performed using Site-Directed Mutagenesis kit (cat. #D0206;
Shanghai Beyotime, China) according to the manufacturer's protocol.
Primers of pGL3-mmp12-MT1 (mutation at −959/−953) and
pGL3-mmp12-MT2 (mutation at −923/−917) were:
5′-GAGGCAGCGAACTTATGGCTCGATTTGAAGGGGTGTTGA-3′ (forward) and
5′-TCAACACCCCTTCAAATCGAGCCATAAGTTCGCTGCCTC-3′ (reverse);
5′-TTGAGTAGAATCTATCCTCGTGCCGCAG CACCACTGCTT-3′ (forward), and
5′-AAGCAGTGGTGCTGCGGCACGAGGATAGATTCTACTCAA-3′ (reverse),
respectively. PGL3-mmp12-MT1+2 (mutation at −959/−953 and
−923/−917) was also constructed. These constructs were verified by
sequencing.
After transfection and/or treatment, the cells were
washed with PBS 3 times. The cells were then lysed in luciferase
cell culture lysis buffer provided with the Dual-Luciferase Assay
kit (Promega). After −80°C cryopreservation for 1 h and rotation at
room temperature for 20 min, the whole cell lysates were collected
into EP tubes and centrifuged at 4°C at 12,000 × g for 10 min. The
supernatant was collected in a fresh tube and 20 µl of the
supernatant was added to the luciferase assay substrate (20
µl). Luminescence was measured as relative light units,
twice for each lysate. Readings for the luciferase assay were
carried out using LUMIstar OPTIMA (BMG Labtech). Each assay was
repeated 3 times. Fold repression values were represented as mean
of three experiments.
Statistical analysis
All of the measurements were collected in triplicate
for each independent operation. The data were statistically
analyzed using the Student's t-test. Statistical analysis was
carried out using SPSS 16.0 statistical software (SPSS, Inc.,
Chicago, IL, USA). Statistical significance was set as a p-value
<0.05.
Results
MMP12 was significantly correlated with
brachyury expression in the NSCLC cell lines
T-box transcription factor brachyury has been
recently reported as a poor prognostic factor in various types of
carcinomas and is mainly characterized as a driver of EMT (5,6). MMP
family members play an important role in tumor cell metastasis
promoted by the brachyury transcription factor in NSCLC. However
the correlation between brachyury and MMPs remains unclear. We
constructed and selected two stable cell lines (H1299-pBra and
H460-pshT) and two negative control stable cell lines (H1299-pCon
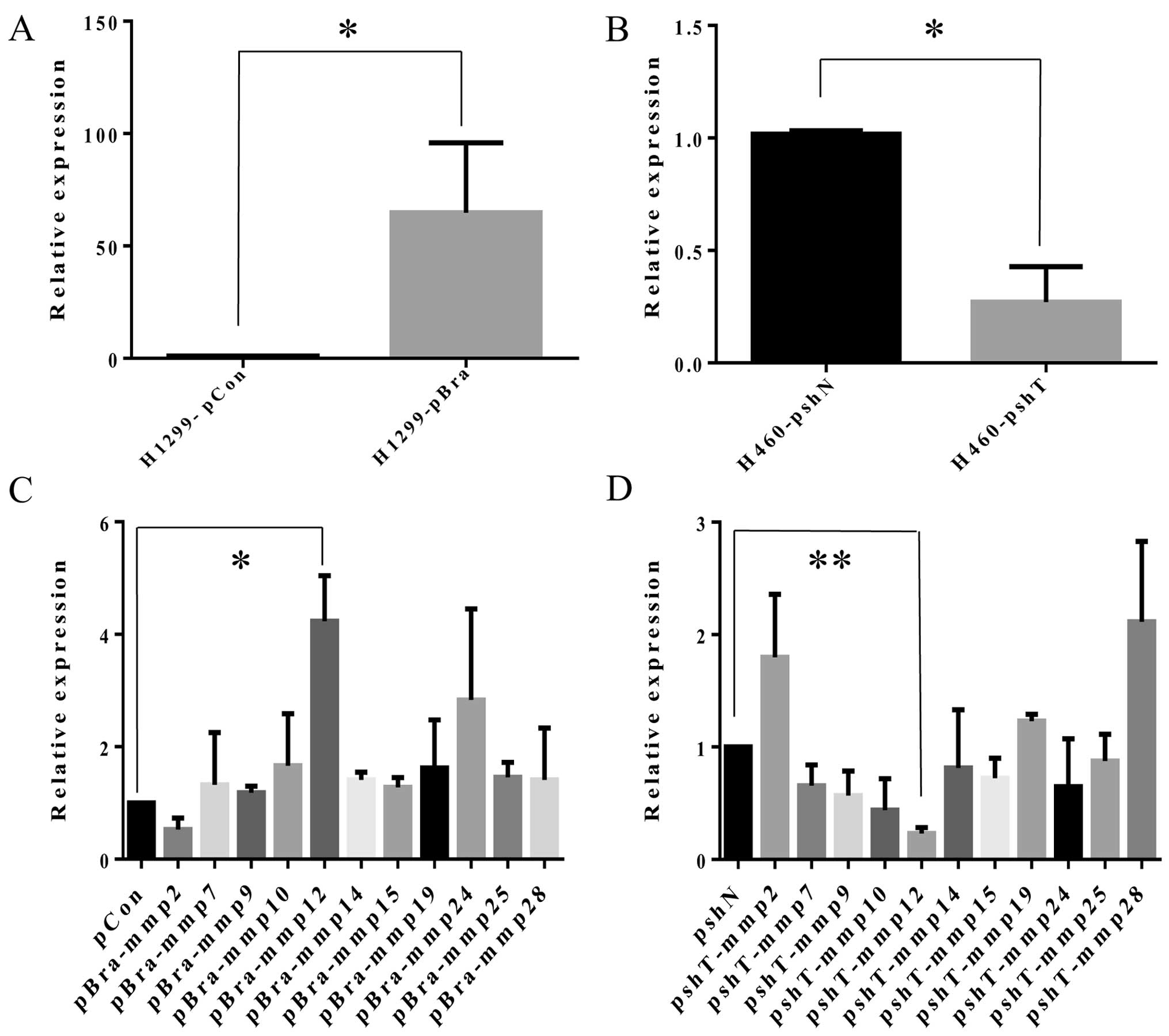
and H460-pshN). mRNA expression of brachyury in the H1299-pBra
cells was significantly increased by 64.8-fold (p<0.001),
compared with that in the H1299-pCon cells (Fig. 1A). mRNA expression of brachyury in
the H460-pshT cells was significantly reduced by 73% (p<0.001),
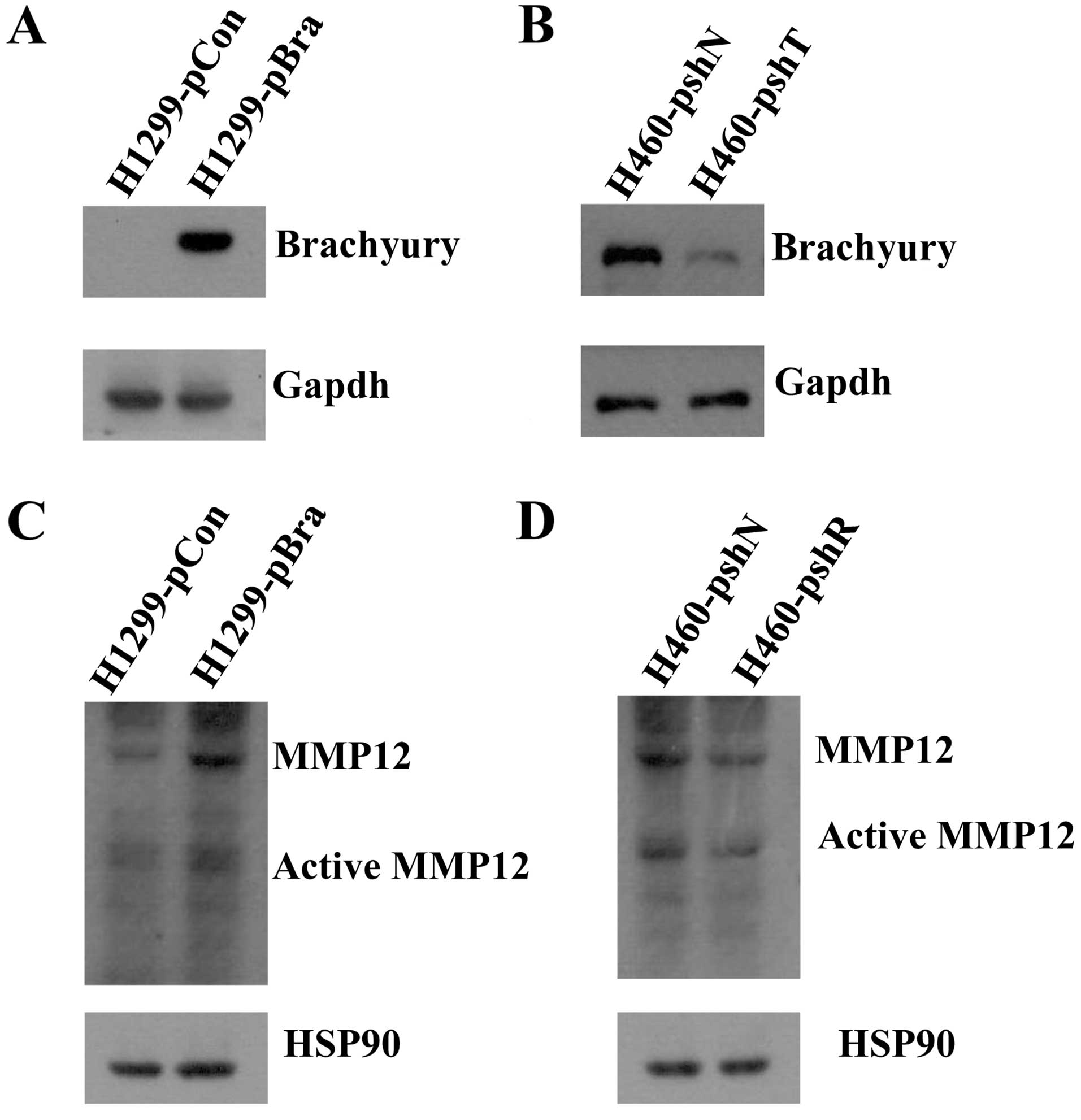
compared with that in the H1299-pCon cells (Fig. 1B). Protein expression of brachyury
in the H1299-pBra and H460-pshT cell lines also showed consistent
results (Fig. 2A and B).
After extraction of total RNA and reverse
transcription of cDNA from the 4 NSCLC stable cell lines
(H1299-pBra, H1299-pCon, H460-pshT and H460-pshN) we investigated
the mRNA expression levels of 23 MMP members by semi-quantitative
PCR. Furthermore, we selected 11 MMP members (MMP2, MMP7, MMP9,
MMP10, MMP12, MMP14, MMP15, MMP19, MMP24, MMP25 and MMP28) for
quantitative real-time PCR according to the agarose gel results
(data not shown), and reported MMP members with increased
expression in NSCLC tissues as reported in previous literature
(10–20). mRNA expression levels of MMPs in the
H1299-pBra cells were as follows. MMP2 exhibited a 47% decrease
(p=0.07), MMP7 exhibited a 32% increase (p=0.61), MMP9 exhibited a
18% increase (p=0.11), MMP10 exhibited a 66% increase (p=0.34),
MMP12 exhibited a 323% increase (p=0.02), MMP14 exhibited a 41%
increase (p=0.88), MMP15 exhibited a 28% increase (p=0.94), MMP19
exhibited a 62% increase (p=0.59), MMP24 exhibited a 183% increase
(p=0.40), MMP25 exhibited a 46% increase (p=0.86), MMP28 exhibited
a 8% increase (p=0.60), compared with the relative levels in the
negative control H1299-pCon cells (Fig.
1C). mRNA expression levels of MMPs in the H460-pBra cells were
as follows. MMP2 exhibited an 80% increase (p=0.13), MMP7 exhibited
a 34% decrease (p=0.09), MMP9 exhibited a 43% decrease (p=0.08),
MMP10 exhibited a 56% decrease (p=0.07), MMP12 exhibited a 77%
decrease (p<0.001), MMP14 exhibited a 19% decrease (p=0.68),
MMP15 exhibited a 28% decrease (p=0.77), MMP19 exhibited a 23%
increase (p=0.04), MMP24 exhibited a 35% decrease (p=0.29), MMP25
exhibited a 12% decrease (p=0.08), MMP28 exhibited a 110% increase
(p=0.06), compared with the levels in the negative control
H1299-pCon cells (Fig. 1D). We
concluded that the expression of MMP12 was the most significantly
correlated with the expression of brachyury. Consistent with the
mRNA expression of MMP12 and brachyury, we observed that the
protein level of MMP12 (54 kDa) and the active protein level of
MMP12 (45 kDa) were increased in the H1299-pBra cells as compared
to the H1299-pCon cells. The protein level of MMP12 and the active
protein level of MMP12 were decreased in the H460-pshT cells as
compared to the H460-pshN cells (Fig.
2C and D).
Brachyury promotes NSCLC cell invasion
via MMP12 expression
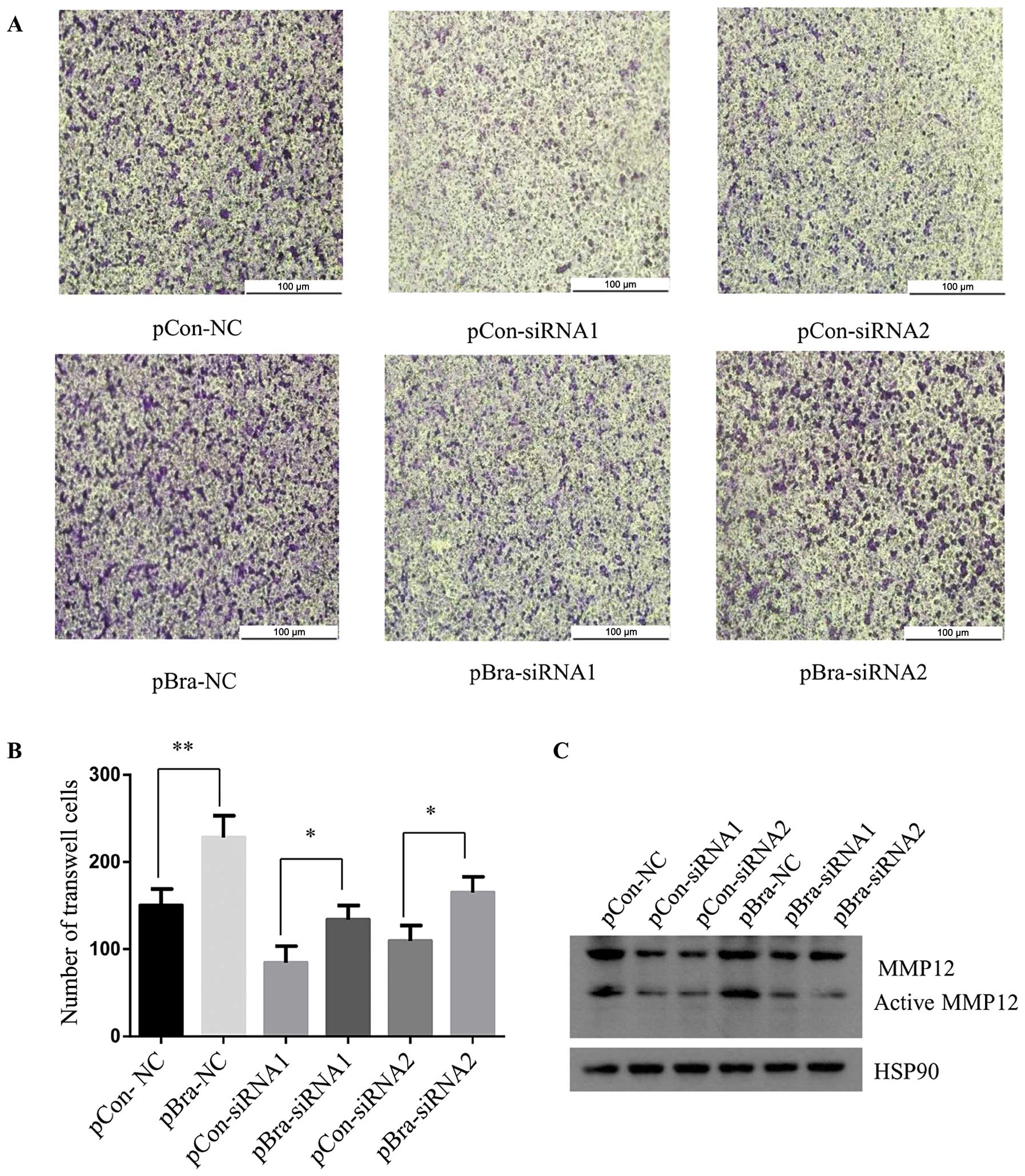
Next, we investigated the role of MMP12 in the
promotion of NSCLC cell invasion by brachyury. We synthesized two
MMP12 siRNAs for transfection into the H1299-pBra and H1299-pCon
stable cells.
As shown in Fig. 3C,
the MMP12 protein levels were reduced in the H1299-pCon and
H1299-pBra cell lines following transfection of the two synthesized
siRNAs targeting MMP12, compared to the levels in the cells
transfected with a control siRNA. Importantly, the H1299-pBra cells
displayed high protein expression of MMP12 compared with the level
in the H1299-pCon cells, when we performed the same MMP12-siRNA
treatment suggesting that brachyury promotes the expression of
MMP12.
Consistent with the protein expression between
brachyury and MMP12, the Matrigel invasion assay revealed higher
invasive ability in the H1299-pBra cells than that in the
H1299-pCon cells following treatment with MMP12-NC or MMP12-siRNA1
and MMP12-siRNA2 (Fig. 3).
Statistical analysis between the H1299-pCon-NC and H1299-pBra-NC
cells; between H1299-pCon-siRNA1 and H1299-pBra-siRNA1 cells;
between H1299-pCon-siRNA2 and H1299-pBra-siRNA2 cells, showed a
significant reduction in invasive ability (p<0.05) (Fig. 3B).
Expression of MMP12 is correlated with
the brachyury level in human primary lung cancer samples and
metastasis samples from lung cancer disease
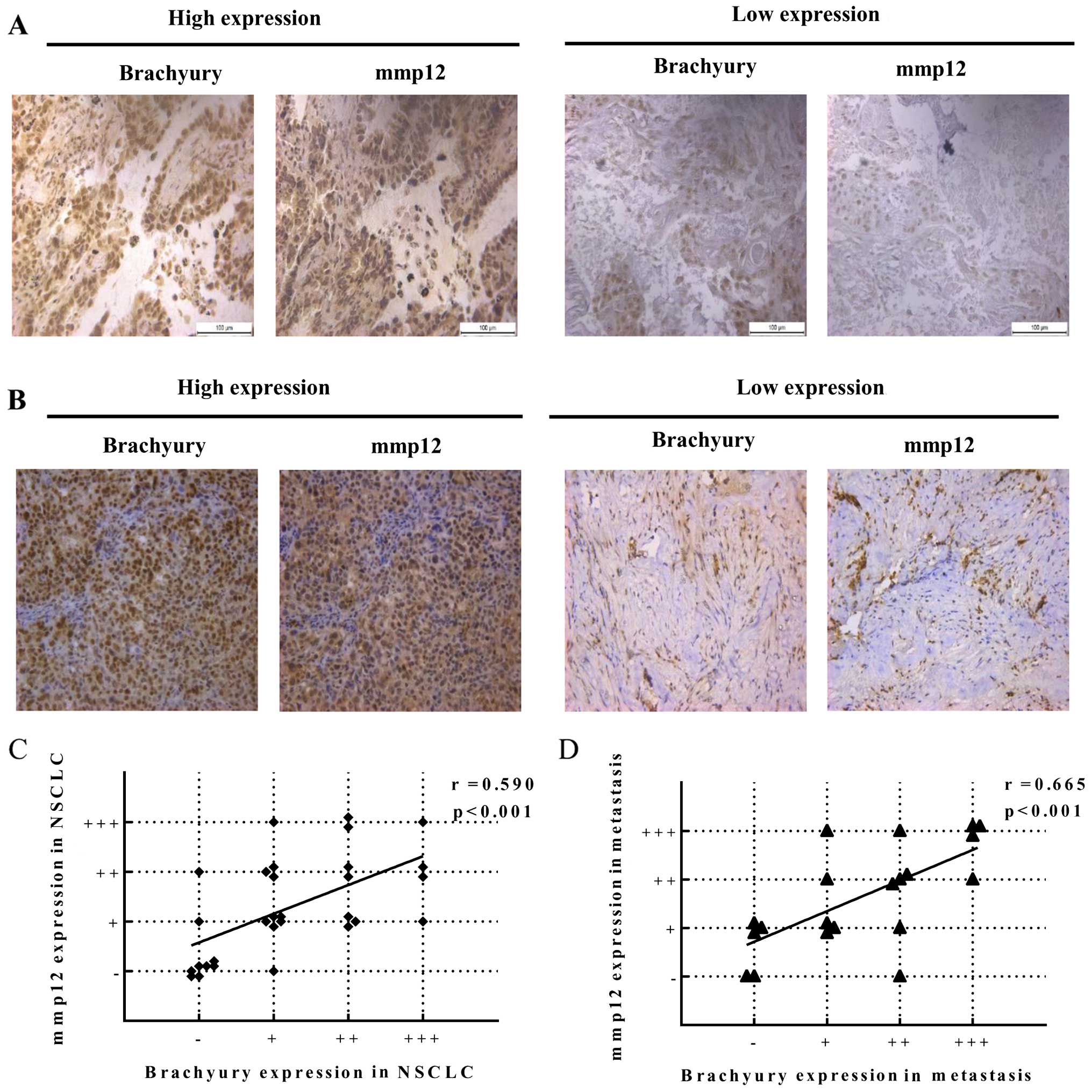
Expression levels of brachyury and MMP12 protein
were assessed by IHC in a series of 30 primary NSCLC carcinomas and
22 NSCLC metastatic tissues. Fig. 4A
and B shows representative results of the intensity scores
observed for brachyury and MMP12 expression, suggesting a positive
correlation between brachyury and MMP12 in the primary NSCLC
carcinomas and metastatic tissues. Brachyury expression exhibited a
high level in 4/30 cases, a medium level in 7/30 cases, a low level
in 10/30 cases and a negative level in 9/30 cases in the primary
NSCLC carcinoma tissues. MMP12 expression exhibited a high level in
4/30 cases, a medium level in 8/30 cases, a low level in 10/30
cases and a negative level in 8/30 cases. Furthermore, brachyury
expression exhibited a high level in 4/22 cases, a medium level in
6/22 cases, a low level in 5/22 cases and a negative level in 5/22
cases in the lung cancer metastatic tissues. MMP12 expression
exhibited a high level in 5/22 cases, a medium level in 5/22 cases,
a low level in 7/22 cases and a negative level in 3/22 cases. We
analyzed the correlation between brachyury and MMP12 in the same
sets of tumors by scatter and agreement plots. These plots
indicated that these two proteins were highly correlated in the
same sets of tumor tissues. Pearson's correlation coefficient was
used as a measure of correlation between brachyury and MMP12, which
was 0.59 for primary lung cancer tissues (Fig. 4C) and 0.665 for lung cancer
metastatic tissues (Fig. 4D).
Brachyury potentially regulates MMP12 in
NSCLC cells
Since the mRNA and protein expression levels of
MMP12 were correlated with brachyury, we sought to explore whether
brachyury regulates MMP12 in the invasion and metastasis of NSCLC
cells. We designed a CHIP-PCR for the MMP12 promoter in the H1299
cells transfected with the pcDNA3.1-Flag-Brachyury vector. The 5′
MMP12 promoter sequence (−3049 bp to −1 bp) was divided into 10
parts, and 10 pairs of primers were designed to amplify each part
of the promoter. The anti-Flag Tag antibody specifically pulled
down DNA fragments corresponding to the −1443 bp to −266 bp region
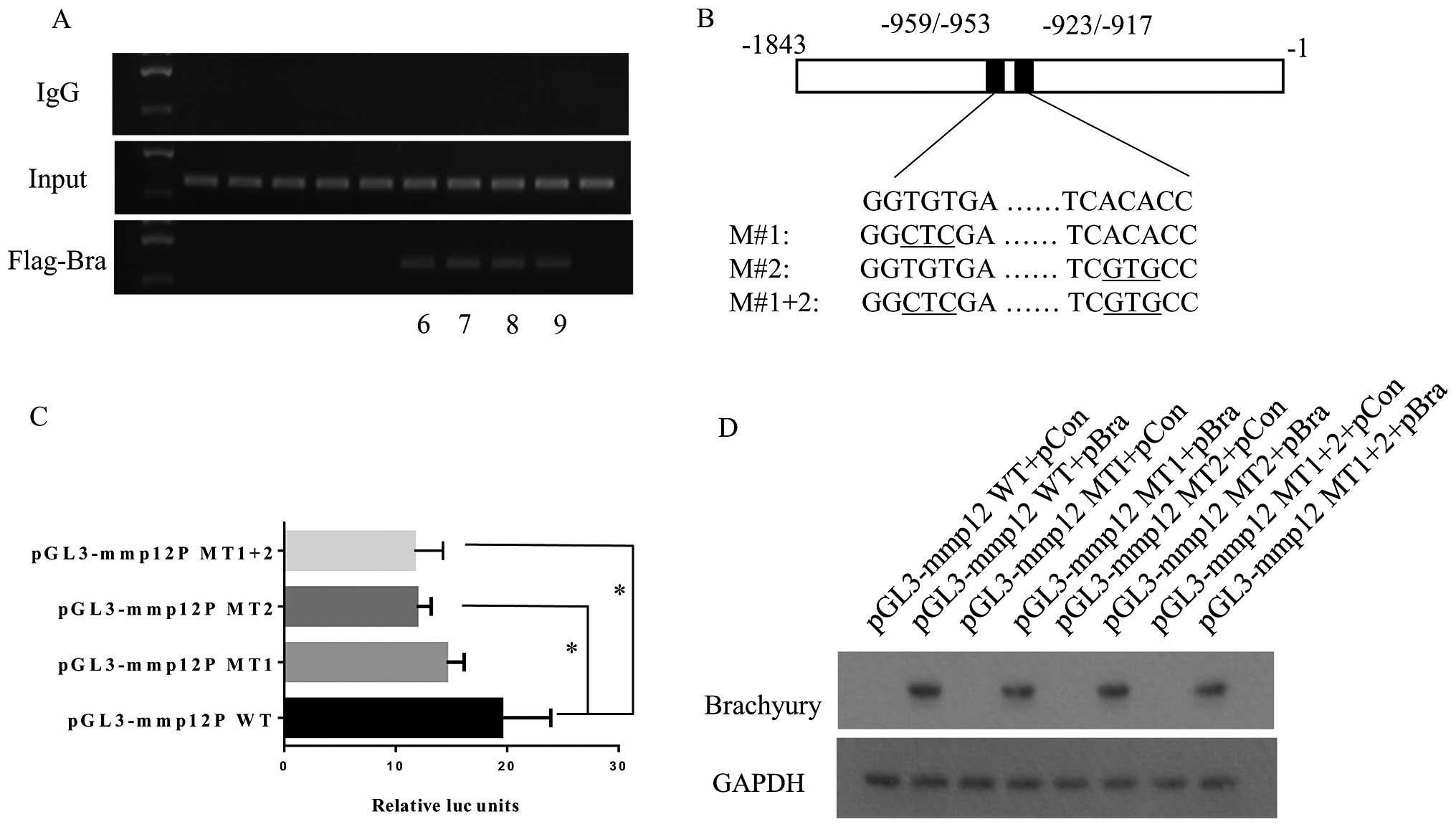
(Fig. 5A). This effect was not
observed when an irrelevant control IgG was used.
Brachyury and other T-box transcription family
members preferentially bind to the consensus elements AATTTCACAC
CTAGGTGTGAAATT (6). Bioinformatic
analysis revealed two putative half-site 5′-GGTGTGA-3′ (−959/−953
bp) and 5′-TCACACC-3′ (−923/−917 bp) within the region (Fig. 5B). Then, we performed 4 cloning
plasmids including pGL3-MMP12-WT (−1/−1,843 bp) and other three
point mutation pGL3-basic plasmids (pGL3-MMP12-MT1, pGL3-MMP12-MT2
and pGL3-MMP12-MT1+2) (Fig. 5B).
MMP12 promoter luciferase reporter assay showed that the point
mutations at each or both potential half-site of the consensus
elements reduced the luciferase units, and the second position
(−923/−917 bp) presented a more marked and significant reduction by
39.1% (p<0.05) (Fig. 5C). These
samples of the luciferase reporter assay were subjected to western
blot assay using GAPDH and brachyury antibodies (Fig. 5D).
Discussion
The product of the brachyury gene is a member of the
T-box family of transcription factors, which is always highly
present in various tumors yet is rarely observed in adult normal
tissues (26,27). The brachyury transcription factor
can promote tumor cell invasion and distant metastasis in various
types of cancer such as colorectal cancer, breast carcinoma,
prostate cancer and NSCLC, and is also an independent prognostic
factor in the clinic (4,28–30).
In the present study, we found overexpression of brachyury and
MMP12 in primary NSCLC and lung cancer metastatic tissues.
Furthermore, brachyury promoted lung cancer cell migration and
invasion mainly through upregulation of MMP12 in vitro.
Moreover we observed a positive correlation between brachyury and
MMP12 by investigating RNA and protein expression levels in
H1299-pBra and H460-pshT stable cells, and in primary NSCLC and
lung cancer metastatic patient tissues.
In accordance with our study, Roselli et al
(5) reported that the expression of
brachyury was significantly increased in human NSCLC samples
compared with the adult normal tissues by PCR and IHC techniques.
Haro et al (2) also found
that brachyury mRNA expression in primary lung carcinoma tissues
was significantly increased and was a predictor of poor prognosis.
Hofmann et al (31)
demonstrated that MMP12 expression was significantly increased in
tumors vs. corresponding lung tissues and MMP12 expression was
significantly correlated with local recurrence and metastatic
disease by examination of the expression of 22 MMPs in 89
surgically treated NSCLC patients. Kettunen et al (16) and Cho et al (17) also demonstrated that MMP12 was
significantly overexpressed in NSCLC patient tissues using c-DNA
microarray technique.
However, previous studies have not investigated the
expression of brachyury and MMP12 in lung cancer metastasis
tissues. In the present study, we also observed a significantly
positive correlation between brachyury and MMP12. The patients were
diagnosed with spine metastasis from NSCLC. Although it has been
debated for decades how cancer cells acquire metastatic capability,
distant metastatic cells not only have similar gene expression
profiles, but also possess various discordant malignant features,
compared with the primary carcinoma (32,33).
In line with our data, we observed a positive correlation of
brachyury and MMP12 between NSCLC metastasis and the normal control
lung tissues was more remarkable than between primary NSCLC tissues
and the normal ones.
In vitro we found that the transcription
factor brachyury had a significant effect on MMP12 expression and
the promotion of migration and invasion of tumor cells by brachyury
was associated with MMP12 in two adenocacinoma cell lines H460 and
H1299. Consistent with our findings, Fernando et al
(6) also found that brachyury
overepression significantly promoted the invasion in tumor cell
lines such as lung cancer cell line H460. Sarkar et al
(34) reported that brachyury was
expressed in a subpopulation of colorectal cancer cells that
resemble invasive front mesenchymal-like cells with characteristics
of cancer stem cells, allowing the cells to respond to signals
prompting invasion or metastasis. Kobayashi et al (29) showed that knockdown of the brachyury
transcription factor increased sensitivity of adenoid cystic
carcinoma cells to chemotherapy and radiation in vitro.
However, these previous studies did not deeply investigate the
correlation between brachyury as an EMT driver and MMPs which play
a crucial role in the process of tumor metastasis including the
process of EMT and degradation of ECM. In our experiments, we found
a distinct significant correlation between brachyury and MMP12 via
PCR experiments, furthermore verifying the association in
vivo. Different with our results, Fernando et al
(6) observed that the H460 cell
line with inhibition of brachyury expression concomitantly had
reduced expression of MMP2 and MMP24. We also observed that MMP24
expression was increased by 183% concomitant following brachyury
overexpression and was reduced by 35% concomitant with inhibiton of
brachyury expression.
In the present study, a high level of MMP12
expression was correlated with increased expression of brachyury in
NSCLC patients, suggesting that MMP12 is at least partially
responsible for the brachyury transcription factor-mediated
metastasis of NSCLC. CHIP and promoter luciferase assays, disclosed
that the segment sequence −216 to −1,083 bp relative to the
transcription start site of MMP12 with positive results of CHIP-PCR
and luciferase units was also significantly increased by 13.6-fold
when pGL3-basic vector was inserted the same segment sequence of
MMP12 promoter. The sequence of ʻAATTCACACCTAGGTGTGAAATTʼ is the
conservative element preferentially bound by brachyury and other
T-box transcription family (6).
Bioinformatic analysis showed two half sites (GGTGTGA and TCACACC)
concentrative to −862 to −820 bp relative to the transcription
start site of MMP12, which possibly were the binding sites by
brachyury transcription factor complex. Point mutation and
luciferase reporter assay showed point mutation at the MMP12
promoter sequence (−923/−917 bp) can significantly cause luciferase
units with 39.1% reduction.
Recent studies have reported that brachyury is a
driver of EMT, which allows the conversion of epithelial polarized
and stationary cells into highly motile and invasive mesenchymal
cells (5,6,35).
Fernando et al (6) reported
that brachyury is able to bind to the E-cadherin promoter promoting
EMT, an effect partially mediated by Slug. The present study
contributes to further clarify the role and pathological mechanism
of brachyury transcription factor in NSCLC migration and
metastasis.
In conclusion, we showed for the first time that
brachyury upregulated MMP12 in NSCLC tumors promoting cancer cell
migration and invasion. Our data thus suggest brachyury as a
potential therapeutic target for NSCLC. However, the detailed
regulatory mechanisms in regards to MMP12 still require further
investigation.
Acknowledgments
The present study was supported by the National
Nature Science Foundation of China (81272943 and 81102036), and the
Science and Technology Commission of Shanghai Municipality
(12JC1411300)..
References
|
1
|
Kispert A, Koschorz B and Herrmann BG: The
T protein encoded by Brachyury is a tissue-specific transcription
factor. EMBO J. 14:4763–4772. 1995.PubMed/NCBI
|
|
2
|
Haro A, Yano T, Kohno M, Yoshida T, Koga
T, Okamoto T, Takenoyama M and Maehara Y: Expression of Brachyury
gene is a significant prognostic factor for primary lung carcinoma.
Ann Surg Oncol. 20(Suppl 3): S509–S516. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Imajyo I, Sugiura T, Kobayashi Y, Shimoda
M, Ishii K, Akimoto N, Yoshihama N, Kobayashi I and Mori Y: T-box
transcription factor Brachyury expression is correlated with
epithelial-mesenchymal transition and lymph node metastasis in oral
squamous cell carcinoma. Int J Oncol. 41:1985–1995. 2012.PubMed/NCBI
|
|
4
|
Kilic N, Feldhaus S, Kilic E, Tennstedt P,
Wicklein D, Wasielewski R, Viebahn C, Kreipe H and Schumacher U:
Brachyury expression predicts poor prognosis at early stages of
colorectal cancer. Eur J Cancer. 47:1080–1085. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roselli M, Fernando RI, Guadagni F, Spila
A, Alessandroni J, Palmirotta R, Costarelli L, Litzinger M,
Hamilton D, Huang B, et al: Brachyury, a driver of the
epithelial-mesenchymal transition, is overexpressed in human lung
tumors: An opportunity for novel interventions against lung cancer.
Clin Cancer Res. 18:3868–3879. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fernando RI, Litzinger M, Trono P,
Hamilton DH, Schlom J and Palena C: The T-box transcription factor
Brachyury promotes epithelial-mesenchymal transition in human tumor
cells. J Clin Invest. 120:533–544. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu K, Liu B and Liu Y: Impact of Brachyury
on epithelial-mesenchymal transitions and chemosensitivity in
non-small cell lung cancer. Mol Med Rep. 12:995–1001.
2015.PubMed/NCBI
|
|
8
|
Kleiner DE and Stetler-Stevenson WG:
Matrix metalloproteinases and metastasis. Cancer Chemother
Pharmacol. 43(Suppl): S42–S51. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sternlicht MD and Werb Z: How matrix
metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol.
17:463–516. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang R, Ke ZF, Wang F, Zhang WH, Wang YF,
Li SH and Wang LT: GOLPH3 overexpression is closely correlated with
poor prognosis in human non-small cell lung cancer and mediates its
metastasis through upregulating MMP-2 and MMP-9. Cell Physiol
Biochem. 35:969–982. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ali-Labib R, Louka ML, Galal IH and Tarek
M: Evaluation of matrix metalloproteinase-2 in lung cancer.
Proteomics Clin Appl. 8:251–257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang J, Luo J, Ni J, Tang L, Zhang HP,
Zhang L, Xu JF and Zheng D: MMP-7 is upregulated by COX-2 and
promotes proliferation and invasion of lung adenocarcinoma cells.
Eur J Histochem. 58:22622014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang Y, Guo S and Zhou Q: Prognostic
value of matrix metalloproteinase-7 expression in patients with
non-small cell lung cancer. Tumour Biol. 35:3717–3724. 2014.
View Article : Google Scholar
|
|
14
|
Ruiz-Morales JM, Dorantes-Heredia R,
Arrieta O, Chavez-Tapia NC and Motola-Kuba D: Neutrophil
gelatinase-associated lipocalin (NGAL) and matrix
metalloproteinase-9 (MMP-9) prognostic value in lung
adenocarcinoma. Tumour Biol. 36:3601–3610. 2015. View Article : Google Scholar
|
|
15
|
Gill JH, Kirwan IG, Seargent JM, Martin
SW, Tijani S, Anikin VA, Mearns AJ, Bibby MC, Anthoney A and
Loadman PM: MMP-10 is overexpressed, proteolytically active, and a
potential target for therapeutic intervention in human lung
carcinomas. Neoplasia. 6:777–785. 2004. View Article : Google Scholar
|
|
16
|
Kettunen E, Anttila S, Seppänen JK,
Karjalainen A, Edgren H, Lindström I, Salovaara R, Nissén AM, Salo
J, Mattson K, et al: Differentially expressed genes in nonsmall
cell lung cancer: Expression profiling of cancer-related genes in
squamous cell lung cancer. Cancer Genet Cytogenet. 149:98–106.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cho NH, Hong KP, Hong SH, Kang S, Chung KY
and Cho SH: MMP expression profiling in recurred stage IB lung
cancer. Oncogene. 23:845–851. 2004. View Article : Google Scholar
|
|
18
|
Wang YZ, Wu KP, Wu AB, Yang ZC, Li JM, Mo
YL, Xu M, Wu B and Yang ZX: MMP-14 overexpression correlates with
poor prognosis in non-small cell lung cancer. Tumour Biol.
35:9815–9821. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou H, Wu A, Fu W, Lv Z and Zhang Z:
Significance of semaphorin-3A and MMP-14 protein expression in
non-small cell lung cancer. Oncol Lett. 7:1395–1400.
2014.PubMed/NCBI
|
|
20
|
Yu G, Herazo-Maya JD, Nukui T, Romkes M,
Parwani A, Juan-Guardela BM, Robertson J, Gauldie J, Siegfried JM,
Kaminski N, et al: Matrix metalloproteinase-19 promotes metastatic
behavior in vitro and is associated with increased mortality in
non-small cell lung cancer. Am J Respir Crit Care Med. 190:780–790.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Zhao H, Wang Y, Lin Y, Tan Y,
Fang X and Zheng L: Non-small cell lung cancer invasion and
metastasis promoted by MMP-26. Mol Med Rep. 4:1201–1209.
2011.PubMed/NCBI
|
|
22
|
Li J, Zhu L, Qu X, Li J, Lin R, Liao L,
Wang J, Wang S, Xu Q and Zhao RC: Stepwise differentiation of human
adipose-derived mesenchymal stem cells toward definitive endoderm
and pancreatic progenitor cells by mimicking pancreatic development
in vivo. Stem Cells Dev. 22:1576–1587. 2013. View Article : Google Scholar
|
|
23
|
Wang HZ, He YX, Yang CJ, Zhou W and Zou
CG: Hepcidin is regulated during blood-stage malaria and plays a
protective role in malaria infection. J Immunol. 187:6410–6416.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sarkar S, Nuttall RK, Liu S, Edwards DR
and Yong VW: Tenascin-C stimulates glioma cell invasion through
matrix metalloproteinase-12. Cancer Res. 66:11771–11780. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Edwards YH, Putt W, Lekoape KM, Stott D,
Fox M, Hopkinson DA and Sowden J: The human homolog T of the mouse
T(Brachyury) gene; Gene structure, cDNA sequence, and assignment to
chromosome 6q27. Genome Res. 6:226–233. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ali A, Wang Z, Fu J, Ji L, Liu J, Li L,
Wang H, Chen J, Caulin C, Myers JN, Zhang P, Xiao J, Zhang B and Li
X: Differential regulation of the REGγ-proteasome pathway by
p53/TGF-β signalling and mutant p53 in cancer cells. Nat Commun.
4:26672013. View Article : Google Scholar
|
|
27
|
Krukovskaia LL, Polev DE, Nosova Iu K,
Baranova AV, Koliubaeva SN and Kozlov AP: Investigation of
transcription factor Brachyury (T) expression in human normal and
tumor tissues. Vopr Onkol. 54:739–743. 2008.In Russian.
|
|
28
|
Palena C, Fernando RI, Hamilton DH, Huang
B and Schlom J: Brachyury, a driver of tumor invasiveness and
resistance to multiple therapies, is a novel immunotherapy target.
J Immunother Cancer. 1(Suppl 1): P2302013. View Article : Google Scholar
|
|
29
|
Kobayashi Y, Sugiura T, Imajyo I, Shimoda
M, Ishii K, Akimoto N, Yoshihama N and Mori Y: Knockdown of the
T-box transcription factor Brachyury increases sensitivity of
adenoid cystic carcinoma cells to chemotherapy and radiation in
vitro: Implications for a new therapeutic principle. Int J Oncol.
44:1107–1117. 2014.PubMed/NCBI
|
|
30
|
Pinto F, Pértega-Gomes N, Pereira MS,
Vizcaíno JR, Monteiro P, Henrique RM, Baltazar F, Andrade RP and
Reis RM: T-box transcription factor brachyury is associated with
prostate cancer progression and aggressiveness. Clin Cancer Res.
20:4949–4961. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hofmann HS, Hansen G, Richter G, Taege C,
Simm A, Silber RE and Burdach S: Matrix metalloproteinase-12
expression correlates with local recurrence and metastatic disease
in non-small cell lung cancer patients. Clin Cancer Res.
11:1086–1092. 2005.PubMed/NCBI
|
|
32
|
Weigelt B, Glas AM, Wessels LF, Witteveen
AT, Peterse JL and van't Veer LJ: Gene expression profiles of
primary breast tumors maintained in distant metastases. Proc Natl
Acad Sci USA. 100:15901–15905. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gomez-Roca C, Raynaud CM, Penault-Llorca
F, Mercier O, Commo F, Morat L, Sabatier L, Dartevelle P, Taranchon
E, Besse B, et al: Differential expression of biomarkers in primary
non-small cell lung cancer and metastatic sites. J Thorac Oncol.
4:1212–1220. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sarkar D, Shields B, Davies ML, Muller J
and Wakeman JA: Brachyury confers cancer stem cell characteristics
on colorectal cancer cells. Int J Cancer. 130:328–337. 2012.
View Article : Google Scholar
|
|
35
|
Palena C, Roselli M, Litzinger MT, Ferroni
P, Costarelli L, Spila A, Cavaliere F, Huang B, Fernando RI,
Hamilton DH, et al: Overexpression of the EMT driver brachyury in
breast carcinomas: Association with poor prognosis. J Natl Cancer
Inst. 106:piidju054. 2014. View Article : Google Scholar
|