Introduction
Pancreatic cancer (PC) is one of the most common
malignant tumors and remains a treatment-refractory cancer due to
its early formation of metastases, poor prognostic and high
mortality (1,2). The incidence and mortality of PC are
increasing world wide. Currently, the most common treatments for PC
patients are surgical resection followed by adjuvant chemotherapy
and radiotherapy. Unfortunately, >80% of PC patients already
have local invasion or distant metastasis when first diagnosed, and
thus have lost the opportunity of surgical therapy. The overall
5-year survival rate of these patients is <5% (3–5). Even
for those patients whose pancreatic cancer can be surgically
resected, followed by adjuvant chemotherapy and radiotherapy, the
overall 5-year survival rate is <25% (6,7). The
high metastatic potential, strong growth and chemotherapy
resistance are the main causes of high mortality of pancreatic
cancer (8,9). However, the multiple metastasis
mechanism of PC remains unclear. Therefore, understanding the key
factors involved in these processes is urgently needed to explore
new therapeutic targets of PC.
Accumulating evidence demonstrates that
epithelial-mesenchymal transition (EMT) could promote cell
proliferation, cell motility and aggression, as well as
anti-apoptotic ability and chemotherapy resistance (10–14).
The key step of EMT is that the expression of epithelial phenotype
E-cadherin is downregulated, which plays an important role in the
maintenance of cell interactions and epithelial cell polarity.
Moreover, the expression of mesenchymal phenotype such as vimentin,
N-cadherin, α-SMA, fibronectin are upregulated (15–17).
The deletion of E-cadherin in pancreatic cancer is associated with
the increase of tumor size, distant metastasis and pathological
staging (18). Upregulation of
mesenchymal phenotype could decrease the sensitivity of PC cells to
chemotherapeutic drugs, and increase the resistance to chemotherapy
(19,20).
Transforming growth factor-β1 (TGF-β1) and bone
morphogenetic proteins 2 (BMP2) are both members of transforming
growth factor-β superfamily, which are involved in cell
proliferation, differentiation, apoptosis, metastasis and
angiogenesis (21–25). Dysregulation of their pathway
contributes to a variety of pathologies, including cancer, fibrosis
and inflammation. In the early stage of tumor, TGF-β1 exhibits
tumor suppressive effects by inhibiting cell cycle progression and
promoting apoptosis. However, in the late stage TGF-β1 exerts tumor
promoting effects by increasing tumor invasiveness and metastasis.
BMP2 also has a similar dual effect. High expression of TGF-β1 and
BMP2 in tumor tissue or blood of patients may be correlated with
disease progression and poor prognosis. A huge number of studies
indicated that both TGF-β1 and BMP2 may be involved in the process
of EMT in PC cells through Smad signaling pathway or non-Smad
signaling pathway so as to increase the ability of invasion and
metastasis (26–30), and the latter may play a more
important role. However, the concrete mechanism has not been
elucidated yet.
Protein phosphatase 1H (PPM1H), a member of the PP2C
family of serine/threonine protein phosphatases, which was
originally identified as a negative regulator of neurite outgrowth,
was comprehensively expressed in healthy human tissues (GeneCards:
http://www.genecards.org/cgi-bin/carddisp.pl?gene=PPM1H&search=PPM1H).
PPM1H was reported to control cell proliferation and
differentiation (31–34). Recently, it has been indicated to
participate in the process of cancer development (31–33).
However, the role of PPM1H in cancer remains controversial.
Previously, PPM1H was found to be downregulated in glioblastoma and
renal cell carcinoma compared to normal brain and kidney but
upregulated in colon adenocarcinoma and prostate adenocarcinoma
compared to normal colon and prostate. It was also demonstrated
that low PPM1H expression in breast cancer trend toward trastuzumab
resistance and worse clinical outcome. These studies suggested that
the role of PPM1H may differ depending on the cancer type and its
exact mechanism of action in cancer remains largely unknown. Until
now, no attempt was made to examine the role of PPM1H in pancreatic
cancer.
In this study, we examined the expression of PPM1H
in human PC cell lines. In addition, we explored the role of PPM1H
in inducing EMT as well as promoting human PC cell migration.
Furthermore, we determined the effect of PPM1H depletion on the
proliferation and apoptosis of human PC cells.
Materials and methods
Cell lines and cell culture
The human pancreatic cancer cell lines PANC-1,
BxPC-3, PANC-03.27, SW1990, MIA-PACA2 and the normal human
pancreatic epithelial cell line HPDE6-C7 were used in this study.
The cell lines PANC-1, BxPC-3 and HPDE6-C7 were purchased from the
cell bank of the Chinese Academy of Sciences (Shanghai, China), and
the other cells were obtained from the American Type Culture
Collection (ATCC). The cell lines BxPC-3, PANC-03.27, MIA-PACA2 and
HPDE6-C7 were cultured in RPMI-1640 medium (Gibco, Rockville, MD,
USA) containing 10% fetal bovine serum (Gibco) and the cell lines
PANC-1 and SW1990 were cultured in DMEM medium (Gibco) containing
10% fetal bovine serum (Gibco). All of the cells were grown at 37°C
in a humidified atmosphere containing 95% O2 and 5%
CO2. BxPC-3 cells were grown to approximately 60%
confluency in RPMI-1640 +10% FBS and were then serum-deprived
overnight in RPMI-1640 medium. Cells were then treated with 10
ng/ml TGF-β1 or 200 ng/ml BMP2 (Peprotech Inc., Rocky Hill, NJ,
USA) for 72 h. The morphology of cells was visualized with a phase
contrast microscope (×200, Nikon, Japan) and imaged with digital
photography.
siRNA transfection
siRNA targeting human PPM1H
(5′-CACGCUUCUUUACCGAGA-3′ and 5′-UCUCGGUAAAGAAGCGUG-3′ duplex) was
synthesized by Ribobio Co. (Guang Zhou, China). A scrambled duplex
siRNA was used as the negative control. BxPC-3 cells were plated at
2×105/well in 6-well plates and incubated until they
reached 50% confluency. Cells were transfected with PPM1H siRNA or
the negative control siRNA at a final concentration of 50 nM with
Lipofectamine 2000 Transfection Reagent (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer's recommendations. After 6 h of
transfection, the medium was replaced with RPMI-1640 medium
containing 10% fetal bovine serum. Cells were then incubated for 72
h for RNA isolation and protein extraction.
RNA isolation and quantitative RT-PCR
(qRT-PCR)
The total RNA from BxPC-3 cells was extracted using
the TRIzol reagent (Takara, Inc., Otsu, Japan) according to the
manufacturer's instructions and was resuspended in nuclease-free
water. Then, 500 ng of total RNA was reverse-transcribed in a final
volume of 10 μl with PrimeScript™ II First-Strand cDNA
Synthesis kit (Takara, Inc.). The reverse transcription reaction
was processed at 37°C for 15 min, 85°C for 5S and 4°C for 10 min.
qRT-PCR was performed using the SYBR Green Master Mix (Takara,
Inc.) on StepOne Real-time PCR systems (Applied Biosystems, Foster
City, CA, USA) in triplicate, and non-template controls were run
for each assay under the same conditions. The PCR reaction was
carried out with an initial denaturation step of 95°C for 30 sec
followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec.
Primers used for real-time PCR were as follows: PPM1H (forward,
5′-GCATTCATGCAGCCTCACTTGC-3′, reverse,
5′-GCCATCCTCTGTCCTTCAGCAC-3′); E-cadherin (forward,
5′-ACAGCCCCGCCTTATGATTCTC-3′, reverse, 5′-AAGCGATTGCCCCATTCGTT-3′);
Vimentin (forward, 5′-CCTTGAACGCAAAGTGGAATC-3′, reverse,
5′-GACATGCTGTTCCTGAATCTGAG-3′); β-actin (forward,
5′-GTTGCGTTACACCCTTTCTTG-3′, reverse, 5′-GACTGCTGTCACCTTCACCGT-3′).
The analysis of qPCR was carried out using the 2−ΔΔCt
method. β-actin was taken as the internal control.
Western blotting
Proteins were extracted from cultured cells and then
quantitated using Pierce BCA Protein Assay kit (Boster Biological
Engineering Co., Ltd., Wuhan, China). Equal amounts of protein from
different samples were separated by 10% SDS/polyacrylamide gel in
Tris-glycin buffer and transferred to PVDF membranes. The membranes
were blocked for 1 h at room temperature in blocking buffer (5%
skim milk in 0.5 Tween-20-TBST), then probed with PPM1H antibody
(Abgent, diluted 1:1000 in TBST), E-cadherin antibody (ProteinTech,
diluted 1:800 in TBST), vimentin antibody (ProteinTech, diluted
1:5000 in TBST), akt antibody (CST, diluted 1:1000 in TBST) and
p-akt antibody (CST, diluted 1:1000 in TBST), respectively,
overnight at 4°C with GAPDH antibody (Promoter Biological Co.,
China, diluted 1:5000 in TBST) as control. Following washes with
TBST, the membranes were incubated with HRP-conjugated goat
anti-rabbit secondary antibody (Promoter Biological Co., diluted
1:5000 in TBST) for 1 h at room temperature. After extensive
washing with TBST, the complex was detected by Super Signal West
Pico Chemiluminescent Substrate (Thermo Fisher Scientific Inc.,
Waltham, MA, USA) according to the manufacturer's instructions.
Blot was scanned and densitometric analysis was done by ImageJ
software (National Institutes of Health, Bethesda, MD, USA).
Transwell cell invasion and migration
assay
The cell invasion and migration was assessed using
24-well polycarbonate filters (membrane pore size 8 μm).
Cells (8×104) were seeded in 100 μl serum-free
medium into the upper chamber and allowed to invade into the lower
chamber, which was filled with 600 μl of RPMI-1640 medium
containing 10% FBS as a chemoattractant. The chamber for invasion
assay was coated by Matrigel (BD Biosciences, Foster City, CA,
USA). The Transwell plate was incubated at 37°C in a 5%
CO2 incubator for 24 h, then the upper chamber was
successively fixed with methanol and stained by 0.1% crystal violet
for 25 min. The upper chamber was then washed with PBS to remove
excess stain and the unmigrated cells staying at the upper layer of
the microporous membrane were gently scraped with a wet cotton
swab. After drying, the stained migrated cells were counted with
the phase contrast microscope. The average number of migrated cells
per field was quantified under high power (×200).
Cell proliferation assay
Cell proliferation was analyzed using the Cell Count
Kit-8 (CCK-8) assay. BxPC-3 cells were seeded in 96-well plates at
a density of 10000 cells per well and transfected with PPM1H siRNA
or the negative control siRNA when the cells reached 50%
confluency, then incubated for 1, 2 or 3 days. At the indicated
time point, CCK8 solution was added to each well and incubated for
1, 2, 3, 4 h. The absorbance value (optical density) of each well
was measured at 450 nm. For each experimental condition, 6-wells
were used. All experiments were performed thrice.
Flow cytometry
BxPC-3 cells were transfected with PPM1H siRNA or
the negative control siRNA for 72 h, then were harvested and washed
with ice-cold PBS. The cells were trypsinized without EDTA and
resuspended with binding buffer. Annexin V and PI staining were
carried out using the Annexin V-FITC Apoptosis Detection kit (BD
Biosciences, San Jose, CA, USA), according to the manufacturer's
instructions. After a 15-min incubation in a dark at room
temperature, the cells were immediately analyzed by FACScan flow
cytometer.
Statistical analysis
Data were presented as mean ± standard deviation
(SD). Experiments were repeated at least three times. SPSS 17.0
software (IBM, Chicago, IL, USA) was used for data analysis. Group
differences were analyzed by Student's t-test or analysis of
variance (ANOVA) according to the data type. P<0.05 was
considered statistically significant.
Results
Expression of PPM1H in pancreatic cancer
cell lines
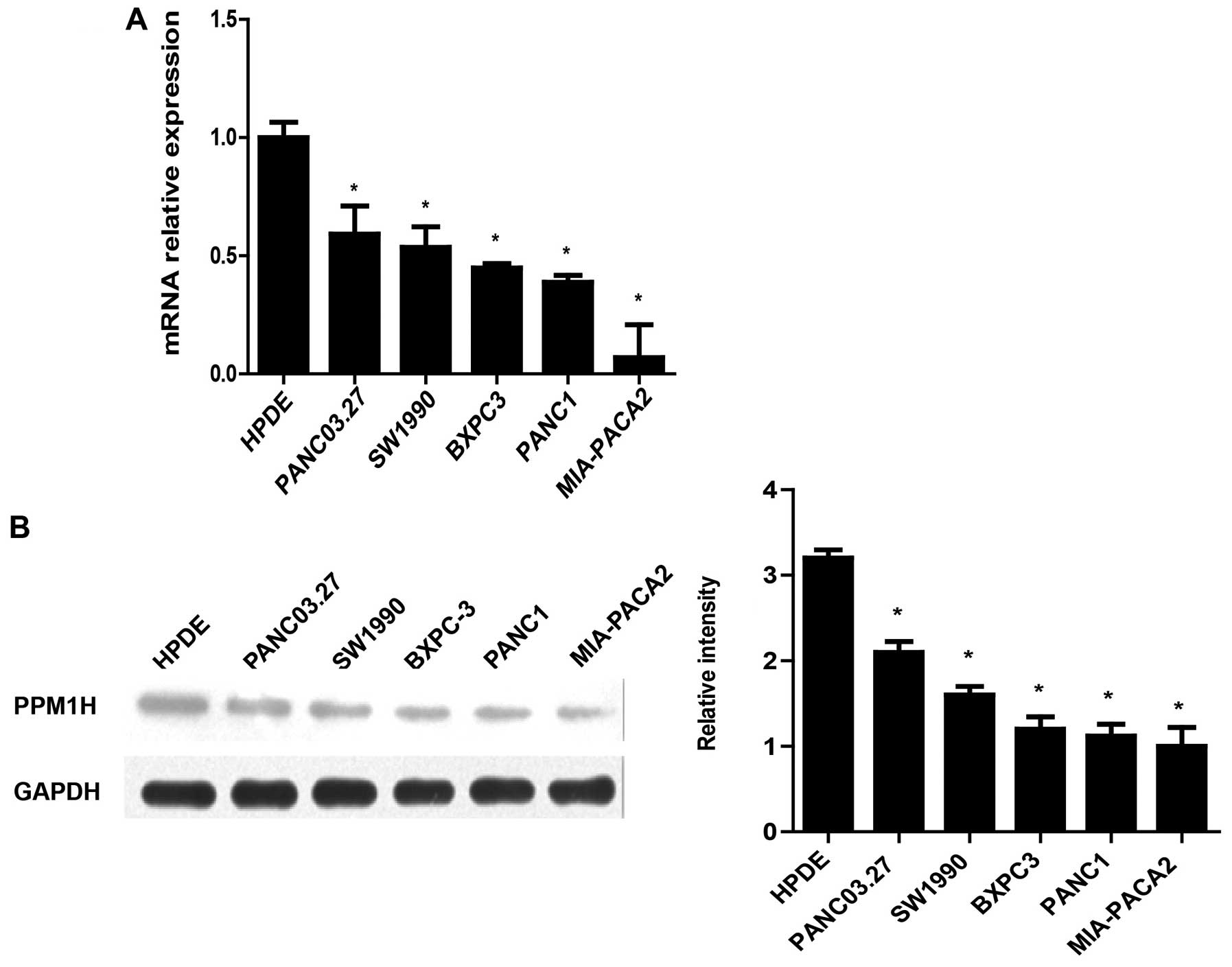
The expression of PPM1H in human pancreatic cancer
cell lines (BxPC-3, MIA-PACA2, SW1990, PANC-1, PANC-03.27) were
analyzed by real-time PCR and western blotting. The real-time PCR
results showed that PPM1H mRNA expression in PC cells were
significantly lower than in normal pancreatic ductal epithelial
cells (HPDE6-C7) (Fig. 1A). In
addition, western blot data also showed lower expression of PPM1H
in PC cell lines compared with HPDE6-C7 cells (Fig. 1B). These data indicated that the
expression of PPM1H was downregulated in PC cells.
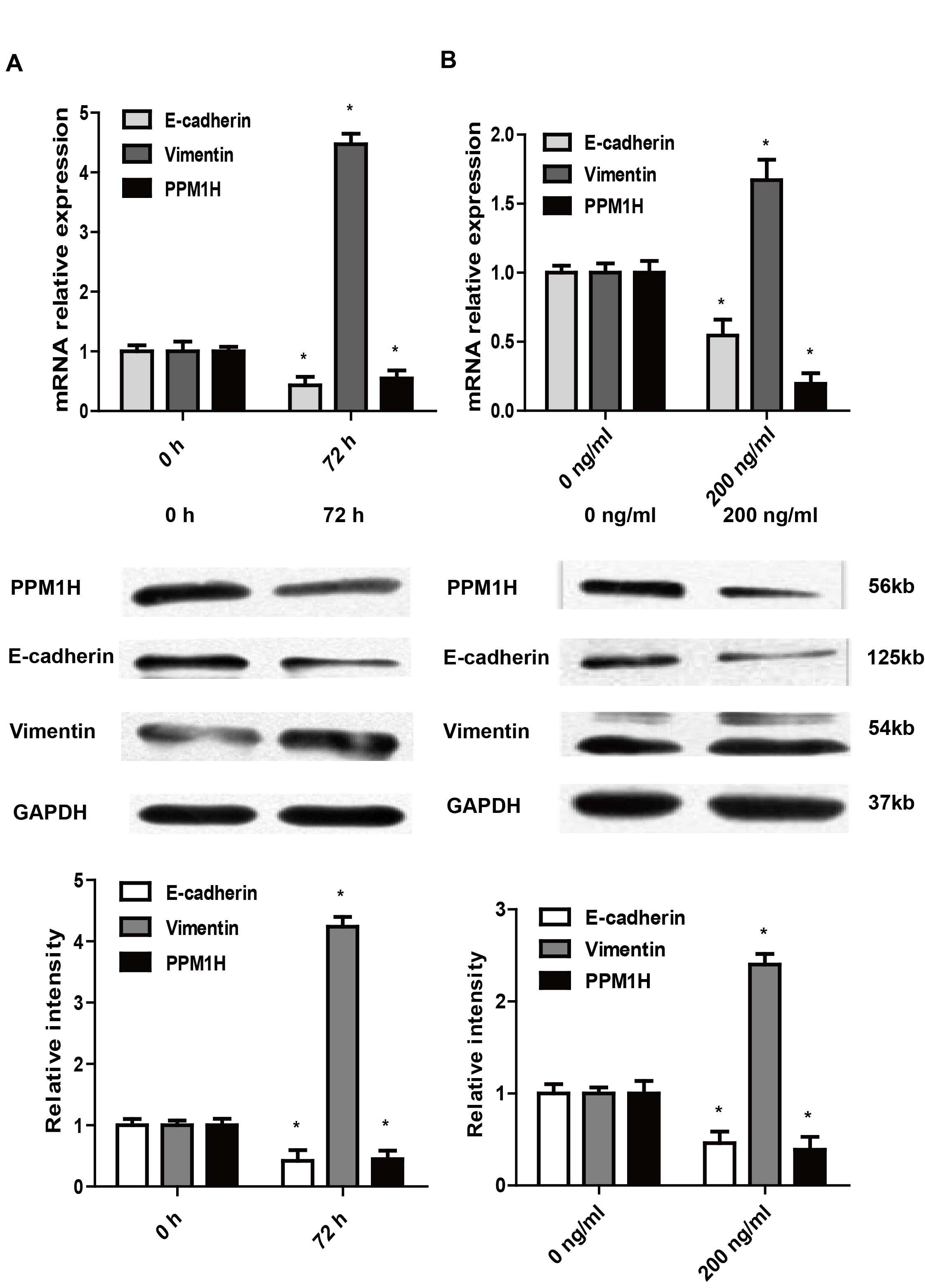
Effect of TGF-β1 and BMP2 on EMT and
PPM1H expression in PC cells
TGF-β1 and BMP2 were demonstrated to be involved in
the process of EMT in tumor cells so as to increase their ability
of invasion and metastasis. As showed by western blotting, after
treatment with 10 ng/ml TGF-β1 or 200 ng/ml BMP2 for 72 h, the
expression of E-cadherin in BxPC-3 cells was decreased, and the
expression of vimentin was increased, indicating EMT of BxPC-3
cells were induced. In addition, the expression of PPM1H was
significantly reduced in TGF-β1 or BMP2 treated BxPC-3 cells
(P<0.05, Fig. 2). These data
suggested that PPM1H may participate in the regulation of EMT.
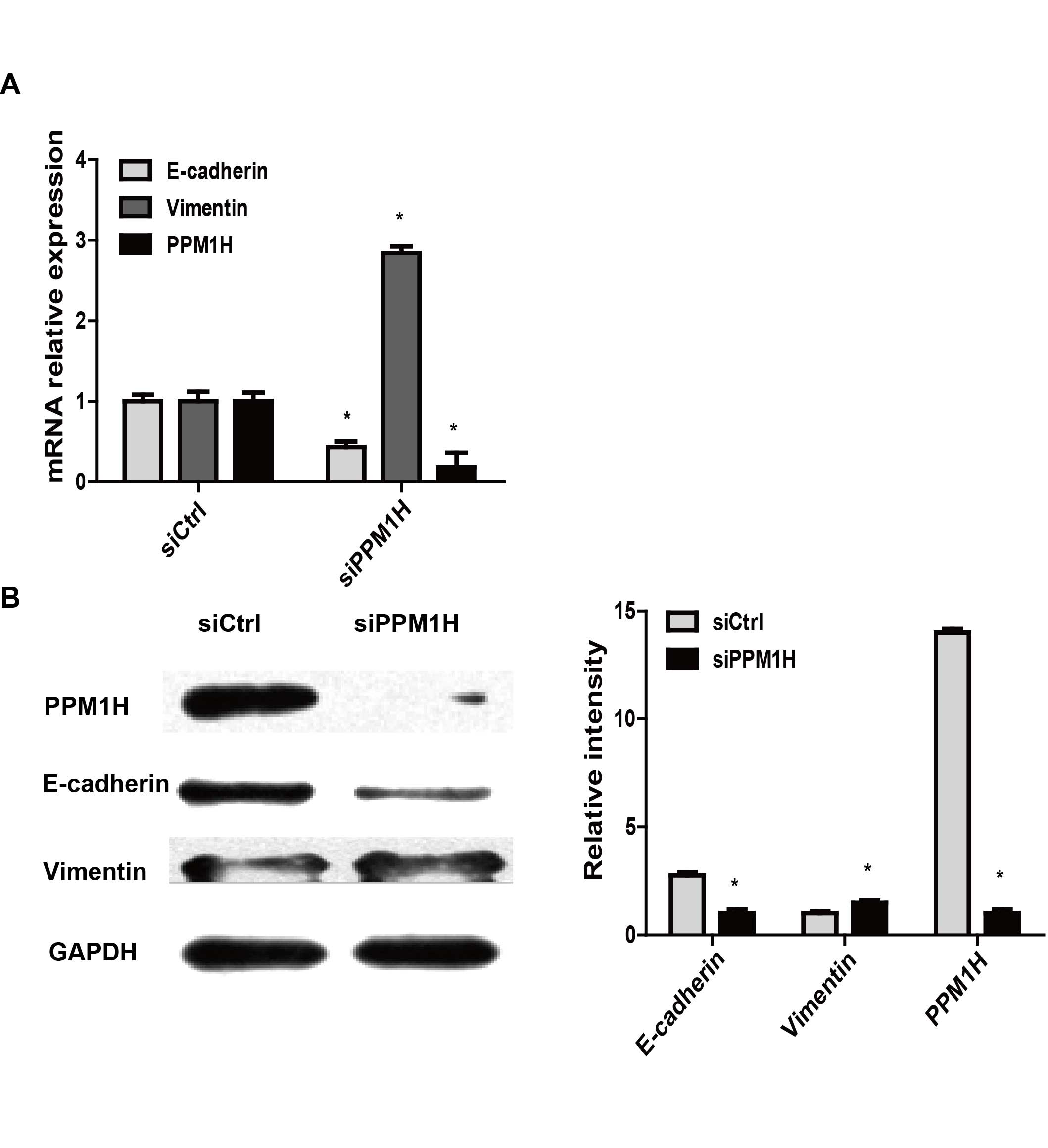
PPM1H gene knockdown induces EMT in PC
cells
To investigate whether PPM1H could induce EMT in PC
cells, PPM1H specific siRNA were used to silence PPM1H expression.
EMT-related markers E-cadherin and vimentin were detected by
real-time PCR and western blotting, respectively, in PPM1H siRNA
(si-PPM1H) transfected BxPC-3 cells. The data showed in BxPC-3
cells that downregulated E-cadherin and upregulated vimentin
expression were induced by PPM1H gene silencing (P<0.05,
Fig. 3), indicating PPM1H gene
silencing may cause EMT in PC cells.
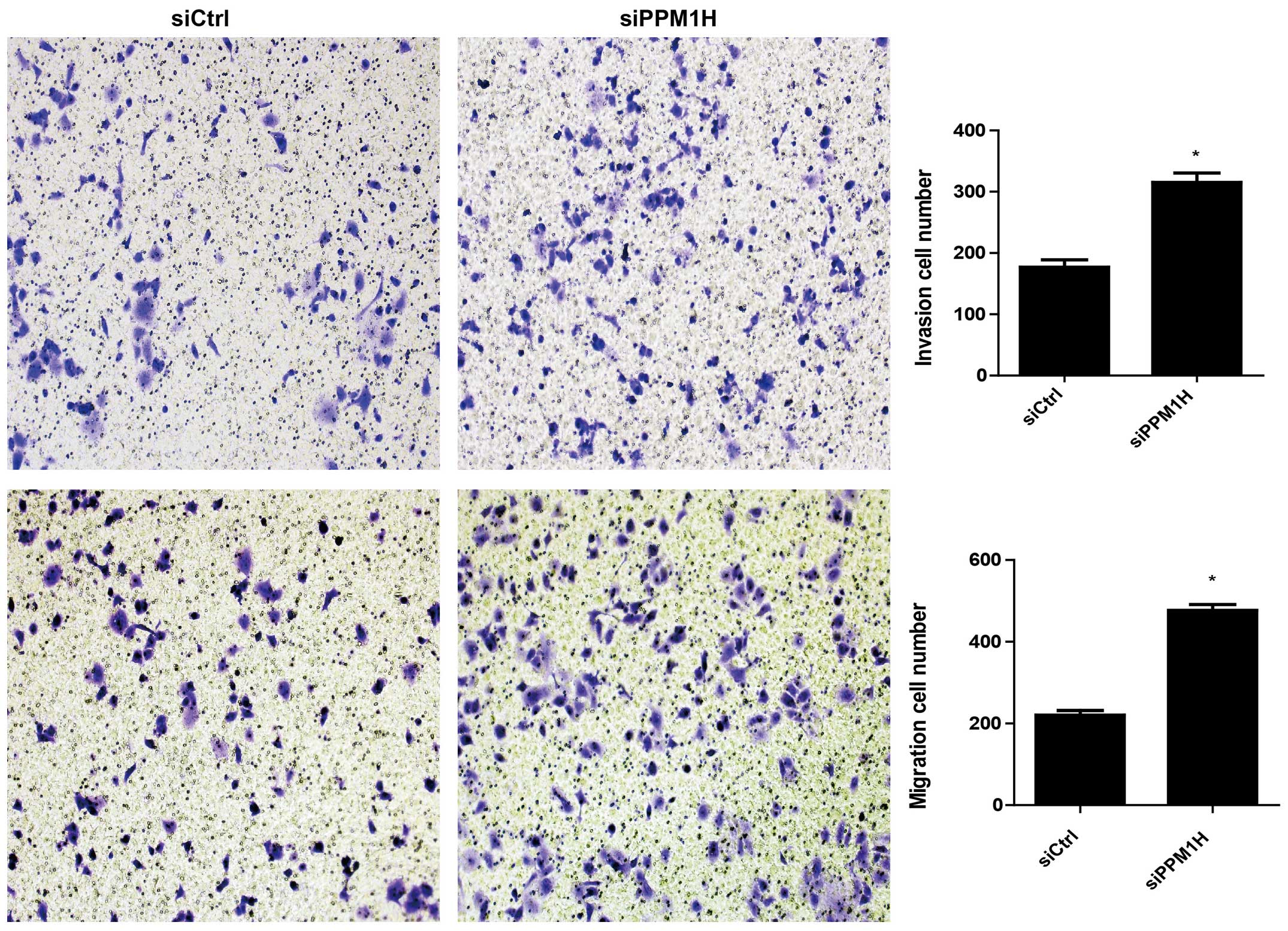
PPM1H knockdown promotes invasion and
migration of PC cells
Next, we used Transwell cell migration assay to
examine the role of PPM1H in invasion and migration of PC cells.
The cell migration rates were compared in PPM1H siRNA treated group
and control group. The data showed that the cells penetrated the
artificial basement membrane in siPPM1H group were two to three
times more than in siCtrl group (P<0.05, Fig. 4), implicating that PPM1H depletion
resulted in increased invasion and migration of PC cells.
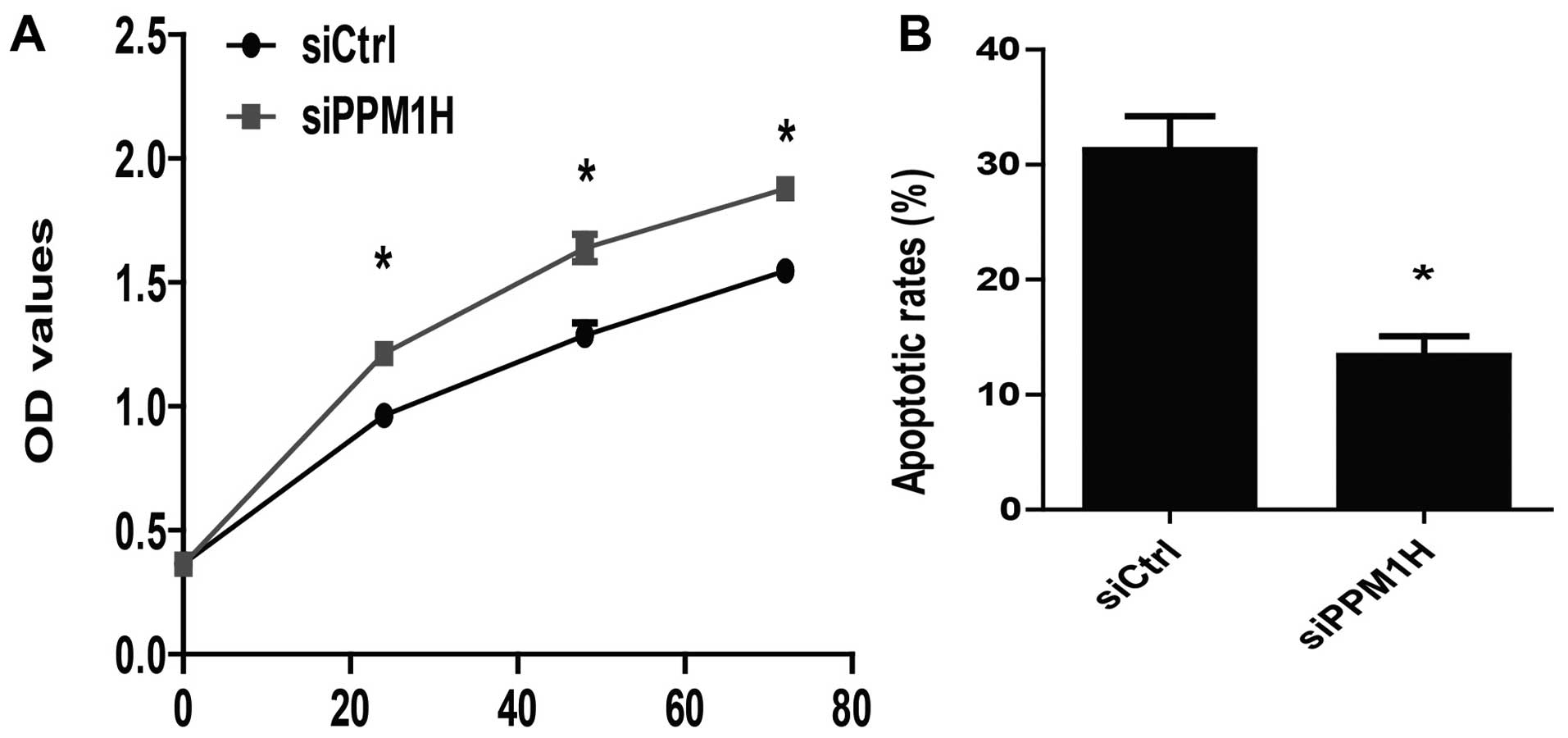
PPM1H knockdown promotes proliferation
and inhibits apoptosis of PC cells
To further determine the role of PPM1H in PC cells,
we assessed the effect of PPM1H depletion on cell proliferation and
apoptosis. It was shown that silencing PPM1H resulted in increased
proliferation rate in BxPC-3 cells (P<0.05, Fig. 5A). We also found that the apoptosis
rate was reduced nearly 20% in siPPM1H group compared to siCtrl
group (P<0.05, Fig. 5B). These
results demonstrated that PPM1H silencing could promote
proliferation and inhibit apoptosis of PC cells.
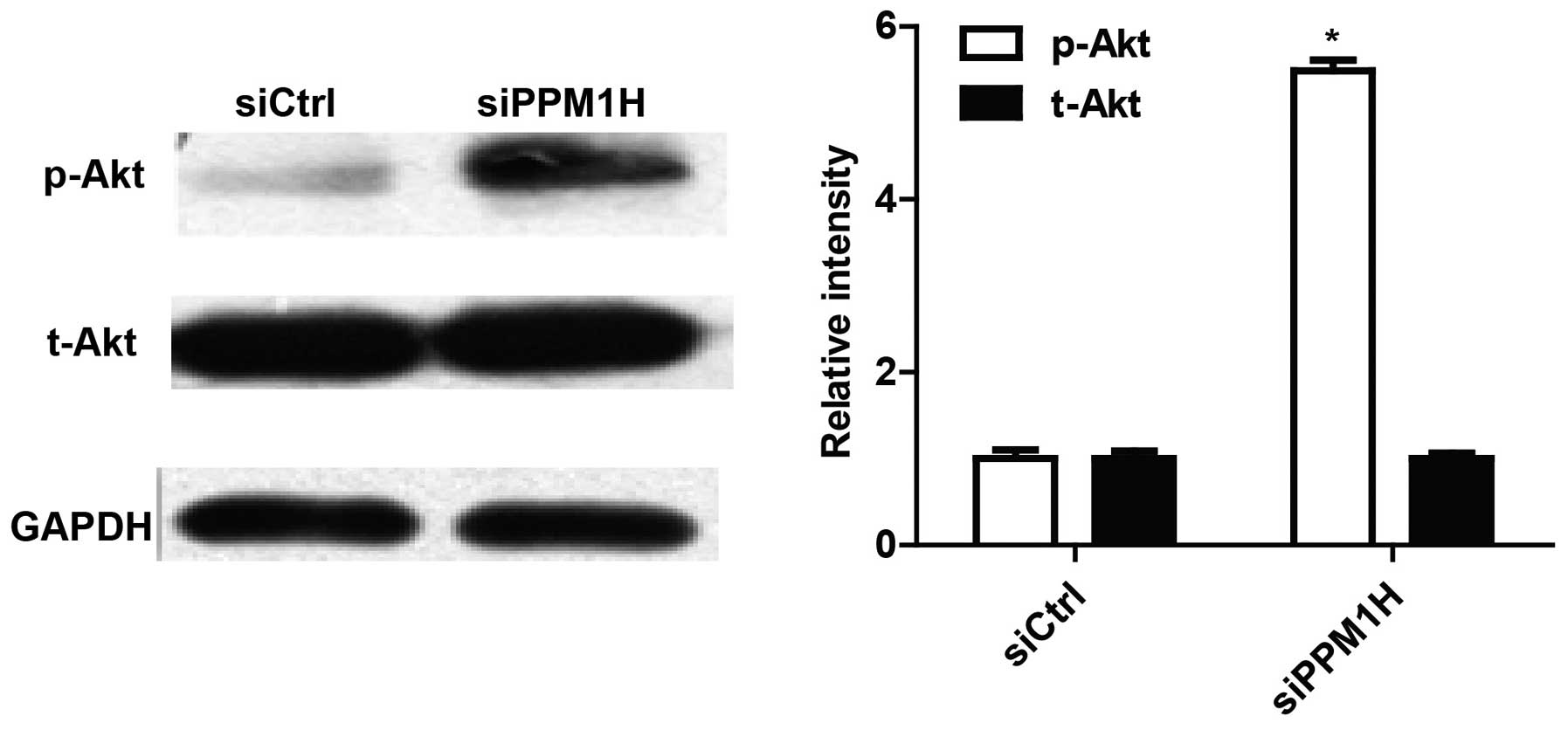
PPM1H knockdown promotes Akt
phosphorylation of PC cells
To explore the mechanisms involved in PPM1H function
in PC cells, we tested Akt activity of BxPC-3 by western blotting.
As shown in Fig. 6, silencing PPM1H
can increase Akt phosphorylation. This result suggested that PPM1H
may function through the Akt signaling pathway.
Discussion
The morbidity and mortality of pancreatic cancer
have been gradually rising, while those of other common cancers
have been declining (7). Although
the detection and management of pancreatic cancer have achieved
certain developments, only less than 5% of patients live 5 years
after diagnosis (6,7). Since its malignant degree is high and
the rate of resection is low, and pancreatic cancer responds poorly
to most chemo therapeutic agents, it is urgent for us to understand
the biological mechanisms that contribute to development and
progression of pancreatic cancer.
EMT was initially considered to be an important
feature of embryonic differentiation and morphology. And now, it
has been demonstrated to be involved in the development of many
diseases, such as inflammation, fibrosis and tumor (35). Accumulating evidence has indicated
that EMT can promote an early stage tumor into aggressive malignant
tumor by increasing the motility and invasiveness of the cells,
this process was consistent with the acquisition of tumor stem cell
phenotype (36–39). Some studies have shown that EMT can
increase the invasive and anti-apoptosis capacity of cells, and may
also affect its ability to resist chemotherapy. In recent years,
EMT has been considered to play a critical role in the high
drug-resistance and invasion of pancreatic cancer, therefore, the
study of EMT in pancreatic cancer is helpful for the development of
new drugs and new targets for molecular therapy.
Phosphorylation and dephosphorylation of proteins
have important effect on the regulation of life activities of
eukaryotic organisms, and the process of phosphorylation is
co-regulated by protein kinase and protein phosphatase. Protein
phosphatase is a kind of key enzyme that controls dephosphorylation
of proteins. Its absence or dysfunction can lead to a variety of
diseases. PPM1H (protein phosphatase,
Mg2+/Mn2+ dependent 1H) is a member of the
PP2C family of Ser/Thr protein phosphatases, which is not sensitive
to Okada acid. Most members of the PP2C family have been shown to
act as inhibitors of cell growth and cellular stress signaling.
These members include PP2Cα, PP2Cβ, and the recently identified
PP2Cs, ILKAP, and PHLPP; ILKAP negatively affects proliferation and
malignant transformation, and PHLPP promotes apoptosis and inhibits
tumor growth. Based on these observations, it is reasonable to
assume that, in general, type 2C phosphatases act as tumor
suppressor proteins. In addition to the PP2Cδ (PPM1D/Wip1), which
behaves as an oncogene (40–43).
At present, there are few studies on PPM1H in tumor,
and the effect of PPM1H is controversial. As the result of this
study shows that PPM1H was downregulated at both transcriptional
and translational levels in five human pancreatic cancer cell lines
compared with normal pancreatic ductal epithelial cell line
(P<0.05), these results were different from the research of
Sugiura et al in colon cancer cell lines (31). These results suggested that PPM1H
plays a complex role in cancer and the effect of PPM1H may vary
among cancers of different organs or tissues. Based on this result,
we speculated that PPM1H may act as a tumor suppressor in
pancreatic cancer.
Both TGF-β1 and BMP2 can regulate EMT through Smad
signaling pathway and non-Smad signaling pathway, the latter is
considered to play a more important role in the process of inducing
EMT (26–27,44–46).
Our study demonstrated that both TGF-β1 and BMP2 can induce EMT in
BxPC-3 cells, interestingly the expression of PPM1H was
significantly downregulated after induction of EMT. It suggests
that PPM1H may participate in the regulation of EMT. To further
confirm the role of PPM1H in EMT of pancreatic cancer cells, we
chose BxPC-3 cell line for PPM1H silencing in which endogenous
PPM1H expression was at relatively high level. We used interference
RNA (RNAi) technique to induce PPM1H gene silencing, and qRT-PCR
and western blotting were used to detect the expression of PPM1H
and EMT related molecules. After silencing PPM1H, the expression of
epithelial mesenchymal phenotype E-cadherin was downregulated,
while the expression of mesenchymal phenotype vimentin was
upregulated in BxPC-3 cells (P<0.05). It was indicated that
deletion of PPM1H gene can promote EMT independently in BxPC-3
cells. Then, we used Transwell cell invasion and migration assay to
examine whether the invasion and metastasis capacity would be
promoted when BxPC-3 cells were induced in EMT. The results showed
that the invasion and metastasis of BxPC-3 cells were enhanced
after silencing PPM1H (P<0.05), which illustrated that silencing
PPM1H could enhance the invasion and metastasis of pancreatic
cancer cells by promoting EMT.
The studies of Lee-Hoeflich et al have found
that the loss of PPM1H gene can lead P27 suppressor gene to degrade
so as to promote the proliferation of breast cancer cells (32). Hence, we speculated that PPM1H might
suppress the proliferation of pancreatic cancer cells. We performed
CCK8 assays to investigate the effect of PPM1H silencing on the
proliferation of BxPC-3 cells. Our results showed that knockdown of
PPM1H resulted in a more dramatic increase of the proliferation
rate in BxPC-3 cells than that in control cells (P<0.05). PPM1H
could suppress cell proliferation of BxPC-3 cells in vitro.
In addition, we assessed that silencing PPM1H can inhibit the
apoptosis of BxPC-3 cells by flow cytometry (P<0.05). Although
we found that silencing PPM1H gene can promote proliferation and
inhibits apoptosis of BxPC-3 cells, its specific mechanism still
needs further exploration.
Since Smad4 is homozygously deleted in BxPC-3 cells
(47), we suspected that PPM1H
affect is through non-Smad pathway. In particular, it has been
reported that mutation or deletion of Smad4 is found in
approximately 50% of pancreatic tumors and is correlated with poor
prognosis (48). Thus, it may be
more important to investigate non-Smad pathways in detail in order
to further understand invasion and metastasis of pancreatic cancer.
In this study, we found that silencing PPM1H can increase Akt
phosphorylation. The Akt signaling pathway is involved in many
cellular processes during the occurrence and development of cancer,
such as proliferation, apoptosis, invasion and metastasis (49,50).
Dephosphorylation of Akt can make it inactive, and thus inhibit the
various biological activities of Akt. This result suggests that the
PPM1H biological effects may be through the Akt signaling
pathway.
Although we have demonstrated that silencing PPM1H
can increase the invasion and metastasis of BxPC-3 cells, as well
as promote its proliferation and inhibit apoptosis, we also need to
explore the specific mechanisms of these effects. Whether silencing
PPM1H can also promote BxPC-3 cells to acquire CSC characteristics
so that it can induce chemotherapeutic resistance of BxPC-3 cells,
and whether overexpression of PPM1H could reverse the above process
is not known.
PPM1H presented low expression in pancreatic cancer
cell lines, and silencing PPM1H can enhance the invasion and
metastasis of BxPC-3 cells by mediating EMT. Silencing PPM1H also
promotes proliferation and inhibits apoptosis of BxPC-3 cells.
These effects may be produced by increasing AKT phosphorylation.
This study suggests that PPM1H may be a novel tumor suppressor gene
for pancreatic cancer. A better understanding of the function of
PPM1H in pancreatic cancer is expected to provide new insight into
the molecular targets of gene therapy for pancreatic cancer.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stathis A and Moore MJ: Advanced
pancreatic carcinoma: Current treatment and future challenges. Nat
Rev Clin Oncol. 7:163–172. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu C, Li H, Su C and Li Z: Viral therapy
for pancreatic cancer: Tackle the bad guys with poison. Cancer
Lett. 333:1–8. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
American Cancer Society: Cancer Facts
& Figures 2009. American Cancer Society; Atlanta, GA: pp.
18–19. 2009
|
|
7
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sarkar FH, Li Y, Wang Z and Kong D:
Pancreatic cancer stem cells and EMT in drug resistance and
metastasis. Minerva Chir. 64:489–500. 2009.PubMed/NCBI
|
|
9
|
Li Y, Kong D, Ahmad A, Bao B and Sarkar
FH: Pancreatic cancer stem cells: Emerging target for designing
novel therapy. Cancer Lett. 338:94–100. 2013. View Article : Google Scholar
|
|
10
|
Tsuji T, Ibaragi S and Hu GF:
Epithelial-mesenchymal transition and cell cooperativity in
metastasis. Cancer Res. 69:7135–7139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arumugam T, Ramachandran V, Fournier KF,
Wang H, Marquis L, Abbruzzese JL, Gallick GE, Logsdon CD, McConkey
DJ and Choi W: Epithelial to mesenchymal transition contributes to
drug resistance in pancreatic cancer. Cancer Res. 69:5820–5828.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rasheed ZA, Yang J, Wang Q, Kowalski J,
Freed I, Murter C, Hong SM, Koorstra JB, Rajeshkumar NV, He X, et
al: Prognostic significance of tumorigenic cells with mesenchymal
features in pancreatic adenocarcinoma. J Natl Cancer Inst.
102:340–351. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ali S, Ahmad A, Banerjee S, Padhye S,
Dominiak K, Schaffert JM, Wang Z, Philip PA and Sarkar FH:
Gemcitabine sensitivity can be induced in pancreatic cancer cells
through modulation of miR-200 and miR-21 expression by curcumin or
its analogue CDF. Cancer Res. 70:3606–3617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Iwatsuki M, Mimori K, Yokobori T, Ishi H,
Beppu T, Nakamori S, Baba H and Mori M: Epithelial-mesenchymal
transition in cancer development and its clinical significance.
Cancer Sci. 101:293–299. 2010. View Article : Google Scholar
|
|
17
|
Castellanos JA, Merchant NB and
Nagathihalli NS: Emerging targets in pancreatic cancer:
Epithelial-mesenchymal transition and cancer stem cells. Onco
Targets Ther. 6:1261–1267. 2013.PubMed/NCBI
|
|
18
|
Shin SJ, Kim KO, Kim MK, Lee KH, Hyun MS,
Kim KJ, Choi JH and Song HS: Expression of E-cadherin and uPA and
their association with the prognosis of pancreatic cancer. Jpn J
Clin Oncol. 35:342–348. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Y, VandenBoom TG II, Kong D, Wang Z,
Ali S, Philip PA and Sarkar FH: Up-regulation of miR-200 and let-7
by natural agents leads to the reversal of
epithelial-to-mesenchymal transition in gemcitabine-resistant
pancreatic cancer cells. Cancer Res. 69:6704–6712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nagaraj NS, Washington MK and Merchant NB:
Combined blockade of Src kinase and epidermal growth factor
receptor with gemcitabine overcomes STAT3-mediated resistance of
inhibition of pancreatic tumor growth. Clin Cancer Res. 17:483–493.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Raida M, Clement JH, Leek RD, Ameri K,
Bicknell R, Niederwieser D and Harris AL: Bone morphogenetic
protein 2 (BMP-2) and induction of tumor angiogenesis. J Cancer Res
Clin Oncol. 131:741–750. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wakefield LM and Hill CS: Beyond TGFβ:
Roles of other TGFβ superfamily members in cancer. Nat Rev Cancer.
13:328–341. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Carreira AC, Alves GG, Zambuzzi WF,
Sogayar MC and Granjeiro JM: Bone morphogenetic proteins:
Structure, biological function and therapeutic applications. Arch
Biochem Biophys. 561:64–73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Senturk S, Mumcuoglu M, Gursoy-Yuzugullu
O, Cingoz B, Akcali KC and Ozturk M: Transforming growth
factor-beta induces senescence in hepatocellular carcinoma cells
and inhibits tumor growth. Hepatology. 52:966–974. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ikushima H and Miyazono K: TGFbeta
signalling: A complex web in cancer progression. Nat Rev Cancer.
10:415–424. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kaminska B, Wesolowska A and Danilkiewicz
M: TGF beta signalling and its role in tumour pathogenesis. Acta
Biochim Pol. 52:329–337. 2005.PubMed/NCBI
|
|
27
|
Wharton K and Derynck R: TGFbeta family
signaling: Novel insights in development and disease. Development.
136:3691–3697. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen X, Liao J, Lu Y, Duan X and Sun W:
Activation of the PI3K/Akt pathway mediates bone morphogenetic
protein 2-induced invasion of pancreatic cancer cells Panc-1.
Pathol Oncol Res. 17:257–261. 2011. View Article : Google Scholar
|
|
29
|
Kang MH, Oh SC, Lee HJ, Kang HN, Kim JL,
Kim JS and Yoo YA: Metastatic function of BMP-2 in gastric cancer
cells: The role of PI3K/AKT, MAPK, the NF-κB pathway, and MMP-9
expression. Exp Cell Res. 317:1746–1762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cano CE, Motoo Y and Iovanna JL:
Epithelial-to-mesenchymal transition in pancreatic adenocarcinoma.
Sci World J. 10:1947–1957. 2010. View Article : Google Scholar
|
|
31
|
Sugiura T, Noguchi Y, Sakurai K and
Hattori C: Protein phosphatase 1H, overexpressed in colon
adenocarcinoma, is associated with CSE1L. Cancer Biol Ther.
7:285–292. 2008. View Article : Google Scholar
|
|
32
|
Lee-Hoeflich ST, Pham TQ, Dowbenko D,
Munroe X, Lee J, Li L, Zhou W, Haverty PM, Pujara K, Stinson J, et
al: PPM1H is a p27 phosphatase implicated in trastuzumab
resistance. Cancer Discov. 1:326–337. 2011. View Article : Google Scholar
|
|
33
|
Aceto N and Bentires-Alj M: On the road to
combinations of targeted therapies: PPM1H phosphatase as a
suppressor of trastuzumab resistance. Cancer Discov. 1:285–286.
2011. View Article : Google Scholar
|
|
34
|
Shen T, Sun C, Zhang Z, Xu N, Duan X, Feng
XH and Lin X: Specific control of BMP signaling and mesenchymal
differentiation by cytoplasmic phosphatase PPM1H. Cell Res.
24:727–741. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mani SA, Guo W, Liao M-J, Eaton EN,
Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et
al: The epithelial-mesenchymal transition generates cells with
properties of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Floor S, van Staveren WC, Larsimont D,
Dumont JE and Maenhaut C: Cancer cells in epithelial-to-mesenchymal
transition and tumor-propagating-cancer stem cells: Distinct,
overlapping or same populations. Oncogene. 30:4609–4621. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Santisteban M, Reiman JM, Asiedu MK,
Behrens MD, Nassar A, Kalli KR, Haluska P, Ingle JN, Hartmann LC,
Manjili MH, et al: Immune-induced epithelial to mesenchymal
transition in vivo generates breast cancer stem cells. Cancer Res.
69:2887–2895. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shreeram S and Bulavin DV: PPM1H - new kid
on the block. Cancer Biol Ther. 7:293–294. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Leung-Hagesteijn C, Mahendra A,
Naruszewicz I and Hannigan GE: Modulation of integrin signal
transduction by ILKAP, a protein phosphatase 2C associating with
the integrin-linked kinase, ILK1. EMBO J. 20:2160–2170. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gao T, Furnari F and Newton AC: PHLPP: A
phosphatase that directly dephosphorylates Akt, promotes apoptosis,
and suppresses tumor growth. Mol Cell. 18:13–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shreeram S, Demidov ON, Hee WK, Yamaguchi
H, Onishi N, Kek C, Timofeev ON, Dudgeon C, Fornace AJ, Anderson
CW, et al: Wip1 phosphatase modulates ATM-dependent signaling
pathways. Mol Cell. 23:757–764. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Moustakas A and Heldin CH: Non-Smad
TGF-beta signals. J Cell Sci. 118:3573–3584. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gordon KJ, Kirkbride KC, How T and Blobe
GC: Bone morphogenetic proteins induce pancreatic cancer cell
invasiveness through a Smad1-dependent mechanism that involves
matrix metalloproteinase-2. Carcinogenesis. 30:238–248. 2009.
View Article : Google Scholar :
|
|
47
|
Deer EL, González-Hernández J, Coursen JD,
Shea JE, Ngatia J, Scaife CL, Firpo MA and Mulvihill SJ: Phenotype
and genotype of pancreatic cancer cell lines. Pancreas. 39:425–435.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wilentz RE, Iacobuzio-Donahue CA, Argani
P, McCarthy DM, Parsons JL, Yeo CJ, Kern SE and Hruban RH: Loss of
expression of Dpc4 in pancreatic intraepithelial neoplasia:
Evidence that DPC4 inactivation occurs late in neoplastic
progression. Cancer Res. 60:2002–2006. 2000.PubMed/NCBI
|
|
49
|
Chang F, Lee JT, Navolanic PM, Steelman
LS, Shelton JG, Blalock WL, Franklin RA and McCubrey JA:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: A target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Altomare DA and Testa JR: Perturbations of
the AKT signaling pathway in human cancer. Oncogene. 24:7455–7464.
2005. View Article : Google Scholar : PubMed/NCBI
|