Introduction
Lung cancer remains the leading cause of
cancer-related deaths worldwide (1). Non-small cell lung cancer (NSCLC)
accounts for ~85% of all lung cancer subtypes and its clinical
course is very crucial with poor patient outcomes (2,3).
Despite recent advances in chemotherapy and surgical procedures,
the overall 5-year survival rate of NSCLC patients (~15%) has not
markedly improved (1). Adjacent
invasion and distant metastases are responsible for the failure of
lung cancer therapy (4). Therefore,
further exploration of the underlying mechanisms of NSCLC
progression is essential.
Epithelial-mesenchymal transition (EMT) is a key
stage in tumor progression. This biological process permits
polarized epithelial cells to assume a mesenchymal cell phenotype,
however this often induces tumor metastasis (5). Increasing evidence supports EMT as a
common event in human advanced NSCLC (6,7).
Consequently, to identify new diagnostic strategies and
specifically targeted drugs, exploring key molecules in EMT that
control NSCLC metastasis is paramount.
MicroRNAs (miRNAs), a family of small non-coding
RNAs, are processed from precursor RNAs with a typical hairpin
secondary structure (8). Several
miRNAs are aberrantly expressed in NSCLC and their dysregulation
has resulted in cancer progression and poor clinical outcome
(9–11). Considering that miRNAs usually act
as EMT-associated downstream effectors of receptor signaling or
protein kinases (12), miRNAs may
represent novel targets for designing antimetastatic drugs due to
their specificity. Moreover, miRNA efficacy data is now available,
making miRNA-based technology applicable against NSCLC metastasis
in vivo (13).
Recently, miR-187 ectopic expression was detected in
various types of cancer including nasopharyngeal (14), renal (15), pancreatic (16), prostate (17) and neuroblastoma (18). In NSCLC, miR-187 is also a potential
tumor suppressor (19), however the
precise molecular mechanism through which miR-187 influences NSCLC
progression remains largely unknown. In the present study, we aimed
to investigate the potential role of miR-187 in NSCLC progression.
We also explored the mechanisms underlying tumor metastasis and
direct targets of miR-187.
Materials and methods
Cell culture
Human NSCLC cell lines A549, H322, GLC-82, SPC-A1,
PC-9, H460 and H1299, and the normal human bronchial epithelial
cell line BEAS-2B, were obtained from the Cell Bank of the Chinese
Academy of Sciences (Shanghai, China). All cells were authenticated
by short tandem repeat (STR) profiling before receipt and were
propagated for <6 months after resuscitation. All cells were
cultured in RPMI-1640 medium (HyClone, Logan, UT, USA) supplemented
with 10% fetal bovine serum (FBS; Gibco-BRL, Invitrogen, Paisley,
UK) at room temperature with a humidity of 90–95%, and 5%
CO2.
miRNA transfection
Refer to our previous study for details (20).
Microarray data set analysis
A microarray data set containing 116 paired primary
lung cancers, and their corresponding adjacent normal lung tissues
(collected a minimum of 5 cm from the tumor) were retrieved from
the Gene Expression Omnibus (GEO) database (accession no. GSE15008)
(21).
RNA isolation, reverse transcription and
quantitative real-time polymerase chain reaction (qRT-PCR). qRT-PCR
for miRNAs and target genes was performed as previously described
(20,22). Primer sequences used for PCR assays
were as follows: U6 forward, 5′-CTCGCTTCGGCAGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′; and miR-187 forward,
5′-TCGTGTCTTGTGTTGCAGCC-3′ and reverse, 5′-TGGTGTCGTGGAGTCG-3′.
Western blot analysis
Protein expression was evaluated by immunoblot
analysis of cell lysates (20–60 µg) in RIPA buffer solution using a
rabbit antibody to E-cadherin, and mouse antibodies to β-catenin,
fibronectin, vimentin and β-actin (1:500; Santa Cruz Biotechnology,
Santa Cruz, CA, USA); rabbit antibodies to p-Akt (Ser473), AKT,
p44/42 MAPK (ERK1/2), and p-p44/42 MAPK (ERK1/2) [1:1,000; Cell
Signaling Technology (CST), Danvers, MA, USA]; and rabbit
antibodies to polymerase I and transcript release factor (PTRF)
(1:500; Abcam, Cambridge, UK).
Immunofluorescence (IF)
Refer to our previous study for details (20). In brief, slides were incubated with
a rabbit antibody to E-cadherin, mouse antibodies to β-catenin and
vimentin (1:500; Santa Cruz Biotechnology), and rabbit antibody to
PTRF (1:100; Abcam) at 4̊C overnight followed by washing with
phosphate-buffered saline (PBS) 3 times. Coverslips were then
incubated with fluorescein isothiocyanate (FITC)- or Texas Red
(TR)-conjugated anti-mouse or anti-rabbit secondary antibodies
(1:120; Santa Cruz Biotechnology) for 30 min at room temperature
prior to staining with 4-6-diamidino-2-phenylindole (DAPI;
Invitrogen, Carlsbad, CA, USA).
Cell migration analysis
Refer to our previous study for details (20).
Wound-healing assay
Refer to our previous study for details (20).
miRNA target validation
The full-length PTRF 3′ untranslated regions
(3′UTRs) were amplified by PCR and cloned downstream of the firefly
luciferase gene in the psiCHECK-2 vector (Promega, Madison, WI,
USA). This vector was named the wild-type (wt) 3′UTR. Site-directed
mutagenesis of the miR-187 binding site in the PTRF 3′UTR was
conducted with the GeneTailor Site-Directed Mutagenesis System
(Invitrogen) and named the mutant (mt) 3′UTR. For reporter assays,
the wt or mt 3′UTR and miR-187 mimic or inhibitor were
cotransfected. The luciferase activity was assessed 2 days after
transfection using the Dual-Luciferase Reporter Assay System
(Promega). Primer sequences used for PCR assays were as follows: WT
PTRF forward, 5′-CCGCTCGAGGAGGCAGCCCCCGCAGTGGACAAC-3′ and reverse,
5′-ATAAGAATGCGGCCGCGAAAAGCAGGTTTATTGGTCGGGC-3′; MutPTRF-1 forward,
5′-TGATTCTGTTTGGACTGGGTTCTCATCTGTGCTCCAGGTTCTCAAGACACGAGTCCCC-3′
and reverse
5′-GGGGACTCGTGTCTTGAGAACCTGGAGCACAGATGAGAACCCAGTCCAAACAGAATCA-3′;
MutPTRF-2 forward,
5′-TTCTCAAGACACGACCAGGTTCTCATCTGTGCTGTCCCCTTGTTCCTCCCCATTAAAG-3′
and reverse
5′-CTTTAATGGGGAGGAACAAGGGGACAGCACAGATGAGAACCTGGTCGTGTCTTGAGAA-3′;
and MutPTRF-3 forward,
5′-TTCTCATCTGTGCTCCAGGTTCTCATCTGTGCTGTCCCCTTGTTCCTCCCCATTAAAG-3′
and reverse
5′-CTTTAATGGGGAGGAACAAGGGGACAGCACAGATGAGAACCTGGAGCACAGATGAGAA-3′.
Statistical analysis
Data were analyzed using SPSS software (version
19.0; SPSS, Inc., Chicago, IL, USA). For the qRT-PCR, Transwell and
luciferase reporter assays, the Students t-test or one-way analysis
of variance (ANOVA) was carried out. All data are presented as the
mean ± standard deviation (SD). Statistical significance was
established when the P-value was <0.05.
Results
miR-187 is upregulated in NSCLC
tissues and cell lines
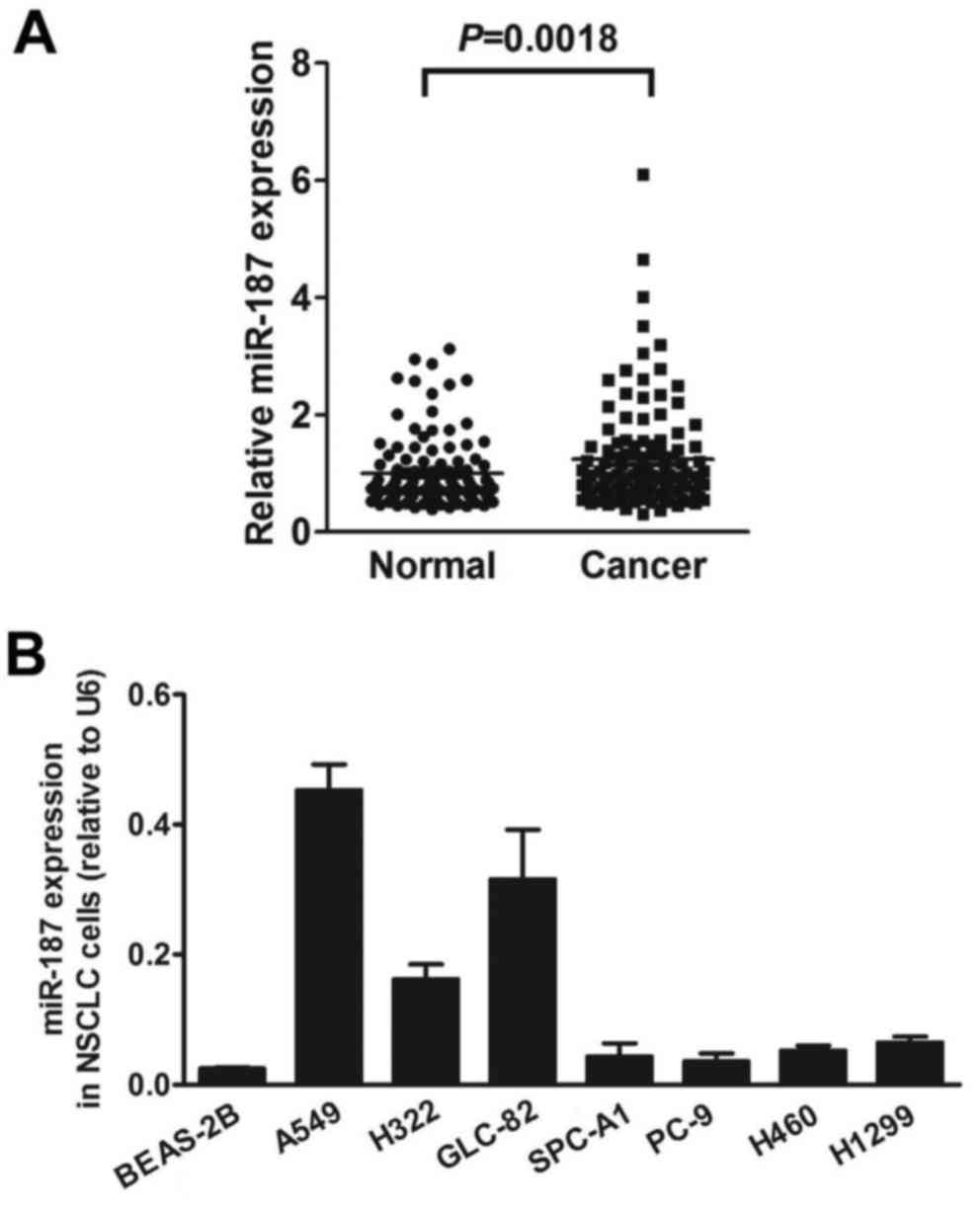
Microarray analysis showed that miR-187 was
upregulated at an average of 1.26-fold (P=0.0018) in the primary
lung neoplasms compared with the corresponding adjacent normal lung
tissues. Up to a 4.99-fold change in miR-187 expression was found
in 71 of all 116 NSCLC samples compared to the controls (Fig. 1A). Furthermore, increased expression
of miR-187 was found in all 7 NSCLC cell lines compared with the
normal human bronchial epithelial cell line BEAS-2B (Fig. 1B).
Exogenous miR-187 promotes NSCLC cell
migration and motility in vitro
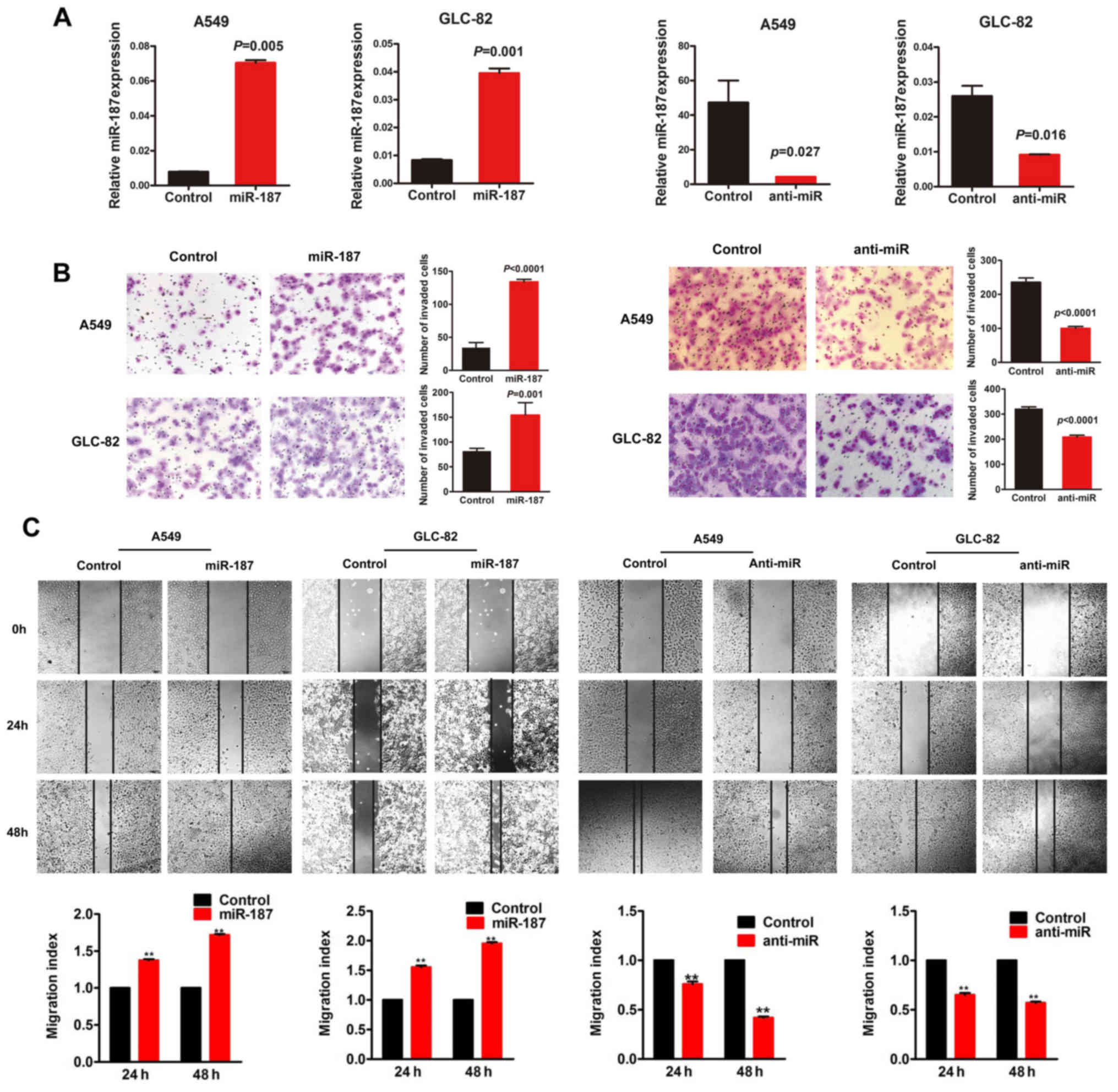
In the present study, miR-187 mimic oligonucleotides
were transfected into A549 and GLC-82 cell lines to determine their
effects on cellular behavior. In contrast, anti-miR-187 was
utilized to observe the effect of miR-187-depletion on cellular
behavior as an miRNA inhibitor. qRT-PCR was performed to detect the
transfection efficiency (P<0.05; Fig. 2A). miR-187-treated cells showed a
remarkable increase in migration and motility potential in
Transwell and wound-healing assays, whereas depletion of endogenous
miR-187 significantly decreased migration and motility (P<0.05;
Fig. 2B and C).
miR-187 mediates EMT and activates
signal transduction pathways
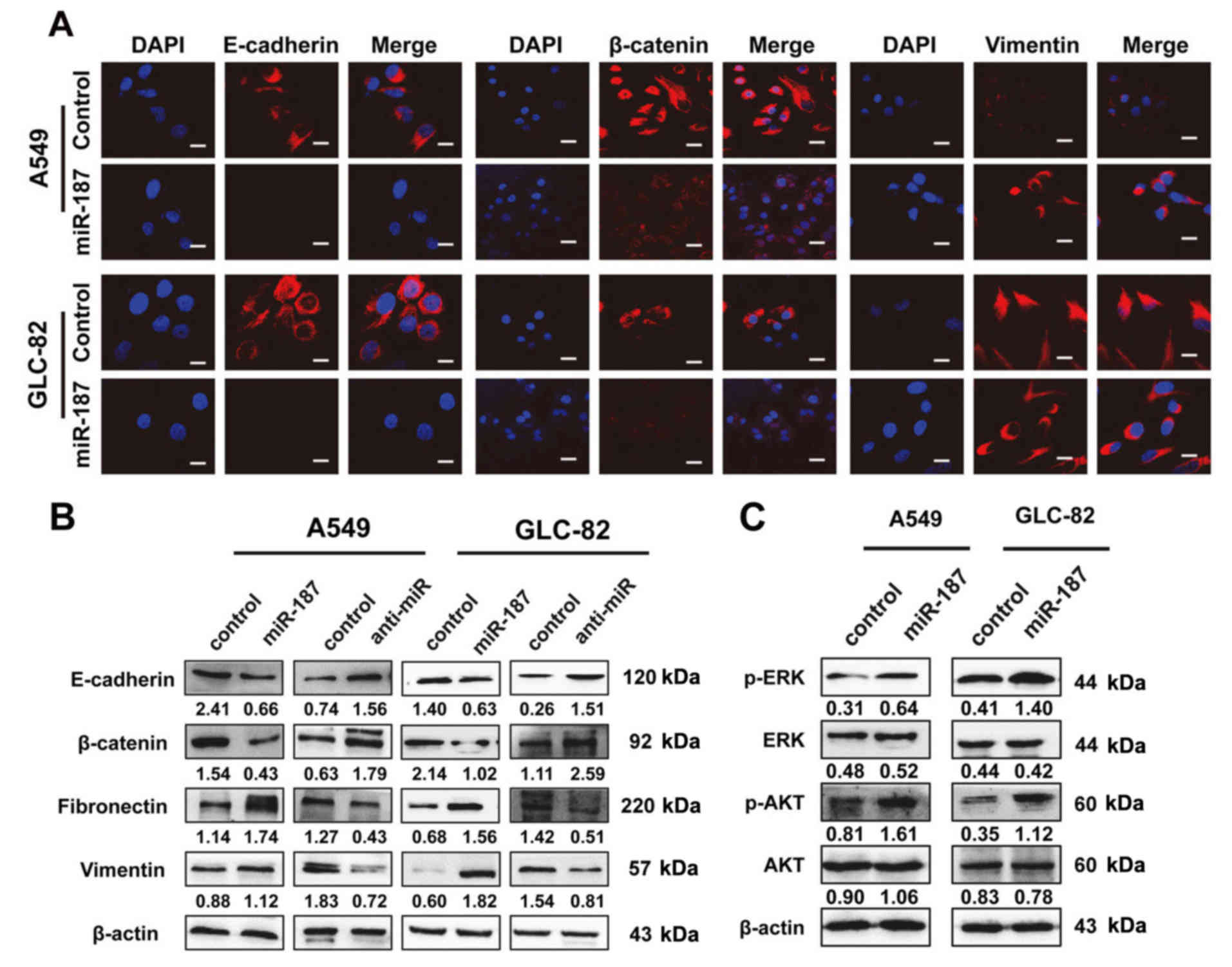
To investigate the mechanisms underlying
miR-187-mediated biological behavior, we investigated its effects
on EMT and signal transduction pathways. IF assays revealed that
exogenous miR-187 overexpression resulted in decreased expression
of the epithelial markers E-cadherin and β-catenin, and increased
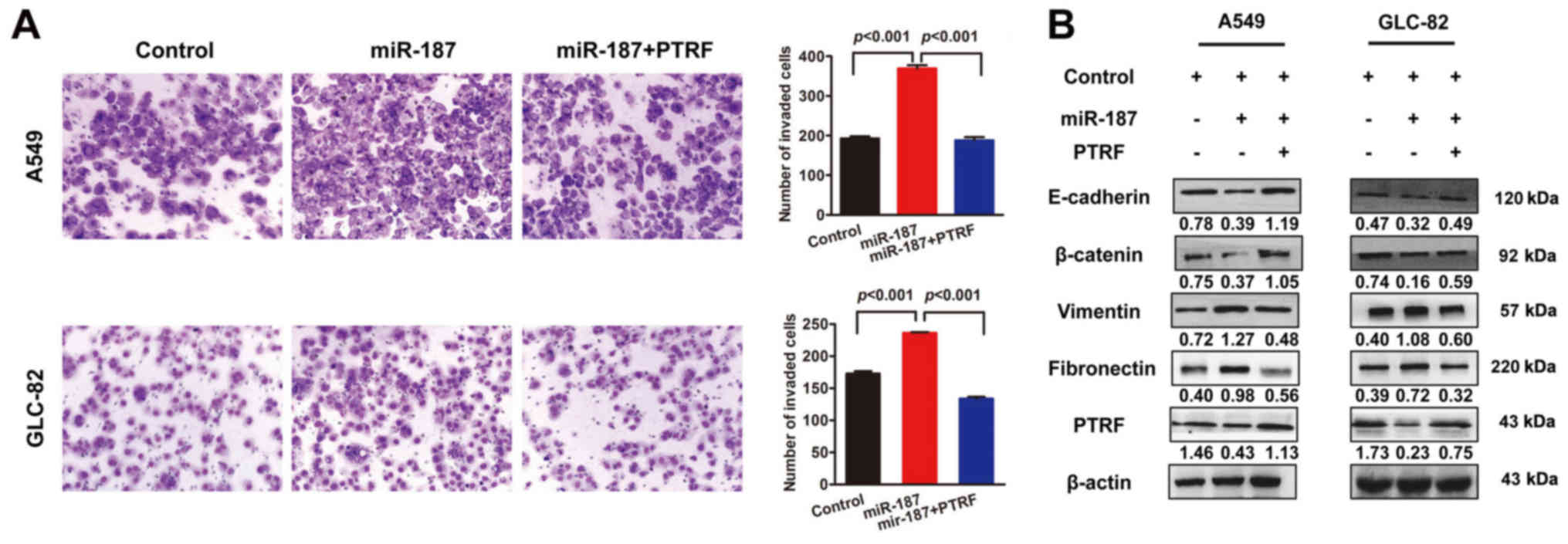
expression of the mesenchymal marker vimentin (Fig. 3A). Western blot analysis revealed
similar changes in EMT markers (Fig.
3B) and indicated that the phosphorylation status of proteins
was involved in EMT signaling. As shown in Fig. 3C, miR-187 activated the
mitogen-activated protein kinase (MAPK) pathway through the
phosphorylation of p44/42 MAPK (ERK1/2) and phosphatidylinositol
3-kinase/protein kinase B (PI3K/AKT) signaling via the
phosphorylation of AKT in A549 and GLC-82 cells.
PTRF is a direct target of
miR-187
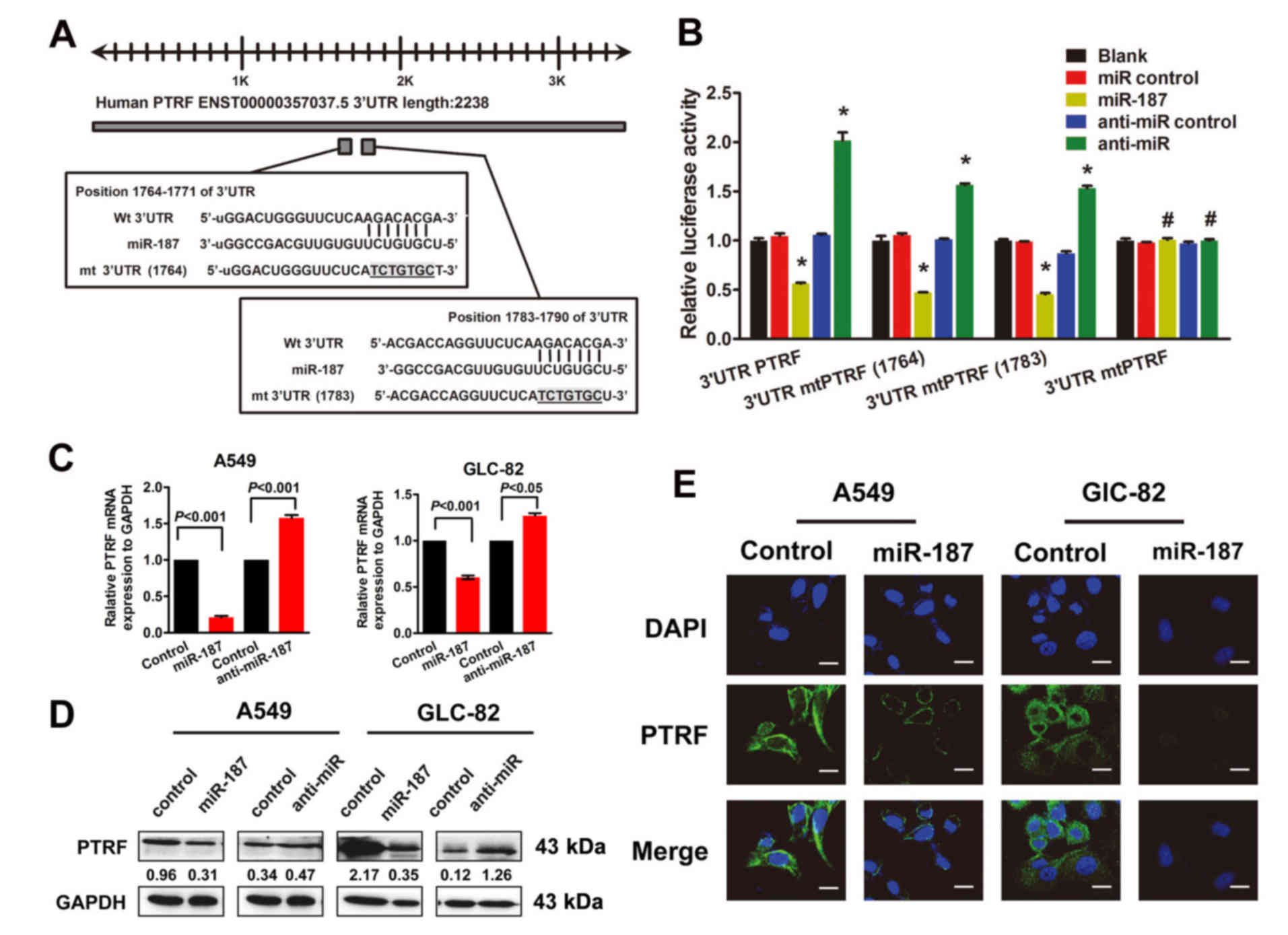
As indicated by computer-based sequence analysis
(based on TargetScan Human 6.2, PicTar and miRanda), PTRF was
identified as a potential target of miR-187 and therefore
considered to be involved in cell migration. The target sequence
(wt 3′UTR) or 2 mutant sequences (mt 3′UTR position 1764–1790) were
cloned into a luciferase reporter vector (Fig. 4A), then 293T cells were transfected
with the wt or mt 3′UTR vector and an miR-187 mimic. A significant
decrease in luciferase activity was observed in both the wt and mt
vectors compared with the miR controls, whereas increased activity
was found in cells cotransfected with anti-miR-187 and the wt or mt
3′UTR vectors (Fig 4B). Unlike for
double mutations, single mutations in the putative binding sites in
the PTRF 3′UTR region could not abrogate this suppression, thereby
providing strong evidence of direct interaction between miR-187 and
PTRF (Fig. 4B). In general, these
results strongly suggest that PTRF is a direct target of miR-187 in
NSCLC cells.
Additionally, qRT-PCR revealed decreased expression
of PTRF in A549 and GLC-82 cells transfected with miR-187 (Fig. 4C). Western blot analysis revealed
that transfection of miR-187 led to a markedly decreased expression
of PTRF proteins in NSCLC cells (Fig.
4D), which were confirmed by IF assays to be potential target
genes regulated by miR-187 (Fig.
4E).
PTRF plays crucial roles in
miR-187-induced EMT and migration of NSCLC cells
To further explore the function and role of PTRF in
NSCLC, we performed gain-of-function assays. Relatively decreased
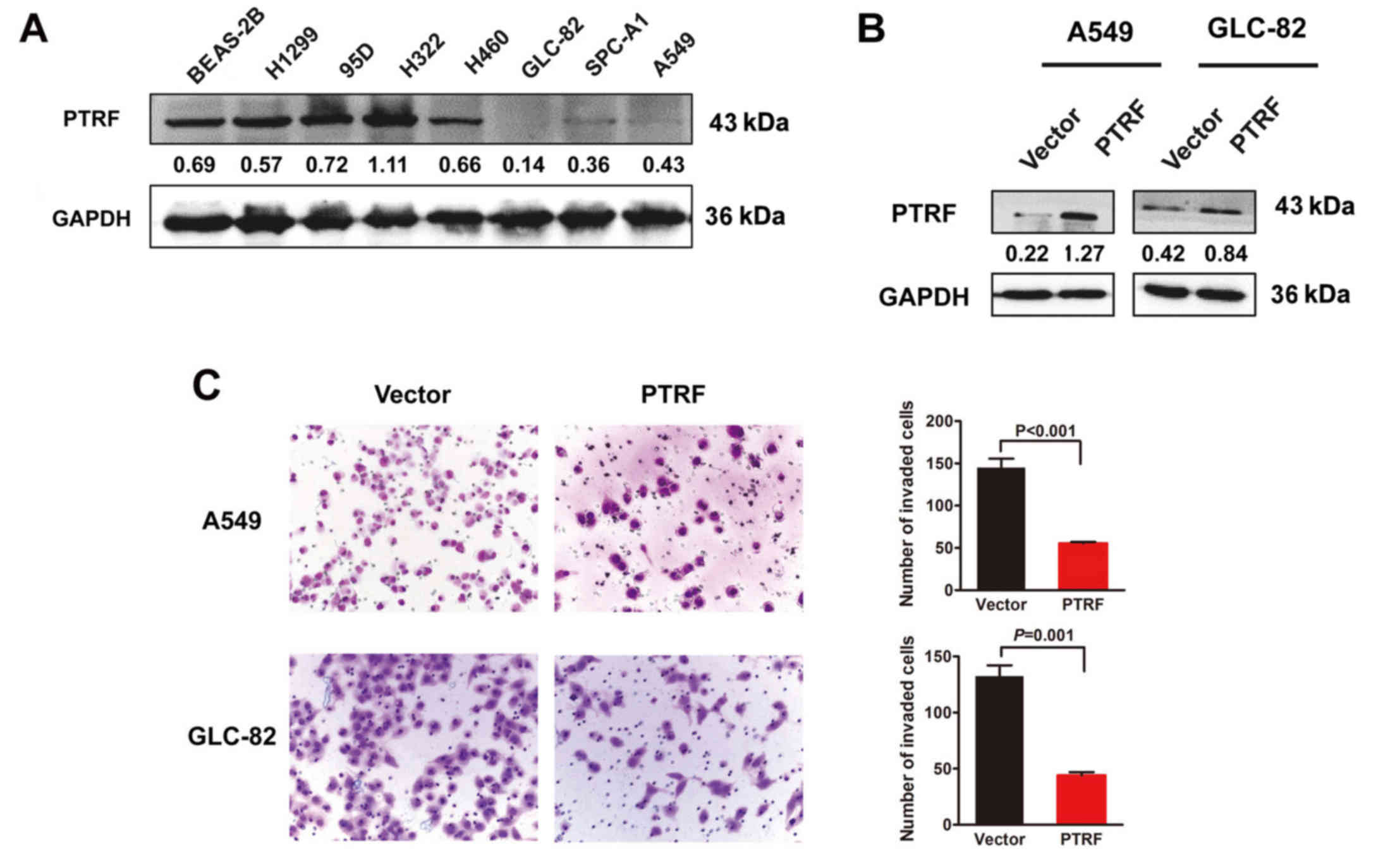
expression of PTRF was found in H460, GLC-82, SPC-A1 and A549 NSCLC
cell lines compared with the normal human bronchial epithelial cell
line BEAS-2B (Fig. 5A). Exogenous
introduction of PTRF markedly enhanced PTRF protein expression
(Fig. 5B) and decreased the
capacity of cell migration in A549 and GLC-82 cells (Fig. 5C).
To evaluate the biological role of PTRF in cell
migration and EMT induced by miR-187, we rescued PTRF expression in
miR-187-transfected NSCLC cells by transfection with PTRF ORF
constructs without 3′UTRs. The results revealed that exogenous
introduction of PTRF clearly suppressed miR-187-induced promotion
of migratory potential in A549 and GLC-82 cells (Fig. 6A). Consistent with the biological
phenotype results, re-expression of PTRF recovered miR-187-induced
EMT (Fig. 6B), strongly supporting
PTRF as a key mediator of miR-187-induced cell migration.
Discussion
Our present results highlight the importance of
upregulated miR-187 expression for NSCLC cells to develop and/or
sustain their aggressive phenotype. Aberrant expression of miR-187
has been reported in various types of cancer (14–18).
Although changes in expression vary greatly, miR-187 may
participate in tumorigenesis in a tissue-specific manner. However,
data on the effects of miR-187 in tumor progression are
controversial. For example, in one study, antisense-induced
suppression of miR-187 caused the inhibition of cell viability in
HeLa cells (23), whereas
overexpression of miR-187 induced apoptosis and led to decreased
proliferation of HeLa cells in another study (24). Hence, enriching our knowledge
regarding specific tumors may contribute to better clinical
management. In NSCLC, miR-187 was found to be a potential tumor
suppressor in tumor development (19), but the precise molecular mechanism
through which miR-187 influences NSCLC progression remains largely
unknown. In the present study, miR-187 was demonstrated to be a
potential promoter of NSCLC progression. However, the mechanism
underlying the abnormal expression of miR-187 remains unclear. The
present data did not support that abnormal histone or DNA
methylation was involved in abnormal expression. Our previous data
revealed that miR-187 may be regulated by cell factors in
colorectal cancer (CRC), such as transforming growth factor-β
(TGFβ). Numerous recent studies have revealed that signaling
pathways or non-coding RNAs also regulate miRNA expression
(25,26). We may investigate the regulatory
mechanism of miR-187 in more detail in future studies.
The mechanism underlying the relationship between
miR-187 and tumor aggressiveness remains to be elucidated. EMT is a
critical step in tumor progression. Recently, the role of miRNAs in
EMT has become a focus in the field of cancer research. Several
studies have demonstrated that miR-200 family members act as key
regulators of EMT to enforce the epithelial phenotype (27). The suppressive role of miR-187 in
ovarian cancer cell lines promoted EMT and was reported to be
achieved through targeting of disabled homolog 2 (Dab2) to decrease
migration and E-cadherin expression (28). However, these are currently the only
observations and suggestions for the underlying molecular
mechanisms. In the present study, miR-187 suppressed the epithelial
phenotype and induced mesenchymal transition, strongly suggesting
its critical role in the EMT process.
TGFβ is a known inducer of EMT via Smads and
complementary non-Smad pathways, such as MAPK (29–31)
and PI3K/AKT (32–34). In our previous study, miR-187
suppressed not only the Smad pathway but also non-Smad pathways in
CRC. Restoring expression of miR-187 only partially neutralized
TGFβ-mediated activation of the Smad pathway by decreasing the
phosphorylation level of Smad2 (20). In NSCLC, our data revealed that
miR-187 could activate both the MAPK and PI3K/AKT pathways,
implying that miR-187 induced NSCLC cell EMT via a non-Smad
pathway, not the classical Smad pathway. Therefore, downstream
targets must be explored in miR-187-induced EMT, and may be useful
in designing novel specific targeted drugs for managing NSCLC
metastasis. However, the specific regulator of miR-187 as a
metastasis promoter is still unclear.
In general, miRNAs exert their biological function
by suppressing their specific target genes at the
post-transcriptional level. Recently, B7-H3 (a novel member of the
B7 family) and Dab2 were reported as target genes of miR-187 in
renal and ovarian cancers, respectively (15,28).
SOX4, NT5E and PTK6 expression was increased in CRC cells treated
with TGFβ, and these effects were offset by the addition of
miR-187, suggesting that they are required for TGFβ-induced EMT
(20). However, we were not able to
identify them as targets directly regulated by miR-187 in NSCLC. As
previously mentioned, miR-187 may act in a cell- or organ-specific
manner through the alteration of the expression of target genes,
further changing their phenotypes. In the present study, all the
evidence indicates that PTRF may be a direct target of miR-187.
Rescued expression of PTRF restored the effect of miR-187 treatment
and inactivated the downstream pathway.
PTRF, also known as cavin-1, was originally
identified as a protein involved in the dissociation of
transcription complexes in vitro (35). PTRF in the cell surface membrane is
associated with vesicular transport, cholesterol homeostasis
(36,37) and lipolysis control (38). To date, the majority of studies
related to PTRF have focused on prostate cancer (PC). Changes in
the cell membrane involving loss of PTRF expression occur with the
development of PC (39).
Overexpression of PTRF in PC3 cells decreased cell motility by
decreasing matrix metalloprotease 9 (MMP9) production (40). The absence of PTRF in PC cells was
found to contribute significantly to tumor progression and
metastasis by promoting the angiogenesis and lymphangiogenesis
potential of cancer cells (41).
One study supports a role for PTRF/cavin-1, through caveolae
formation, as an attenuator of the non-caveolar functionality of
Cav1 in Gal3-Cav1 signaling and regulation of focal adhesion
dynamics and cancer cell migration (42). In addition, loss of PTRF expression
has been demonstrated to be related with tumor progression in
breast (43), pancreatic (44) and glioblastoma (33) cancers. Only one publication based on
proteomic assays reported PTRF/cavin-1 loss-of-expression in NSCLC
tissue at the protein level, suggesting a potential role for PTRF
in NSCLC development (45). Our
results demonstrated for the first time that PTRF contributes to
NSCLC progression through its involvement in EMT development.
In short, our results provide a basis for the
concept that increased expression of miR-187 in human NSCLC may be
significant in the acquisition of an aggressive phenotype. We
believe that miR-187 functions as a promoter in NSCLC progression
and may serve as a novel therapeutic biomarker. Moreover, the
functional and/or mechanistic studies of miR-187 presented in the
present study, indicate that miR-187 may play a critical role in
controlling non-Smad-mediated EMT by regulating PTRF expression.
This suggests that activating EMT during tumor metastasis in NSCLC
may be counteracted by suppressing miR-187, a notion that can be
readily tested in the clinic.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 81572813, 81272762 and
81401874), and the Guangdong Natural Science Foundation (nos.
S2013010014254 and 2014A030313490).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paez JG, Jänne PA, Lee JC, Tracy S,
Greulich H, Gabriel S, Herman P, Kaye FJ, Lindeman N, Boggon TJ, et
al: EGFR mutations in lung cancer: Correlation with clinical
response to gefitinib therapy. Science. 304:1497–1500. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eccles SA and Welch DR: Metastasis: Recent
discoveries and novel treatment strategies. Lancet. 369:1742–1757.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin SY, Lee YX, Yu SL, Chang GC and Chen
JJ: Phosphatase of regenerating liver-3 inhibits invasiveness and
proliferation in non-small cell lung cancer by regulating the
epithelial-mesenchymal transition. Oncotarget. 7:21799–21811.
2016.PubMed/NCBI
|
|
7
|
Yao Y, Shi M, Liu S and Li Y, Guo K, Ci Y,
Liu W and Li Y: MARVELD1 modulates cell surface morphology and
suppresses epithelial-mesenchymal transition in non-small cell lung
cancer. Mol Carcinog. 55:1714–1727. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ambros V, Lee RC, Lavanway A, Williams PT
and Jewell D: MicroRNAs and other tiny endogenous RNAs in C.
elegans. Curr Biol. 13:807–818. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu K, Ding H, Wang W, Liao Z, Fu Z, Hong
Y, Zhou Y, Zhang CY and Chen X: Tumor-suppressive miR-218-5p
inhibits cancer cell proliferation and migration via EGFR in
non-small cell lung cancer. Oncotarget. 7:28075–28085.
2016.PubMed/NCBI
|
|
10
|
Ma N, Zhang W, Qiao C, Luo H, Zhang X, Liu
D, Zang S, Zhang L and Bai J: The tumor suppressive role of
miRNA-509-5p by targeting FOXM1 in non-small cell lung cancer. Cell
Physiol Biochem. 38:1435–1446. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou L, Di Q, Sun B, Wang X, Li M and Shi
J: MicroRNA-194 restrains the cell progression of non-small cell
lung cancer by targeting human nuclear distribution protein C.
Oncol Rep. 35:3435–3444. 2016.PubMed/NCBI
|
|
12
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kasinski AL and Slack FJ: Epigenetics and
genetics. MicroRNAs en route to the clinic: Progress in validating
and targeting microRNAs for cancer therapy. Nat Rev Cancer.
11:849–864. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen HC, Chen GH, Chen YH, Liao WL, Liu
CY, Chang KP, Chang YS and Chen SJ: MicroRNA deregulation and
pathway alterations in nasopharyngeal carcinoma. Br J Cancer.
100:1002–1011. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao J, Lei T, Xu C, Li H, Ma W, Yang Y,
Fan S and Liu Y: MicroRNA-187, down-regulated in clear cell renal
cell carcinoma and associated with lower survival, inhibits cell
growth and migration though targeting B7-H3. Biochem Biophys Res
Commun. 438:439–444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bloomston M, Frankel WL, Petrocca F,
Volinia S, Alder H, Hagan JP, Liu CG, Bhatt D, Taccioli C and Croce
CM: MicroRNA expression patterns to differentiate pancreatic
adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA.
297:1901–1908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Casanova-Salas I, Rubio-Briones J,
Calatrava A, Mancarella C, Masiá E, Casanova J, Fernández-Serra A,
Rubio L, Ramírez-Backhaus M, Armiñán A, et al: Identification of
miR-187 and miR-182 as biomarkers of early diagnosis and prognosis
in patients with prostate cancer treated with radical
prostatectomy. J Urol. 192:252–259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Y and Stallings RL: Differential
patterns of microRNA expression in neuroblastoma are correlated
with prognosis, differentiation, and apoptosis. Cancer Res.
67:976–983. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun C, Li S, Yang C, Xi Y, Wang L, Zhang F
and Li D: MicroRNA-187-3p mitigates non-small cell lung cancer
(NSCLC) development through down-regulation of BCL6. Biochem
Biophys Res Commun. 471:82–88. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Luo Y, Shao Z, Xu L, Liu X, Niu
Y, Shi J, Sun X, Liu Y, Ding Y, et al: MicroRNA-187, a downstream
effector of TGFβ pathway, suppresses Smad-mediated
epithelial-mesenchymal transition in colorectal cancer. Cancer
Lett. 373:203–213. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tan X, Qin W, Zhang L, Hang J, Li B, Zhang
C, Wan J, Zhou F, Shao K, Sun Y, et al: A 5-microRNA signature for
lung squamous cell carcinoma diagnosis and hsa-miR-31 for
prognosis. Clin Cancer Res. 17:6802–6811. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang H, An H, Wang B, Liao Q, Li W, Jin X,
Cui S, Zhang Y, Ding Y and Zhao L: miR-133a represses tumour growth
and metastasis in colorectal cancer by targeting LIM and SH3
protein 1 and inhibiting the MAPK pathway. Eur J Cancer.
49:3924–3935. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park SY, Lee JH, Ha M, Nam JW and Kim VN:
miR-29 miRNAs activate p53 by targeting p85 alpha and CDC42. Nat
Struct Mol Biol. 16:23–29. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun C, Li S, Zhang F, Xi Y, Wang L, Bi Y
and Li D: Long non-coding RNA NEAT1 promotes non-small cell lung
cancer progression through regulation of miR-377-3p-E2F3 pathway.
Oncotarget. 7:51784–51814. 2016.PubMed/NCBI
|
|
26
|
Zhang X, Wang C, Shan S, Liu X, Jiang Z
and Ren T: TLR4/ROS/miRNA-21 pathway underlies lipopolysaccharide
instructed primary tumor outgrowth in lung cancer patients.
Oncotarget. 7:42172–42182. 2016.PubMed/NCBI
|
|
27
|
Gregory PA, Bracken CP, Bert AG and
Goodall GJ: MicroRNAs as regulators of epithelial-mesenchymal
transition. Cell Cycle. 7:3112–3118. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chao A, Lin CY, Lee YS, Tsai CL, Wei PC,
Hsueh S, Wu TI, Tsai CN, Wang CJ, Chao AS, et al: Regulation of
ovarian cancer progression by microRNA-187 through targeting
Disabled homolog-2. Oncogene. 31:764–775. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li NY, Weber CE, Wai PY, Cuevas BD, Zhang
J, Kuo PC and Mi Z: An MAPK-dependent pathway induces
epithelial-mesenchymal transition via Twist activation in human
breast cancer cell lines. Surgery. 154:404–410. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tomlinson DC, Baxter EW, Loadman PM, Hull
MA and Knowles MA: FGFR1-induced epithelial to mesenchymal
transition through MAPK/PLCγ/COX-2-mediated mechanisms. PLoS One.
7:e389722012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gui T, Sun Y, Shimokado A and Muragaki Y:
The roles of mitogen-activated protein kinase pathways in
TGF-β-induced epithelial-mesenchymal transition. J Signal
Transduct. 2012:2892432012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ning J, Liu W, Zhang J, Lang Y and Xu S:
Ran GTPase induces EMT and enhances invasion in non-small cell lung
cancer cells through activation of PI3K-AKT pathway. Oncol Res.
21:67–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Liu T, Bai Y, Liao H, Qiu S, Chang
Z, Liu Y, Yan X and Guo H: Polymerase I and transcript release
factor acts as an essential modulator of glioblastoma
chemoresistance. PLoS One. 9:e934392014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Massagué J: TGFβ signalling in context.
Nat Rev Mol Cell Biol. 13:616–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jansa P, Mason SW, Hoffmann-Rohrer U and
Grummt I: Cloning and functional characterization of PTRF, a novel
protein which induces dissociation of paused ternary transcription
complexes. EMBO J. 17:2855–2864. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cohen AW, Hnasko R, Schubert W and Lisanti
MP: Role of caveolae and caveolins in health and disease. Physiol
Rev. 84:1341–1379. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hamoudane M, Maffioli S, Cordera R, Maggi
D and Salani B: Caveolin-1 and polymerase I and transcript release
factor: New players in insulin-like growth factor-I receptor
signaling. J Endocrinol Invest. 36:204–208. 2013.PubMed/NCBI
|
|
38
|
Aboulaich N, Ortegren U, Vener AV and
Strålfors P: Association and insulin regulated translocation of
hormone-sensitive lipase with PTRF. Biochem Biophys Res Commun.
350:657–661. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gould ML, Williams G and Nicholson HD:
Changes in caveolae, caveolin, and polymerase 1 and transcript
release factor (PTRF) expression in prostate cancer progression.
Prostate. 70:1609–1621. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Aung CS, Hill MM, Bastiani M, Parton RG
and Parat MO: PTRF-cavin-1 expression decreases the migration of
PC3 prostate cancer cells: Role of matrix metalloprotease 9. Eur J
Cell Biol. 90:136–142. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nassar ZD, Moon H, Duong T, Neo L, Hill
MM, Francois M, Parton RG and Parat MO: PTRF/Cavin-1 decreases
prostate cancer angiogenesis and lymphangiogenesis. Oncotarget.
4:1844–1855. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Meng F, Joshi B and Nabi IR: Galectin-3
overrides PTRF/Cavin-1 reduction of PC3 prostate cancer cell
migration. PLoS One. 10:e01260562015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bai L, Deng X, Li Q, Wang M, An W, Deli A,
Gao Z, Xie Y, Dai Y and Cong YS: Down-regulation of the cavin
family proteins in breast cancer. J Cell Biochem. 113:322–328.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu L, Xu HX, Wang WQ, Wu CT, Chen T, Qin
Y, Liu C, Xu J, Long J, Zhang B, et al: Cavin-1 is essential for
the tumor-promoting effect of caveolin-1 and enhances its
prognostic potency in pancreatic cancer. Oncogene. 33:2728–2736.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gámez-Pozo A, Sánchez-Navarro I, Calvo E,
Agulló-Ortuño MT, López-Vacas R, Díaz E, Camafeita E, Nistal M,
Madero R, Espinosa E, et al: PTRF/cavin-1 and MIF proteins are
identified as non-small cell lung cancer biomarkers by label-free
proteomics. PLoS One. 7:e337522012. View Article : Google Scholar : PubMed/NCBI
|