Introduction
Hepatocellular carcinoma (HCC) is one of the most
common cancers worldwide and causes approximately 600,000 deaths
every year (1,2). Although liver transplantation and
surgical resection are the most efficient therapeutic strategies
for early stage HCC (3,4), the majority of patients, who are
usually diagnosed at an advanced stage, must rely on traditional
chemotherapies (5). Currently,
conventional systemic chemotherapies are still not effective to
treat patients with advanced HCC (6). Therefore, novel therapeutic agents
with high efficacy are urgently required for the clinical treatment
of advanced HCC.
As is known, the abnormal proliferation of cancer
cells plays an important role in the development of cancers such as
HCC (7). Therefore, apoptosis is
considered as a potent therapeutic target for cancer treatment. It
is believed that apoptosis can be triggered by diverse
pro-apoptosis stimuli converging on mitochondria, which induces
mitochondrial dysfunction such as mitochondrial depolarization,
cytochrome c release, and caspase enzyme activation (8,9). In
addition, mitochondria are the primary source of endogenous
reactive oxygen species (ROS) and the targets of ROS in turn
(9). Tumor cells with a higher
level of ROS can cause redox imbalance, oxidative DNA damage and
caspase-dependent or -independent activation of the
mitochondria-mediated intrinsic apoptotic pathway (10). Therefore, novel therapeutic agents
with high potential in promoting ROS production are urgently
required for the clinical treatment of advanced HCC.
Thioredoxin interaction protein (TXNIP) is a 46 kDa
protein that was first identified in an HL-60 leukemia cell line
treated with 1α, 25-dihydroxyvitamin D3 [1,
25(OH2) D3] (11). As a negative regulator of
thioredoxin (TRX), TXNIP was found to be an inducer of the
intracellular level of reactive oxygen species (ROS) (12). A number of studies have confirmed
that TXNIP shows strong growth suppressive, metastasis inhibitory
and proapoptotic functions (13,14).
Subsequently, it has been identified as a tumor suppressor gene in
various hematological malignancies and solid tumors, such as breast
cancer and thyroid cancer (15–17).
Overexpression of TXNIP inhibited tumor growth and caused cell
cycle arrest and apoptosis. TXNIP was also shown to play a role as
a transcriptional repressor of cyclin A2. TXNIP-deficient cells
were shown to upregulate Wnt pathway, reduce p21 and regulate
hematopoietic stem cell proliferation and cell quiescence (18,19).
However, the potential role of TXNIP in the development of HCC
remains unclear.
In this study, we first examined the expression of
TXNIP in adjacent non-tumor liver tissue and HCC tissues. We found
that the expression of INPP4B directly correlated with TNM stage in
patients with HCC. Furthermore, we transfected SMMC7221 cells with
a pcDNA-3.1-TXNIP plasmid to study the effect and mechanism of
TXNIP on the proliferation and apoptosis of HCC cells in
vitro. Our clinical and mechanistic data indicated that TXNIP
may be a tumor suppressor involved in the progression of HCC.
Targeting the TXNIP pathway may be a potential treatment modality
for HCC.
Materials and methods
Patients and tissue samples
From January 2012 to December 2015, 105 HCC patients
with liver tumor who underwent surgical resection in Department of
General Surgery, the First Affiliated Hospital, Xinxiang Medical
University were collected. Thirty matched HCC tissue and the
adjacent non-tumor liver tissue (ANLT) specimens were obtained from
these HCC patients. Normal liver tissues were obtained from 30
patients with giant hemangioma during hepatic resection. Tissue
specimens were fixed by formalin immediately upon collection and
then paraffin-embedded. The pathological diagnosis for all cases
was made by at least two Board Certified pathologists working at
the Department of Pathology in the First Affiliated Hospital of
Xinxiang Medical University. Informed consent was obtained from
each patient and was approved by the Institute Research Ethics
Committee at Cancer Center (Xinxiang Medical University).
Immunohistochemistry
The HCC samples were sectioned at 4 µm. Sections
were deparaffinized with xylene and hydrated using graded alcohol;
antigen retrieval and blocking were then performed, and slides were
incubated with polyclonal rabbit anti-human TXNIP (Bioworld
Technology), followed by incubation with secondary antibodies.
Detection was performed by 3,3′-diaminobenzidine (DAB) and
hematoxylin. The slides were next evaluated by two pathologists in
a blinded fashion. All immunohistochemistry staining was
independently assessed by two experienced pathologists. The
staining intensity was graded from 0 to 2 (0, no staining; 1, weak;
2, strong). The staining extent was graded from 0 to 4 based on the
percentage of immunoreactive tumor cells (0, 1–5, 6–25, 26–75 and
76–100%). A score ranging from 0 to 8 was calculated by multiplying
the staining extent score with the staining intensity score,
resulting in a low (0–4) level or a high (6–8) level
for each sample (20).
Cell lines
L02 cells were obtained from the Cancer Research
Institute of Xi'an Jiaotong University. HepG2 cells were purchased
from the American Type Culture Collection (ATCC, Rockville, MD,
USA). SMCC7221 and HCCLM3 cells were obtained from the Cell Bank of
Shanghai Institute of Biochemistry and Cell Biology, Chinese
Academy of Sciences (Shanghai, China) and cultured in RPMI-1640
(Invitrogen, Carlsbad, CA, USA) medium containing 10% fetal bovine
serum, 100 U/ml penicillin, and 10 µg/ml streptomycin at 37°C in
humidified air containing 5% CO2.
Plasmid constructs and
transfection
The Full-length TXNIP coding region was obtained by
RT-PCR amplification of normal human liver cDNA and then subcloned
into a pcDNA3.1 vector. For transfection, SMMC7221 cells were
cultured to 80% confluence and transfected with 4 µg
pcDNA-3.1-TXNIP plasmid using Lipofectamine 2000 (Invitrogen)
according to the manufacturer's recommendations. The
pcDNA-3.1-scramble plasmid was used as a negative control. Cells
were harvested after 48 h for subsequent analysis.
Cell proliferation analysis
Cells (1×105 cells/well) were seeded in
96-well plates and cultured for 24, 48 and 72 h. After the cell
culture medium was removed, cells were washed three times with
phosphate-buffered saline (PBS), and then incubated in 100 µl
freshly DMEM medium containing MTT (0.2 mg/ml) at 37°C for 4 h. The
supernatant was removed and 200 µl DMSO was added for 30 min to
dissolve the formazan crystals. The absorbance was measured at a
wavelength of 570 nm using a microplate reader (Thermo Fisher
Scientific, Inc., Waltham, UK), and the results were used to
calculate the viability of the cells.
Apoptosis analysis
Cell apoptosis was analyzed by flow cytometry based
on the Annexin V-FITC/propidium iodide (PI) apoptosis kit (Strong
Biotech Corp., Taipei, Taiwan) according to the manufacturer's
instructions. Briefly, cells were seeded in a 6-well plate at the
density 3×105 cells/well and cultured for 24 h, then
harvested by trypsinization and centrifuged at 2000 rpm for 5 min,
followed by washing twice with ice-cold PBS. After adding 100 µl
Annexin V binding buffer, 5 µl Annexin V and 5 µl PI, the samples
were incubated at room temperature for 15 min in the dark, followed
by the addition of Annexin V binding buffer to bring the total
volume to 1 ml. The cells were then transferred into a flow
cytometry tube and analyzed using a flow cytometer (Cytomics FC
500, Beckman Coulter, Atlanta, CA, USA).
Determination of ROS production
The oxidant sensitive fluorescent probe DCFH-DA was
used to determine intracellular ROS generation. Cells were seeded
in a 96-well plate at the density 1×104 cells/well and
cultured for 24 h, then incubated with DCFH-DA for 30 min at 37°C
in the dark. After being washed twice with PBS, fluorescence images
of the cells were acquired using the InCell 2000 confocal
microscope. For the quantitative evaluation of intracellular ROS
production efficacy, the fluorescence was measured by the software
modules supplied with the InCell 2000.
Measurement of the mitochondrial
membrane potential
The mitochondrial membrane potential was measured
using JC-1, which was a mitochondria-specific lipophilic cationic
fluorescence dye and was capable of selectively entering the
mitochondria. Cells were seeded in a 96-well plate at the density
1×104 cells/well and cultured for 24 h, then the cell
culture media was removed and incubated with 100 µl JC-1 stain
solution (200 µM) for 20 min at 37°C in the dark. After washing
twice with PBS, the cells on coverslips were mounted onto slides
and examined with the InCell 2000 confocal microscope. Quantitative
image analysis was carried out using the software modules supplied
with the InCell 2000.
Mitochondrial complex II activity
For mitochondrial complex II activity assay,
SMMC7221 cells were cultured in 6-well plates at the density
1×104 cells/well for 24 h. The cells were collected and
the activity was detected according to Succinate Dehydrogenase
Activity Assay kit (BioVision, Palo Alto, CA, USA). The absorbance
was measured at 600 nm.
Quantitative real-time polymerase
chain reaction
For HCC and ANLT tissues, the total RNA was isolated
using the RecoverAll™ Total Nucleic Acid Isolation kit (Ambion,
Austin, TX, USA) according to the manufacturer's protocol. Total
mRNA of cells was extracted with TRIzol reagent (Invitrogen)
according to the manufacturer's instructions. First strand cDNA was
generated from 2 µg total RNA using RevertAid™ First Strand cDNA
Synthesis kit (Fermentas, Glen Burnie, MD, USA). qRT-PCR was
performed in triplicate utilizing the CFX Real-time PCR Detection
System (Bio-Rad, Hercules, CA, USA) and Absolute Blue QPCR SYBR
Green Mix (Applied Biosystems, Foster City, CA, USA). The cycle
number when the fluorescence first reached a preset threshold (Ct)
was used to quantify the initial concentration of individual
templates for expression of the mRNA of genes of interest.
Transcripts of the housekeeping gene GAPDH in the same incubations
were used for internal normalization. Primer pairs were as follows:
TXINP-F: 5′-TGGCAAAGGAGCAGATTAGTTGCTTAGG-3′, and TXNIP-R:
5′-CTGCCACAAGAACTCTGTGAAATTGGATATTAGA-3′; and GAPDH-F:
5′-CATGCGAACAGCAGTCACTTGCTGATGTAT-3′, and GAPDH-R:
5′-ATTTTAGGACCCTGCGTCGGCCGTCTG-3′.
Western blot analysis
Cells were homogenized in ice-cold radio
immunoprecipitation assay lysis buffer (Santa Cruz Biotechnology,
Santa Cruz, CA, USA) for 30 min and centrifuged at 12,000 × g at
4°C for 10 min, and the supernatants were collected. Protein
concentrations were determined using a protein assay dye reagent
(Bio-Rad). Equal amounts (usually 50 µg) were loaded onto a 10%
SDS-PAGE gel and separated electrophoretically. Then, the proteins
were transferred to a PVDF membrane (Millipore, Boston, MA, USA).
After blocking with 5% non-fat dry milk in PBS with 0.1% Tween-20
(PBS-T) for 2 h at room temperature, the PVDF membrane was
incubated overnight at 4°C with primary antibodies, including
anti-SDHA (1:1500, Santa Cruz Biotechnology), anti-Bax (1:1000,
Cell Signaling Technology, Danvers, MA, USA), anti-Bcl-2 (1:1000,
Cell Signaling Technology), anti-cleaved caspase-9 (1:1000, Cell
Signaling Technology), anti-cleaved caspase-3 (1:1000, Cell
Signaling Technology), anti-p38 (1:1000, Cell Signaling
Technology), anti-p-p38 (1:1000, Cell Signaling Technology),
anti-ERK (1:1000, Cell Signaling), anti-p-ERK (1:1000, Cell
Signaling Technology), anti-JNK (1:1500, Cell Signaling
Technology), and anti-p-JNK (1:1000, Cell Signaling Technology),
respectively, followed by incubation with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG antibodies (1:2000, Sigma,
St. Louis, MO, USA) for 1 h at room temperature. Protein detection
was performed based on an enhanced chemiluminescence (ECL) method
and photographed using a BioSpectrum Gel Imaging System (HR410,
UVP, Upland, CA, USA). Detection of β-actin on the same membrane
was used as a loading control.
Statistical analysis
Data are presented as the mean ± SD. Differences
between two groups were analyzed by Student's t-test, while
differences between multiple groups were analyzed by ANOVA.
P-values <0.05 were considered to indicate a statistically
significant difference.
Results
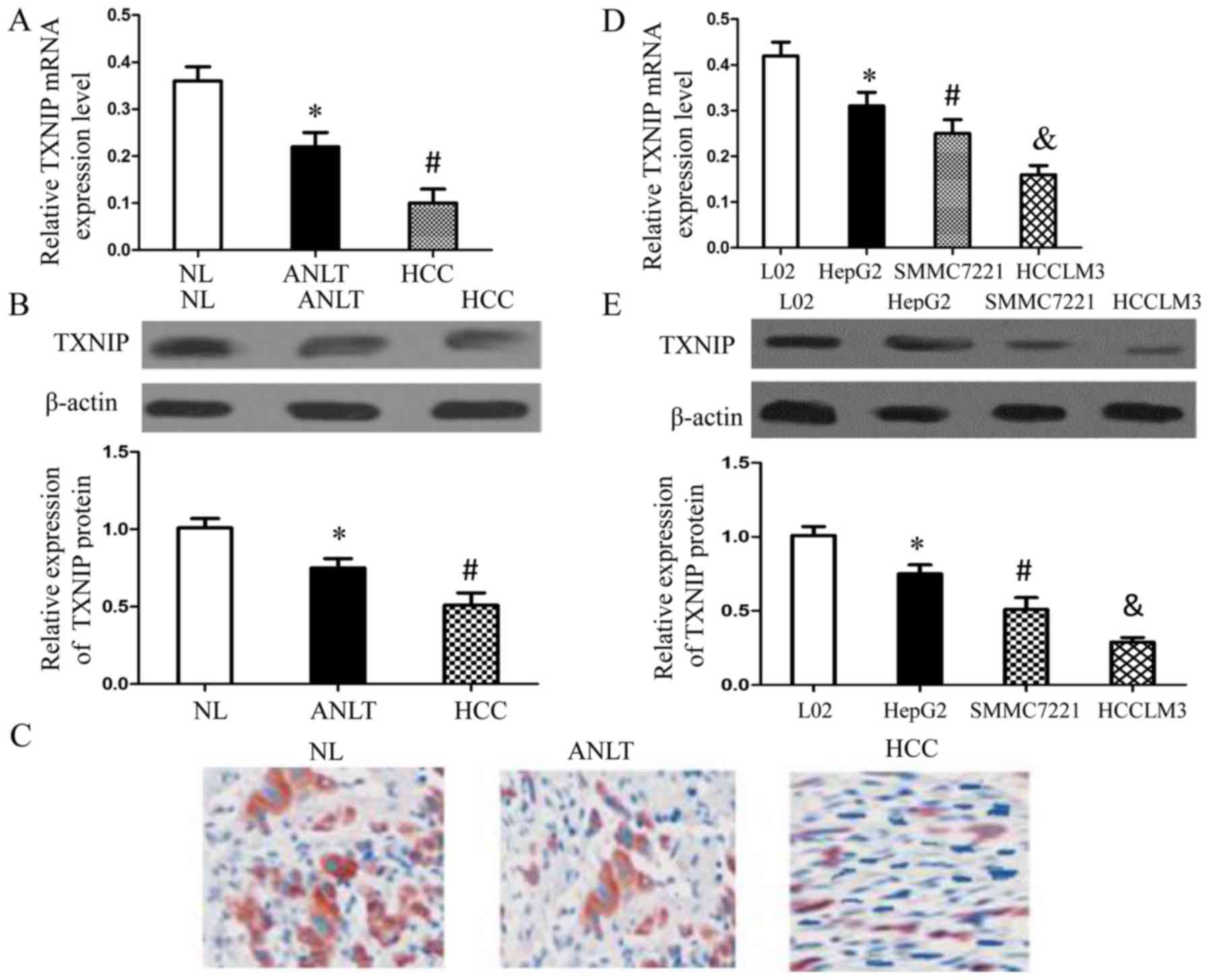
TXNIP expression is reduced in human
hepatocellular carcinoma
To determine whether TXNIP expression was changed in
human hepatocellular carcinoma, we first examined its mRNA
expression and protein expression in HCC tissues and the adjacent
non-tumor liver tissues (ANLT) isolated from the same patient as
well as in normal liver tissues (NL) obtained from patients with
giant hemangioma by qRT-PCR and western blotting, respectively.
There were 30 HCC patients randomly selected for the study. At the
same time, immunohistochemistry was also performed to analyze the
expression of TXNIP in HCC tissues, ANLT and NL. Noteworthy, when
compared with the mRNA level and protein level of TXNIP in the ANLT
tissues, both were significantly downregulated in HCC tissues
(Fig. 1A and B). The results of
immunohistochemistry showed that none or few cells showing
positively-stained cytoplasm were detected in HCC tissues and
diffuse strong brown in ANLT (Fig.
1C).
We further examined the differences of TXNIP
expression between three HCC cell lines and a normal liver cell
line (L02 cells) by qRT-PCR (Fig.
1D) and western blotting (Fig.
1E). Of note, TXNIP expression in cancer cells was negatively
correlated with cell metastatic potential, since HCCLM3 cells
possessed the highest capability for metastasis, while HepG2 cells
had the least capacity for metastasis. Combined with the above
results, we concluded that TXNIP expression was reduced in the
human hepatocellular carcinoma.
Correlation of TXNIP expression with
clinicopathological characteristics of HCC
Subsequently, the association of TXNIP expression
with the clinicopathological features of HCC was analyzed. Overall,
105 HCC patients were stratified into the low and high expression
groups according to the results of immunohistochemistry. The
correlations of TXNIP expression with clinicopathological
characteristics were analyzed. It was found that the TXNIP
expression was related with tumor nodule number, vascular invasion
and TNM (Table I). These results
together revealed that decreased TXNIP expression possibly
participated in HCC progression.
 | Table I.The correlations of TXNIP with
clinicopathological features of HCC. |
Table I.
The correlations of TXNIP with
clinicopathological features of HCC.
|
|
| TXNIP |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
variable | n | Low expression | High
expression | P-value |
|---|
| Gender |
|
|
| 0.150 |
|
Female | 67 | 38 | 29 |
|
|
Male | 38 | 16 | 22 |
|
| Age |
|
|
| 0.296 |
|
<60 | 46 | 21 | 25 |
|
|
≥60 | 59 | 33 | 26 |
|
| AFP |
|
|
| 0.393 |
| <20
ng/ml | 91 | 45 | 46 |
|
| ≥20
ng/ml | 14 | 9 | 5 |
|
| HBsAg |
|
|
| 0.872 |
|
Negative | 85 | 44 | 41 |
|
|
Positive | 20 | 10 | 10 |
|
| Tumor nodule
number |
|
|
| 0.003 |
|
Solitary | 31 | 9 | 22 |
|
|
Multiple (>2) | 74 | 45 | 29 |
|
| Vascular
invasion |
|
|
| 0.029 |
|
Absence | 93 | 34 | 59 |
|
|
Presence | 12 | 10 | 2 |
|
| TNM |
|
|
| 0.039 |
| I | 55 | 23 | 32 |
|
|
II–III | 50 | 31 | 19 |
|
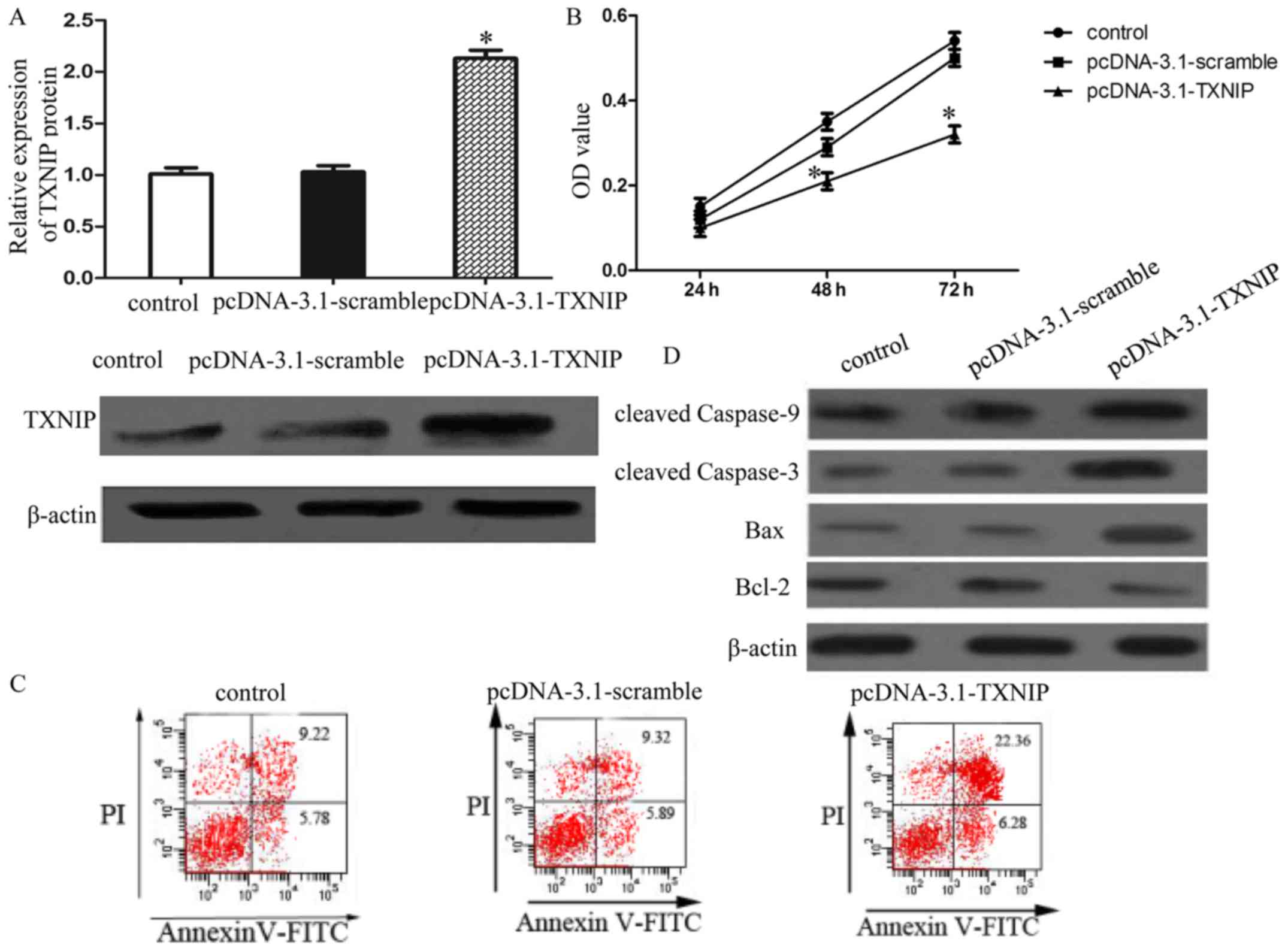
TXNIP overexpression significantly
inhibits SMMC7221 cell proliferation and induces apoptosis
To functionally characterize the role of TXNIP in
HCC cell proliferation and apoptosis, MTT assay and flow cytometry
were performed. SMMC7221 cells were transfected with
pcDNA-3.1-TXNIP plasmid and the TXNIP expression was confirmed by
western blot analysis (Fig. 2A).
The results of proliferation analysis and apoptosis showed that
enforced TXNIP expression inhibited SMMC7221 cells proliferation
and induced apoptosis (Fig. 2B and
C). Furthermore, TXNIP overexpression increased the expression
of cleaved caspase-9 and −3, as well as increased the expression of
pro-apoptotic protein Bax and suppressed the anti-apoptotic protein
Bcl-2 (Fig. 2D). Collectively,
these data suggested that TXNIP overexpression induced growth
arrest and mitochondrial-related apoptosis in SMMC7221 cells.
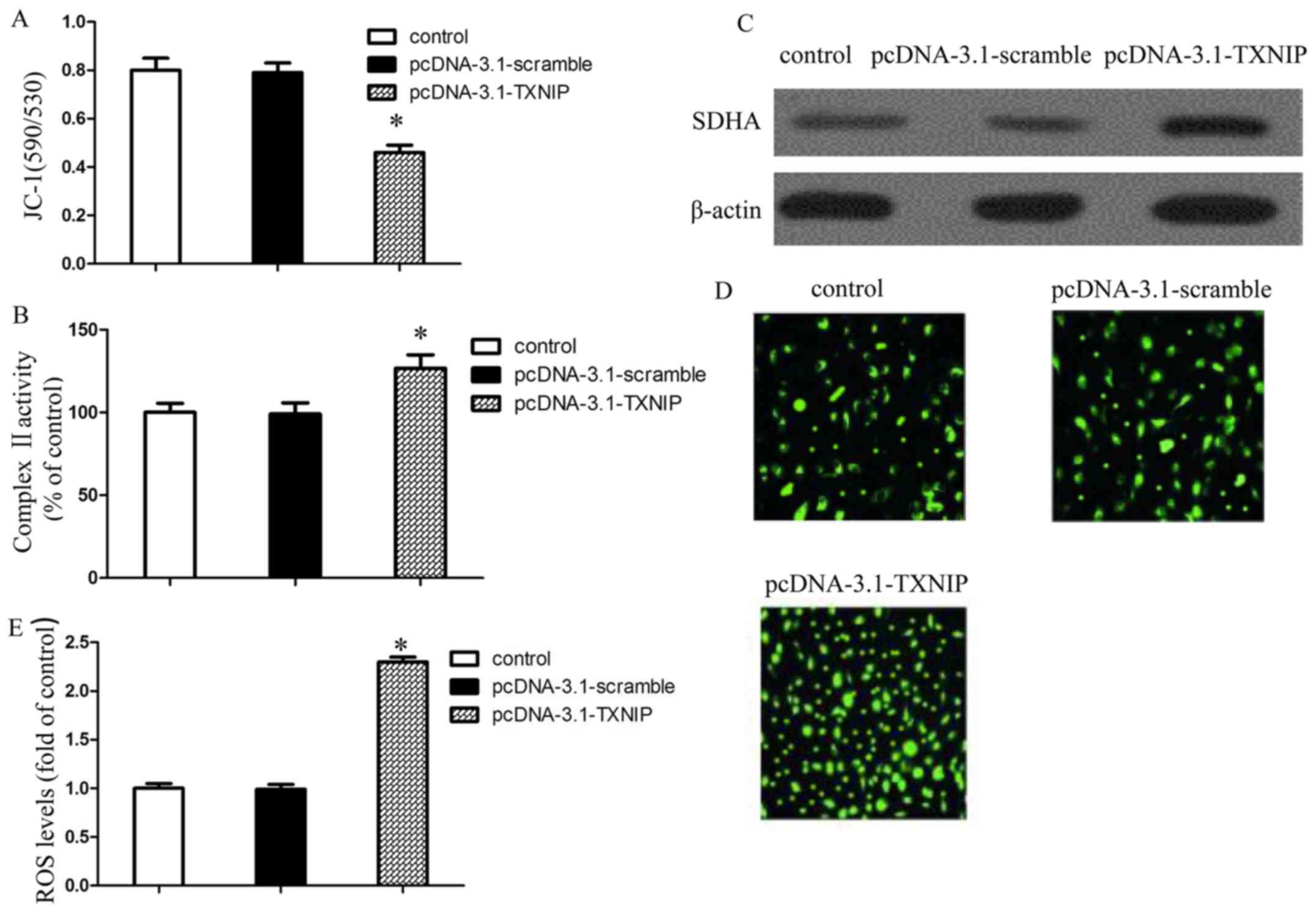
TXNIP overexpression increases ROS
generation via impaired mitochondrial function
Mitochondrial dysfunction is closely related to the
mitochondrial-related apoptosis. In this study, JC-1 staining was
performed to detect the mitochondrial membrane potential in
SMMC7221 cells. Fig. 3A shows that
the mitochondrial membrane potential of SMMC7221 was decreased
after transfection with pcDNA-3.1-TXNIP plasmid, which indicated
that TXNIP overexpression induced mitochondrial dysfunction.
Moreover, TXNIP overexpression significantly increased the
mitochondrial complex II activity of SMMC7221 and promoted the
expression of SDHA, a subunit of complex II which is a major site
for ROS generation (Fig. 3B and C).
To determine whether ROS generation was involved in TXNIP
overexpression-induced apoptosis, we measured the levels of ROS in
SMMC7221-treated cells using DCF-DA reagent (Fig. 3D). Our results revealed that TXNIP
overexpression could increase the ROS generation (Fig. 3E). Combined with the above results,
we concluded that TXNIP overexpression could increase the ROS
generation via impaired mitochondrial function.
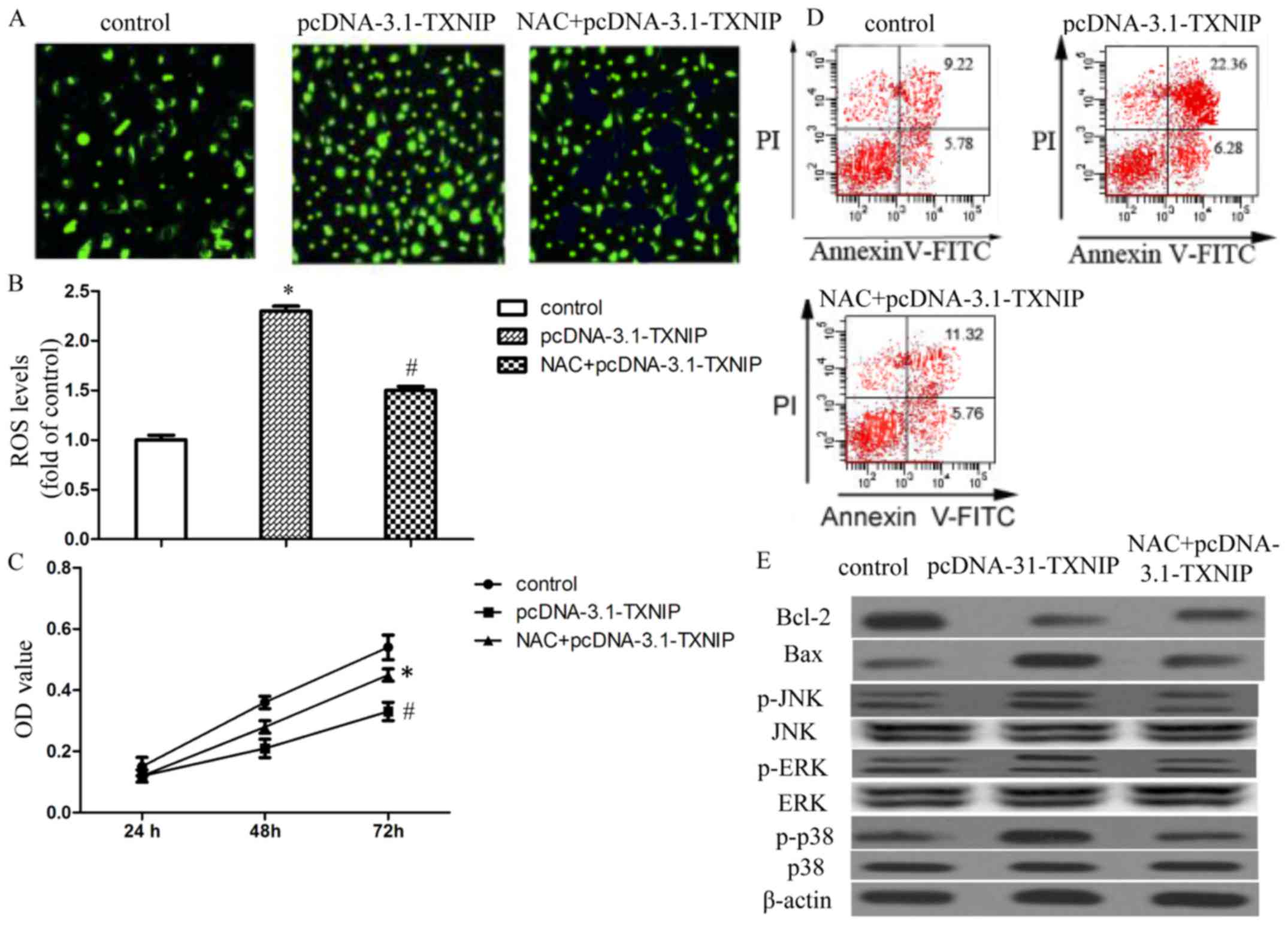
ROS generation is involved in TXNIP
overexpression-induced SMMC7221 cell growth arrest and
apoptosis
In order to assess the roles of ROS generation in
TXNIP overexpression-induced SMMC7221 cell growth arrest and
apoptosis, we treated SMMC7221 cells with ROS scavenger NAC before
transfection with the pcDNA-3.1-TXNIP plasmid. Our results revealed
that NAC markedly attenuated TXNIP-induced ROS generation in
SMCC7221 cells (Fig. 4A and B). NAC
also alleviated the inhibition effects of TXNIP overexpression on
SMMC7221 proliferation, as well as cancer cell apoptosis (Fig. 4C and D). We also examined the
effects of NAC on TXNIP overexpression-induced variation of
apoptosis-related proteins. Our results revealed that NAC obviously
attenuated TXNIP overexpression-induced downregulation of Bcl-2 and
upregulation of Bax (Fig. 4E). The
results suggested that ROS generation may be involved in TXNIP
overexpression-induced growth arrest and apoptosis.
TXNIP overexpression activates MAPK
pathways in a ROS- dependent manner
We next sought to identify the signal transduction
pathways involved in proliferation inhibition in response to TXNIP
overexpression. We monitored the expression levels and activation
status of MAPKs, which are also the major oxidative
stress-sensitive signal transducing pathways and serve as upstream
signals for the initiation of apoptosis. The results showed that
TXNIP overexpression significantly increased the levels of
phosphorylated JNK, ERK and p38 without affecting the expression of
total proteins. Interestingly, the ROS scavenger NAC significantly
attenuated the TXNIP overexpression-induced activation of JNK, ERK
and p38 MAPK (Fig. 4E). Our data
suggested that TXNIP overexpression could activate the JNK, ERK and
p38 MAPK via ROS.
Discussion
TXNIP was initially identified as a
thioredoxin-binding protein that inhibits thioredoxin (TRX),
thereby contributing to redox homeostasis (21). Since TRX also promotes tumor
progression by angiogenesis induction (22) and apoptosis inhibition (23,24),
TXNIP, as its inhibitor, has great potential as a tumor suppressor
gene. Several studies have shown that the expression of TXNIP was
significantly reduced in different types of aggressive cancers such
as acute myeloid leukemia (AML), breast cancer and bladder cancer,
which emphasized its relevance in tumor prevention (25–29).
In this study, we found that TXNIP expression was decreased
significantly in most HCC tissues, suggesting a potential
involvement of TXNIP in the development of HCC. It has been
reported that TNXIP is an inhibitor of TRX, which could suppress
tumor cell apoptosis. Hence, we transfected pcDNA-3.1-TXNIP plasmid
into SMMC7221 cells with the purpose of exploring the effect of
TXNIP overexpression on HCC cell proliferation and apoptosis.
Our results showed that TXNIP overexpression
significantly inhibited SMCC7221 cell proliferation and induced
apoptosis (Fig. 2B and C). It has
been confirmed that apoptosis could be induced through an intrinsic
or extrinsic pathway (30–32), and activation of the caspase cascade
was also involved in apoptosis (33). Only when caspase is cleaved can the
activation of caspase occur. Furthermore, activation of caspase-9,
an initiator caspase, is required for the activation of executioner
caspase (i.e., caspase-3) to induce apoptosis (34). In our study, TXNIP overexpression
obviously activated caspase-9 and −3, which led to the induction of
apoptosis. Although it is still unknown whether an intrinsic or
extrinsic pathway was the key pathway inducing apoptosis in
SMCC7221 cells transfected with pcDNA-3.1-TXNIP plasmid, TXNIP
overexpression-induced apoptosis involved the intrinsic pathway
that was associated with mitochondrial dysfunction.
Mitochondria-dependent apoptosis is regulated by proteins of the
Bcl-2 family. Our results showed that TXNIP overexpression promoted
the expression of a proapoptotic protein (Bax) and inhibits the
expression of an antiapoptotic protein (Bcl-2) (Fig. 2D). In addition, SMMC7221 cells
transfected with pcDNA-3.1-TXNIP plasmid exhibited a lower
mitochondrial membrane potential (Fig.
3A); combined with the above results, we concluded that TXNIP
overexpression could induce apoptosis in HCC cells through
mitochondrial dysfunction.
Mitochondria are not only vital organelles for
producing ATP and intermediates for eukaryotic cancer cells, they
are also key sources of ROS generation (35–37).
Mitochondrial complex II, known as succinate dehydrogenase (SDH),
is the only complex that participates in the Krebs cycle as well as
the electron transport chain (38).
Ralph et al (39) reported
that the succinate dehydrogenase (SDH)/complex II system could act
as a key redox regulator of ROS production via an electron driving
mechanism. In our study, TXNIP overexpression significantly
increased the activity of complex II and promoted the expression of
SDHA, as well as increasing the production of intracellular ROS. It
is well known that the intrinsic apoptotic pathway is especially
susceptible to ROS (40). In the
present study, the inhibition of ROS production by the antioxidant
NAC significantly alleviated TXNIP overexpression-mediated growth
arrest and apoptosis (Fig. 4C and
D), suggesting that TXNIP overexpression-induced apoptosis in
SMMC7221 cells was closely associated with the production of ROS,
and may act as upstream signaling molecules to initiate
mitochondria-mediated cell apoptosis.
MAPKs are the major oxidative stress-sensitive
signal transducing pathways and are also a family of
serine/threonine kinases that regulate a variety of cellular events
such as proliferation and apoptosis (41), at the same time serving as upstream
signals for the initiation of apoptosis (42). Further, ROS functions as second
messengers in diverse signaling pathways (43). Our results showed that TXNIP
overexpression significantly increased the levels of phosphorylated
JNK, ERK and p38 MAPK without affecting the expression of total
proteins (Fig. 4E), indicating that
these MAPK pathways were activated in the process of TXNIP
overexpression-induced apoptosis in SMMC7221 cells. Further, our
study also showed that pre-treatment with NAC prevented the
phosphorylation of JNK, ERK and p38 MAPK. Therefore, we concluded
that JNK, ERK and p38 MAPK were activated in response to TXNIP
overexpression-induced ROS generation, which are frequently
associated with the induction of apoptosis.
In summary, we demonstrated that TXNIP
overexpression induced apoptotic cell death in SMMC7221 cells
through intrinsic pathways via triggering mitochondrial-mediated
ROS generation and activating MAPK pathways. This study provided
insight into the molecular mechanisms of TXNIP
overexpression-induced apoptosis in liver cancer cells and
presented that TXNIP may serve as a potential therapeutic target
for HCC.
References
|
1
|
Bao H, Liu P, Jiang K, Zhang X, Xie L,
Wang Z and Gong P: Huaier polysaccharide induces apoptosis in
hepatocellular carcinoma cells through p38 MAPK. Oncol Lett.
12:1058–1066. 2016.PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Signoriello S, Annunziata A, Lama N,
Signoriello G, Chiodini P, De Sio I, Daniele B, Di Costanzo GG,
Calise F, Olivieri G, et al: Survival after locoregional treatments
for hepatocellular carcinoma: A cohort study in real-world
patients. Sci World J. 2012:5647062012. View Article : Google Scholar
|
|
4
|
Chan SL and Yeo W: Targeted therapy of
hepatocellular carcinoma: Present and future. J Gastroenterol
Hepatol. 27:862–872. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu SC: Where are we in the chemoprevention
of hepatocellular carcinoma? Hepatology. 51:734–736.
2010.PubMed/NCBI
|
|
6
|
Wang X, Wang N, Cheung F, Lao L, Li C and
Feng Y: Chinese medicines for prevention and treatment of human
hepatocellular carcinoma: Current progress on pharmacological
actions and mechanisms. J Integr Med. 13:142–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li H, Miao Q, Xu CW, Huang JH, Zhou YF and
Wu MJ: OTX1 contributes to hepatocellular carcinoma progression by
regulation of ERK/MAPK pathway. J Korean Med Sci. 31:1215–1223.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sepand MR, Ghahremani MH,
Razavi-Azarkhiavi K, Aghsami M, Rajabi J, Keshavarz-Bahaghighat H
and Soodi M: Ellagic acid confers protection against
gentamicin-induced oxidative damage, mitochondrial dysfunction and
apoptosis-related nephrotoxicity. J Pharm Pharmacol. 68:1222–1232.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zeng CC, Lai SH, Yao JH, Zhang C, Yin H,
Li W, Han BJ and Liu YJ: The induction of apoptosis in HepG-2 cells
by ruthenium(II) complexes through an intrinsic ROS-mediated
mitochondrial dysfunction pathway. Eur J Med Chem. 122:118–126.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Diao QX, Zhang JZ, Zhao T, Xue F, Gao F,
Ma SM and Wang Y: Vitamin E promotes breast cancer cell
proliferation by reducing ROS production and p53 expression. Eur
Rev Med Pharmacol Sci. 20:2710–2717. 2016.PubMed/NCBI
|
|
11
|
Zhu HJ, Wang DG, Yan J and Xu J:
Up-regulation of microRNA-135a protects against myocardial
ischemia/reperfusion injury by decreasing TXNIP expression in
diabetic mice. Am J Transl Res. 7:2661–2671. 2015.PubMed/NCBI
|
|
12
|
Hong K, Xu G, Grayson TB and Shalev A:
Cytokines regulate β-cell thioredoxin-interacting protein (TXNIP)
via distinct mechanisms and pathways. J Biol Chem. 291:8428–8439.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hong SY and Hagen T: 2-Deoxyglucose
induces the expression of thioredoxin interacting protein (TXNIP)
by increasing O-GlcNAcylation - Implications for targeting the
Warburg effect in cancer cells. Biochem Biophys Res Commun.
465:838–844. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen D, Dang BL, Huang JZ, Chen M, Wu D,
Xu ML, Li R and Yan GR: MiR-373 drives the
epithelial-to-mesenchymal transition and metastasis via the
miR-373-TXNIP-HIF1α-TWIST signaling axis in breast cancer.
Oncotarget. 6:32701–32712. 2015.PubMed/NCBI
|
|
15
|
Baldan F, Mio C, Lavarone E, Di Loreto C,
Puglisi F, Damante G and Puppin C: Epigenetic bivalent marking is
permissive to the synergy of HDAC and PARP inhibitors on TXNIP
expression in breast cancer cells. Oncol Rep. 33:2199–2206.
2015.PubMed/NCBI
|
|
16
|
Nie W, Huang W, Zhang W, Xu J, Song W,
Wang Y, Zhu A, Luo J, Huang G, Wang Y, et al: TXNIP interaction
with the Her-1/2 pathway contributes to overall survival in breast
cancer. Oncotarget. 6:3003–3012. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morrison JA, Pike LA, Sams SB, Sharma V,
Zhou Q, Severson JJ, Tan AC, Wood WM and Haugen BR: Thioredoxin
interacting protein (TXNIP) is a novel tumor suppressor in thyroid
cancer. Mol Cancer. 13:622014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Filios SR, Xu G, Chen J, Hong K, Jing G
and Shalev A: MicroRNA-200 is induced by thioredoxin-interacting
protein and regulates Zeb1 protein signaling and beta cell
apoptosis. J Biol Chem. 289:36275–36283. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ho B, Huang G and Golubovskaya VM: Focal
adhesion kinase regulates expression of thioredoxin-interacting
protein (TXNIP) in cancer cells. Anticancer Agents Med Chem.
14:3–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Zhang JB, Qin Y, Wang W, Wei L,
Teng Y, Guo L, Zhang B, Lin Z, Liu J, et al: PROX1 promotes
hepatocellular carcinoma metastasis by way of up-regulating
hypoxia-inducible factor 1α expression and protein stability.
Hepatology. 58:692–705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nishiyama A, Matsui M, Iwata S, Hirota K,
Masutani H, Nakamura H, Takagi Y, Sono H, Gon Y and Yodoi J:
Identification of thioredoxin-binding protein-2/vitamin D(3)
up-regulated protein 1 as a negative regulator of thioredoxin
function and expression. J Biol Chem. 274:21645–21650. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Welsh SJ, Bellamy WT, Briehl MM and Powis
G: The redox protein thioredoxin-1 (Trx-1) increases
hypoxia-inducible factor 1alpha protein expression: Trx-1
overexpression results in increased vascular endothelial growth
factor production and enhanced tumor angiogenesis. Cancer Res.
62:5089–5095. 2002.PubMed/NCBI
|
|
23
|
Saitoh M, Nishitoh H, Fujii M, Takeda K,
Tobiume K, Sawada Y, Kawabata M, Miyazono K and Ichijo H: Mammalian
thioredoxin is a direct inhibitor of apoptosis signal-regulating
kinase (ASK) 1. EMBO J. 17:2596–2606. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y and Min W: Thioredoxin promotes ASK1
ubiquitination and degradation to inhibit ASK1-mediated apoptosis
in a redox activity-independent manner. Circ Res. 90:1259–1266.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou J, Bi C, Cheong LL, Mahara S, Liu SC,
Tay KG, Koh TL, Yu Q and Chng WJ: The histone methyltransferase
inhibitor, DZNep, up-regulates TXNIP, increases ROS production, and
targets leukemia cells in AML. Blood. 118:2830–2839. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hirotsu C, Tufik S, Bergamaschi CT,
Tenorio NM, Araujo P and Andersen ML: Sleep pattern in an
experimental model of chronic kidney disease. Am J Physiol Renal
Physiol. 299:F1379–F1388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sheth SS, Bodnar JS, Ghazalpour A,
Thipphavong CK, Tsutsumi S, Tward AD, Demant P, Kodama T, Aburatani
H and Lusis AJ: Hepatocellular carcinoma in Txnip-deficient mice.
Oncogene. 25:3528–3536. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Butler LM, Zhou X, Xu WS, Scher HI,
Rifkind RA, Marks PA and Richon VM: The histone deacetylase
inhibitor SAHA arrests cancer cell growth, up-regulates
thioredoxin-binding protein-2, and down-regulates thioredoxin. Proc
Natl Acad Sci USA. 99:pp. 11700–11705. 2002; View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nishizawa K, Nishiyama H, Matsui Y,
Kobayashi T, Saito R, Kotani H, Masutani H, Oishi S, Toda Y, Fujii
N, et al: Thioredoxin-interacting protein suppresses bladder
carcinogenesis. Carcinogenesis. 32:1459–1466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fox JL and MacFarlane M: Targeting cell
death signalling in cancer: Minimising ‘Collateral damage’. Br J
Cancer. 115:5–11. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matthews GM, Newbold A and Johnstone RW:
Intrinsic and extrinsic apoptotic pathway signaling as determinants
of histone deacetylase inhibitor antitumor activity. Adv Cancer
Res. 116:165–197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wajant H: The Fas signaling pathway: More
than a paradigm. Science. 296:1635–1636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shimizu H, Banno Y, Sumi N, Naganawa T,
Kitajima Y and Nozawa Y: Activation of p38 mitogen-activated
protein kinase and caspases in UVB-induced apoptosis of human
keratinocyte HaCaT cells. J Invest Dermatol. 112:769–774. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu YJ, Wong BS, Yea SH, Lu CI and Weng SH:
Sinularin induces apoptosis through mitochondria dysfunction and
inactivation of the pI3K/Akt/mTOR pathway in gastric carcinoma
cells. Mar Drugs. 14:142016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zong WX, Rabinowitz JD and White E:
Mitochondria and Cancer. Mol Cell. 61:667–676. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Weinberg SE and Chandel NS: Targeting
mitochondria metabolism for cancer therapy. Nat Chem Biol. 11:9–15.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Byun HO, Kim HY, Lim JJ, Seo YH and Yoon
G: Mitochondrial dysfunction by complex II inhibition delays
overall cell cycle progression via reactive oxygen species
production. J Cell Biochem. 104:1747–1759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang L, Zhang X, Cui G, Chan JY, Wang L,
Li C, Shan L, Xu C, Zhang Q, Wang Y, et al: A novel agent exerts
antitumor activity in breast cancer cells by targeting
mitochondrial complex II. Oncotarget. 7:32054–32064.
2016.PubMed/NCBI
|
|
39
|
Ralph SJ, Moreno-Sánchez R, Neuzil J and
Rodríguez-Enríquez S: Inhibitors of succinate: Quinone
reductase/Complex II regulate production of mitochondrial reactive
oxygen species and protect normal cells from ischemic damage but
induce specific cancer cell death. Pharm Res. 28:2695–2730. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang C, Jia X, Bao J, Chen S, Wang K,
Zhang Y, Li P, Wan JB, Su H, Wang Y, et al: Polyphyllin VII induces
apoptosis in HepG2 cells through ROS-mediated mitochondrial
dysfunction and MAPK pathways. BMC Complement Altern Med.
16:582016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Takeda K, Matsuzawa A, Nishitoh H and
Ichijo H: Roles of MAPKKK ASK1 in stress-induced cell death. Cell
Struct Funct. 28:23–29. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zebelo SA and Maffei ME: Role of early
signalling events in plant-insect interactions. J Exp Bot.
66:435–448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Avisetti DR, Babu KS and Kalivendi SV:
Activation of p38/JNK pathway is responsible for embelin induced
apoptosis in lung cancer cells: Transitional role of reactive
oxygen species. PLoS One. 9:e870502014. View Article : Google Scholar : PubMed/NCBI
|