Introduction
Osteosarcoma (OS) is a common bone cancer that
mainly affects children and adolescents (1,2).
Although multiple treatments including local control interventions
and chemotherapy have improved the survival of patients with OS,
the survival rate is still ~60–80% in OS patients (3). Moreover, the 5-year event-free
survival for patients with high grade OS is still less than 50%
(3). Therefore, it is imperative to
develop more effective therapeutic strategies for the treatment of
OS. It has been demonstrated that genetic and cytogenetic
abnormalities are critically involved in the development and
progression of OS (4). For example,
mutations in tumor suppressors and activation of oncogenes are
associated with OS (5). Therefore,
it is essential to understand the molecular mechanism of OS
tumorigenesis and to validate novel therapeutic targets for OS.
Emerging evidence has revealed that several
biological molecules could be effective therapeutic targets for OS.
For instance, multiple genes such as Notch, Wnt, nuclear factor-κB
(NF-κB), p53, phosphoinositide 3-kinase (PI3K)/Akt and
mitogen-activated protein kinase (MAPK) are critical in the
development of OS (4). Tao et
al identified that Notch activation is a driver of osteogenic
sarcoma (6). Recent studies have
validated that S-phase kinase-associated protein 2 (Skp2) plays an
essential role in the development of various human malignancies.
Skp2 has been characterized to exhibit its oncogenic function via
targeting of its substrates including p27 (7,8), p21
(9), p57 (10) and Forkhead box protein O1 (FOXO1)
(11,12). Engineered mouse models revealed that
conditional depletion of Skp2 in mice suppressed tumor growth in T
cell lineage (13), B cell lineage
(14), bone marrow (15), liver (16,17),
breast (18), prostate (19) and skin cancer (20). Consistently, upregulation of Skp2 in
mice enhanced tumor growth in lymphoma (21), prostate cancer (19) and breast tumor (18). Importantly, high expression of Skp2
has been observed in a wider spectrum of cancers including
lymphomas (22,23), pancreatic cancer (24), breast carcinomas (25–29),
prostate (30,31) and gastric cancer (32), melanoma (33–35),
hepatocarcinoma (36) and
nasopharyngeal carcinoma (37,38).
Skp2 was also highly expressed and was correlated with relapse,
metastasis and survival in OS patients (39). However, the role of Skp2 in OS has
not been fully elucidated.
In the present study to explore the role of Skp2 in
OS, we depleted Skp2 in MG-63 and SW 1353 cells. Then, we
investigated the biological function of Skp2 in cell growth,
apoptosis, and in the cell cycle of OS cells after knockdown of
Skp2. Moreover, we assessed the effects of Skp2 depletion on OS
cell migration. Methodically, we determined whether the downstream
targets of Skp2 are involved in the progression of OS. We found
that depletion of Skp2 inhibited cell growth, induced apoptosis,
arrested the cell cycle and suppressed cell migration in OS cells.
Moreover, depletion of Skp2 exerted its function partly through the
regulation of Akt and p27 in OS cells. Our findings suggest that
targeting Skp2 may be a promising therapeutic strategy for the
treatment of OS.
Materials and methods
Cell culture, reagents and
antibodies
The human OS MG-63 and SW 1353 cells (ATCC,
Manassas, VA, USA) were cultured at 37°C in 5% CO2 in
Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS).
3-(4,5-Dimethythiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was obtained from Sigma-Aldrich (St. Louis, MO, USA). Anti-Akt and
anti-pAkt antibodies were obtained from Cell Signaling Technology
(Danvers, MA, USA). Antibodies against β-actin, Skp2 and p27 were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Transfection
Cells were seeded into 6-well plates and transfected
with control siRNA or Skp2 siRNAs (GenePharma, Shanghai, China)
using Lipofectamine 2000 according to the manufacturer's
protocol.
MTT assay
Cells were seeded at equal densities into 96-well
culture plates. After 24 h, the cells were then treated with Skp2
siRNA for 48 h. An MTT assay was conducted to determine the
absorbance at 560 nm using a Benchmark Microplate Reader (Bio-Rad,
Hercules, CA, USA). All values were normalized to those of the
control.
Cell cycle analysis
The transfected cells were seeded into 100-mm dishes
for 48 h. Then, the cells were fixed in ice-cold 70% ethanol in
phosphate-buffered saline (PBS) for 12 h. The fixed cells were
treated with 0.1 mg/ml RNase A for 20 min. The cells were
re-suspended in propidium iodide (PI) (50 µg/ml). Cell cycle
distribution was assessed using a FACScan flow cytometer and
analyzed.
Apoptosis assay
The transfected cells were cultured into a 6-well
plate for 48 h. Then, the cells were harvested and washed with PBS,
resuspended in 500 µl of binding buffer with 5 µl of PI and 5 µl of
FITC-conjugated anti-Annexin V antibody. Apoptosis was assessed by
a FACSCalibur flow cytometer.
Real-time RT-PCR analysis
Total RNA from transfected cells was isolated using
TRIzol and reversed-transcribed into cDNA by RevertAid First Strand
cDNA Synthesis kit according to the manufacturer's protocol. The
primers used in the PCR reactions were: Skp2 forward, 5′-GCT GCT
AAA GGT CTC TGG TGT-3′ and reverse, 5′-AGG CTT AGA TTC TGC AAC
TTG-3′; GAPDH forward, 5′-ACC CAG AAG ACT GTG GAT GG-3′ and
reverse, 5′-CAG TGA GCT TCC CGT TCA G-3′. The expression of GAPDH
was used as an internal control.
Western blot analysis
Cells were lysed in lysis buffer and protein
concentrations were detected by Brandford assay reagent. Equal
amounts of proteins were resolved on SDS-PAGE, and then transferred
to membranes. The membranes were immunoblotted with primary
antibodies including Skp2, Akt, phosphor-Akt-Ser473, p27 and
β-actin, followed by secondary antibodies conjugated with
horseradish peroxidase. Then, the bands were revealed using an
enzyme-linked chemiluminescence (ECL) detection kit assay.
Wound healing assay
Cells were cultured in 6-well plates and grown to
confluency. When cells converged to ~100%, monolayers of cells were
scratched with 200 µ yellow pipette tips and washed with PBS. The
scratched area was photographed with a microscope at 0 and 20 h,
respectively.
Transwell invasion assay
An invasion assay was carried out using BD BioCoat
Matrigel invasion chambers. Briefly, tranfected cells were seeded
in DMEM without serum in the upper chamber. DMEM containing 10% FBS
was added to the lower chamber. After overnight incubation, the
non-invading cells were removed. The cells that had invaded through
the Matrigel matrix membrane were stained with 4 µg/ml calcein AM
in Hanks' buffered saline at 37°C for 1 h. The fluorescence of the
invaded cells was read on Ultra Multifunctional Microplate Reader
(Tecan, Durham, NC, USA) at excitation/emission wavelengths of
530/590 nm. Invasiveness was assessed by calculating the invading
cells with a microscope.
Statistical analysis
The data are expressed as the mean ± SD. A Student's
t-test was performed to evaluate statistical significance between
the Skp2-siRNA transfection group and the NC-siRNA treated group.
The level of significance was considered as P<0.05.
Results
Skp2 expression is inhibited by its
siRNAs in OS cells
To determine whether Skp2 is involved in the
regulation of cell proliferation in OS cells, we depleted Skp2
using its specific siRNAs in both MG-63 and SW 1353 cells.
Real-time reverse transcription-polymerase chain reaction (RT-PCR)
and western blot analysis were applied for the detection of the
efficacy of multiple Skp2 siRNAs on inhibition of Skp2 in OS cells.
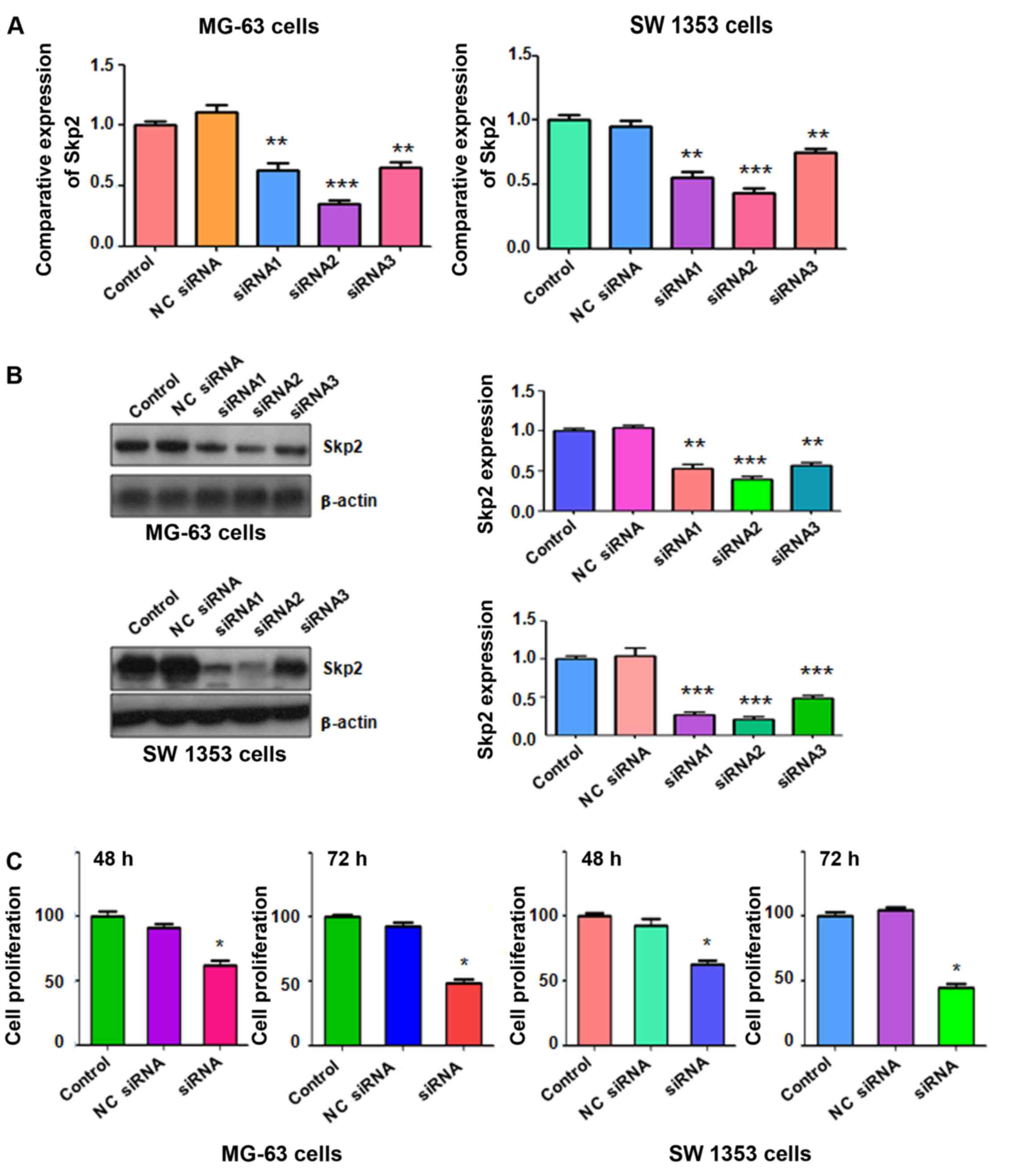
The results from RT-PCR revealed that Skp2 siRNAs significantly
downregulated Skp2 mRNA levels in both MG-63 and SW 1353 cells
(Fig. 1A). Consistently, our
western blot analysis demonstrated that Skp2 siRNAs markedly
inhibited the expression of Skp2 in both OS cell lines (Fig. 1B). In the following experiments,
Skp2 siRNA2 was used to deplete Skp2 expression in the two OS cell
lines.
Depletion of Skp2 inhibits cell
proliferation in OS cells
It has been demonstrated that downregulation of Skp2
suppressed cell proliferation in human cancer cells. To determine
whether Skp2 regulates cell proliferation in OS cells, an MTT assay
was performed to assess cell proliferation in OS cells transfected
with Skp2 siRNA. We found that depletion of Skp2 significantly
inhibited cell proliferation in MG-63 cells compared with the
control group (Fig. 1C). Similarly,
Skp2 siRNA treatment led to inhibition of cell proliferation in SW
1353 cells (Fig. 1C). Our findings
revealed that depletion of Skp2 suppressed cell proliferation in OS
cells.
Depletion of Skp2 triggers apoptosis
in OS cells
Next, we used an Annexin V-FITC/PI apoptosis
detection kit to assess apoptosis in OS cells after Skp2-siRNA
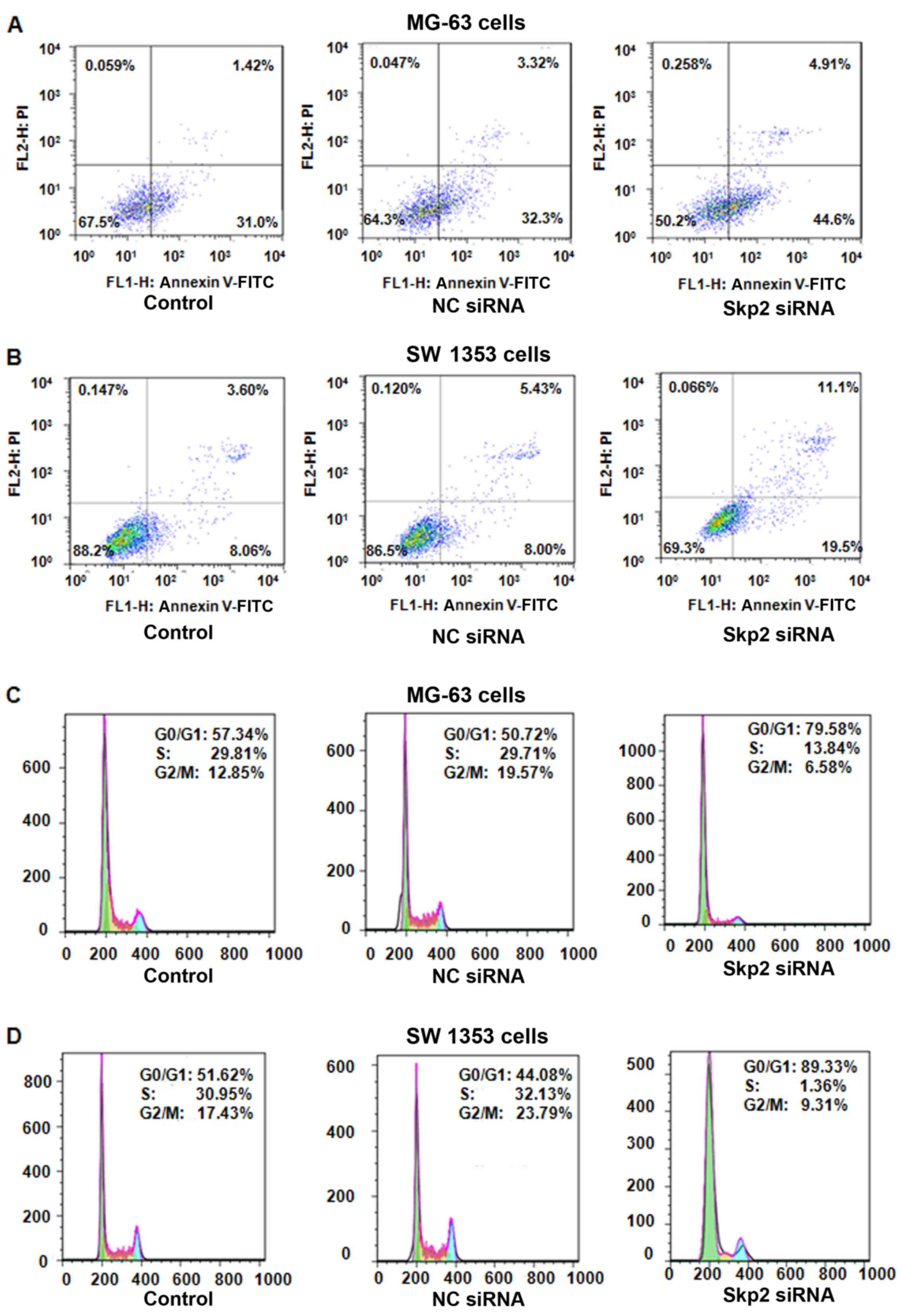
transfection. We found that depletion of Skp2 induced apoptosis in
both MG-63 and SW 1353 cells (Fig. 2A
and B). The percentage of apoptotic cells increased from 13.43%
in the control siRNA-treated group to 30.6% in the Skp2
siRNA-treated group in the SW 1353 cells (Fig. 2B). We also observed that
downregulation of Skp2 enhanced apoptosis in the MG-63 cells
(Fig. 2A). These data indicated
that depletion of Skp2 led to increased apoptosis, which
contributed to cell growth inhibition in both OS cell lines.
Depletion of Skp2 induces cell cycle
arrest in OS cells
To explore whether Skp2 regulates cell cycle
progression, PI staining and flow cytometry assays were conducted
in OS cells treated with Skp2 siRNA. We found a typical G0/G1
arrest pattern in both Skp2 siRNA-treated OS cell lines (Fig. 2C and D). The G0/G1 phase fraction
increased from 50.72% in the control siRNA-treated cells to 79.58%
in the Skp2 siRNA-treated MG-63 cells (Fig. 2C). Similarly, G0/G1 cell cycle
arrest was found in the Skp2 siRNA-treated SW 1353 cells (Fig. 2D). These results demonstrated that
depletion of Skp2 induced G0/G1 arrest in OS cells.
Depletion of Skp2 suppresses cell
migration and invasion in OS cells
To explore whether depletion of Skp2 suppresses the
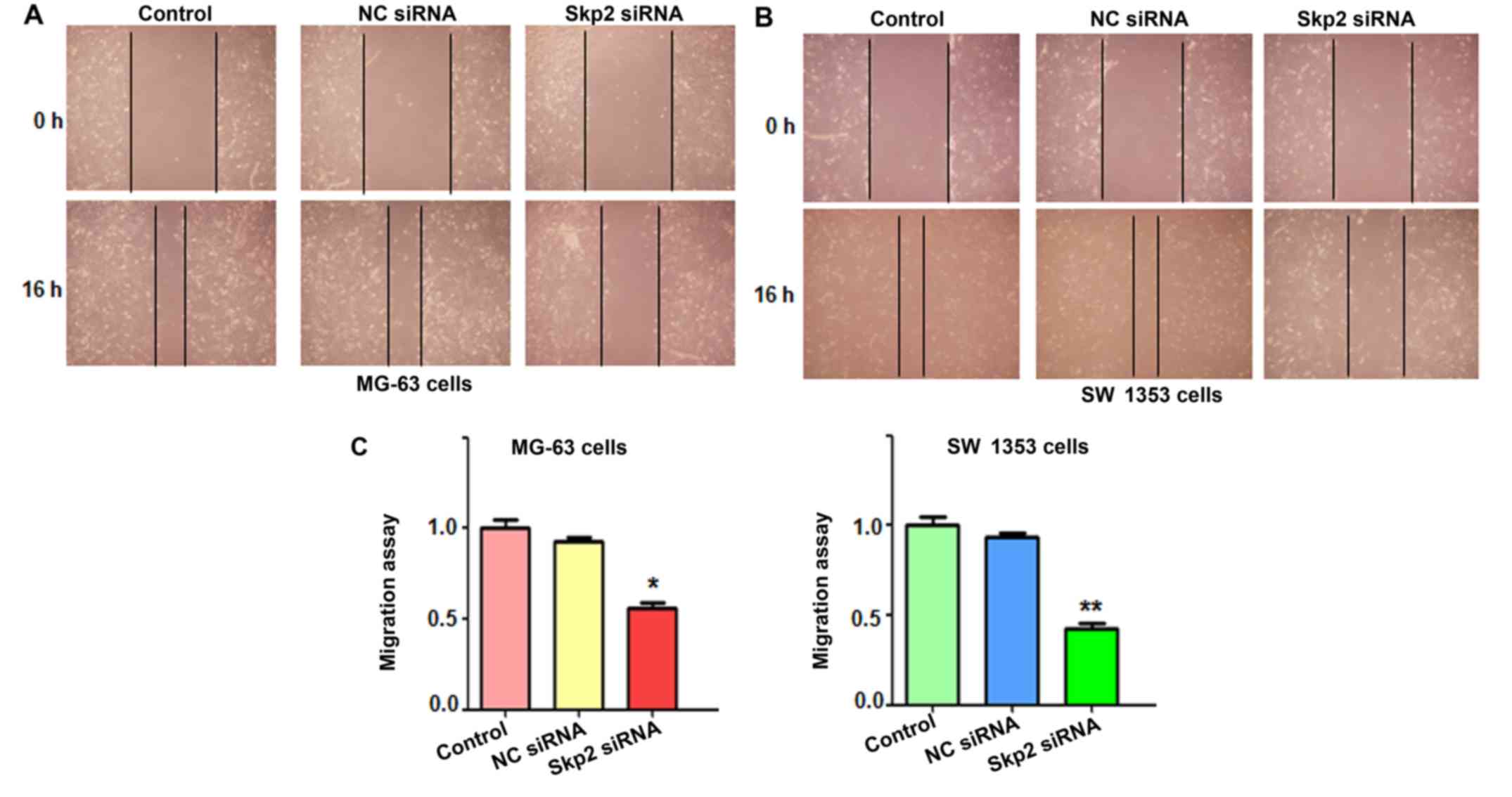
motility of OS cells, wound healing assays were conducted to assess
the migration of OS cells following Skp2-siRNA transfection. Our
results from the wound healing assays demonstrated that inhibition
of Skp2 significantly decreased cell migration in both OS cell
lines (Fig. 3). To further validate
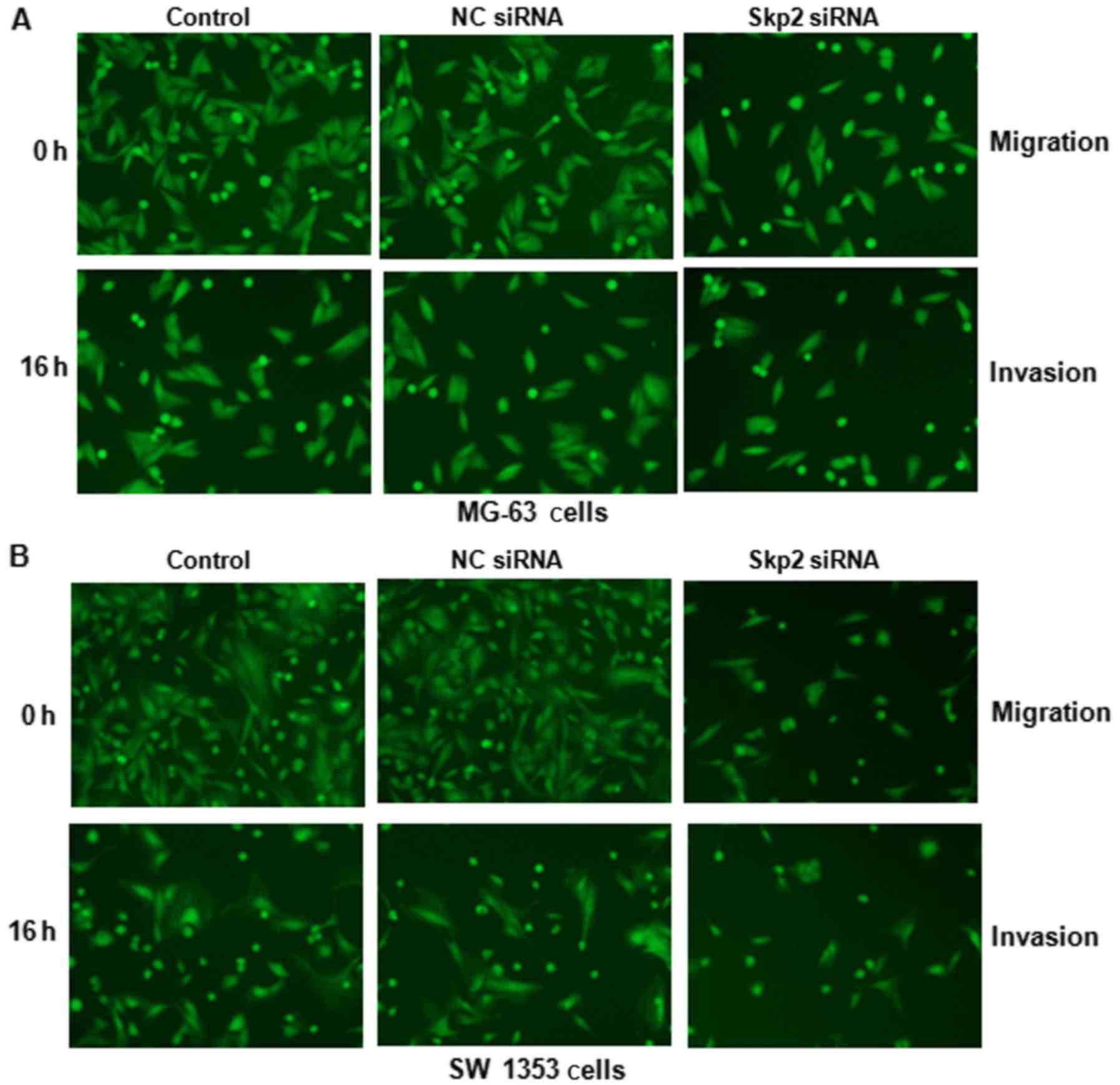
this finding, migration and invasion assays were performed to
determine the cell migratory and invasive activities of OS cells
treated with Skp2 siRNA using Transwell chamber assays. We observed
that downregulation of Skp2 markedly inhibited the migration and
invasion (Fig. 4) in both OS cell
lines. Altogether, depletion of Skp2 may inhibit motility in OS
cells.
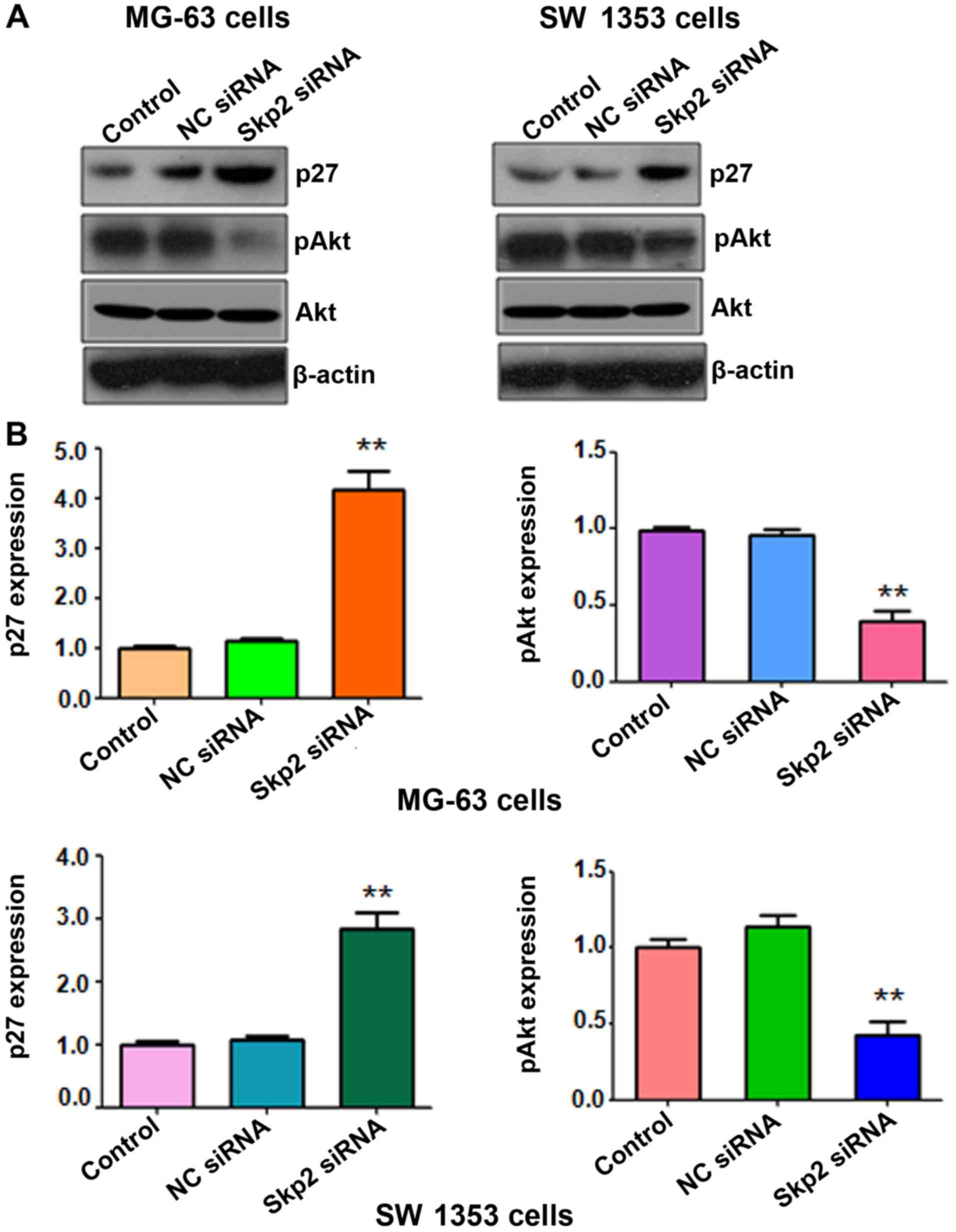
Depletion of Skp2 increases the
expression level of p27, but decreases the expression level of pAkt
in OS cells
It has been demonstrated that Skp2 regulates the
levels of p27 and pAkt, two key targets of Skp2, in several types
of human cancers. To further determine whether depletion of Skp2
regulates the expression of p27 and pAkt in OS cells, we assessed
the levels of these two genes in the Skp2 siRNA-treated OS cell
lines. Our western blot analysis data revealed that downregulation
of Skp2 increased p27 expression in both OS cell lines (Fig. 5). Furthermore, depletion of Skp2
decreased the expression of pAkt in OS cells, but not the total
level of Akt (Fig. 5). These
findings revealed that Skp2 suppressed the expression level of
pAkt, but increased the expression of p27 in OS cells.
Discussion
Various studies have discovered that Skp2 plays an
important role in tumorigenesis in a variety of human types of
cancer (40,41). However, the function of Skp2 in OS
remains largely obscure. Indirect evidence has indicated the
function of Skp2 in OS cells. For example, the depletion of
Forkhead box M1 (FoxM1) inhibited cell growth in human OS U2OS
cells due to dysreguation of Skp2 and Cks1, leading to a mitotic
block and accumulated levels of p21 and p27 (42). A previous study indicated that FoxM1
regulated the transcription of cell cycle genes including genes
encoding the SCF (Skp2-Cks1) ubiquitin ligase complex (42). Another study demonstrated that
inhibition of the Notch pathway by γ-secretase inhibitor suppressed
the growth of OS in vitro and in vivo due to a
decrease in the expression of accelerators of the cell cycle such
as Skp2, cyclin D1, and cyclin E1 and E2 (43). Similarly, inhibition of smoothened,
a key molecule in the Hedgehog pathway, slowed the growth of OS
cells via the suppression of Skp2 and cyclin D1 and E1, but
upregulation of p21 (44). In line
with this, knockdown of Gli2, a key mediator of the Hedgehog
pathway, prevented OS growth and anchorage-independent growth, and
promoted G1 phase arrest through inhibition of Skp2 and cyclin D1
(45). In the present study,
depletion of Skp2 in OS cells inhibited cell growth, arrested the
G1 cell cycle, triggered cell apoptosis, and suppressed cell
migration. This is direct evidence validating the biological
function of Skp2 in OS cells. Our findings demonstrated that
inactivation of Skp2 could be a useful approach for the treatment
of OS patients.
Accumulated evidence has revealed that Akt directly
binds to Skp2 and subsequently enhances the translocation of Skp2
from the nucleus to the cytoplasm, leading to activation of Skp2
function (46,47). Notably, one research group reported
that Akt could promote Skp2 translocation to the cytoplasm
(12,48). A recent study revealed that Skp2
stimulated Akt activation in human breast cancer cells (49). In line with this finding, our
results revealed that depletion of Skp2 inhibited the activation of
pAkt in OS cells. This result demonstrated that Skp2 exerted its
oncogenic function partly through the Akt pathway in OS cells.
Further detailed investigations are warranted to determine how Skp2
regulates Akt activation in OS cells.
Since inhibition of Skp2 represents an effective
therapeutic approach for patients with OS, it is critical to
discover and develop Skp2 inhibitors. To this end, several Skp2
inhibitors have been developed. For instance,
5-bromo-8-toluylsulfonamidoquinoline-1 inhibited the interaction of
Skp2 with cyclin-dependent kinase regulatory subunit 1 (Cks1),
leading to the inhibition of cell growth in lung cancer cells
(50). Another compound M1 targeted
the p300-binding site of Skp2 and disrupted the Skp2/p300
interaction, leading to apoptosis and cell death in cancer cells
(51). Arsenic trioxide exhibited
its antitumor activity via suppression of Skp2 in pancreatic cancer
cells (52). Caffeic acid phenethyl
ester suppressed cell proliferation and arrested the cell cycle
through inactivation of Skp2 in prostate cancer cells (53). Notably, multiple natural agents have
been characterized as Skp2 inhibitors (53–56).
Flavokawain A (FKA) selectively inhibited Skp2 in a
proteasome-dependent manner in prostate cancer (57). Quercetin, curcumin and lycopene
enhanced cell cycle arrest via the targeting of Skp2 expression in
breast cancer cells (54).
Moreover, curcumin was validated to regulate the
PI3K/Akt-SKP2-Cip/Kips pathway in breast cancer cells (55,56).
Furthermore, curcumin inhibited tumorigenesis via the
downregulation of Skp2 in glioma cells (58). 15,16-Dihydrotanshinone I (DHTI) has
been found to inhibit cell proliferation and migration, induce cell
apoptosis and G1 phase arrest partly through the suppression of
cell cycle regulators including Skp2 in OS cells (59). Li et al found that
saurolactam inhibited proliferation, migration and invasion of
human OS cells in part through the inactivation of Skp2 and pAkt
(60). Collectively, Skp2 may be a
potential therapeutic target for the treatment of OS.
References
|
1
|
Anderson ME: Update on survival in
osteosarcoma. Orthop Clin North Am. 47:283–292. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vos HI, Coenen MJ, Guchelaar HJ and Loo Te
DM: The role of pharmacogenetics in the treatment of osteosarcoma.
Drug Discov Today. 21:1775–1786. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kushlinskii NE, Fridman MV and Braga EA:
Molecular mechanisms and microRNAs in osteosarcoma pathogenesis.
Biochemistry. 81:315–328. 2016.PubMed/NCBI
|
|
5
|
Li C, Cong Y, Liu X, Zhou X, Zhou G, Lu M,
Shi X and Wu S: The progress of molecular diagnostics of
osteosarcoma. Front Biosci. 21:20–30. 2016. View Article : Google Scholar
|
|
6
|
Tao J, Jiang MM, Jiang L, Salvo JS, Zeng
HC, Dawson B, Bertin TK, Rao PH, Chen R, Donehower LA, et al: Notch
activation as a driver of osteogenic sarcoma. Cancer Cell.
26:390–401. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carrano AC, Eytan E, Hershko A and Pagano
M: SKP2 is required for ubiquitin-mediated degradation of the CDK
inhibitor p27. Nat Cell Biol. 1:193–199. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsvetkov LM, Yeh KH, Lee SJ, Sun H and
Zhang H: p27Kip1 ubiquitination and degradation is
regulated by the SCFSkp2 complex through phosphorylated
Thr187 in p27. Curr Biol. 9:661–664. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu ZK, Gervais JL and Zhang H: Human CUL-1
associates with the SKP1/SKP2 complex and regulates
p21CIP1/WAF1 and cyclin D proteins. Proc Natl Acad Sci
USA. 95:11324–11329. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kamura T, Hara T, Kotoshiba S, Yada M,
Ishida N, Imaki H, Hatakeyama S, Nakayama K and Nakayama KI:
Degradation of p57Kip2 mediated by
SCFSkp2-dependent ubiquitylation. Proc Natl Acad Sci
USA. 100:10231–10236. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang H, Regan KM, Wang F, Wang D, Smith
DI, van Deursen JM and Tindall DJ: Skp2 inhibits FOXO1 in tumor
suppression through ubiquitin-mediated degradation. Proc Natl Acad
Sci USA. 102:1649–1654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Cui J, Bauzon F and Zhu L: A
comparison between Skp2 and FOXO1 for their cytoplasmic
localization by Akt1. Cell Cycle. 9:1021–1022. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nakayama K, Nagahama H, Minamishima YA,
Matsumoto M, Nakamichi I, Kitagawa K, Shirane M, Tsunematsu R,
Tsukiyama T, Ishida N, et al: Targeted disruption of Skp2
results in accumulation of cyclin E and p27Kip1,
polyploidy and centrosome overduplication. EMBO J. 19:2069–2081.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kratzat S, Nikolova V, Miething C,
Hoellein A, Schoeffmann S, Gorka O, Pietschmann E, Illert AL,
Ruland J, Peschel C, et al: Cks1 is required for tumor cell
proliferation but not sufficient to induce hematopoietic
malignancies. PLoS One. 7:e374332012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Agarwal A, Bumm TG, Corbin AS, O'Hare T,
Loriaux M, VanDyke J, Willis SG, Deininger J, Nakayama KI, Druker
BJ, et al: Absence of SKP2 expression attenuates BCR-ABL-induced
myeloproliferative disease. Blood. 112:1960–1970. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakayama K, Nagahama H, Minamishima YA,
Miyake S, Ishida N, Hatakeyama S, Kitagawa M, Iemura S, Natsume T
and Nakayama KI: Skp2-mediated degradation of p27 regulates
progression into mitosis. Dev Cell. 6:661–672. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Minamishima YA and Nakayama K and Nakayama
K: Recovery of liver mass without proliferation of hepatocytes
after partial hepatectomy in Skp2-deficient mice. Cancer Res.
62:995–999. 2002.PubMed/NCBI
|
|
18
|
Chander H, Halpern M, Resnick-Silverman L,
Manfredi JJ and Germain D: Skp2B attenuates p53 function by
inhibiting prohibitin. EMBO Rep. 11:220–225. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shim EH, Johnson L, Noh HL, Kim YJ, Sun H,
Zeiss C and Zhang H: Expression of the F-box protein SKP2 induces
hyperplasia, dysplasia, and low-grade carcinoma in the mouse
prostate. Cancer Res. 63:1583–1588. 2003.PubMed/NCBI
|
|
20
|
Sistrunk C, Kim SH, Wang X, Lee SH, Kim Y,
Macias E and Rodriguez-Puebla ML: Skp2 deficiency inhibits
chemical skin tumorigenesis independent of p27Kip1
accumulation. Am J Pathol. 182:1854–1864. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pagano M: Control of DNA synthesis and
mitosis by the Skp2-p27-Cdk1/2 axis. Mol Cell. 14:414–416. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kullmann MK, Grubbauer C, Goetsch K, Jäkel
H, Podmirseg SR, Trockenbacher A, Ploner C, Cato AC, Weiss C,
Kofler R, et al: The p27-Skp2 axis mediates glucocorticoid-induced
cell cycle arrest in T-lymphoma cells. Cell Cycle. 12:2625–2635.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lim MS, Adamson A, Lin Z, Perez-Ordonez B,
Jordan RC, Tripp S, Perkins SL and Elenitoba-Johnson KS: Expression
of Skp2, a p27Kip1 ubiquitin ligase, in malignant
lymphoma: Correlation with p27Kip1 and proliferation
index. Blood. 100:2950–2956. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schüler S, Diersch S, Hamacher R, Schmid
RM, Saur D and Schneider G: SKP2 confers resistance of pancreatic
cancer cells towards TRAIL-induced apoptosis. Int J Oncol.
38:219–225. 2011.PubMed/NCBI
|
|
25
|
Chan CH, Lee SW, Wang J and Lin HK:
Regulation of Skp2 expression and activity and its role in cancer
progression. Sci World J. 10:1001–1015. 2010. View Article : Google Scholar
|
|
26
|
Hulit J, Lee RJ, Li Z, Wang C, Katiyar S,
Yang J, Quong AA, Wu K, Albanese C, Russell R, et al:
p27Kip1 repression of ErbB2-induced mammary tumor
growth in transgenic mice involves Skp2 and Wnt/beta-catenin
signaling. Cancer Res. 66:8529–8541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fujita T, Liu W, Doihara H, Date H and Wan
Y: Dissection of the APCCdh1-Skp2 cascade in breast
cancer. Clin Cancer Res. 14:1966–1975. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Voduc D, Nielsen TO, Cheang MC and Foulkes
WD: The combination of high cyclin E and Skp2 expression in breast
cancer is associated with a poor prognosis and the basal phenotype.
Hum Pathol. 39:1431–1437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu J, Wei XL, Huang WH, Chen CF, Bai JW
and Zhang GJ: Cytoplasmic Skp2 expression is associated with p-Akt1
and predicts poor prognosis in human breast carcinomas. PLoS One.
7:e526752012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei S, Chu PC, Chuang HC, Hung WC, Kulp SK
and Chen CS: Targeting the oncogenic E3 ligase Skp2 in prostate and
breast cancer cells with a novel energy restriction-mimetic agent.
PLoS One. 7:e472982012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao H, Bauzon F, Fu H, Lu Z, Cui J,
Nakayama K, Nakayama KI, Locker J and Zhu L: Skp2 deletion
unmasks a p27 safeguard that blocks tumorigenesis in the absence of
pRb and p53 tumor suppressors. Cancer Cell. 24:645–659. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Masuda TA, Inoue H, Sonoda H, Mine S,
Yoshikawa Y, Nakayama K, Nakayama K and Mori M: Clinical and
biological significance of S-phase kinase-associated protein
2 (Skp2) gene expression in gastric carcinoma:
Modulation of malignant phenotype by Skp2 overexpression, possibly
via p27 proteolysis. Cancer Res. 62:3819–3825. 2002.PubMed/NCBI
|
|
33
|
Benevenuto-de-Andrade BA, León JE, Carlos
R, Delgado-Azañero W, Mosqueda-Taylor A and Paes-de-Almeida O:
Immunohistochemical expression of Skp2 protein in oral nevi and
melanoma. Med Oral Patol Oral Cir Bucal. 18:e388–e391. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qu X, Shen L, Zheng Y, Cui Y, Feng Z, Liu
F and Liu J: A signal transduction pathway from TGF-β1 to SKP2 via
Akt1 and c-Myc and its correlation with progression in human
melanoma. J Invest Dermatol. 134:159–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen G, Cheng Y, Zhang Z, Martinka M and
Li G: Cytoplasmic Skp2 expression is increased in human melanoma
and correlated with patient survival. PLoS One. 6:e175782011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu M, Ma J, Xue W, Cheng C, Wang Y, Zhao
Y, Ke Q, Liu H, Liu Y, Li P, et al: The expression and prognosis of
FOXO3a and Skp2 in human hepatocellular carcinoma. Pathol Oncol
Res. 15:679–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu HM, Liang Y, Chen Q, Wu QN, Guo YM,
Shen GP, Zhang RH, He ZW, Zeng YX, Xie FY, et al: Correlation of
Skp2 overexpression to prognosis of patients with nasopharyngeal
carcinoma from South China. Chin J Cancer. 30:204–212. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fang FM, Chien CY, Li CF, Shiu WY, Chen CH
and Huang HY: Effect of S phase kinase-associated protein 2
expression on distant metastasis and survival in nasopharyngeal
carcinoma patients. Int J Radiat Oncol Biol Phys. 73:202–207. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liao QD, Zhong D and Chen Q: Protein
expression of Skp2 in osteosarcoma and its relation with prognosis.
Zhong Nan Da Xue Xue Bao Yi Xue Ban. 33:606–611. 2008.(In Chinese).
PubMed/NCBI
|
|
40
|
Xu D, Li CF, Zhang X, Gong Z, Chan CH, Lee
SW, Jin G, Rezaeian AH, Han F, Wang J, et al: Skp2-macroH2A1-CDK8
axis orchestrates G2/M transition and tumorigenesis. Nat Commun.
6:66412015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee SW, Li CF, Jin G, Cai Z, Han F, Chan
CH, Yang WL, Li BK, Rezaeian AH, Li HY, et al: Skp2-dependent
ubiquitination and activation of LKB1 is essential for cancer cell
survival under energy stress. Mol Cell. 57:1022–1033. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang IC, Chen YJ, Hughes D, Petrovic V,
Major ML, Park HJ, Tan Y, Ackerson T and Costa RH: Forkhead box M1
regulates the transcriptional network of genes essential for
mitotic progression and genes encoding the SCF (Skp2-Cks1)
ubiquitin ligase. Mol Cell Biol. 25:10875–10894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tanaka M, Setoguchi T, Hirotsu M, Gao H,
Sasaki H, Matsunoshita Y and Komiya S: Inhibition of Notch pathway
prevents osteosarcoma growth by cell cycle regulation. Br J Cancer.
100:1957–1965. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hirotsu M, Setoguchi T, Sasaki H,
Matsunoshita Y, Gao H, Nagao H, Kunigou O and Komiya S: Smoothened
as a new therapeutic target for human osteosarcoma. Mol Cancer.
9:52010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nagao H, Ijiri K, Hirotsu M, Ishidou Y,
Yamamoto T, Nagano S, Takizawa T, Nakashima K, Komiya S and
Setoguchi T: Role of GLI2 in the growth of human osteosarcoma. J
Pathol. 224:169–179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gao D, Inuzuka H, Tseng A, Chin RY, Toker
A and Wei W: Phosphorylation by Akt1 promotes cytoplasmic
localization of Skp2 and impairs APCCdh1-mediated Skp2 destruction.
Nat Cell Biol. 11:397–408. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin HK, Wang G, Chen Z, Teruya-Feldstein
J, Liu Y, Chan CH, Yang WL, Erdjument-Bromage H, Nakayama KI, Nimer
S, et al: Phosphorylation-dependent regulation of cytosolic
localization and oncogenic function of Skp2 by Akt/PKB. Nat Cell
Biol. 11:420–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang H: Skip the nucleus, AKT drives Skp2
and FOXO1 to the same place? Cell Cycle. 9:868–869. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chan CH, Li CF, Yang WL, Gao Y, Lee SW,
Feng Z, Huang HY, Tsai KK, Flores LG, Shao Y, et al: The Skp2-SCF
E3 ligase regulates Akt ubiquitination, glycolysis, herceptin
sensitivity, and tumorigenesis. Cell. 149:1098–1111. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Singh R, Sran A, Carroll DC, Huang J,
Tsvetkov L, Zhou X, Sheung J, McLaughlin J, Issakani SD, Payan DG,
et al: Developing structure-activity relationships from an HTS hit
for inhibition of the Cks1-Skp2 protein-protein interaction. Bioorg
Med Chem Lett. 25:5199–5202. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Oh M, Lee JH, Moon H, Hyun YJ and Lim HS:
A chemical inhibitor of the Skp2/p300 interaction that promotes
p53-mediated apoptosis. Angew Chem Int Ed Engl. 55:602–606. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gao JK, Wang LX, Long B, Ye XT, Su JN, Yin
XY, Zhou XX and Wang ZW: Arsenic trioxide inhibits cell growth and
invasion via down-regulation of Skp2 in pancreatic cancer cells.
Asian Pac J Cancer Prev. 16:3805–3810. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lin HP, Lin CY, Huo C, Hsiao PH, Su LC,
Jiang SS, Chan TM, Chang CH, Chen LT, Kung HJ, et al: Caffeic acid
phenethyl ester induced cell cycle arrest and growth inhibition in
androgen-independent prostate cancer cells via regulation of Skp2,
p53, p21Cip1 and p27Kip1. Oncotarget. 6:6684–6707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Huang HC, Lin CL and Lin JK:
1,2,3,4,6-penta-O-galloyl-β-D-glucose, quercetin,
curcumin and lycopene induce cell-cycle arrest in MDA-MB-231 and
BT474 cells through downregulation of Skp2 protein. J Agric Food
Chem. 59:6765–6775. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Jia T, Zhang L, Duan Y, Zhang M, Wang G,
Zhang J and Zhao Z: The differential susceptibilities of MCF-7 and
MDA-MB-231 cells to the cytotoxic effects of curcumin are
associated with the PI3K/Akt-SKP2-Cip/Kips pathway. Cancer Cell
Int. 14:1262014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sun SH, Huang HC, Huang C and Lin JK:
Cycle arrest and apoptosis in MDA-MB-231/Her2 cells induced by
curcumin. Eur J Pharmacol. 690:22–30. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li X, Yokoyama NN, Zhang S, Ding L, Liu
HM, Lilly MB, Mercola D and Zi X: Flavokawain A induces
deNEDDylation and Skp2 degradation leading to inhibition of
tumorigenesis and cancer progression in the TRAMP transgenic mouse
model. Oncotarget. 6:41809–41824. 2015.PubMed/NCBI
|
|
58
|
Wang L, Ye X, Cai X, Su J, Ma R, Yin X,
Zhou X, Li H and Wang Z: Curcumin suppresses cell growth and
invasion and induces apoptosis by down-regulation of Skp2 pathway
in glioma cells. Oncotarget. 6:18027–18037. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen X, Li Q, He Y, Du H, Zhan Z, Zhao H,
Shi J, Ye Q and Hu J: 15,16-dihydrotanshinone I induces apoptosis
and inhibits the proliferation, migration of human osteosarcoma
cell line 143B in vitro. Anticancer Agents Med Chem. 15:12015.
View Article : Google Scholar
|
|
60
|
Li Z, Liu H, Li B, Zhang Y and Piao C:
Saurolactam inhibits proliferation, migration, and invasion of
human osteosarcoma cells. Cell Biochem Biophys. 72:719–726. 2015.
View Article : Google Scholar : PubMed/NCBI
|