Introduction
Cancer of the brain and the central nervous system
is one of the 10 most common types of cancer in China, and its
incidence rate has evidently increased from 2000 to 2011 (1). Glioma is the most common malignant
primary brain tumor, and accounts for the vast majority of all
intracranial tumors (2). At
present, standard therapeutic options include maximal safe
resection, followed by radiation therapy and adjuvant chemotherapy,
which are not particularly effective (3). Despite the improvement of treatment
modalities, the majority of patients with malignant glioma may have
poor prognosis within 2 years of diagnosis (4).
Structure-specific recognition protein 1 (SSRP1) is
a subunit of the facilitates chromatin transcription (FACT)
complex, which attaches to the nucleosome and reassembles the
nucleosome (5). SSRP1 was initially
identified in 1991 as a high-mobility group protein 1
(HMG1)-related DNA binding protein, and the capacity of SSRP1 for
biological functions can be attributed to its HMG domain (6). As an important histone chaperone, it
plays a role as a transcription factor in the regulation of several
targets to modulate cellular processes such as DNA replication, DNA
damage repair, apoptosis and cell cycle regulation (7–11). It
has been documented that the expression of SSRP1 was associated
with the stage of cellular differentiation. High SSRP1 levels were
observed in stem or less-differentiated cells, while low SSRP1
levels were observed in more differentiated cells (12). The high expression of SSRP1 was
detected in multiple human types of cancer, including
non-small-cell lung cancer, renal cell carcinoma, pancreatic ductal
and colorectal adenocarcinoma, and hepatocellular carcinoma
(13–15). In addition, it is a potential marker
and target of aggressive types of cancer (13,16,17).
The mitogen-activated protein kinase (MAPK)
signaling pathway is a highly conserved module that regulates
various cellular functions, including proliferation,
differentiation and malignant transformation (18). The MAPK pathway has been reported to
be activated in over 88% of gliomas (19). The suppression of MAPK signaling
synergizes the cytotoxicity of receptor tyrosine kinase inhibitors
in glioma tumor-initiating cells (20). Research on the function of SSRP1 has
focused on directing nucleosome reorganization as the histone
chaperone, while the expression and potential molecular mechanism
of SSRP1 in glioma remains unknown.
In the present study, we evaluated the expression of
SSRP1 in human patient samples and investigated its function in the
progression of glioma. In addition, we explored the effect of SSRP1
silencing on glioma cell proliferation and invasion. The results
revealed that SSRP1 may function as a regulator, and promote cell
proliferation and invasion by enhancing the activity of the MAPK
signaling pathway. Therefore, the present study revealed that SSRP1
played a role in the progression of glioma, and also provided
valuable information for understanding the mechanism of glioma.
Materials and methods
Tissue samples
Human glioma tissues were obtained during surgery at
the Department of Neurosurgery, Renmin Hospital of Wuhan University
(Wuhan, China), from 2010 to 2016. These control non-glioma human
brain tissues were collected from unmatched patients undergoing
surgery for intracranial hypertension. A total of 83
paraffin-embedded tissue samples, which included 77 glioma tissues,
and 6 normal brain (NB) tissues, were histopathologically diagnosed
by two neuropathologists. For real-time quantitative reverse
transcription-polymerase chain reaction (RT-PCR), 30 frozen (stored
in liquid nitrogen) glioblastoma and 10 NB tissues were evaluated.
All tissues were classified according to the 2016 World Health
Organization (WHO) classification of tumors of the central nervous
system. Prior patient consent and approval from the Ethics
Committee of Wuhan University were obtained for the use of these
clinical materials. The authors assert that all experiments
complied with the ethical standards of relevant national and
institutional guidelines as well as the laws of the People's
Republic of China. The detailed demographics of patients are
presented in Table I.
 | Table I.Correlation between the
clinicopathological characteristics and expression of SSRP1 protein
in glioma. |
Table I.
Correlation between the
clinicopathological characteristics and expression of SSRP1 protein
in glioma.
|
|
| SSRP1 expression |
|
|---|
|
|
|
|
|
|---|
| Characteristics | No. of patients | High | Low | P-value |
|---|
| Age (years) |
|
|
|
|
| ≥50 | 42 | 28 | 14 | 0.223 |
|
<50 | 35 | 21 | 14 |
|
| Sex |
|
|
|
|
| Male | 39 | 27 | 12 | 0.535 |
|
Female | 38 | 22 | 16 |
|
| WHO grade |
|
|
|
|
|
I+II | 11 | 3 | 8 | 0.009 |
|
III+IV | 66 | 46 | 20 |
|
Cells and cell culture
Human glioblastoma-derived cancer cell lines U118
and U251 were purchased from the Cell Bank Type Culture Collection
of the Chinese Academy of Sciences (Shanghai, China). The American
Type Culture Collection (ATCC; Manassas, VA, USA) suspects that
U118 is a contaminated cell line which has similar cytogenetics and
similar origin with glioblastoma cell line U138MG, since there is
no doubt that U118 is a cell line derived from malignant gliomas,
we thus used U118 as experimental cells. These cells were cultured
in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) (both from Gibco Grand Island, NY, USA),
100 U/ml of penicillin and 100 µg/ml of streptomycin
(Sigma-Aldrich, St. Louis, MO, USA) at 37°C with 5%
CO2.
Gene expression profiles
SSRP1 expression datasets were obtained from the
Gene Expression Omnibus (GEO) (http://www.ncbi.nlm.nih.gov/geo/). Three datasets were
retrieved for the study: GSE50161 (13 NB, 15 pilocytic astrocytoma,
46 ependymoma, 22 medulloblastoma and 34 glioblastoma tissues were
collected for microarray analysis), GSE4290 (23 NB and 157 glioma
tissues were analyzed; glioma tissues comprised of 45 grade II, 31
grade III and 81 grade IV cases), and GSE3185 (10 astrocytomas and
10 glioblastoma tissues were analyzed). The expression values of
SSRP1 were transformed to relative expression.
Immunohistochemical staining
The paraffin-embedded tissues were placed in xylene
3 times for 15 min at room temperature, hydrated in a series of
100, 95, 90, 80, 70 and 60% ethanol solutions, and washed in
phosphate-buffered saline (PBS). The sections were treated with 3%
H2O2 and subjected to antigen retrieval by
citrate buffer (10 mmol/l, pH 6.0) for 15 min. Then, the sections
were incubated with primary rabbit anti-SSRP1 polyclonal antibody
(1:200; GeneTex, Inc., Irvine, CA,. USA) overnight at 4°C. After
washing with PBS, the slides were incubated with poly-HRP goat
anti-rabbit antibody (Maixin Bio, Fujian, China) for 30 min, and
incubated with diaminobenzidine for 5 min. Next, the sections were
counterstained with hematoxylin, dehydrated in ethanol and
coverslips were placed on the slides. Images were captured using an
Olympus BX40 microscope and the CC-12 Soft Imaging System (Olympus,
Tokyo, Japan).
Immunohistochemical evaluation
SSRP1-positive cells displayed brownish yellow
granules on the cytoplasm. These results were evaluated through
immunohistochemical scores (IHC scores), according to the intensity
of the staining and the percentage of immunoreactive cells. The
estimate of the staining intensity was scored as follows: 0, no
staining; 1, weak staining; 2, moderate staining; and 3, strong
staining. The percentage was rated on a scale of 0–4, as follows:
0, <5%; 1, 5–25%; 2, 26–50%; 3, 51–75%; and 4, 76–100%. The IHC
score was obtained by multiplying the percentage and intensity
score. These IHC scores ranged from 0 to 12, in which scores 0–4
were considered as low expression, while scores 5–12 were
considered as high expression. These scores were independently
determined by two independent senior pathologists.
Western blot analysis
Cells were lysed in ice-cold RIPA buffer (50 mM of
Tris-HCl pH 7.4, 150 mM of NaCl, 1% Triton X-100, 0.25%
deoxycholate, 1.5 mM of MgCl2, 1 mM of EGTA, 1 mM of
phenylmethylsulfonyl fluoride, 10 mM of ZnAF, 10 mM of pervanadate,
10 µg/ml of leupeptin and 10 µg/ml of aprotinin) and incubated for
30 min. The protein concentration was determined using the BCA
method. The cell lysate was heated at 100°C for 10 min after being
mixed with sample loading buffer. The protein samples were equally
loaded on 10% SDS-PAGE, and transferred onto nitrocellulose
membranes. After being blocked with 5% non-fat milk in Tris-buffer,
the membrane strips were incubated with a primary antibody
overnight at 4°C, followed by Alex Fluor 680/790-labeled goat
anti-rabbit or goat anti-mouse IgG (Li-COR Biosciences, Lincoln,
NE, USA). The strips were visualized using the LI-COR Odyssey
Infrared Imaging System (Li-COR Biosciences).
The primary antibodies were as follows: p38,
phospho-p38, ERK, phospho-ERK, JNK and phospho-JNK (1:1,000; Cell
Signaling Technology, Danvers, MA, USA); MMP2, VEGF, EGFR, cyclin D
and E (1:1,000; Abcam, Cambridge, MA, USA); p65, Bcl2, Snail,
c-MYC, SSRP1 and GAPDH (1:1,000; GeneTex, Inc.).
RNA isolation and RT-PCR
Total RNA was isolated from U118 and U251 cell
lines, and extracted from NB and glioma tissues with TRIzol
reagents (Invitrogen, Carlsbad, CA, USA). Then, cDNA was prepared
from 2 µg of total RNA using the PrimeScript RT reagent kit with
gDNA Eraser (Takara, Tokyo, Japan) according to the manufacturer's
instructions. The primers were as follows: SSRP1 forward,
5′-GGATTGAAAGAGGGCATGAA-3′ and reverse, 5′-AGAGGCGTTGCTGTCAAACT-3′;
and GAPDH forward, 5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse,
5′-GCCATCACGCCACAGTTTC-3′. RT-PCR was performed to quantify the
mRNA expression using the SYBR-Green PCR Master Mix (Takara). GAPDH
was used for SSRP1 normalization.
Transient transfection with
siRNAs
Small interfering RNAs (siRNAs) were designed and
synthesized by Suzhou GenePharma, Inc. (Shanghai, China). Two
siRNAs targeting the SSRP1 gene were designed and purchased, and
the most effective siRNAs were identified using both RT-PCR and
western blotting. The sequences of the siRNAs against SSRP1 were as
follows: siRNA1 sense, 5′-GCCAUGUCUACAAGUAUGATT-3′ and antisense,
5′-UCAUACUUGUAGACAUGGCTT-3′; siRNA2 sense,
5′-CCCAGAAUGGUGUUGUCAAATT-3′ and antisense,
5′-UUUGACAACACAUUCUGGGTT-3′. The sequence of the negative control
siRNA was as follows: sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
antisense, 5′-ACGUGACACGUUCGGAGAATT-3′. The siRNA sequences were
derived from another research group at our laboratory, and the
specificity had been verified (14). Glioma cells were plated onto a 6- or
a 96-well plate at 40–50% confluency. After 24 h, siRNA
transfections were conducted using HyperFect (Qiagen, Hilden,
Germany), according to the manufacturer's protocol. Cells were
harvested after 48–72 h. All siRNAs were used at a final
concentration of 20 nM.
Cell proliferation assay
Cell proliferation was analyzed using Cell Counting
Kit-8 (CCK-8; Dojindo, Tokyo, Japan). Cells were seeded in 96-well
plates on day 0, and the cell growth was assessed at days 1, 2 and
3 after culturing, according to the manufacturer's instructions. On
average, 5 replicates for each time-point were statistically
analyzed.
5-Ethynyl-2′-deoxyuridine (EdU)
incorporation assay
Cells were seeded into 96-well plates containing
complete medium, and were allowed to attach overnight. Then, the
cell growth was determined using the CellLight Edu imaging
detecting kit (RiboBio, Guangzhou, China) according to the
manufacturer's protocol. In brief, the cells were cultured in
medium with 10 µM of EdU for 24 h, and fixed with 4%
paraformaldehyde for 20 min. Subsequently, the cells were incubated
in an Apollo reaction cocktail for 30 min, and the cell nuclei were
stained with 5 µg/ml of Hoechst 33342 for 30 min. The cells were
visualized under a fluorescence microscope (Olympus BX51; Olympus,
Tokyo, Japan). Experiments were performed in triplicate.
Cell cycle analysis
Cells were harvested with 0.25% trypsin and washed
twice with ice-cold PBS. After centrifugation, the cells were
suspended with 75% methanol overnight at −20°C. Then, the cells
were washed twice with PBS, incubated in PBS with 1 mg/ml of RNAase
at 37°C, and stained with 50 mg/ml of propidium iodide (PI) in PBS.
For each experiment, 2.5×104 cells were analyzed using
BD FACSAria (BD Biosciences, Franklin Lakes, NJ, USA). Six
replicates for each time-point were statistically analyzed.
Apoptosis analysis
Cells in each group were plated in 6-well plates,
and harvested at 48 h after transient transfection with siRNA.
Then, the cells were stained with ApoScreen Annexin V and PI,
according to the manufacturer's instructions (BD Biosciences). For
each experiment, 2.5×104 cells were analyzed using BD
FACSAria (BD Biosciences). Six replicates for each time-point were
statistically analyzed.
Cell migration and invasion
assays
The cell migration assay was conducted using 8-µm
pore size Transwell chambers (Corning, Corning, NY, USA). Cells in
each group were suspended in serum-free DMEM. Then,
5×104 cells in 100 µl of DMEM were plated into the upper
chamber, and 600 µl of DMEM containing 10% FBS was added to the
lower chamber. These chambers were cultured at 37°C with 5%
CO2 for 24 h. The cells in the upper chamber of the
insert were collected using a cotton swab. Cells that had adhered
to the lower surface were fixed with 4% paraformaldehyde, stained
with 0.1% crystal violet and counted under a microscope (Olympus
BX51). The invasion assay was the same, except that the Transwell
chambers were precoated with Matrigel (R&D Systems,
Minneapolis, MN, USA), and the cells were cultured for 36 h.
Statistical analysis
All statistical analyses are presented as the
average of at least triplicate samples and as the mean ± standard
deviation. SPSS 19.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad
Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA, USA)
were used for statistical analysis. A P-value <0.05 was
considered statistically significant.
Results
SSRP1 mRNA and protein are
overexpressed in glioma tissues
In order to observe the mRNA expression level of
SSRP1 in glioma, 3 independent GEO datasets (GSE50161, GSE4290 and
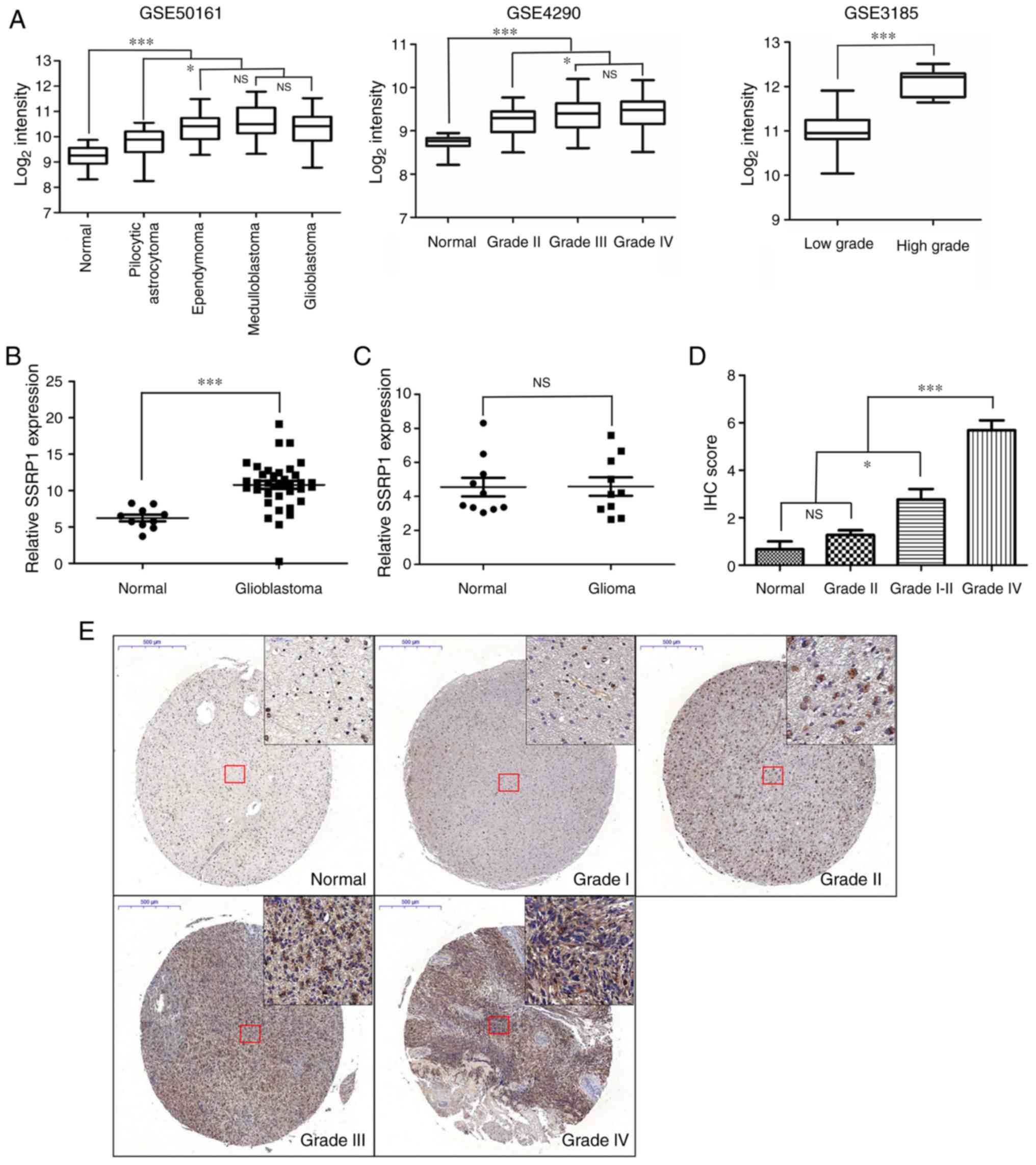
GSE3185) were analyzed. As shown in Fig. 1A, the expression of SSRP1 was
upregulated in tumor tissues, compared to NB tissues; and the
expression levels of SSRP1 were significantly higher in the
glioblastoma multiform group, compared with the low-grade glioma
group. The mRNA expression of SSRP1 in 10 NB and 30 glioblastoma
tissues were examined by RT-PCR. Compared with NB tissues,
glioblastoma tissues exhibited higher SSRP1 mRNA expression levels
(P<0.001; Fig. 1B). Next, we
attempted to elucidate the clinicopathological significance of the
mRNA expression of SSRP1 in the peripheral blood of glioma
patients, and found that the difference between glioma patients and
normal controls was not statistically significant (P=0.97; Fig. 1C). In addition, the protein
expression levels of SSRP1 were assessed in an independent cohort
of 83 cases, which included 6 NB cases, 3 grade I, 8 grade II, 22
grade III and 44 grade IV cases, using immunohistochemical
staining. The staining density of SSRP1 in the high-grade group
exhibited stronger coloring and broader distribution, compared to
that in the low-grade group (Fig.
1E). As shown in Table I, among
the 11 cases of glioma tissues at grades I–II, 3 (27.3%) cases
revealed positive staining, while 11 out of 22 (50.0%) cases of
glioma tissues at grade III and 35 out of 44 (79.5%) cases of
glioma tissues at grade IV were detected with positive staining of
SSRP1 via immunohistochemical analysis. Quantitative analysis also
revealed that the average score of SSRP1 staining increased from
histologic grade I to grade IV (P<0.05; Fig. 1D). However, it was found that there
was no significant association between the expression of SSRP1 and
the age and sex of these patients (Table I).
Downregulated SSRP1 expression
suppresses cell proliferation in vitro
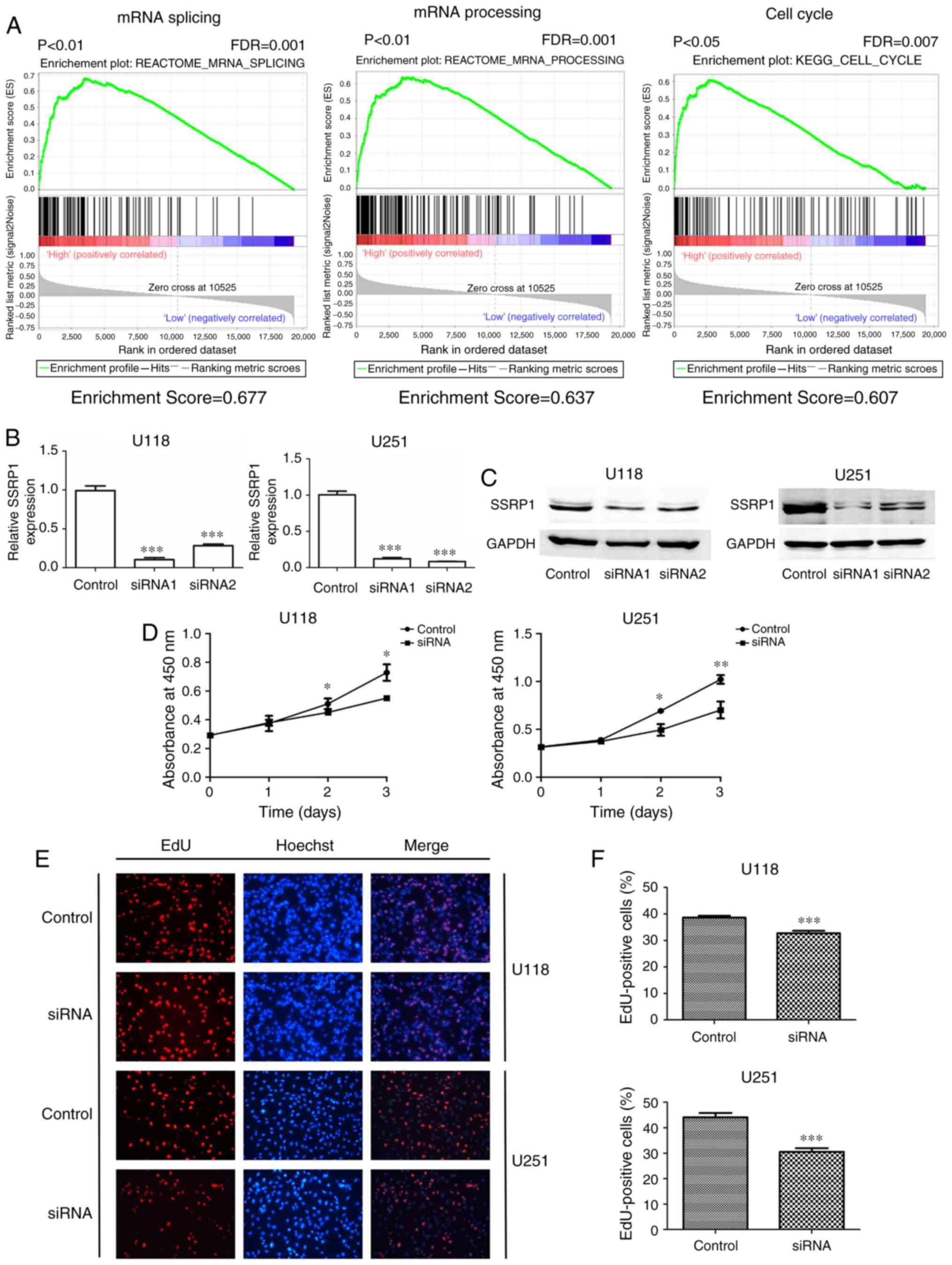
Gene set enrichment analysis (GSEA) was performed to
identify biological processes potentially modulated by SSRP1. As
shown in Fig. 2A, the GSEA results
from gene profiling data (GSE50161) demonstrated that a mass of
gene sets were enriched by SSRP1, and the enriched expression of
gene sets included mRNA splicing, mRNA processing and the cell
cycle (Fig. 2A).
A plasmid pcDNA3.1-SSRP1 was constructed to increase
the expression of SSRP1 in glioma cells. U251 cells transfected
with pcDNA3.1-SSRP1 plasmid (U251-SSRP1) has a higher mRNA
expression level, while the protein level was only 1.20-fold. The
average percentage of EdU-positive U251-SSRP1 cells was similar
with that in the U251 cells transfected with empty vector (38.30
and 39.51%; P=0.502). Flow cytometric analysis revealed that the
average percentage of U251-SSRP1 cells in the S + G2/M phase was
52.17%, which had no significant differences with the control group
(P=0.102). Overexpression of SSRP1 could not exert its biological
effects in U251, and the reason may be attributed to the high
expression of exogenous expression of SSRP1. In addition, further
increased expression of SSRP1 was at variance with objective
reality. Thus, we did not perform a functional study of the
overexpression SSRP1.
siRNA was used to specifically knockdown the
expression of SSRP1 in U118 and U251 cell lines, which were
established from high-grade glioma tumors. The efficient knockdown
of SSRP1 expression was assessed through both RT-PCR and western
blotting, and siRNA1 revealed more interference efficiency
(P<0.001; Fig. 2B and C).
Therefore, siRNA1 was selected to knockdown the expression of SSRP1
in U118 and U251.
Subsequently, we verified the effect of the
downregulated expression of SSRP1 in glioma cell growth in
vitro. A CCK-8 assay was used to evaluate U118 and U251 cell
growth at 24, 48 and 72 h after siRNA transfection. The results
revealed that the cell growth was significantly decreased in cells
transfected with si-SSRP1 compared with the cells transfected with
scramble siRNA (negative control) (P<0.05; Fig. 2D). In addition, cell proliferation
was also assessed using the EdU incorporation assays. As shown in
Fig. 2E and F, the average
percentage of EdU-positive cells in the control group and the SSRP1
knockdown group were 38.60 and 32.73% in the U118 cells, and 44.1
and 30.6% in the U251 cells (P<0.001). The knockdown of SSRP1
expression led to a marked decrease in the percentage of
EdU-positive cells, as compared with the control group
(P<0.001).
All the aforementioned results revealed that the
downregulation of SSRP1 suppressed the proliferation of glioma
cells in vitro.
siRNA-mediated knockdown of SSRP1
arrests the cell cycle and causes apoptosis in glioma cells
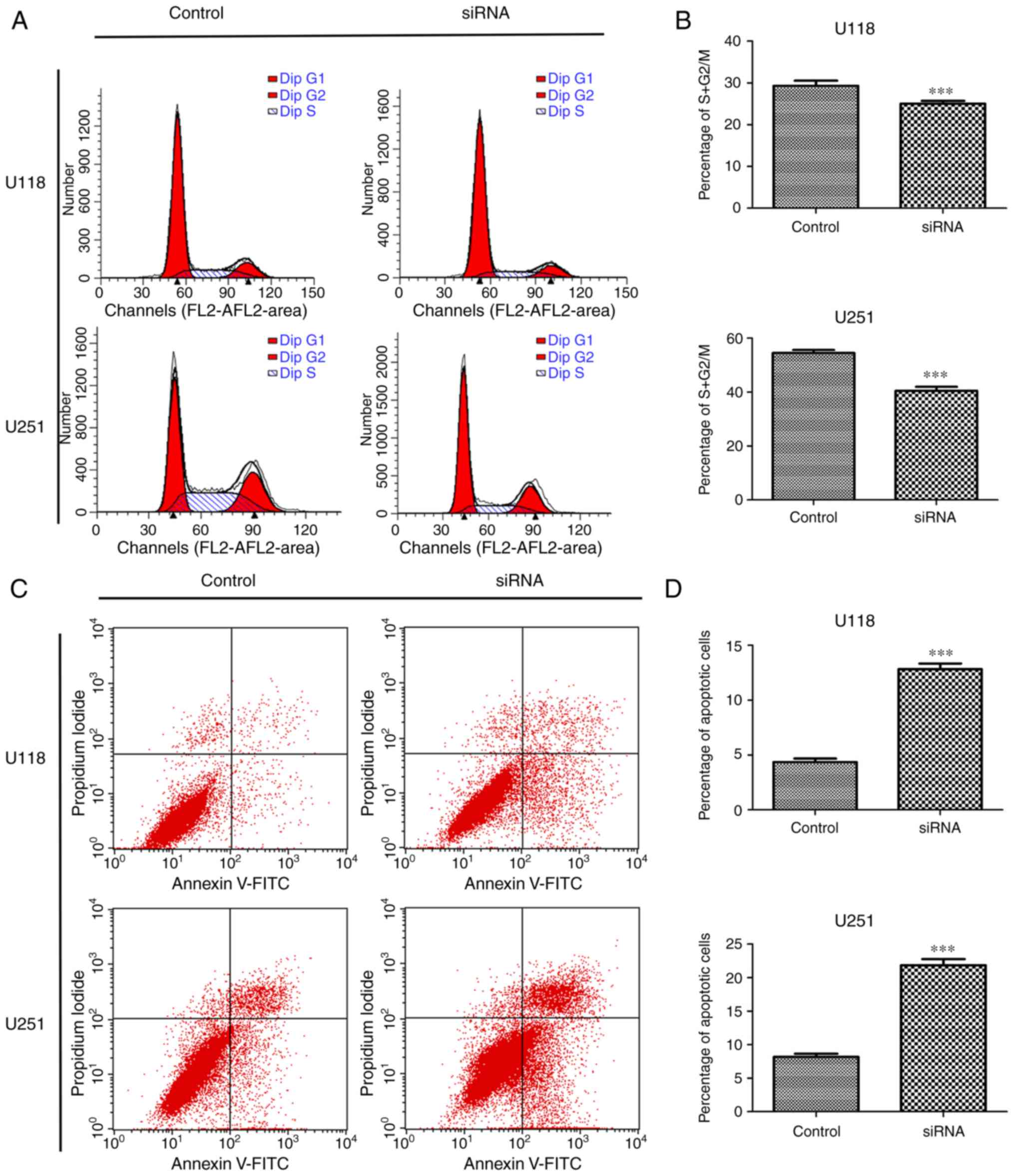
In order to explore whether the effect of SSRP1
depletion on cell proliferation was related to the cell cycle and
apoptosis, flow cytometric analysis was first performed. Cells
transfected with si-SSRP1 exhibited a significant decrease in the
percentage of cells in the S + G2/M phase compared with the cells
transfected with scramble siRNA in the U118 (29.31 vs. 24.05%) and
U251 cell lines (54.53 vs. 40.47%) (Fig. 3A and B). Moreover, the function of
SSRP1 in apoptosis was also investigated. After transfection with
si-SSRP1 or scramble siRNA for 48 h, the average percentage of
apoptotic U118 cells treated with si-SSRP1 and the control group
was 4.36 and 12.83%, respectively (P<0.001; Fig. 3C and D). Experiments using U251
yielded similar results, and the average percentage of apoptotic
cells was significantly increased after treatment with si-SSRP1.
The percentage of apoptotic U251 cells in the two groups was 8.18
and 21.83% (P<0.001; Fig. 3C and
D). These results revealed that SSRP1 was involved in the cell
cycle process and apoptosis of glioma cells.
Decrease of SSRP1 by siRNA inhibits
glioma cell migration and invasion
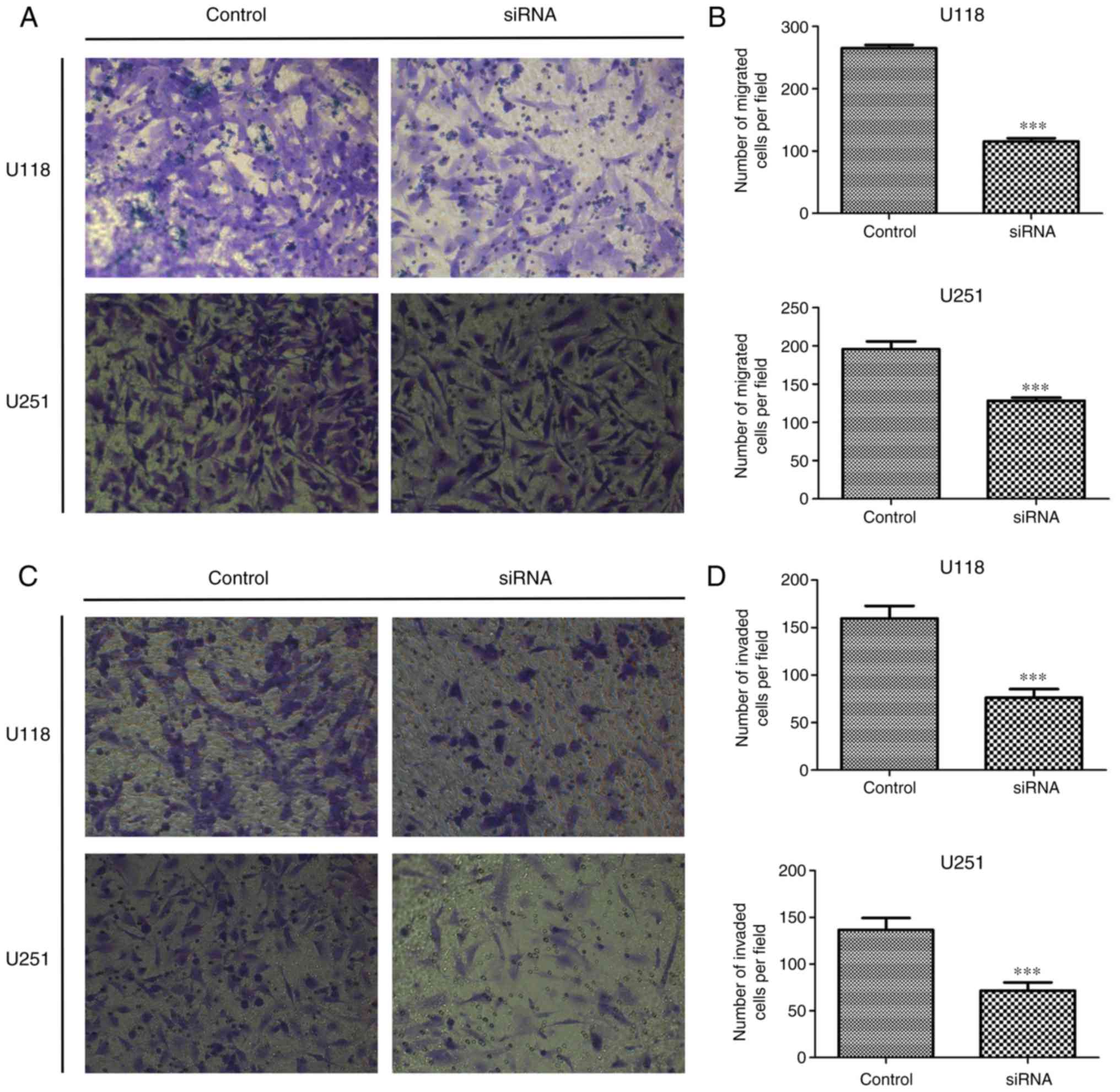
In order to investigate the role of SSRP1 in glioma
metastasis, we examined whether SSRP1 played a role in cell
migration and invasion using the Transwell apparatus. After 24 h of
incubation, the number of migrated cells in both the U118 and U251
cells transfected with si-SSRP1 was significantly lower than that
in the negative control group (P<0.001, for each; Fig. 4A and B). The Transwell matrix
penetration assay was performed to evaluate the effects of SSRP1 on
cell invasion. Compared with the control group, the cells with
downregulated SSRP1 expression by siRNA demonstrated a
significantly decreased invasion in both U118 and U251 cell lines
(P<0.001, for both; Fig. 4C and
D). All these results demonstrated that SSRP1 modulates glioma
cell migration and invasion in vitro.
SSRP1 controls the expression of
proliferation-and migration-associated genes in glioma
In order to explore the mechanism by which SSRP1
modulated the progression of cell proliferation and metastasis, the
proteins involved in the cell cycle, migration and invasion in U118
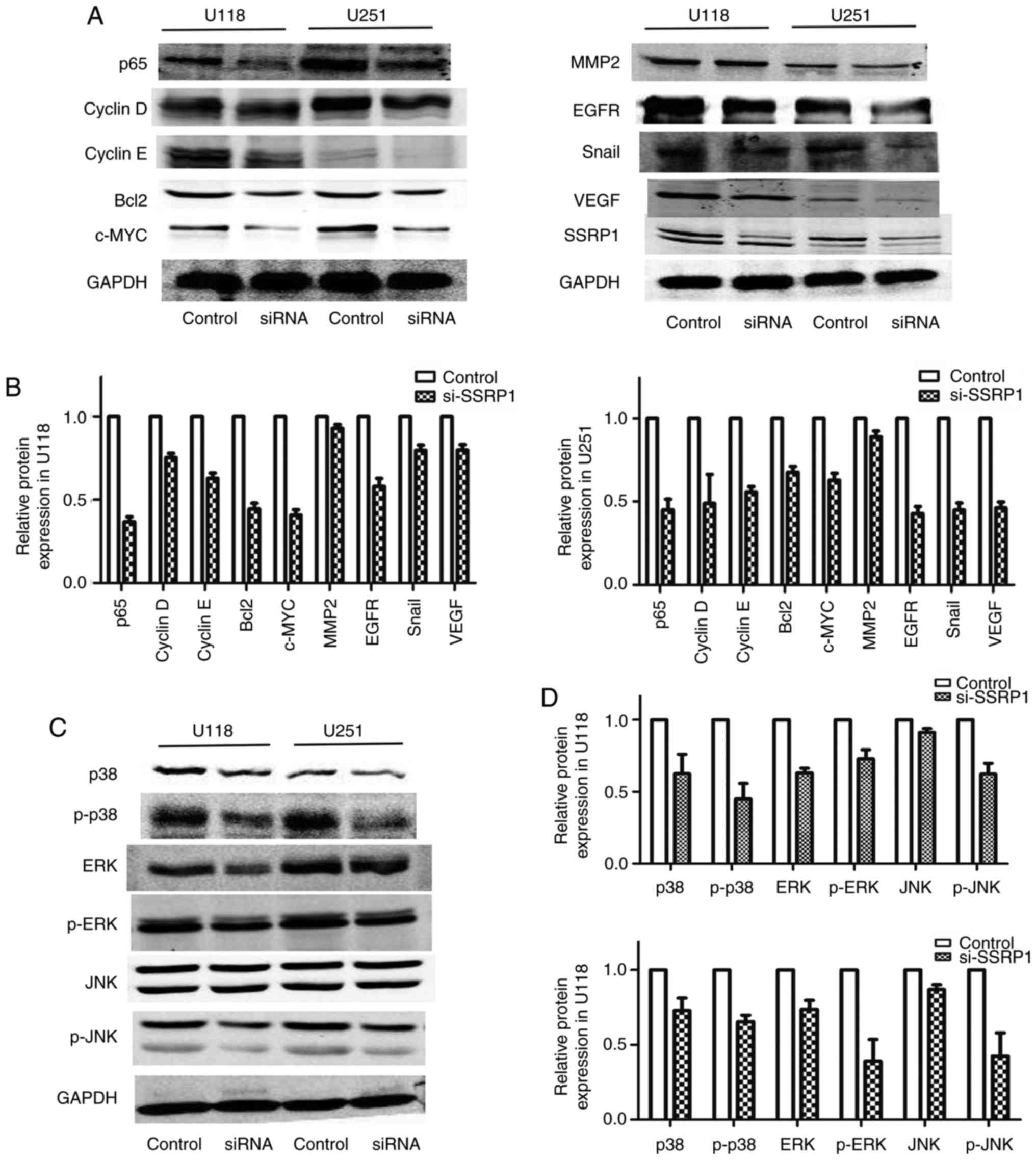
and U251 cells were analyzed. The downregulated expression of SSRP1
by si-SSRP1 suppressed the protein levels of p65, c-myc and the
cell cycle regulators including cyclin D and E (Fig. 5A). Compared to the negative control
group, the protein levels of epithelial-mesenchymal transition and
metastasis, including EGFR, VEGF and Snail, decreased in the group
transfected with si-SSRP1, but MMP2 did not decrease (Fig. 5A).
SSRP1 regulates the MAPK signaling
pathway
The MAPK signaling pathway has an important role in
regulating the proliferation and migration of glioblastoma cells.
In order to evaluate the underlying mechanisms of SSRP1 in
regulating glioma cell proliferation and migration, western blot
assay was carried out to detect the alterations in the main
proteins in the MAPK pathway. As shown in Fig. 5B, the downregulated expression of
SSRP1 significantly decreased the phosphorylation of p38, ERK and
JNK. Furthermore, the total p38 and ERK protein expression was also
decreased in the group transfected with si-SSRP1, but not the total
JNK protein level (Fig. 5B). These
results revealed that SSRP1 is an upstream factor that modulated
the MAPK signaling pathway in glioma.
Discussion
SSRP1 has been reported as a subunit of histone
chaperone protein FACT, which causes the disruption of nucleosome
and histone replacements (21,22).
It has been reported to be involved in the progression of DNA
replication, DNA damage repair, cell proliferation and cell
apoptosis (14,15,23).
Previous studies have focused on the molecular mechanism directing
transcription elongation. However, few studies have reported the
correlation between SSRP1 and cancer, and no study has focused on
the clinical significance and oncogenicity effect of SSRP1 in
glioma.
Our studies revealed that both the mRNA and protein
levels of SSRP1 were upregulated in glioma tissues, compared with
those in NB tissues, and that the SSRP1 protein level was higher in
the high-grade glioma patients than in the low-grade glioma
patients, which were identical to the results of the GEO datasets
analyses. These findings suggest that SSRP1 may play an important
role in the development of human glioma, and that the
overexpression level may predict high-grade glioma.
The overexpression of SSRP1 in glioblastoma
predicted the oncogenic role of SSRP1 in glioma tumorigenesis.
First, we used multiple methods, including the CCK-8 and EdU
incorporation assays, and FACS flow cytometric analysis, to
identify the role of SSRP1 in GBM cell growth progression.
Furthermore, we demonstrated that the viability and proliferation
abilities were significantly decreased in glioma cells with the
silencing of SSRP1 by siRNA transfection. In addition, we also
ascertained the biological function of SSRP1 on GBM cell motility,
and detected that the knockdown of SSRP1 by siRNA inhibited glioma
cell migration and invasion. Glioblastoma is the most common
aggressive phenotype of glioma, and is identified by the
characteristics of cellular proliferation, diffuse infiltration,
necrosis, angiogenesis and intense resistance to apoptosis
(24). The results of the present
study reflected that the decreased expression of SSRP1 inhibited
the ability of cell proliferation, anti-apoptosis and metastasis,
which were the hallmark features of glioblastoma.
In order to further confirm the function of SSRP1,
we examined some cell growth regulators and metastasis markers. It
was observed that the protein levels of proliferation-associated
genes, including p65, c-myc, cyclin D and E, were inhibited after
SSRP1 downregulation in glioma U118 and U251 cells. Furthermore, we
demonstrated that SSRP1 knockdown suppressed the expression of
metastasis-associated genes such as EGFR, VEGF and Snail.
The MAPK pathway regulated multiple cellular
programs including embryogenesis, proliferation, apoptosis and
differentiation, based on cues derived from the surface and
metabolic state of the cells (25).
The 3 major MAPK pathways are ERKs, JNKs and the p38 families
(26). In the present study, we
also demonstrated that the downregulated expression of SSRP1
significantly decreased the phosphorylation of p38, ERK and JNK,
and that total p38 and ERK protein expression was also decreased,
but not the total JNK protein level. Hence, the present study
revealed that SSRP1 may be involved in tumor progression via the
MAPK pathway.
In conclusion, the present study indicated that
SSRP1 may regulate the function of the MAPK pathway to promote cell
proliferation and metastasis in gliomas. SSRP1 overexpression in
glioma specimens could be used for the diagnosis of glioma, and our
results revealed the potential role for SSRP1 in glioma therapy. In
addition, further research to clarify the molecular mechanism of
SSRP1 involved in glioma tumorigenesis is warranted.
Acknowledgements
The present study was supported by grants from the
National Science Foundation of China (nos. 81502175 and
81572489).
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohgaki H and Kleihues P: Genetic pathways
to primary and secondary glioblastoma. Am J Pathol. 170:1445–1453.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baumert BG, Hegi ME, Van den Bent MJ, von
Deimling A, Gorlia T, Hoang-Xuan K, Brandes AA, Kantor G, Taphoorn
MJ, Hassel MB, et al: Temozolomide chemotherapy versus radiotherapy
in high-risk low-grade glioma (EORTC 22033–26033): A randomised,
open-label, phase 3 intergroup study. Lancet Oncol. 17:1521–1532.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Norden AD, Drappatz J and Wen PY: Novel
anti-angiogenic therapies for malignant gliomas. Lancet Neurol.
7:1152–1160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Belotserkovskaya R, Oh S, Bondarenko VA,
Orphanides G, Studitsky VM and Reinberg D: FACT facilitates
transcription-dependent nucleosome alteration. Science.
301:1090–1093. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Röttgers K, Krohn NM, Lichota J, Stemmer
C, Merkle T and Grasser KD: DNA-interactions and nuclear
localisation of the chromosomal HMG domain protein SSRP1 from
maize. Plant J. 23:395–405. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Birch JL, Tan BC, Panov KI, Panova TB,
Andersen JS, Owen-Hughes TA, Russell J, Lee SC and Zomerdijk JC:
FACT facilitates chromatin transcription by RNA polymerases I and
III. EMBO J. 28:854–865. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumari A, Mazina OM, Shinde U, Mazin AV
and Lu H: A role for SSRP1 in recombination-mediated DNA damage
response. J Cell Biochem. 108:508–518. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mason PB and Struhl K: The FACT complex
travels with elongating RNA polymerase II and is important for the
fidelity of transcriptional initiation in vivo. Mol Cell Biol.
23:8323–8333. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan BC, Liu H, Lin CL and Lee SC:
Functional cooperation between FACT and MCM is coordinated with
cell cycle and differential complex formation. J Biomed Sci.
17:112010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang W, Zeng F, Liu Y, Shao C, Li S, Lv
H, Shi Y, Niu L, Teng M and Li X: Crystal structure of human SSRP1
middle domain reveals a role in DNA binding. Sci Rep. 5:186882015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Garcia H, Fleyshman D, Kolesnikova K,
Safina A, Commane M, Paszkiewicz G, Omelian A, Morrison C and
Gurova K: Expression of FACT in mammalian tissues suggests its role
in maintaining of undifferentiated state of cells. Oncotarget.
2:783–796. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garcia H, Miecznikowski JC, Safina A,
Commane M, Ruusulehto A, Kilpinen S, Leach RW, Attwood K, Li Y,
Degan S, et al: Facilitates chromatin transcription complex is an
‘accelerator’ of tumor transformation and potential marker and
target of aggressive cancers. Cell Reports. 4:159–173. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding Q, He K, Luo T, Deng Y, Wang H, Liu
H, Zhang J, Chen K, Xiao J, Duan X, et al: SSRP1 contributes to the
malignancy of hepatocellular carcinoma and is negatively regulated
by miR-497. Mol Ther. 24:903–914. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dermawan JK, Gurova K, Pink J, Dowlati A,
De S, Narla G, Sharma N and Stark GR: Quinacrine overcomes
resistance to erlotinib by inhibiting FACT, NF-κB, and cell-cycle
progression in non-small cell lung cancer. Mol Cancer Ther.
13:2203–2214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koman IE, Commane M, Paszkiewicz G,
Hoonjan B, Pal S, Safina A, Toshkov I, Purmal AA, Wang D, Liu S, et
al: Targeting FACT complex suppresses mammary tumorigenesis in
Her2/neu transgenic mice. Cancer Prev Res. 5:1025–1035. 2012.
View Article : Google Scholar
|
|
17
|
Carter DR, Murray J, Cheung BB, Gamble L,
Koach J, Tsang J, Sutton S, Kalla H, Syed S, Gifford AJ, et al:
Therapeutic targeting of the MYC signal by inhibition of histone
chaperone FACT in neuroblastoma. Sci Transl Med. 7:312ra1762015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lake D, Corrêa SA and Müller J: Negative
feedback regulation of the ERK1/2 MAPK pathway. Cell Mol Life Sci.
73:4397–4413. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Z, Guo Q, Wang R, Xu G, Li P, Sun Y,
She X, Liu Q, Chen Q, Yu Z, et al: The D Domain of LRRC4 anchors
ERK1/2 in the cytoplasm and competitively inhibits MEK/ERK
activation in glioma cells. J Hematol Oncol. 9:1302016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shingu T, Holmes L, Henry V, Wang Q, Latha
K, Gururaj AE, Gibson LA, Doucette T, Lang FF, Rao G, et al:
Suppression of RAF/MEK or PI3K synergizes cytotoxicity of receptor
tyrosine kinase inhibitors in glioma tumor-initiating cells. J
Transl Med. 14:462016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takahata S, Yu Y and Stillman DJ: FACT and
Asf1 regulate nucleosome dynamics and coactivator binding at the HO
promoter. Mol Cell. 34:405–415. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsunaka Y, Fujiwara Y, Oyama T, Hirose S
and Morikawa K: Integrated molecular mechanism directing nucleosome
reorganization by human FACT. Genes Dev. 30:673–686. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao XJ, Feng JX, Zhu S, Liu XH, Tardieux I
and Liu LX: Protein phosphatase 2C of toxoplasma gondii interacts
with human SSRP1 and negatively regulates cell apoptosis. Biomed
Environ Sci. 27:883–893. 2014.PubMed/NCBI
|
|
24
|
Furnari FB, Fenton T, Bachoo RM, Mukasa A,
Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, et al:
Malignant astrocytic glioma: Genetics, biology, and paths to
treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Raman M, Chen W and Cobb MH: Differential
regulation and properties of MAPKs. Oncogene. 26:3100–3112. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Owens DM and Keyse SM: Differential
regulation of MAP kinase signalling by dual-specificity protein
phosphatases. Oncogene. 26:3203–3213. 2007. View Article : Google Scholar : PubMed/NCBI
|