Introduction
Colorectal cancer is one of the malignant tumors
that affects human health and life worldwide, and it is the third
most common cancer and the fourth cause of cancer-related mortality
in western countries (1). At
present, other than surgery, chemotherapy and radiotherapy are the
two dominant strategies for treating colorectal cancer.
Unfortunately, these strategies are associated with undesirable
side-effects including nausea, diarrhea, poor quality of life and
others. Thus, the development of novel anticancer agents against
colorectal cancer from nature is urgently needed. In particular,
those with a history of medicinal use from the rich traditional
Chinese medicine (TCM) resources are of great interest.
The medicinal mushroom G. lucidum, also
called Reshi in Japan and Lingzhi in China, has been consumed for
thousands of years in East Asia to promote longevity and improve
overall health (2). G.
lucidum has been reported to possess anti-inflammatory
activities (3,4), antitumor activities (5,6),
alleviating hepatotoxicity activities (7,8),
analgesic effects (9,10) and anti-HIV-1 activities (11). Currently, over 300 active compounds
have been isolated from G. lucidum fruiting bodies, mycelia
and spores (12). The bio-active
substances of G. lucidum include polysaccharides,
triterpenoids, amino acids, peptides, fatty acids, oligosaccharides
and trace elements, especially over 150 triterpenoids have been
isolated from G. lucidum (13). Among these active components,
triterpenoids (major active component of the ethanol extracts of
G. lucidum) and polysaccharides (major active component of
the water extracts of G. lucidum) have been extensively
studied for their anticancer effects against many types of cancers.
At present, a large number of studies have found that ethanol
extracts of G. lucidum has a broad spectrum anticancer
effects how human gastric (14,15),
urothelial (16), ovarian (17), colon (18) and liver (19) cancers. However, it still remains
unclear about the exact mechanism by which the ethanol extracts of
G. lucidum exert for its anticancer effects in these
cancers. In addition, most of the above studies examined
triterpenoids extracted from fruiting bodies or mycelia of G.
lucidum. Only few studies examined the triterpenoids extracted
from the sporoderm-broken spores of G. lucidum for their
anticancer effects. Min et al (20) reported that the spores contain more
triterpenoids compared with other parts of G. lucidum.
However, it has not been reported whether the sporoderm-broken
spores of G. lucidum ethanol extracts (BSGLEE) could inhibit
colorectal cancer carcinogenesis either in vitro or in
vivo.
Therefore, in the present study we examined the
anticancer effects of BSGLEE on colorectal cancer and the potential
molecular mechanisms underlying these activities of BSGLEE were
also explored. From in vitro and in vivo studies we
demonstrate that BSGLEE is effective in inhibiting HCT116 cancer
cell proliferation and tumor growth through regulating key genes
and proteins involved in apoptosis, migration and cell cycle
arrest.
Materials and methods
Materials
FITC Annexin V apoptosis detection kit and propidium
iodide (PI)/RNase staining solution were purchased from BD
Biosciences (San Diego, CA, USA). Hoechst 33342 was obtained from
Invitrogen (Carlsbad, CA, USA). [3-(4,
5-dimethylthia-zol-2-yl)-2,5-diphenyltetrazolium bromide] (MTT) was
obtained from HXBIO (Hangzhou, China). Polyclonal β-actin and PARP
antibodies, and monoclonal pro-caspase-3, cleaved caspase-3 and
pro-caspase-7 antibodies were obtained from Cell Signaling
Technology (Danvers, MA, USA). RNA extraction kit was purchased
from Aidlab Biotechnologies Co., Ltd. (Beijing, China). The iScript
cDNA Synthesis kit and SYBR Master Mix were purchased from Bio-Rad
Laboratories (Hercules, CA, USA). The bicinchoninic acid (BCA)
assay kit was purchased from Pierce (Rockford, IL, USA). The
Western Lightening™ Plus-ECL Enhanced chemiluminescence substrate
assay kit was purchased from Perkin-Elmer (Waltham, MA, USA).
Ki-67, Bax, Bcl-2 and cyclin D1 antibodies for immunohistochemistry
were obtained from Wuhan Goodbio Technology Co., Ltd. (Wuhan,
China). Transwell plates were purchased from Costar, Inc.,
(Kennebunk, ME, USA).
BSGL ethanol extract preparation
Powder of sporoderm-broken spores of G.
lucidum (BSGL) were purchased from Taian Zhengxin Science and
Technology Co., Ltd. (Anhui, China). The tritepenoids from the
powder of sporoderm-broken spores of G. lucidum were
extracted by modified protocol based on ethanol extraction method
described before (21). The
modification was based on results of orthogonal experiments.
Briefly, we adopted the following conditions: 95% of ethanol, 85°C
of extraction temperature, 2 h of extraction time, ratio of
material to liquid as 1:60 (g/ml) and 2 times of extraction. The
extraction solution was centrifuged at 3000 × g for 3 min and then
the supernatant was collected. The ethanol solvent in the
supernatant was removed using a vacuum evaporator. The dried
extracts were weighed and stored at −20°C for further analysis for
subsequent experiments. BSGLEE was weighed and dissolved in
dimethyl sulfoxide (DMSO) and further diluted using the
corresponding cell culture medium immediately at stock solution of
10 mg/ml.
Cell culture
The colon cancer cell line HCT116 was purchased from
the American Type Culture Collection (ATCC; Manassas, VA, USA).
HCT116 cells were maintained in Dulbeccos modified Eagles medium
(DMEM; Gibco, Gaithersburg, MD, USA) containing 10% fetal bovine
serum (FBS; Gibco) and 100 units/ml penicillin (Invitrogen), 0.1
mg/ml streptomycin (Invitrogen) and cultured at 37°C in a
humidified atmosphere with 5% CO2.
Morphological observation and MTT
assay
In order to explore whether HCT16 cells may be
killed by BSGLEE, morphological observation was conducted in the
test. HCT116 cells were seed in 6-well plate at 2×105
cells/well and incubated at 37°C in the presence of 5%
CO2. After 24-h incubation when cells reached ~50%
confluence, cells were treated with different concentrations of
BSGLEE (0, 0.64, 1.6, 4.0 and 10.0 mg/ml) for 48 h. Phase contrast
images of the conditioned cells were captured by Motic phase
contrast microscope equipped with a digital camera (Motic, Xiamen,
China) to obtain the effects of different concentrations of BSGLEE
on the number and morphology of HCT116 cells. In addition, cell
viability was detected by MTT assay. Briefly, HCT116 cells were
seeded in 96-well plates at 1×104 cells/well. Cells were
treated with various concentrations of BSGLEE (0, 0.64, 1.6, 4 and
10 mg/ml) in DMEM for 24, 48 and 72 h. Next, 20 µl of MTT solution
(5 mg/ml) was added to each well followed by incubation for 4 h at
37°C. Then the medium was discarded and 200 µl DMSO was added to
dissolve the formazan crystals. Viable cells were detected by
measuring absorbance at 490 nm using a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA). As BSGLEE at 0.64 mg/ml
failed to kill HCT116 as significantly as other concentrations,
0.64 mg/ml was eliminated in the subsequent experiments.
Flow cytometric analysis of apoptosis
and cell cycle arrest
The distribution of numbers of apoptotic cells and
cells in different cell cycle phases upon BSGLEE (0, 1.6, 4 and 10
mg/ml) treatments in HCT116 cells were detected by flow cytometry.
Briefly, cells (2×105 cells/well) were seeded in 6-well
plates. For apoptosis analysis, after 24-h incubation while cells
reached ~50% confluence, HCT116 cells were exposed to BSGLEE for 36
h. The treated cells were collected and washed twice with cold
phosphate-buffered saline (PBS). At least 1×105 cells
were resuspended in 100 µl 1X binding buffer containing 5 µl of
FITC Annexin V and 5 µl propidium iodide (PI). The cells were
gently vortexed, and then incubated for 15 min at RT in the dark
and 400 µl of 1X binding buffer was added to each tube. The cell
apoptosis rates were analyzed by Guava easyCyte HT flow cytometry
system (Merck KGaA, Darmstadt, Germany). For cell cycle analysis,
after 24-h incubation (~50% confluence), the cells were treated
with BSGLEE (0, 1.6, 4 and 10 mg/ml) for 48 h. According to
manufacturers protocol (BD Biosciences), cells were harvested and
fixed in 70% ethanol and then stored at −20°C for 2 h minimum. The
cells were washed twice to remove the ethanol residue and stained
in 0.5 ml of PI/RNase staining buffer in the dark at room
temperature (RT) for 30 min. The cell cycle was analyzed using the
same flow cytometer as stated above. The DNA content in the G0/G1,
S and G2/M phase was analyzed by ModFit 3.2 LT software (Verity
Software House, Topsham, ME, USA).
Hoechst staining
DNA staining was performed using the Hoechst 33342
staining to confirm the alterations of nuclei morphology in HCT116
cells after BSGLEE treatment. Briefly, HCT116 cells were treated
with BSGLEE (0, 1.6, 4 and 10 mg/ml) for 36 h and then stained with
Hoechst 33342 (10 µg/ml) for 15 min at RT in the dark. The nuclear
morphology of HCT116 cells was observed using fluorescence
microscope at a magnification of 200-fold (Nikon, Tokyo, Japan)
after being washed with PBS. Image from 5 randomly selected
microscopic fields were captured and only one representative
picture was presented.
Cell migration analysis
Cell migration ability was evaluated by wound
healing assay as previously described (22). Briefly, HCT116 cells were seeded in
6-well plates (2.0×105 cells/well), and when cells
reached ~90% confluence, wound was created by scratching the cell
monolayer using a pipette tip and then washed with PBS for removal
of cell debris. Cells were exposed to BSGLEE and allowed to migrate
into the wound area for 12 and 24 h. Images of the wound area were
captured using the inverted microscope (Nikon, Tokyo, Japan).
Transwell migration assay
Cell migration ability was evaluated by 24-well
Transwell chamber according to the manufacturers instruction.
Briefly, HCT116 cells were seeded in 6-well plates, and when cells
reached ~90% confluence, the cells were treated with different
concentrations of BSGLEE (0, 1.6, 4.0 and 10.0 mg/ml) for 24 h.
Then, cells were trypsinized and counted. Next, 2.0×105
HCT116 cells in 100 µl of serum-free medium were placed into top
chambers of Transwell plate, and 600 µl medium with 20% FBS was
added in the lower chambers. The plate was incubated in 5%
CO2 for additional 24 h at 37°C. Cells penetrating
through the porous membrane were detected by hematoxylin and eosin
staining (H&E), and then observed with light microscope (Leica
Microsystems, Wetzlar, Germany). The number of cells stained were
counted in five random fields of each chamber.
Western blot analysis
After treatment with BSGLEE for 48 h, cells were
washed twice with PBS and fully lysed with ice-cold RIPA buffer
(Sigma-Aldrich, St. Louis, MO, USA). Then cell lysates were
centrifuged at 13,000 rpm for 10 min at 4°C. Supernatants were
collected and concentration of total protein was quantified by BCA
assay. Protein samples (40 µg/lane) were separated by 10–12%
SDS-PAGE gel and then transferred to polyvinylidene fluoride
membranes (PVDF) (Merck Millipore, Darmstadt, Germany) using a
transfer system (Bio-Rad Laboratories, 100 v, 120 min). After
blocking with 5% skim milk in TBST for 1 h at RT, the membranes
were incubated overnight at 4°C in the primary antibody solution
against pro-caspase-3, cleaved caspase-3, pro-caspase-7 and PARP
antibodies according to the manufacturers instruction. Membranes
were washed 3 times with TBST for 10 min and incubated in the
HRP-conjugated secondary antibody solution (Cell Signaling
Technology, Inc., Beverly, MA, USA) for 1 h at room temperature.
The signals were detected using ECL chemiluminescence reagent
(Perkin-Elmer) and β-actin was used as loading control.
Quantitative real-time PCR
(qRT-PCR)
Total RNA was isolated using RNA kit from Aidlab
following instruction. The concentration and purity of total RNA
was determined by NanoDrop 2000 (Thermo Fisher Scientific, Waltham,
MA, USA). cDNA was synthesized with iScript cDNA Synthesis Kit
(Bio-Rad Laboratories). PCR amplification was performed using a
SYBR Master Mix (Bio-Rad Laboratories) on CFX-96 real-time PCR
system (Bio-Rad Laboratories). β-actin was used as an internal
control. Fold changes relative to the control were calculated
according to the 2−∆∆Ct method. The primer sequences are
listed in detail in Table I.
 | Table I.Primer sequences used in qRT-PCR. |
Table I.
Primer sequences used in qRT-PCR.
| Primer | Forward primer | Reverse primer |
|---|
| c-Met |
TTCTGACCGAGGGAATCATCA |
CCTTCACTTCGCAGGCAGAT |
| Cyclin D1 |
GTGGCCTCTAAGATGAAGGAGA |
GGAAGTGTTCAATGAAATCGTG |
| Cyclin E |
CAGCCTTGGGACAATAATGC |
TTGCACGTTGAGTTTGGGTA |
| CDK1 |
TAGCGCGGATCTACCATACC |
CATGGCTACCACTTGACCTGT |
| CDK2 |
CAGGATGTGACCAAGCCAGT |
TGAGTCCAAATAGCCCAAGG |
| CDK4 |
CTGGACACTGAGAGGGCAAT |
TGGGAAGGAGAAGGAGAAGC |
| p16 |
ACCAGAGGCAGTAACCATGC |
TGATCTAAGTTTCCCGAGGTTT |
| p21 |
TTAGCAGCGGAACAAGGAGT |
CGTTAGTGCCAGGAAAGACA |
| Bcl-2 |
AAGAGCAGACGGATGGAAAAAGG |
GGGCAAAGAAATGCAAGTGAATG |
| NAG-1 |
ACCTGCACAGCCATGCCCGGGCA |
CAGTGGAAGGACCAGGACTGCTC |
| Bax |
CCGATGGCAACTTCAACTGGG |
GTCAGCACTCCCGCCACAAAG |
| Fra-1 |
CAGCTCATCGCAAGAGTAGCA |
CAAAGCGAGGAGGGTTGGA |
| E-cad |
GGTCTCTCTCACCACCTCCA |
CCTCGGACACTTCCACTCTC |
| MMP-1 |
GGGAGATCATCGGGACAACTC |
GGGCCTGGTTGAAAAGCAT |
| MMP-2 |
AGTTTCCATTCCGCTTCCAG |
CGGTCGTAGTCCTCAGTGGT |
| Vimentin |
AGAGAACTTTGCCGTTGAAGC |
ACGAAGGTGACGAGCCATT |
| β-actin |
CTGGAACGGTGAAGGTGACA |
AAGGAACTTCCTTGAACAATGCA |
In vivo xenograft nude mouse
study
To determine the inhibitory effects of BSGLEE on
occurrence and development of colon tumor in vivo, in total
44, 5-week-old male BALB/c nude mice were fed in specific pathogen
free (SPF) environment. All experimental procedures were conducted
following the Guide for the Use and Care of Laboratory Animals of
the National Institutes of Health. This study was approved by the
Committee on the Ethics of Animal Experiments of Zhejiang Chinese
Medical University (Permit no: SYXK 2012–0002). HCT116 cells
(5×106) suspended in 200 µl PBS were transplanted into
the right flank of the nude mice. The nude mice were randomly
divided into normal group (no HCT116 cell injection, n=8, PBS),
model group (n=12, PBS), low-dose group (n=12, 75 mg/kg BSGLEE) and
high-dose group (n=12, 150 mg/kg BSGLEE) on the day of cell
transplantation. The day after HCT116 injection, the mice were
treated with BSGLEE once a day by oral gavage. Palpable tumor was
examined every day until no more tumors formed in each group. The
mice were weighed and tumor sizes were measured using a digital
vernier caliper (0.01 mm) twice a week. After 5 weeks of treatment,
the mice were sacrificed and dissected. Tumor tissues were weighed
and fixed in 4% formalin for 24 h and then processed and embedded
in paraffin. Tumor volume was calculated as follows: Tumor volume =
(length × width × width)/2.
Immunohistochemistry and H&E
staining
Paraffin-embedded tissue blocks were cut into serial
sections (4 µm) and the expression of Ki-67, Bax, Bcl-2 and cyclin
D1 were determined by immunohistochemistry staining. Briefly, the
sections were deparaffinized using citric acid buffer (pH 6.0), and
then the slides were treated with 3% hydrogen peroxide to block
endogenous peroxidase activity before incubation with Ki-67
(GB13030-2, 1:1,000 diluted in 1% BSA), Bcl-2 (GB12008-1, 1:400),
Bax (GB11007, 1:300) and cyclin D1 (GB GB13079, 1:200) primary
antibodies. The slides were then incubated with 5 µg/ml
biotinylated anti-goat IgG secondary antibody (Dako, Carpinteria,
CA, USA) for 30 min at room temperature. After washing, slides were
stained with 3,3-diaminobenzidin (DAB; Dako), and then
counterstained with hematoxylin, dehydrated and mounted with a
coverslip. For hematoxylin and eosin (H&E) staining, sections
were deparaffinized with xylene for 10 min and then rehydrated. The
sections were stained with hematoxylin for 10 min and then stained
with eosin for 1 min. The sections were dehydrated and mounted. All
images were captured using an inverted fluorescence microscope
(Nikon, Tokyo, Japan).
Statistical analysis
The SPSS 17.0 software (SPSS, Inc., Chicago, IL,
USA) was used for statistical analysis. The data are presented as
mean ± standard deviation or standard error. Comparisons among the
different groups were performed by the one-way analysis of variance
(ANOVA). A probability value of P<0.05 was considered
significant.
Results
BSGLEE inhibits cell proliferation in
HCT116 cells
MTT assay and microscope observation were used to
examine anti-proliferation effects of BSGLEE (0–10 mg/ml) in HCT116
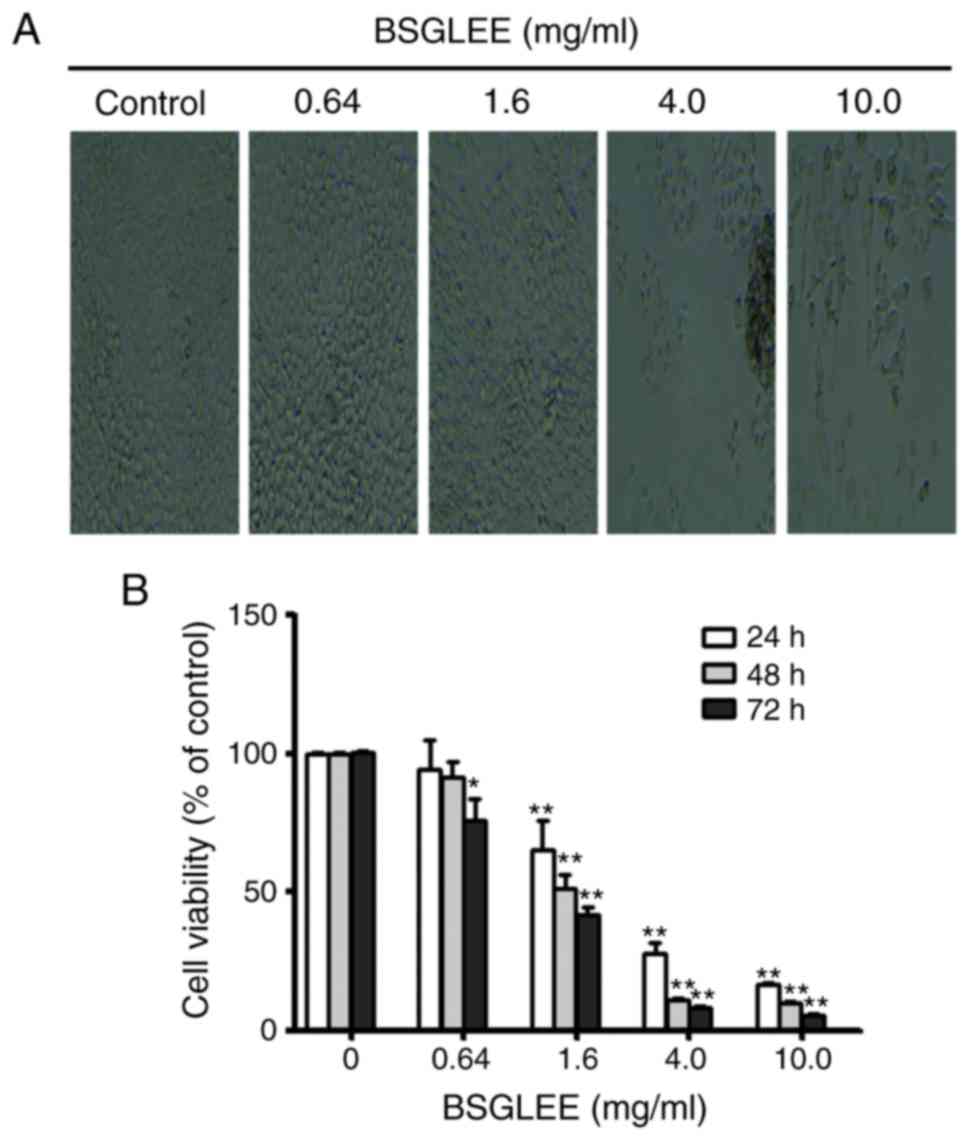
cells. As shown in Fig. 1A, the
number of HCT116 cells with BSGLEE incubation for 48 h decreased
significantly compared to control cells, which is inversely
correlated with the dose of BSGLEE. MTT assay further confirmed
that within the concentration of BSGLEE range from 1.6 to 10 mg/ml,
the proliferation of HCT116 cells was significantly inhibited with
the increase of the dosage of BSGLEE (P<0.01; Fig. 1B). We found that the inhibitory
effect of BSGLEE on HCT116 cell proliferation is also
time-dependent. As shown in Fig.
1B, at each dose of BSGLEE, cell proliferation decreased
significantly with prolonged treatment from 24 to 72 h (P<0.01).
To further determine the relationship between time and efficacy of
BSGLEE activity, the IC50 values of BSGLEE in HCT116
cells were calculated by the CompuSyn software to be 2.79, 1.77 and
1.29 mg/ml at 24, 48 and 72 h, respectively (data not shown). Taken
together, these results suggest that BSGLEE could significantly
inhibit HCT116 cancer cell proliferation at a time- and
dose-dependent manner. As the BSGLEE at 0.64 mg/ml did not show
significant inhibition of cell viability as determined by MTT
assay, we eliminated this concentration in subsequent
experiments.
BSGLEE induces G0/G1 cell cycle arrest
in HCT116 cells
We next examined the cell cycle distribution of
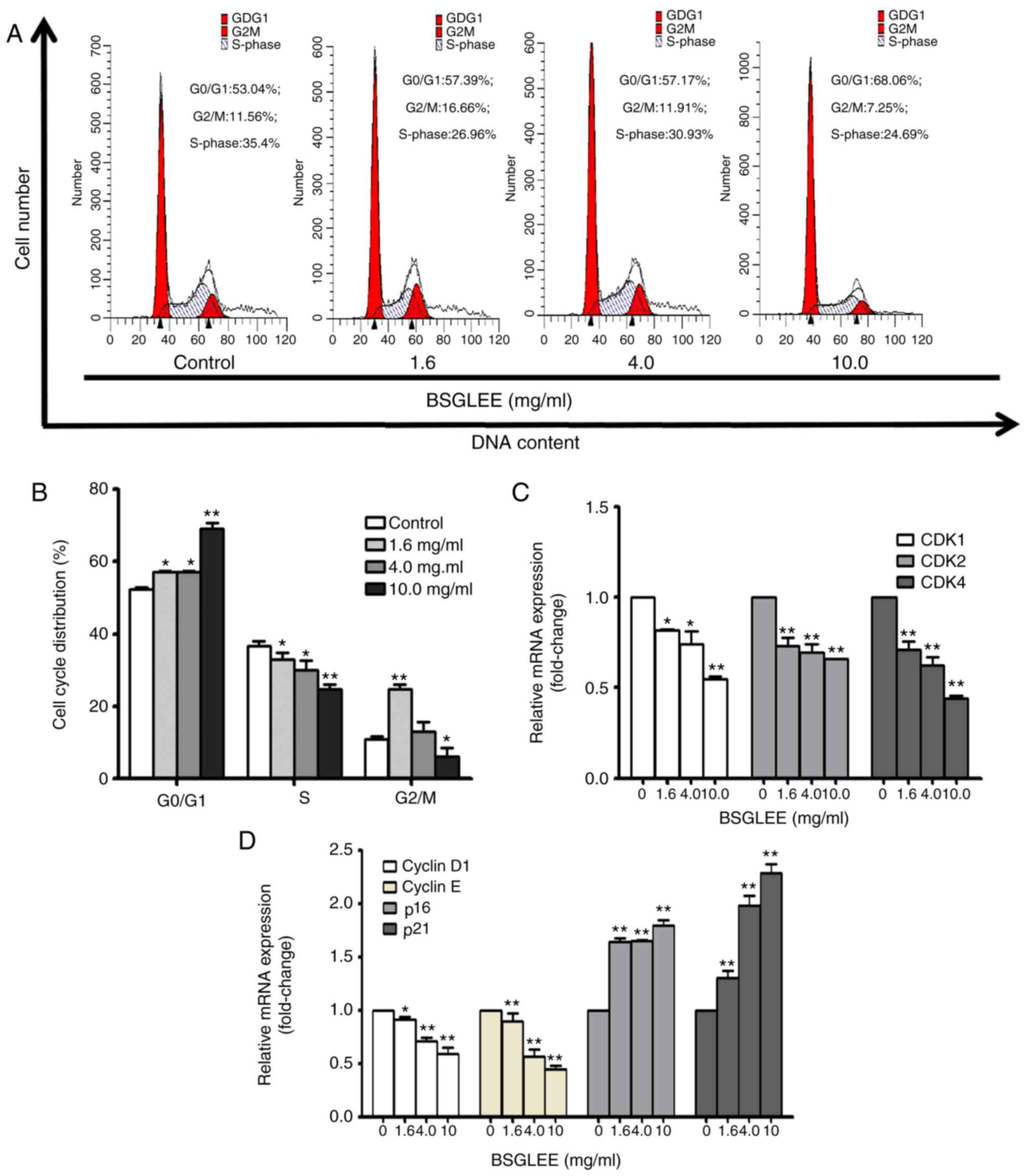
HCT116 cells treated with different concentrations of BSGLEE at 48
h. Our result indicated that BSGLEE treatment caused a significant
accumulation of cell population in the G0/G1 phase compared with
untreated cells (Fig. 2A and B;
P<0.05). In particular, the highest concentration of BSGLEE (10
mg/ml) increased up to 28% more cell accumulated in G0/G1 phase
(Fig. 2A), suggesting a G0/G1 phase
arrest by BSGLEE in HCT116 cells. Furthermore, BSGLEE also
significantly reduced cell accumulation in S phase (Fig. 2A and B). To further examine the
underlying mechanism of the anticancer effects of BSGLEE, we
conducted qRT-PCR assay to detect the expression of key genes that
regulate cell cycle progression in HCT116 cells. Our results showed
that the relative mRNA expression levels of CDK1, CDK2, CDK4,
cyclin D1 and cyclin E were downregulated, while the relative mRNA
expression levels of p16 and p21 were upregulated upon BSGLEE
treatments in a dose-dependent manner (P<0.05; Fig. 2C and D). Collectively, these data
indicate that BSGLEE could significantly inhibit HCT116 cell
proliferation through inducing cell cycle arrest at G0/G1 phase
which is associated with regulating key genes that modulate cell
cycle progression.
BSGLEE induces apoptosis in HCT16
cells
It is well recognized that one important type of
cell death caused by anticancer agents is due to apoptosis. Thus,
we examined whether BSGLEE could induce apoptosis in HCT116 cells.
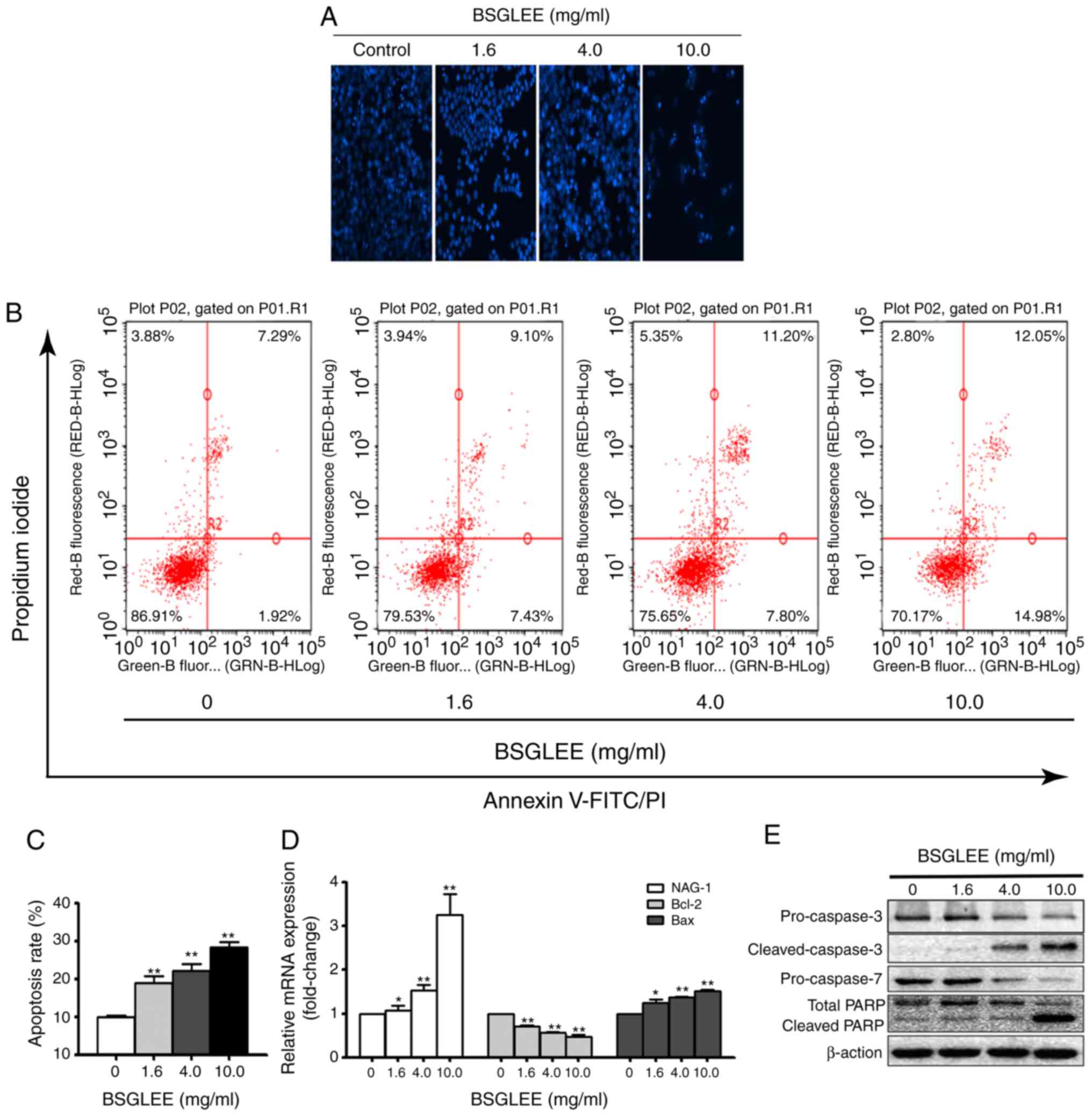
As shown in Fig. 3A, we found that
HCT116 cells exposed to BSGLEE at 36 h exhibited morphological
features of apoptotic cells as observed with an increased
percentage of cells with brighter nuclear and fragments that were
determined by Hoechst staining assay (Fig. 3A). Next, the distribution of
apoptotic cells was quantified by flow cytometry. As shown in
Fig. 3B, BSGLEE induced a
dose-dependent increase in the proportion of early and late
apoptotic cells in HCT116 cells at 36 h of treatment (P<0.05;
Fig. 3C). Data presented in
Fig. 3B is one representative image
out of three experiments. To further explore underlying mechanism
of BSGLEE induced apoptosis in HCT116 cells, qRT-PCR and western
blot analysis were conducted to examine the key molecules involved
in apoptosis cascade. As shown in Fig.
3D, the relative mRNA expression level of Bcl-2 was
downregulated (P<0.05), while the mRNA expression level of Bax
was upregulated compared with the untreated cells (P<0.05). A
number of studies have shown that non-steroidal anti-inflammatory
drug-activated gene-1 (NAG-1), a pro-apoptotic gene, is upregulated
by many anticancer agents and may play an important role in
apoptosis induced by these anticancer agents (23). Indeed, we found that the relative
mRNA level of NAG-1 was upregulated upon BSGLEE treatment,
especially by the higher concentrations of BSGLEE (P<0.05;
Fig. 3D). Western blot results
showed that the expression of pro-caspase-3 and pro-caspase-7 were
reduced upon BSGLEE treatments while cleaved caspase-3 was
upregulated, in particular by 10 mg/ml of BSGLEE, suggesting
caspase activation by BSGLEE. In addition, total PARP level was
reduced while the cleaved-PARP was upregulated in HCT116 cells upon
BSGLEE treatment (Fig. 3E). Taken
together, BSGLEE significantly induced apoptosis through
de-regulating key genes and proteins that regulate apoptosis
cascade in HCT116 cells.
BSGLEE inhibits migration in HCT116
cells
To explore whether the anticancer effect of BSGLEE
in vitro is related to cell migration, we examined the
motility of HCT116 cells via the scratch wound healing assay.
Immediately after cells reached 90% confluency, the cells were
scratched to create wounds. Cells were then treated with a various
concentration of BSGLEE (1.6, 4.0 and 10.0 mg/ml) and wound-healing
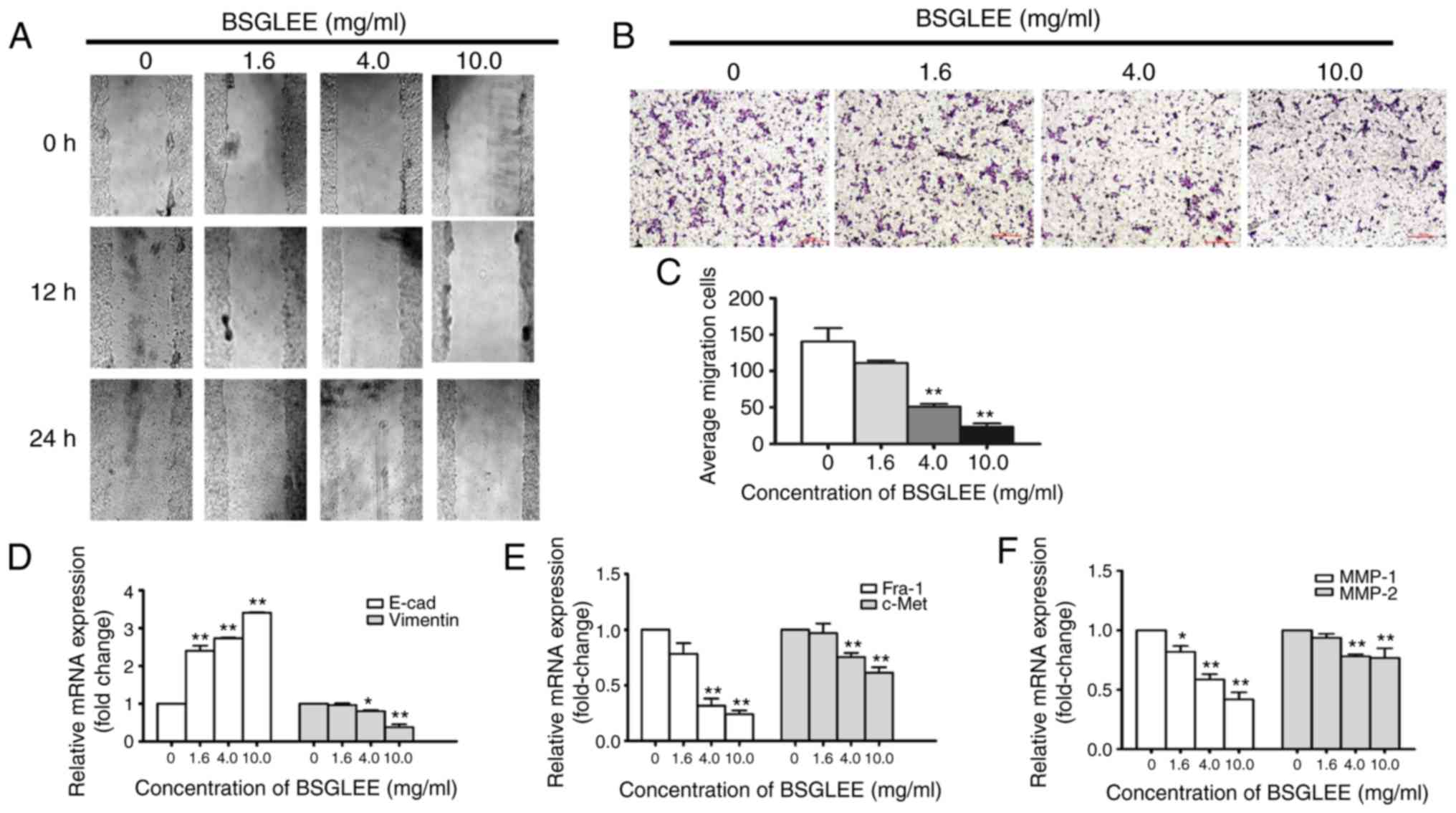
was observed at 12 and 24 h. As depicted in Fig. 4A, treatment with BSGLEE
time-dependently delayed cell motility when compared with the
controls. In addition, we found that the healing ability of cells
was gradually reduced upon BSGLEE treatment in a dose-dependent
manner (Fig. 4A). To confirm
results from wound healing experiment, we further performed
Transwell assay. Compared to control cells, the number of cells
treated with BSGLEE that penetrated through the porous membrane (as
stained purple) was significantly decreased in a dose-dependent
manner (Fig. 4B and C). These data
demonstrated that BSGLEE could significantly inhibit HCT116 cell
migration in dose-dependent manner. These observations suggest that
the migration of HCT116 cells could be inhibited by BSGLEE in a
dose- and time-dependent manner. Next, we examined the expression
of key genes that are related with cell migration upon BSGLEE
treatment in HCT116 cells. As determined by qRT-PCR assay, the
relative mRNA expression levels of Fra-1, c-Met and vimentin were
downregulated in the HCT116 cells upon BSGLEE treatment, especially
at significant level by higher doses (Fig. 4D and E). The relative mRNA
expression of MMP-1 and MMP-2, but not MMP-9 (data not shown), was
significantly reduced upon BSGLEE treatment with MMP-1 and has a
better dose-response (P<0.05; Fig.
4F). Besides, we found the mRNA level of E-cadherin, a tumor
suppressor gene that controls cell adhesion, was significantly
upregulated in a dose-dependent manner by BSGLEE (P<0.05;
Fig. 4D). However, there was no
significant change of the expression upon BSGLEE treatments of
Snail, Twist, and Slug which are all cell adhesion and migration
regulators in HCT116 cells (data not shown). Taken together, these
results suggest that BSGLEE could suppress migration of HCT116
cells through downregulating the expression of MMP-1 and MMP-2 and
upregulating E-cadherin.
BSGLEE inhibits xenograft tumor growth
in vivo
To evaluate the antitumor effects of BSGLEE on
initiation and development of colorectal cancer in vivo, we
examined rate of tumor formation, tumor volume and tumor weight in
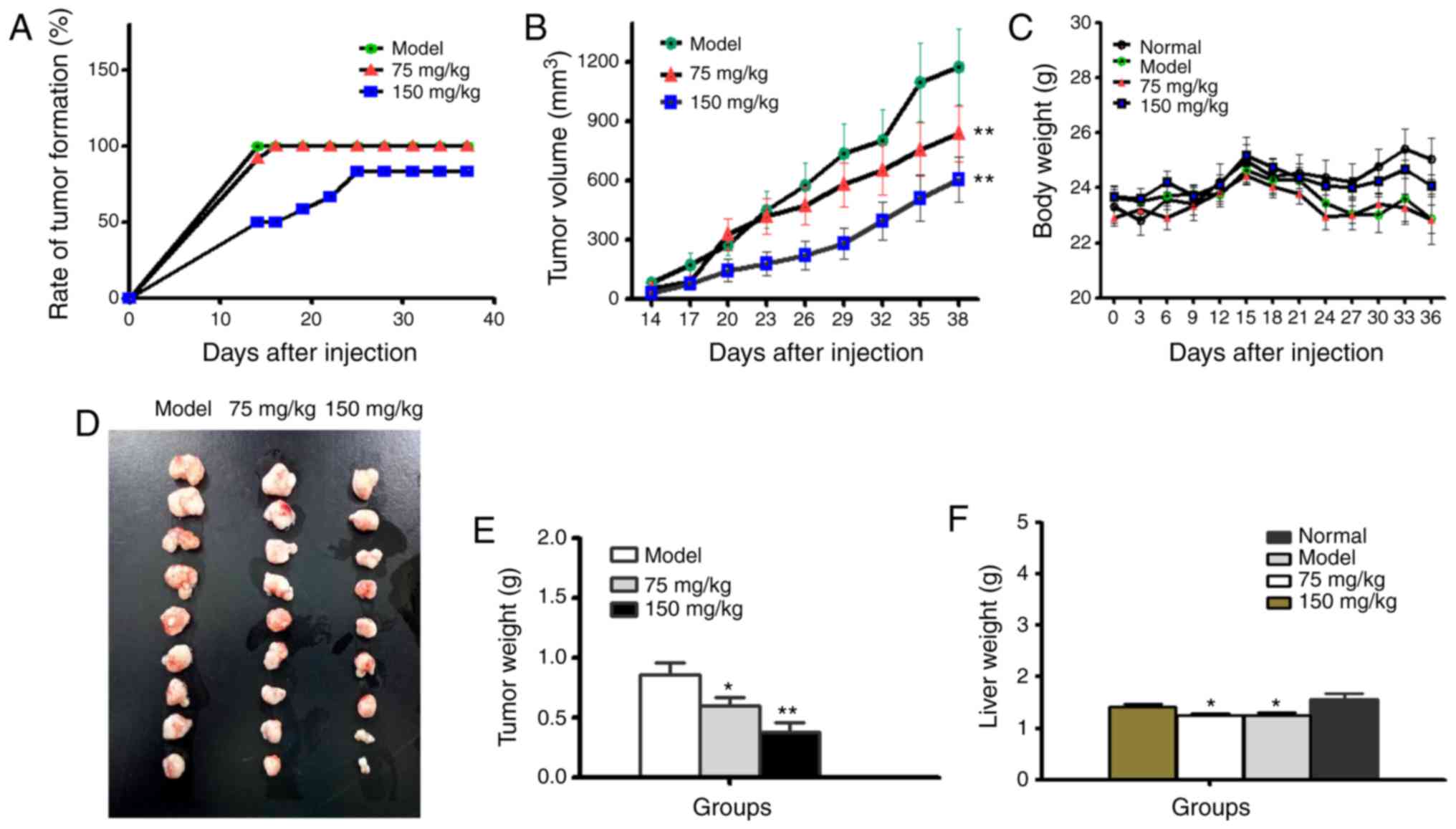
xenograft nude mice upon BSGLEE treatment. As shown in Fig. 5A, rate of tumor formation were
dose-dependently delayed by BSGLEE treatments. In the model group,
100% (12/12) of mice developed palpable tumor 14 days after
injection of HCT116 cells. In contrast, only 58.3% (7/12) of mice
in 75 mg/kg and 50% (6/12) of mice in 150 mg/kg BSGLEE treatment
groups developed palpable tumor 14 days after injection (Fig. 5A). In addition, by day 19, 75 mg/kg
BSGLEE group reached maximum tumor formation rate as 91.3% (11/12),
while until 25 days after HCT116 injection, 150 mg/kg group reached
maximum tumor formation rate at 83.3% (10/12) (Fig. 5A). These data indicate that oral
gavage of BSGLEE could effectively delay tumor formation in a
dose-dependent manner, suggesting a chemopreventive role in colon
cancer at early stage. During the experiment, the volume of tumors
in each group increased continuously (Fig. 5B). However, the tumor volume of
BSGLEE treated groups was significantly smaller than that of model
group (Fig. 5B). Although the body
weights of tumor-bearing mice are all lower than normal group (no
HCT116 cell injection), there were no significant differences in
body weights among tumor-bearing mice throughout the experiment
(Fig. 5C). At the end of the
experiment, the body weight of normal group (without HCT116 cell
injection), model group, 75 mg/kg group and 150 mg/kg group were as
follows: 25.06±1.08, 23.34±2.52, 22.84±1.5 and 23.14±2.47 g,
respectively (Fig. 5C).
At necropsy, all tumors were carefully excised and
displayed according to their sizes as shown in Fig. 5D. It is obvious that tumors in
BSGLEE treated groups are much smaller than model group. The mean
tumor weights were 0.86±0.28, 0.59±0.20 and 0.38±0.23 g for model
group, 75 mg/kg group and 150 mg/kg group, respectively (P<0.05;
Fig. 5E). In addition, we found the
weights of liver tissues of the nude mice in the model and 75 mg/kg
BSGLEE groups were significantly lower than normal group
(P<0.05; Fig. 5F), suggesting
xenograft tumor had a negative effect on liver weights. However,
150 mg/kg of BSGLEE treatment seemed to alleviate this effect which
was probably due to smaller tumor burden. Notably, the body weight
of nude mice in 150 mg/kg group seemed also closer to normal group
compared to model group and 75 mg/kg BSGLEE treatment group
(Fig. 5C), which may probably also
due to smaller tumor sizes in 150 mg/kg group.
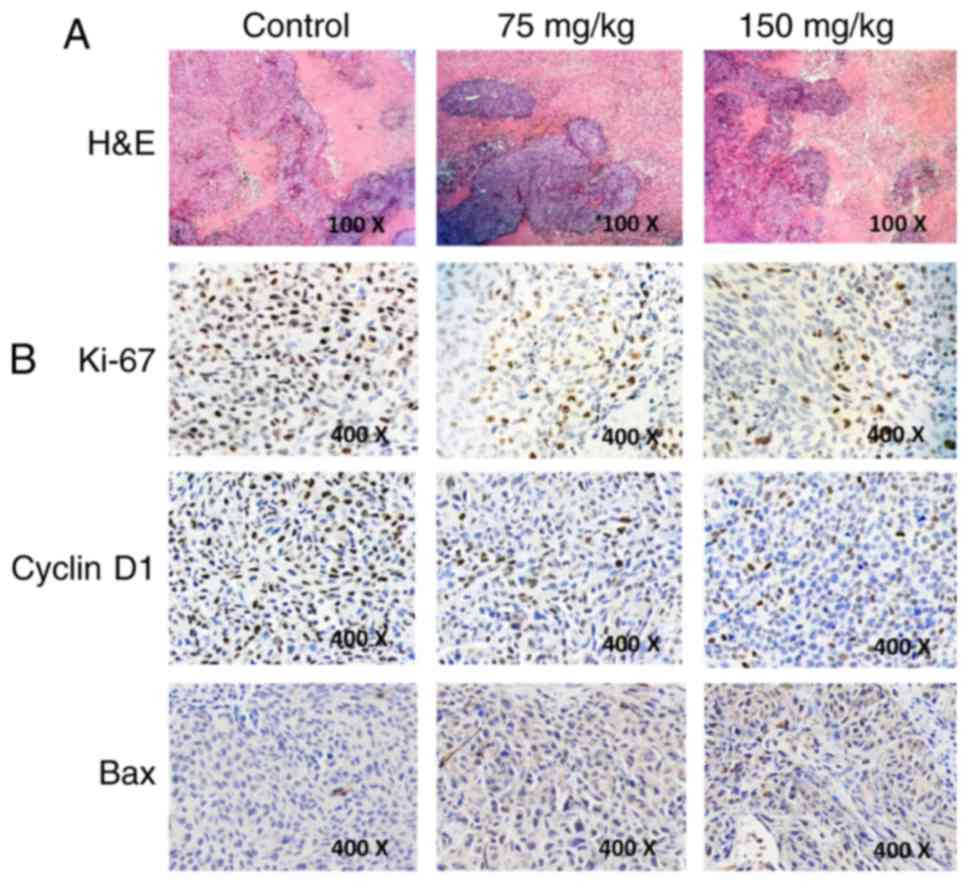
Since necrosis is often induced in tumors upon
treatment with anticancer agents, we examined whether BSGLEE
treatment could induce more necrosis than model group in xenograft
tumors by H&E staining. However, H&E staining revealed that
BSGLEE did not induce more necrosis in xenograft tumors as
characterized by large blurred, massive, unstructured red-stained
material as compared with dark purple stained living cells in
non-necrotic area (Fig. 6A).
Besides, we examined the molecular mechanisms through which BSGLEE
inhibits tumor growth in HCT116 xenografts by immunochemistry
staining. Ki-67 staining of tumor sections was performed to examine
whether proliferation was reduced upon BSGLEE treatment. We found
that both 75 and 150 mg/kg of BSGLEE dramatically decreased Ki-67
staining, suggesting a reduction of proliferation in tumor samples
upon BSGLEE treatments (Fig. 6B).
In addition, we found the expression of cell cycle regulator cyclin
D1 was dramatically reduced upon BSGLEE treatment, especially by
150 mg/kg of the ethanol extracts (Fig.
6B). We also performed TUNEL staining to determine whether
BSGLEE could induce apoptosis in xenograft tumors. However, TUNEL
staining failed to show BSGLEE could induce apoptosis in tumor
samples (data not shown), but we did observe an increased intensity
of Bax staining in the tumor sections treated with BSGLEE in a
dose-dependent manner (Fig. 6B),
while we could not detect Bcl-2 expression (data not shown). Taken
together, our in vivo study suggests that BSGLEE is
effective in inhibiting HCT116 xenograft development and
progression through multiple mechanisms in nude mice.
Discussion
Cancer patients treated by chemotherapy and/or
radiotherapy often suffer serious side-effects. The use of
traditional Chinese medicine (TCM) in cancer prevention and therapy
has received recognition by the West as adjunct/alternative to
conventional cancer therapy and prevention (24). An understanding of the molecular
basis and highlighting their potential applications for cancer
treatment is crucial (24). Among
numerous TCM, G. lucidum is one of the most widely studied
TCM. G. lucidumis a medicinal mushroom that has been used in
East Asian for over 2000 years for promotion of vitality and
longevity. At present, a large number of studies have demonstrated
that the ethanol extracts of G. lucidum from fruiting bodies
or mycelia that mainly contain triterpenoids have a variety of
anti-carcinogenesis effects in many types of cancer. Recently, with
the advance in sporoderm-breaking technology, ethanol extracts from
the sporoderm-broken spores of G. lucidum (BSGL), which
contains more triterpenoids (12,20)
became a new field of interest for prevention studies. However, the
anticancer effects and mechanism by which the BSGL ethanol extract
(BSGLEE) exerts on colorectal cancer have not yet been reported.
The present study demonstrates that BSGLEE is very potent in
inhibiting colorectal cancer HCT116 cell proliferation and
xenograft tumor development through inducing cell cycle arrest,
inhibiting migration and increasing apoptosis, which are associated
with deregulation of key molecules that regulating these
pathways.
During the multistep development of human tumors, at
least six biological capabilities are acquired by cells undergoing
tumorigenesis, which are considered as hallmarks of cancer
(25). These hallmarks include
sustaining proliferative signaling, evading growth suppressors,
resisting cell death, enabling replicative immortality, inducing
angiogenesis and activating invasion and metastasis (25). It has been well recognized that the
inhibition of proliferation, migration and induction of apoptosis
are significant mechanisms through which anticancer agents exert to
inhibit carcinogenesis. In this study, we found that BSGLEE
significantly suppressed HCT116 cell proliferation in a dose- and
time-dependent manner. This inhibitory effect of BSGLEE on colon
cancer cells is associated with blocking the cell cycle progression
at the G0/G1 phase and induction of apoptosis.
Cell cycle progression is regulated by a complex of
cell-division cyclins and cyclin-dependent kinases (CDKs) (26). It has been reported that tumor cells
frequently display an upregulated expression of cyclin D1 (27,28),
cyclin E (29), CDK1, CDK2 and CDK4
(30,31). p16 and p21 are members of CDK
inhibitor (CKI) family which can specifically inhibit activity of
cyclins and CDKs and thus delay cell cycle progression. In various
tumor cell lines and solid tumors, the expression of CKIs was
suppressed (32). Consistent with
literature, we found that the mRNA expression of p16 and p21 was
upregulated upon BSGLEE treatment in HCT116 cells. In addition, a
reduction of cyclin D, cyclin E, CDK1, CDK2 and CDK4 expression at
mRNA levels was also observed. These data suggest that
anti-proliferative effects of BSGLEE in HCT116 cells may be through
inhibiting cell cycle progression. Correlating with our study, Ruan
et al (33) reported that a
mixture of triterpenoids of G. lucidum induced cell
accumulation at G1 phase in HeLa cells. A recent study found G.
lucidum triterpenoid-induced primarily cell cycle arrest at
G1/G0 phase was due to upregulation of p21 expression and the
downregulation of CDK4 expression in prostate cancer cells
(34). These data suggest that cell
cycle arrest is one of the potential mechanisms by which BSGLEE
elicits its anti-proliferation activity in colorectal cancer
cells.
Apoptosis is an independent cell ordered death
process, which is regulated by strict and complex signal network
with a series of related genes needing to be coordinated (25). Anticancer agent that can effectively
induce apoptosis of cancer cells could serve as a promising
candidate for cancer chemoprevention or therapy. In the present
study, we found that BSGLEE was effective in inducing apoptosis in
HCT116 cells in a dose-dependent manner. Many studies reported that
Bcl-2 is highly expressed in colon cancer (35,36)
and overexpression of Bax increases apoptosis induced by
chemotherapeutic agents (35). The
pro-apoptotic gene NAG-1 is a divergent member of the transforming
growth factor-beta (TGF-β) family. Shim and Eling reported that
NAG-1 expression in VES-treated PC-3 human prostate carcinoma cells
was upregulated and NAG-1 plays an important role in VES-induced
apoptosis (37). Notably, we also
found that the relative mRNA expression level of NAG-1 and Bax in
BSGLEE-induced HCT116 cells were upregulated, while Bcl-2
expression is downregulated. In addition, western blot analysis
showed that the expression of pro-caspase-3 and pro-caspase-7 were
downregulated, suggesting a caspase activation, which play a
pro-apoptotic role in cells. Consistent with our results, Chen
et al (38) reported that
BSGLEE suppressed the growth of human lung cancer cells through
inducing apoptosis, which may be partially regulated through
inhibition of the Akt/mTOR signaling pathway. Autophagy, another
form of cell death has been shown to play a role in anticancer
agents-induced cell death in cancer cells as well. Some scholars
reported that triterpenoids from G. lucidum could induce
autophagy in colon cancer cells through inhibition of the
expression of p38 mitogen-activated kinase (p38 MAPK) (39). However, whether BSGLEE could induce
autophagy in HCT 116 cells need to be examined in future
studies.
In addition, we found that BSGLEE inhibited cell
migration as determined by wound healing and Transwell assays, and
this effect was associated with a significant upregulation of
E-cadherin (E-cad) and downregulation of MMP-1 and MMP-2.
E-cadherin, a tumor suppressor gene, plays an important role in the
process of cell adhesion, which is downregulated in many tumors
(40). MMP-1 is an interstitial
collagenase and member of the zinc-dependent endopeptidase family
(41). MMP-1 expression is
upregulated in many cancers which is associated with poor patient
outcome (41). MMP-2 also plays an
important role during carcinogenesis. Consistent with our results,
Martínez-Montemayor et al (42) reported that G. lucidum whole
extract significantly inhibited breast cancer cell invasion through
upregulating E-cadherin and downregulating MMP-9. Study by Jang
et al (14) found ethanol
extracts of G. lucidum induced anti-invasiveness in human
gastric cancer AGS cells through inhibiting MMP-2. Taken together,
this study suggests that BSGLEE inhibited migration of HCT116 cells
mainly through downregulating MMP-1 and MMP-2, and upregulating
E-cadherin expression.
To further examine the anti-carcinogenesis effect of
BSGLEE in colon cancer, we conducted in vivo study using
xenograft nude mice. We found that BSGLEE could significantly
inhibit formation and growth of xenograft tumors in nude mice.
Treatment with BSGLEE obviously decreased the proliferation and
slightly induced apoptosis in xenograft tumors, as evidenced by the
Ki-67 and Bax staining assay, which was consistent with our results
in vitro. Correlating with our study, a methanol extract
from G. lucidum significantly inhibited B16 mouse melanoma
growth in vivo (43).
Several studies suggest that G. lucidum has a
hepatoprotective effect (7,43,44).
One study found that the ethanol extract of G. lucidum
treatment is effective in protecting against ethanol-induced acute
hepatic injury in SD rats by modulating the activities of
ethanol-metabolizing enzymes and by attenuating oxidative stress
(7). Shi et al (44) found pretreatment of mice with G.
lucidum peptides reduced D-galactosamine (D-GalN)-induced
hepatic injury, including a significant decrease in the activity of
superoxide dismutase (SOD) and in the glutathione (GSH) level in
the liver. In this study, we were not able to examine whether G.
lucidum have the above effects in liver of the nude mice. We
did observe an increase of liver tissue weight upon 150 mg/kg
BSGLEE treatment. However, the increase of liver weight by 150
mg/kg BSGLEE may only be due to less tumor burden in this group. In
addition, we found that BSGLEE supplementation did not show any
toxicity to the mice. Several clinical studies demonstrated that
supplementation with G. lucidum is overall safe in human
subjects. For example, a prospective, randomized, double-blind,
placebo-controlled study reported that 4-week intake of 1.5 g/day
G. lucidum did not impair platelet and global hemostatic
function and was demonstrated to be safe in healthy volunteers
(45). In one double-blind,
placebo-controlled, randomized clinical study, Noguchi et al
(46) reported that G.
lucidum ethanol extract (6 mg once a day) was well tolerated
and significantly improved lower urinary tract symptoms in men.
Another study found that G. lucidum intake for 4 weeks in a
controlled human supplementation study shows no toxicity to
subjects (47). Although many
studies have examined anticancer effects of G. lucidum
triterpenoids in laboratory studies, clinical studies are still
lacking. Compared to several available studies using water extract
of G. lucidum (mainly contains polysaccharides), few or no
studies examined health benefits of G. lucidum ethanol
extracts in cancer patients. One randomized, double-blind,
placebo-controlled study reported a Chinese medicinal herb complex
that contains whole extracts of G. lucidum has significantly
improved the immune function of cancer patients receiving
chemotherapy and/or radiotherapy (48). However, evidence from well-designed
human clinical trials is still scarce. In particular, clinical
studies using ethanol extracts of G. lucidum as new
nutraceutical or drug for the prevention and treatment of
colorectal cancer are needed in future studies.
In conclusion, the present study demonstrated that
the ethanol extracts that mainly contain triterpenoids of the
sporoderm-broken spores of G. lucidum significantly
inhibited colorectal cancer cell proliferation and xenograft tumor
growth through deregulating expression of the key molecules of cell
cycle, apoptosis and proliferation. To the best of our knowledge,
this is the first study to examine the chemopreventive effects of
BSGLEE that mainly contains a mixture of triterpenoids in
colorectal cancer. Our results also indicate that BSGLEE may serve
as a novel anticancer agent for colorectal cancer prevention and
therapy.
Acknowledgements
We thank Yu Huang at the Animal Facility of Zhejiang
Medical University for maintenance of the nude mice. The present
study was supported by the National Natural Science Foundation of
China (grant no. 81473397).
Glossary
Abbreviations
Abbreviations:
|
G. lucidum
|
Ganoderma lucidum
|
|
BSGL
|
powder of sporoderm-broken spores of
G. lucidum
|
|
pBSGLEE
|
ethanol extracts of sporoderm-broken
spores of G. lucidum
|
|
NAG-1
|
non-steroidal anti-inflammatory
drug-activated gene-1
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bishop KS, Kao CH, Xu Y, Glucina MP,
Paterson RR and Ferguson LR: From 2000 years of Ganoderma lucidum
to recent developments in nutraceuticals. Phytochemistry.
114:56–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Akihisa T, Nakamura Y, Tagata M, Tokuda H,
Yasukawa K, Uchiyama E, Suzuki T and Kimura Y: Anti-inflammatory
and anti-tumor-promoting effects of triterpene acids and sterols
from the fungus Ganoderma lucidum. Chem Biodivers. 4:224–231. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ko HH, Hung CF, Wang JP and Lin CN:
Antiinflammatory triterpenoids and steroids from Ganoderma lucidum
and G. tsugae. Phytochemistry. 69:234–239. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu H, Ahn NS, Yang X, Lee YS and Kang KS:
Ganoderma lucidum extract induces cell cycle arrest and apoptosis
in MCF-7 human breast cancer cell. Int J Cancer. 102:250–253. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu G, Qian Z, Guo J, Hu D, Bao J, Xie J,
Xu W, Lu J, Chen X and Wang Y: Ganoderma lucidum extract induces G1
cell cycle arrest, and apoptosis in human breast cancer cells. Am J
Chin Med. 40:631–642. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jang SH, Cho SW, Yoon HM, Jang KJ, Song CH
and Kim CH: Hepatoprotective evaluation of Ganoderma lucidum
pharmacopuncture: In vivo studies of ethanol-induced acute liver
injury. J Pharmacopuncture. 17:16–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mowsumi FR, Rahman MM, Rahaman A, Selina
FA, Islam MJ and Bhuiyan MHS: Preventive effect of Ganoderma
lucidum on paraetamol-induced acute hepatotoxicity in rats.
Bangladesh J Sci Res. 5:573–578. 2013.
|
|
9
|
Lam FF, Ko IW, Ng ES, Tam LS, Leung PC and
Li EK: Analgesic and anti-arthritic effects of Lingzhi and San Miao
San supplementation in a rat model of arthritis induced by Freunds
complete adjuvant. J Ethnopharmacol. 120:44–50. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koyama K, Imaizumi T, Akiba M, Kinoshita
K, Takahashi K, Suzuki A, Yano S, Horie S, Watanabe K and Naoi Y:
Antinociceptive components of Ganoderma lucidum. Planta Med.
63:224–227. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
el-Mekkawy S, Meselhy MR, Nakamura N,
Tezuka Y, Hattori M, Kakiuchi N, Shimotohno K, Kawahata T and Otake
T: Anti-HIV-1 and anti-HIV-1-protease substances from Ganoderma
lucidum. Phytochemistry. 49:1651–1657. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu GS, Guo JJ, Bao JL, Li XW, Chen XP, Lu
JJ and Wang YT: Anti-cancer properties of triterpenoids isolated
from Ganoderma lucidum - a review. Expert Opin Investig Drugs.
22:981–992. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Boh B, Berovic M, Zhang J and Zhi-Bin L:
Ganoderma lucidum and its pharmaceutically active compounds.
Biotechnol Annu Rev. 13:265–301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jang KJ, Son IS, Shin DY, Yoon HM and Choi
YH: Anti-invasive activity of ethanol extracts of Ganoderma lucidum
through tightening of tight junctions and inhibition of matrix
metalloproteinase activities in human gastric carcinoma cells. J
Acupunct Meridian Stud. 4:225–235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jang KJ, Han MH, Lee BH, Kim BW, Kim CH,
Yoon HM and Choi YH: Induction of apoptosis by ethanol extracts of
Ganoderma lucidum in human gastric carcinoma cells. J Acupunct
Meridian Stud. 3:24–31. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuen JW, Gohel MD and Au DW:
Telomerase-associated apoptotic events by mushroom Ganoderma
lucidum on premalignant human urothelial cells. Nutr Cancer.
60:109–119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu HS, Yang XL, Wang LB, Zhao DX and Chen
L: Effects of extracts from sporoderm-broken spores of Ganoderma
lucidum on HeLa cells. Cell Biol Toxicol. 16:201–206. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hong KJ, Dunn DM, Shen CL and Pence BC:
Effects of Ganoderma lucidum on apoptotic and anti-inflammatory
function in HT-29 human colonic carcinoma cells. Phytother Res.
18:768–770. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin SB, Li CH, Lee SS and Kan LS:
Triterpene-enriched extracts from Ganoderma lucidum inhibit growth
of hepatoma cells via suppressing protein kinase C, activating
mitogen-activated protein kinases and G2-phase cell cycle arrest.
Life Sci. 72:2381–2390. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Min BS, Nakamura N, Miyashiro H, Bae KW
and Hattori M: Triterpenes from the spores of Ganoderma lucidum and
their inhibitory activity against HIV-1 protease. Chem Pharm Bull
(Tokyo). 46:1607–1612. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu QY, Jin YS, Zhang Q, Zhang Z, Heber D,
Go VL, Li FP and Rao JY: Ganoderma lucidum extracts inhibit growth
and induce actin polymerization in bladder cancer cells in vitro.
Cancer Lett. 216:9–20. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wynne S and Djakiew D: NSAID inhibition of
prostate cancer cell migration is mediated by Nag-1 induction via
the p38 MAPK-p75(NTR) pathway. Mol Cancer Res. 8:1656–1664. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang X, Baek SJ and Eling TE: The diverse
roles of nonsteroidal anti-inflammatory drug activated gene
(NAG-1/GDF15) in cancer. Biochem Pharmacol. 85:597–606. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Parekh HS, Liu G and Wei MQ: A new dawn
for the use of traditional Chinese medicine in cancer therapy. Mol
Cancer. 8:212009.https://doi.org/10.1201/b16611-11 View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alao JP: The regulation of cyclin D1
degradation: Roles in cancer development and the potential for
therapeutic invention. Mol Cancer. 6:242007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu Y, Bi T, Wang Z, Wu G, Qian L, Gao Q
and Shen G: Oxymatrine synergistically enhances antitumor activity
of oxaliplatin in colon carcinoma through PI3K/AKT/mTOR pathway.
Apoptosis. 21:1398–1407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gong J, Ardelt B, Traganos F and
Darzynkiewicz Z: Unscheduled expression of cyclin B1 and cyclin E
in several leukemic and solid tumor cell lines. Cancer Res.
54:4285–4288. 1994.PubMed/NCBI
|
|
30
|
Cho HJ and Park JH: Kaempferol induces
cell cycle arrest in HT-29 human colon cancer cells. J Cancer Prev.
18:257–263. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee DE, Lee KW, Jung SK, Lee EJ, Hwang JA,
Lim TG, Kim BY, Bode AM, Lee HJ and Dong Z:
6,7,4-trihydroxyisoflavone inhibits HCT-116 human colon cancer cell
proliferation by targeting CDK1 and CDK2. Carcinogenesis.
32:629–635. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Graña X and Reddy EP: Cell cycle control
in mammalian cells: Role of cyclins, cyclin dependent kinases
(CDKs), growth suppressor genes and cyclin-dependent kinase
inhibitors (CKIs). Oncogene. 11:211–219. 1995.PubMed/NCBI
|
|
33
|
Ruan W, Wei Y and Popovich DG: Distinct
responses of cytotoxic Ganoderma lucidum triterpenoids in human
carcinoma cells. Phytother Res. 29:1744–1752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang T, Xie ZP, Huang ZS, Li H, Wei AY, Di
JM, Xiao HJ, Zhang ZG, Cai LH, Tao X, et al: Total triterpenoids
from Ganoderma Lucidum suppresses prostate cancer cell growth by
inducing growth arrest and apoptosis. J Huazhong Univ Sci Technolog
Med Sci. 35:736–741. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kobayashi T, Sawa H, Morikawa J, Zhang W
and Shiku H: Bax induction activates apoptotic cascade via
mitochondrial cytochrome c release and Bax overexpression enhances
apoptosis induced by chemotherapeutic agents in DLD-1 colon cancer
cells. Jpn J Cancer Res. 91:1264–1268. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Paul-Samojedny MI, Kokocińska D, Samojedny
A, Mazurek U, Partyka R, Lorenz Z and Wilczok T: Expression of cell
survival/death genes: Bcl-2 and Bax at the rate of colon cancer
prognosis. Biochim Biophys Acta. 25-29:2005.
|
|
37
|
Shim M and Eling TE: Vitamin E succinate
induces NAG-1 expression in a p38 kinase-dependent mechanism. Mol
Cancer Ther. 7:961–971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen Y, Lv J, Li K, Xu J, Li M, Zhang W
and Pang X: Sporoderm-broken spores of Ganoderma lucidum inhibit
the growth of lung cancer: Involvement of the Akt/mTOR signaling
pathway. Nutr Cancer. 68:1151–1160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thyagarajan A, Jedinak A, Nguyen H, Terry
C, Baldridge LA, Jiang J and Sliva D: Triterpenes from Ganoderma
Lucidum induce autophagy in colon cancer through the inhibition of
p38 mitogen-activated kinase (p38 MAPK). Nutr Cancer. 62:630–640.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Alves C Castro, Rosivatz E, Schott C,
Hollweck R, Becker I, Sarbia M, Carneiro F and Becker KF: Slug is
overexpressed in gastric carcinomas and may act synergistically
with SIP1 and Snail in the down-regulation of E-cadherin. J Pathol.
211:507–515. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shin DH, Dier U, Melendez JA and Hempel N:
Regulation of MMP-1 expression in response to hypoxia is dependent
on the intracellular redox status of metastatic bladder cancer
cells. Biochim Biophys Acta. 1852:2593–2602. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Martínez-Montemayor MM, Acevedo RR,
Otero-Franqui E, Cubano LA and Dharmawardhane SF: Ganoderma lucidum
(Reishi) inhibits cancer cell growth and expression of key
molecules in inflammatory breast cancer. Nutr Cancer. 63:1085–1094.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu LY, Chen H, Liu C, Wang HQ, Kang J, Li
Y and Chen RY: Triterpenoids of Ganoderma theaecolum and their
hepatoprotective activities. Fitoterapia. 98:254–259. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shi Y, Sun J, He H, Guo H and Zhang S:
Hepatoprotective effects of Ganoderma lucidum peptides against
D-galactosamine-induced liver injury in mice. J Ethnopharmacol.
117:415–419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kwok Y, Ng KF, Li CC, Lam CC and Man RY: A
prospective, randomized, double-blind, placebo-controlled study of
the platelet and global hemostatic effects of Ganoderma lucidum
(Ling-Zhi) in healthy volunteers. Anesth Analg. 101:423–426, table
of contents. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Noguchi M, Kakuma T, Tomiyasu K, Yamada A,
Itoh K, Konishi F, Kumamoto S, Shimizu K, Kondo R and Matsuoka K:
Randomized clinical trial of an ethanol extract of Ganoderma
lucidum in men with lower urinary tract symptoms. Asian J Androl.
10:777–785. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wachtel-Galor S, Szeto YT, Tomlinson B and
Benzie IF: Ganoderma lucidum (‘Lingzhi’); acute and short-term
biomarker response to supplementation. Int J Food Sci Nutr.
55:75–83. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhuang SR, Chen SL, Tsai JH, Huang CC, Wu
TC, Liu WS, Tseng HC, Lee HS, Huang MC, Shane GT, et al: Effect of
citronellol and the Chinese medical herb complex on cellular
immunity of cancer patients receiving chemotherapy/radiotherapy.
Phytother Res. 23:785–790. 2009. View Article : Google Scholar : PubMed/NCBI
|