Introduction
Gastric cancer is one of the most prevalent types of
cancer worldwide (1). The prognosis
of gastric cancer is poor because the majority of patients are
diagnosed at the advanced stage (2), and the average 5-year survival rate of
advanced gastric cancer is 20–30%, despite a favorable prognosis
for early-stage gastric cancer, with an average 5-year survival
rate of >90% (3). Therefore, the
identification of molecular targets for developing novel treatment
is essential in gastric cancer.
MicroRNAs (miRNAs) are noncoding RNAs that consist
of 22–25 nucleotides, and modulate various cellular processes,
including cell growth, development, differentiation, metabolism and
apoptosis (4). Increasing evidence
supports a role of miRNAs as oncogenes or suppressor in solid
tumors (5). miR-181a is a member of
the miR-181 family that serves a role in the pathogenesis of
various cancer types (6–9). Particularly, it is reported to be
expressed at a high level in gastric cancer and may promote gastric
carcinogenesis (10,11). Inconsistent with these studies, Lin
et al (12) reported that
miR-181a inhibits cell proliferation, migration and metastasis, and
is downregulated in gastric cancer. Therefore, the function of
miR-181a in the pathogenesis of gastric cancer remains
controversial, and the exact molecular mechanisms by which miR-181a
modulate the process remain to be elucidated.
The Ras association domain family protein1 isoform A
(RASSF1A), encoded by the RASSF1A gene, is localized at chromosome
3p21.3 (13). In various cancer
types, including non-small cell lung and gastric cancer,
suppression of RASSF1A expression has been reported (14–16),
and RASSF1A therefore is theorized to function as a tumor
suppressor. Aberrant promoter methylation is the most common
molecular mechanism of silencing RASSF1A (17,18).
Furthermore, miRNAs, including miR-602 and miR-181a/b, have been
demonstrated to target and downregulate RASSF1A in hepatocellular
carcinoma and acute promyelocytic leukemia (16,19).
This suggests that miRNA-mediated suppression of RASSF1A may serve
an essential role in the carcinogenesis and cancer progression. The
present study aimed to investigate the interaction between miR-181a
and RASSF1A, and their respective roles in gastric cancer.
Materials and methods
Clinical samples and cell
cultures
A total of 42 pairs of gastric cancer samples and
adjacent non-cancer tissue samples (5 cm away from the tumor) were
collected from patients (31 males and 11 females; aged 40–78 years
old) who had undergone surgery for primary gastric cancer at The
First Affiliated Hospital of Xi'an Jiaotong University (Xi'an,
China) between March 2014 and July 2014. No patient had received
preoperative radiotherapy or chemotherapy. Written informed consent
was obtained from all patients, and the study protocol was approved
by the Ethics Committee of The First Affiliated Hospital of Xi'an
Jiaotong University (Xi'an, China).
AGS, SGC-7901 and 293 cells were purchased from the
Shanghai Institute of Cell Biology, Chinese Academy of Sciences
(Shanghai, China). Cells were maintained at 37°C in RPMI-1640
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented
with 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences,
Logan, UT, USA) in a humidified incubator with 5%
CO2.
Cell transfection
The miR-181a mimics, negative control (NC), miR-181a
inhibitor, inhibitor NC, siRNA-RASSF1A and siRNA-NC were
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The
sequences were as follows: miR-181a mimic,
5′-AACAUUCAACGCUGUCGGUGAGUUCACCGACAGCG-3′; miR-181a inhibitor,
5′-ACUCACCGACAGCGUUGAAUGUU-3′; siRNA-RASSF1A forward,
5′-GACCUCUGUGGCGACUU-3′ and reverse, 5′-UGAAGUCGCCACAGAG-3′; NC and
siRNA-NC forward, 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′; inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAA-3′. For RNA delivery, cells were seeded at
a density of 1×105 cells/well in 6-well plates, and
Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
was used to transfect the cells with 100 nM miR-181a mimic or NC,
200 nM miR-181a inhibitor or inhibitor NC, and 50 nM siRNA-RASSF1A,
following the manufacturer's protocol. Each experiment was repeated
three times.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract total RNA from the cell lines
and tissue samples, following the manufacturer's protocol. The
RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.) was used to convert the RNA into cDNA with the
following temperature protocol: 25°C for 5 min, followed by 42°C
for 60 min and 70°C for 5 min. The following primers were used for
qPCR: RASSF1A forward, 5′-AGTGCGCGCATTGCAAGTT-3′ and reverse,
5′-AAGGTCAGGTGTCTCCCAC-3′; miR-181a forward,
5′-ACACTCCAGCTGGGAACATTCAACGCTGTCG-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGACTCACCG-3; RNU6B forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The primers of miR-181a and RNU6B were purchased from Guangzhou
RiboBio Co., Ltd. (Guangzhou, China). For the chain reaction, 2 µl
of cDNA, 1 µl forward primer and 1 µl reverse primer were mixed in
SYBR Premix Ex Taq II (Takara Biotechnology Co., Ltd., Dalian,
China) reagent. RT-qPCR was performed with a CFX96™ Real-Time PCR
Detection system (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
using SYBR Premix Ex Taq II at 95°C for 10 min, followed by 40
cycles of 95°C for 10 sec and 60°C for 1 min. The specific mRNA
expression level was quantified by using the 2−∆∆Cq
method (20). Each experiment was
repeated three times.
miR-181a target prediction
The miR-181a sequence was obtained from miRBase
(http://www.microrna.sanger.ac.uk). The
target genes were predicted using bioinformatics analysis tools,
including TargetScan (http://www.targetscan.org/vert_72/), miRBase
(http://www.mirbase.org/) and PICTA (https://pictar.mdc-berlin.de/).
Luciferase reporter assay
The 795 bp RASSF1A 3′-untranslated region (UTR)
fragment was amplified by qPCR from the DNA of AGS cells and
inserted into the pGL3-control vector (Promega Corporation,
Madison, WI, USA). The following primers were used: RASSF1A 3′-UTR
forward, 5′-GTCTAGACCTCTTGTACCCCAGGTGG-3′ and reverse,
5′-GTCTAGAGAGGATCTTGAAATCTTTATTGAG-3′. Mutagenesis of the miR-181a
binding site was performed using a Site-directed Mutagenesis kit
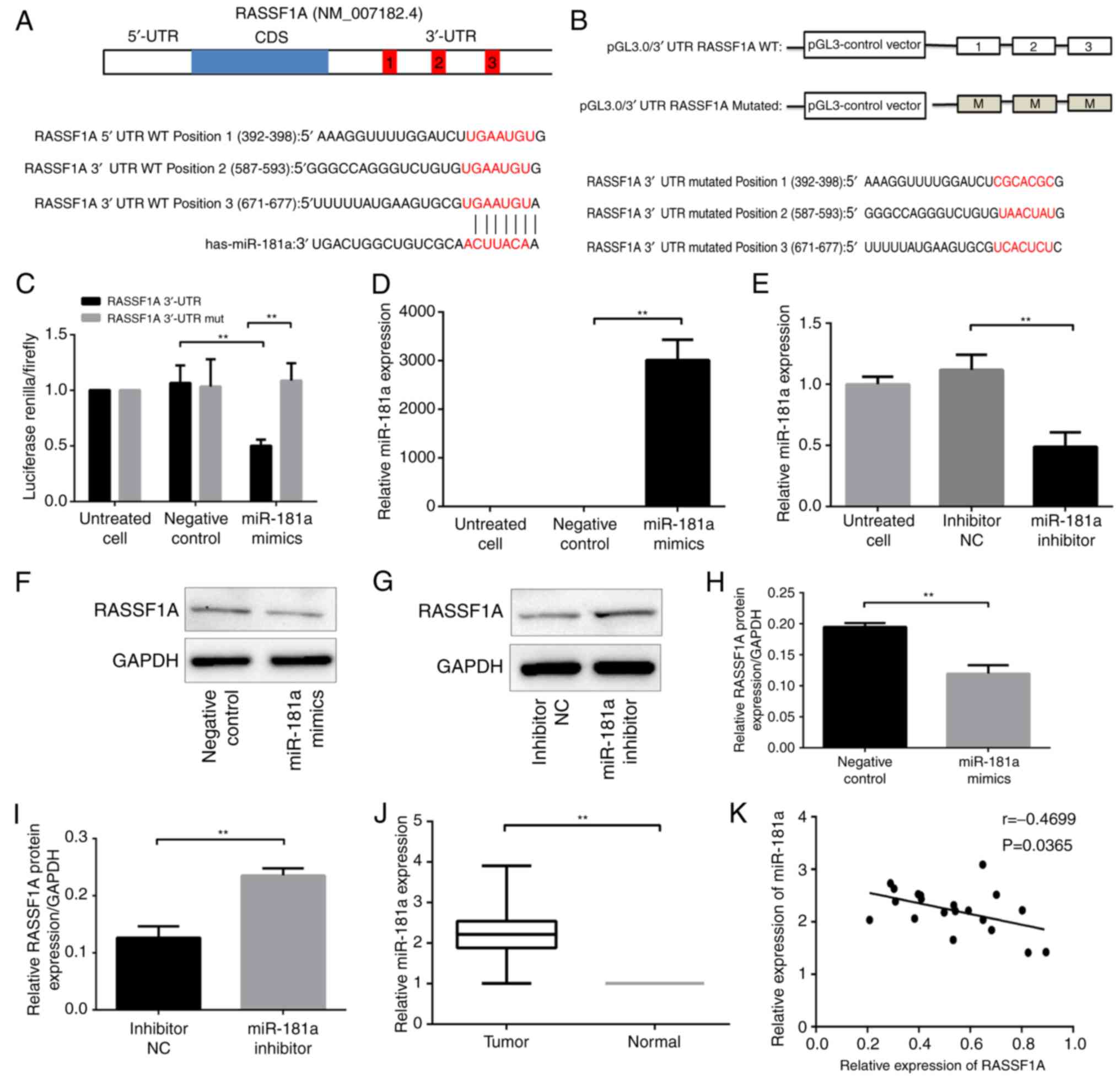
(Takara Biotechnology Co., Ltd.) (16) (Fig. 1A
and B). For Luciferase reporter assay, 239 cells were
transiently cotransfected with 0.5 µg pGL3-RASSF1A 3′UTR or
pGL3-RASSF1A 3′UTR mutated, and 50 nM miR-181a mimics or NC using
Lipofectamine 2000. Then, 24 h after transfection, Firefly and
Renilla luciferase activity were measured using the
Dual-Luciferase Reporter assay system, following the manufacturer's
protocol (Promega Corporation). Each experiment was repeated three
times.
Cell growth assay
SGC-7901 and AGS cells were transfected with
oligonucleotides as aforementioned. Cells were seeded into 96-well
culture plates (6×103 cells/well) in 100 µl RPMI-1640
medium containing 10% FBS. Cell growth was assessed using a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) following the manufacturer's protocol at 24, 48,
72 and 96 h. Each experiment was repeated three times.
Cell cycle and apoptosis assay
Following transfection for 48 h, SGC-7901 and AGS
cells were collected by trypsinization and washed with PBS twice.
Cycle analyses were performed with the following protocol: Cells
were fixed in 75% cold ethanol at 4°C overnight. Subsequently, the
cells were treated with RNase A for 30 min at 37°C and stained with
propidium iodide (PI) in the dark at room temperature for 30 min.
For apoptosis analysis, cells were labeled with Annexin V-FITC and
PI (BD Biosciences, Franklin Lakes, NJ, USA) according to the
manufacturer's protocol. Cell cycle and apoptosis were assessed
using FACS Calibur flow cytometry (BD Biosciences), and the data
were analyzed using Flowjo 10.0 software (FlowJo LLC, Ashland, OR,
USA). Each experiment was repeated three times.
Protein extraction and western blot
analysis
Fresh tissues and cells were lysed using RIPA buffer
(Beyotime Institute of Biotechnology, Shanghai, China) containing a
protease inhibitor cocktail (Roche Diagnostics GmbH, Mannheim,
Germany) and were quantified using a bicinchoninic acid assay
(Pierce; Thermo Fisher Scientific, Inc.). Tissue or cell lysates
containing 30 µg total protein/lane were loaded and separated on a
10% SDS-PAGE gel, followed by transference to polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA). The
membranes were blocked with 5% fat-free dry milk at room
temperature for 1 h and then incubated with appropriate primary
antibodies at 1:1,000 dilution at 4°C overnight. The primary
antibodies against human cell division cycle 25A (CDC25A; mouse
IgG; cat. no. sc-70823), p21 (mouse IgG, cat. no. sc-71811),
Bcl-2-associated X protein (BAX; mouse IgG; cat. no. sc-20067),
B-cell lymphoma 2 (Bcl-2; mouse IgG; cat. no. sc-509) and GAPDH
(mouse IgG; cat. no. sc-47724) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Anti-cyclin A2 (mouse IgG;
cat. no. 4656) and cyclin D1 (rabbit IgG; cat. no. 2922) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Anti-RASSF1A (mouse IgG; cat. no. eB114-10H1) was purchased from
Thermo Fisher Scientific, Inc. (eBioscience). The membrane was
washed with TBS-Tween-20 buffer at room temperature three times,
each time for 10 min, following incubation with the primary
antibodies, and subsequently incubated with goat anti-rabbit
horseradish peroxidase-conjugated secondary antibody (Abgent, San
Diego, CA, USA) at room temperature for 2 h. To visualize the
protein bands, SuperSignal West Femto Maximum Sensitivity substrate
(Pierce; Thermo Fisher Scientific, Inc.) was added. Protein
expression was quantified by ImageJ software version 1.46 (National
Institutes of Health; Bethesda, MD, USA). Each experiment was
repeated three times.
Statistical analysis
All data presented are mean ± standard deviation
unless otherwise noted. A Student's t-test or one-way analysis of
variance with Duunett's post hoc test were used to detect
statistical differences between groups. The correlation between
RASSF1A and miR-181a expression was analyzed using the Pearson's
correlation test. The association between the miR-181a level and
clinicopathological characteristics in gastric cancer tissues was
analyzed using a Student's t-test or one-way analysis of variance
with the Least significant difference post hoc test. All
statistical analyses were performed using SPSS 18.0 software (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Identifying RASSF1A as a direct target
of miR-181a
In order to investigate the role of miR-181a in
gastric carcinogenesis, search for its putative target genes with
bioinformatics analysis tools, including TargetScan, miRBase and
PICTAR was performed. RASSF1A was identified as a possible target
(Fig. 1A). The interaction between
miR-181a and RASSF1A was then assessed. The possible binding sites
for miR-181a in the 3′-UTR of RASSF1A was mutated, and luciferase
reporter constructs with the mutated sequence and the wild type
sequence were constructed (Fig.
1B). The mutant or wild-type RASSF1A construct was
co-transfected with miR-181a mimics in 293 cells, and the
luciferase activity of the RASSF1A 3′-UTR constructs was measured
with the dual luciferase reporter assay. Luciferase activity in
cells that were transfected with wild-type constructs and miR-181a
mimics was significantly suppressed compared with cells transfected
with wild-type constructs and NC (P<0.01; Fig. 1C). In contrast, cells co-transfected
with the mutant constructs and miR-181a mimics exhibited a recovery
of luciferase activity (P<0.01; Fig.
1C). These results indicated a direct interaction between
miR-181a and RASSF1A at the 3′-UTR binding sites.
The effects of miR-181a on RASSF1A protein
expression was then explored. The miR-181a expression in AGS cells
transfected with miR-181a mimics was significantly higher compared
with that in cells transfected with negative control (P<0.01;
Fig. 1D); whereas the miR-181a
expression in SCG-7901 cells transfected with miR-181a inhibitor
was lower compared with that in cells transfected with inhibitor NC
(P<0.01; Fig. 1E). RASSF1A
protein level was significantly lower in AGS cells transfected with
miR-181a mimics compared with that in cells transfected with
negative control (P<0.01; Fig. 1F
and H). Conversely, the RASSF1A protein level was significantly
higher in SGC-7901 cells transfected with miR-181a inhibitor
compared with that in cells transfected with inhibitor NC
(P<0.01; Fig. 1G and I).
Therefore, miR-181a level was negatively associated with the
RASSF1A protein level. Notably, miR-181a did not significantly
affect RASSF1A mRNA level in AGS and SGC-7901 cells as assessed by
qPCR analyses (data not shown), indicating that miR-181a directly
suppressed RASSF1A expression at the translational level.
RASSF1A protein expression negatively
correlates with miR-181a level in gastric cancer tissue
To investigate whether the regulation of RASSF1A
level by miR-181a had clinical implications, miR-181a and RASSF1A
levels in tumor tissues and adjacent normal tissues obtained from
patients with gastric cancer were examined.
The relative level of miR-181a in 42 paired tumor
and normal tissues was measured, and it was demonstrated that the
miR-181a level was significantly higher in tumor tissues compared
with that in adjacent normal tissue (P<0.01; Fig. 1J). Further analyses did not identify
a correlation between the miR-181a level and patient age, sex,
tumor size, the locations of tumor, cell differentiation, the tumor
node metastasis (TNM) stage or the depth of tumor invasion
(Table I).
 | Table I.Correlation between miR-181a level
and clinicopathological characteristics in gastric cancer
tissues. |
Table I.
Correlation between miR-181a level
and clinicopathological characteristics in gastric cancer
tissues.
| Clinicopathological
parameters | Patients
(n=42) | Relative
expression | Test
statistics | P-value |
|---|
| Sex |
|
| t=0.854 | 0.398 |
|
Male | 31 | 2.320±1.643 |
|
|
|
Female | 11 | 1.861±1.129 |
|
|
| Age at diagnosis,
years |
|
| t=−0.967 | 0.340 |
|
<60 | 20 | 1.961±1.026 |
|
|
|
≥60 | 22 | 2.417±1.868 |
|
|
| T stage |
|
| t=−0.984 | 0.331 |
|
T1+T2 | 5 | 1.569±0.801 |
|
|
|
T3+T4 | 37 | 2.285±1.587 |
|
|
| Lymphatic
metastasis |
|
| t=−1.706 | 0.096 |
| No | 12 | 1.579±0.876 |
|
|
|
Yes | 30 | 2.448±1.667 |
|
|
| M stage |
|
| t=−1.178 | 0.246 |
| M0 | 35 | 1.583±0.587 |
|
|
| M1 | 7 | 2.323±1.629 |
|
|
| TNM stage |
|
| t=−1.641 | 0.109 |
|
I+II | 9 | 1.475±0.508 |
|
|
|
III+IV | 33 | 2.398±1.652 |
|
|
| Tumor location |
|
| F=3.188 | 0.052 |
|
Proximal gastric | 17 | 2.873±1.907 |
|
|
| Gastric
body | 8 | 1.939±0.921 |
|
|
| Distal
gastric | 17 | 1.649±1.058 |
|
|
| Tumor size, cm |
|
| t=0.500 | 0.620 |
|
>5 | 20 | 2.324±1.226 |
|
|
| ≤5 | 22 | 2.087±1.777 |
|
|
|
Differentiation |
|
| t=−1.215 | 0.233 |
|
Low/Poorly | 17 | 1.936±1.196 |
|
|
|
Moderate/high | 18 | 2.606±1.955 |
|
|
| Pathological
type |
|
| F=0.286 | 0.753 |
|
Adenocarcinoma | 35 | 2.280±1.643 |
|
|
|
Mucinous carcinoma | 5 | 1.769±0.603 |
|
|
|
Neuroendocrine carcinoma | 2 | 1.867±0.936 |
|
|
miR-181a expression was then assessed with qPCR and
RASSF1A protein levels with western blotting in 10 paired tumor and
normal tissues. Furthermore, correlation analysis between miR-181a
and RASSF1A expression levels was performed. A moderate correlation
between the miR-181a level and RASSF1A protein level identified
(P=0.0365; Fig. 1K).
RASSF1A knockdown promotes cell
proliferation and G1/S transition, and inhibits apoptosis in AGS
cells
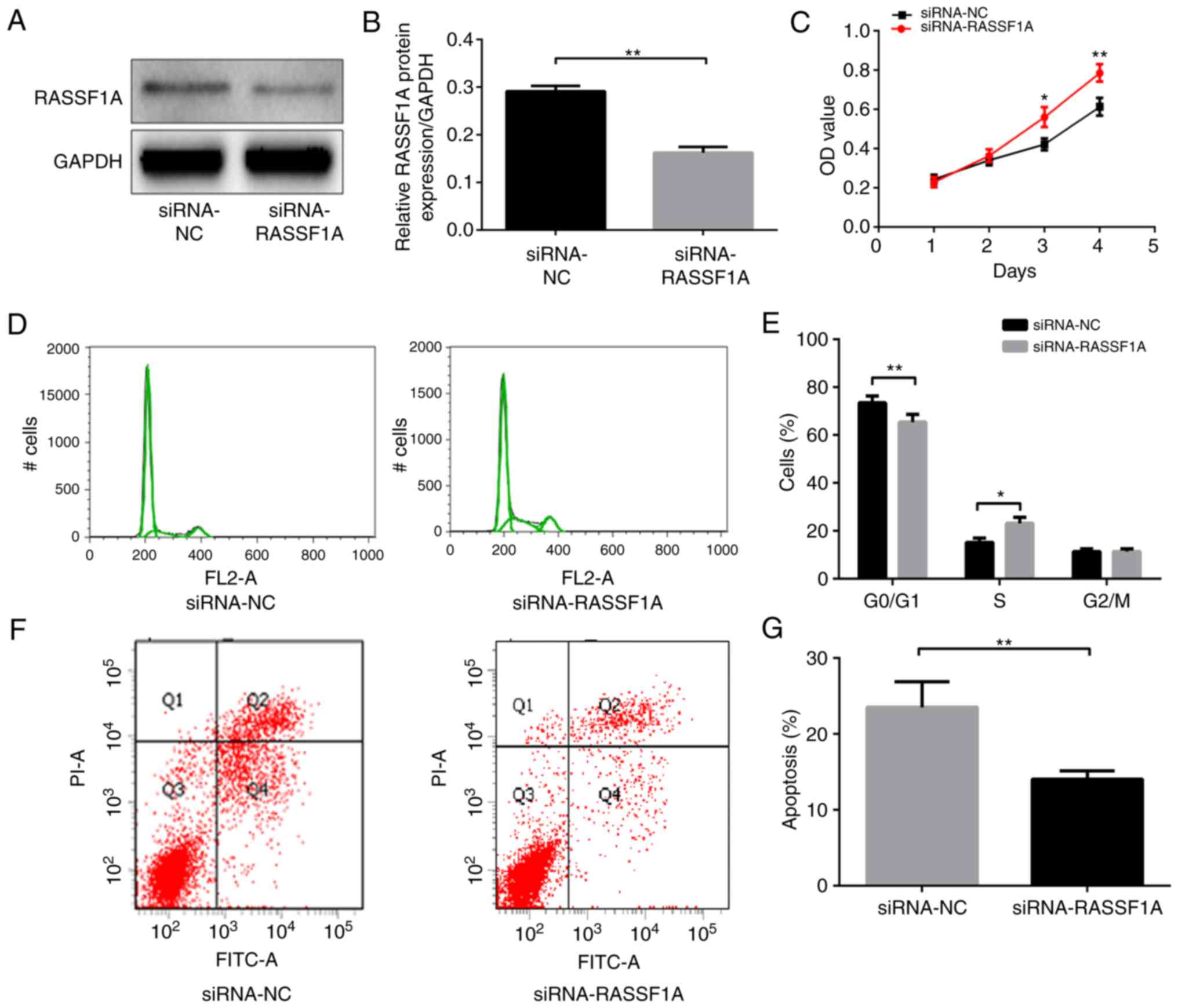
Whether RASSF1A activities have a role in gastric
carcinogenesis was investigated. Cell growth, cell cycle and
apoptosis were examined in AGS cells transfected with siRNA that
targeted RASSF1A. The RASSF1A protein expression level was
significantly lower in AGS cells transfected with siRNA-RASSF1A
compared with that in cells transfected with siRNA-NC (P<0.01;
Fig. 2A and B). Knockdown of
RASSF1A with siRNA significantly enhanced cell growth in AGS cells
as assessed using the CCK-8 assay (P<0.01; Fig. 2C). In addition, the
siRNA-RASSF1A-transfected cells contained a significantly lower
proportion of cells in the G0/G1 phase and a significantly higher
proportion of cells in the S phase compared with the siRNA-NC
transfected cells (P<0.01 and P<0.05, respectively; Fig. 2D and E), whereas the percentages of
cells in the G2/M phase did not differ significantly between the
two groups (P>0.05).
Lastly, the apoptotic rates of AGS cells were
assessed with cell flow cytometry. The apoptotic rate of cells
transfected with siRNA-RASSF1A was significantly lower compared
with that of the cells transfected with siRNA-NC (P<0.01;
Fig. 2F and G). Therefore, the
aforementioned results suggest that the knockdown of RASSF1A
promoted cell proliferation and reduced apoptosis in AGS cells.
Effects of miR-181a on gastric cancer
cells
As indicated by the aforementioned results, miR-181a
negatively modulated RASSF1A expression, and RASSF1A suppressed
cell proliferation, but promoted apoptosis. Thus, the effects of
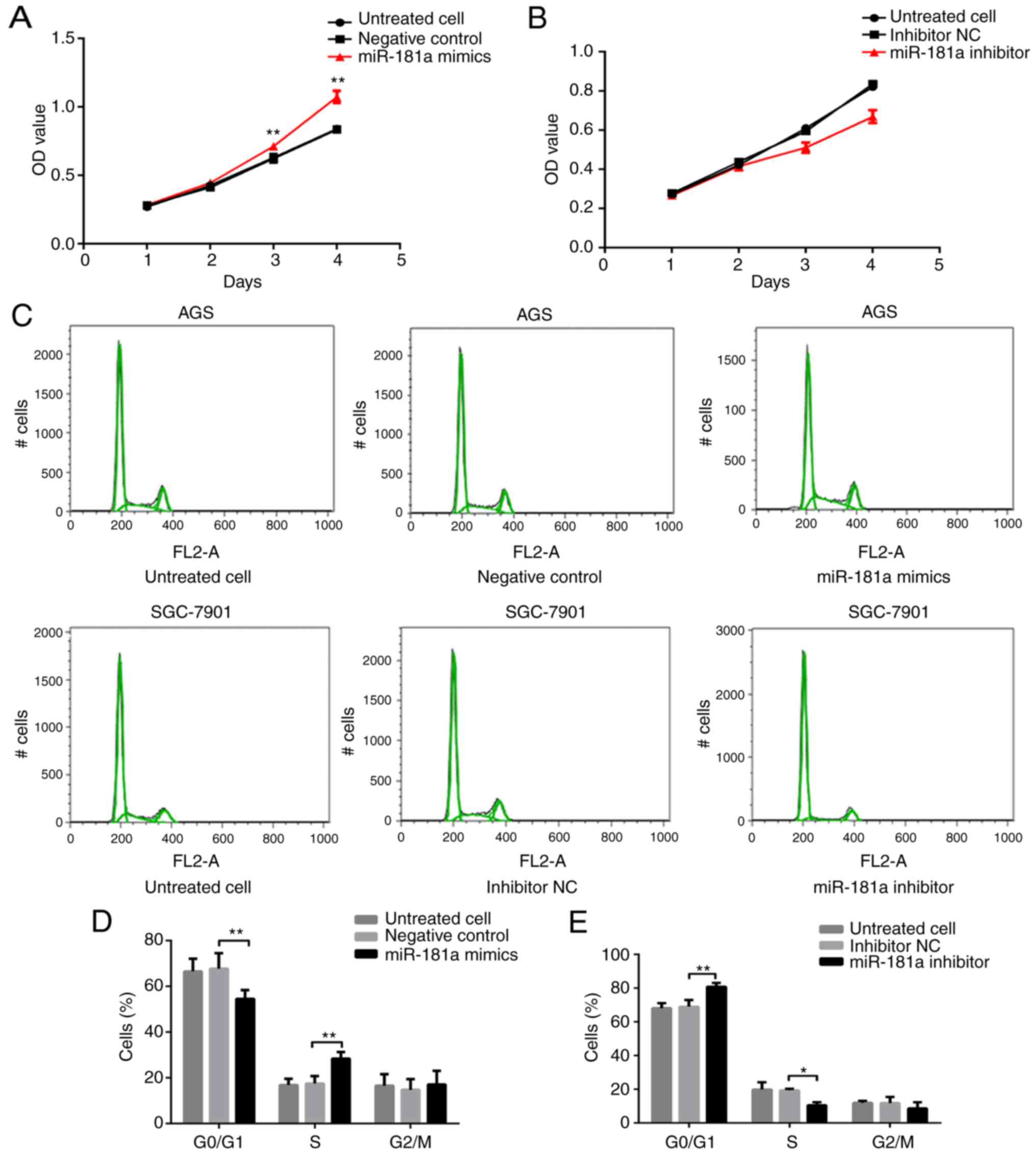
miR-181a on gastric cancer cell lines were examined next. Cell
growth was assessed with a CCK-8 assay. AGS cells transfected with
miR-181a mimics exhibited significantly greater growth rates
compared with cells transfected with negative control (P<0.01;
Fig. 3A). Conversely, SGC-7901
cells transfected with miR-181a inhibitor demonstrated
significantly less growth compared with cells transfected with
inhibitor NC (P<0.01; Fig.
3B).
The percentage of cells in each cell cycle phase was
measured. The percentage of cells in the G0/G1 phase was
significantly lower, while the percentage of cells in the S phase
was significantly higher in the miR-181a mimics-transfected AGS
cells compared with that in cells transfected with negative control
(both P<0.01; Fig. 3C and D).
The percentage of cells in the G2/M phase did not differ
significantly among the three groups (P>0.05; Fig. 3C and D). Transfection of miR-181a
inhibitor revealed the opposite effects, where the percentage of
cells in the G0/G1 phase was significantly higher, and in the S
phase significantly lower in SGC-7901 cells transfected with
miR-181a inhibitor compared with cells treated with inhibitor NC
(P<0.01 and P<0.05, respectively; Fig. 3C and E). The percentage of cells in
the G2/M phase; however, was not significantly different among the
three groups of cells (P>0.05; Fig.
3C and E).
Lastly, the apoptotic rate of cells was assessed
with cell flow cytometry (Fig. 3F).
The apoptotic rate in AGS cells transfected with miR-181a mimic was
significantly lower compared with that in cells transfected with
negative control (P<0.01; Fig. 3F
and G). In contrast, the apoptotic rate was significantly
higher in SGC-7901 cells transfected with miR-181a inhibitor
compared with that in cells transfected with the inhibitor NC
(P<0.01; Fig. 3F and H).
Taken together, these data indicated that miR-181a
and RASSF1A exhibit opposing effects on cell proliferation and
apoptosis.
RASSF1A knockdown attenuates the
effects of miR-181a downregulation on the proliferation and
apoptosis of SGC-7901 cells
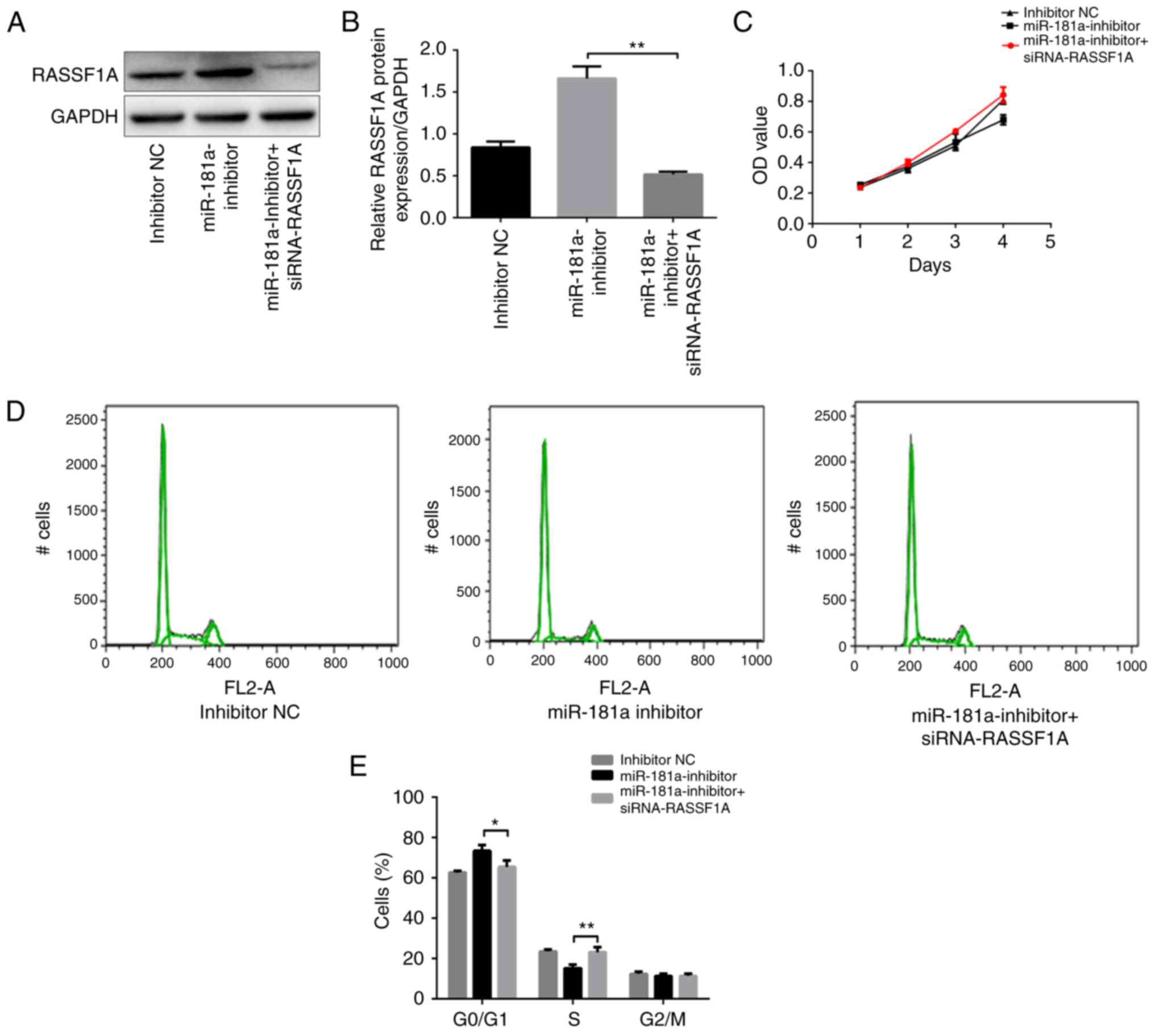
Whether RASSF1A knockdown attenuated the effect of
miR-181a downregulation on proliferation and apoptosis of SGC-7901
cells was investigated. SGC-7901 cells were co-transfected with
miR-181a inhibitor and siRNA-RASSF1A. Western blot analysis was
used to confirm that RASSF1A expression was downregulated by
siRNA-RASSF1A in SGC-7901 cells transfected with miR-181a inhibitor
(P<0.01; Fig. 4A and B). Next,
cell growth was assessed using a CCK-8 assay. The growth of
SGC-7901 cells co-transfected with miR-181a inhibitor and
siRNA-RASSF1A was significantly increased compared with cells
transfected with miR-181a inhibitor only (P<0.01; Fig. 4C). Furthermore, the percentage of
cells in the G0/G1 phase was significantly lower, while the
percentage of cells in the S phase was significantly higher in the
cells co-transfected with miR-181a inhibitor and siRNA-RASSF1A
compared with cells transfected with miR-181a inhibitor only
(P<0.05 and P<0.01, respectively; Fig. 4D and E). The percentage of cells in
the G2/M phase did not differ significantly between the two groups
(P>0.05; Fig. 4D and E). These
results indicated that the inhibitory effects of downregulation of
miR-181a on proliferation in SGC-7901 cells were attenuated by
RASSF1A knockdown.
The apoptotic rate in cells co-transfected with
miR-181a inhibitor and siRNA-RASSF1A was significantly lower
compared with that in cells transfected with miR-181a inhibitor
only (P<0.01; Fig. 4F and G).
Taken together, these data suggested that RASSF1A knockdown
attenuated the effects of miR-181a downregulation on cell
proliferation and apoptosis.
Effects of miR-181a on the expression
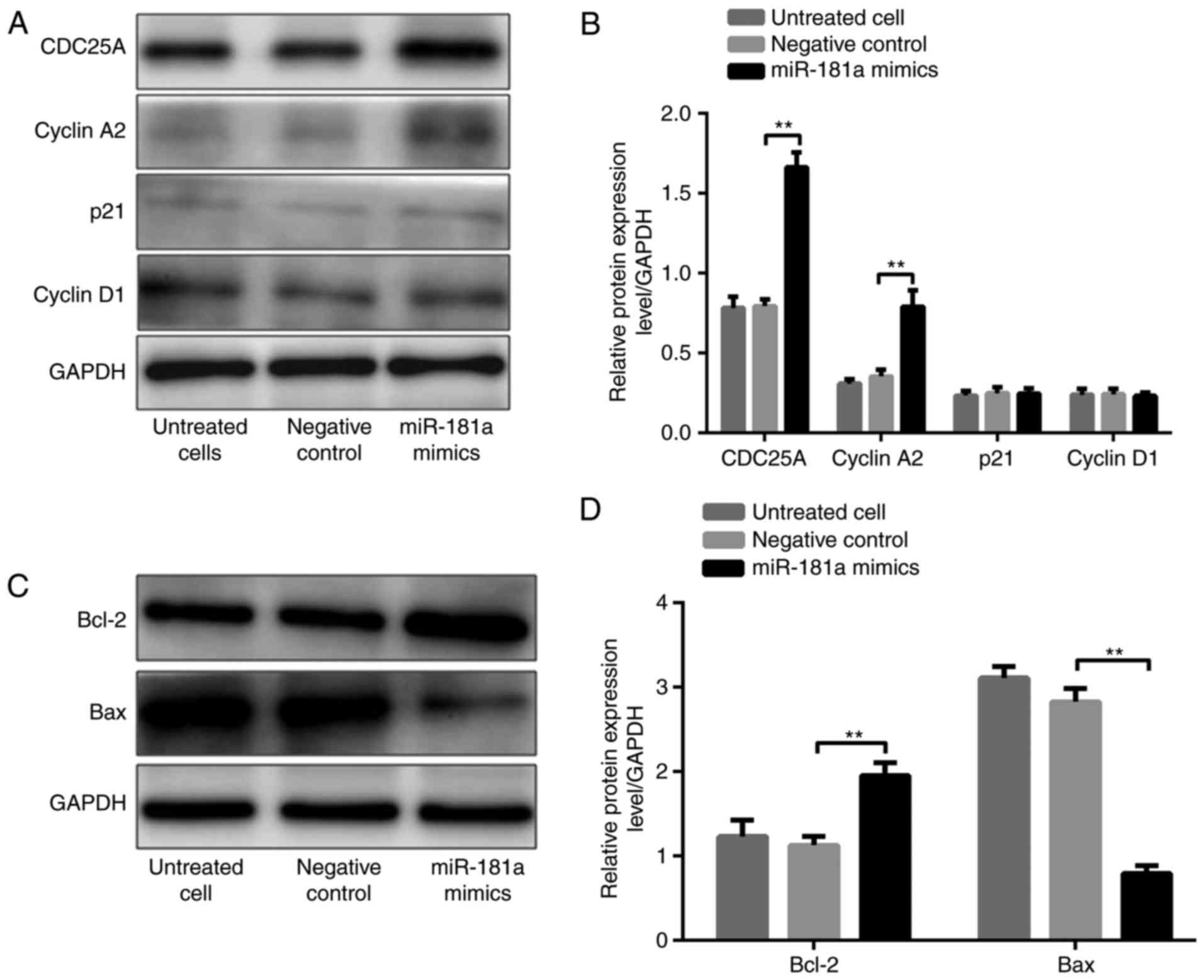
of tumorigenesis proteins
To further investigate the role of miR-181a in
gastric cancer, the effects of miR-181a on the expression of
tumorigenesis-associated proteins, including CDC25A, cyclin A2,
p21, cyclin D1, Bcl-2, and Bax, were investigated.
It was demonstrated that CDC25A, cyclin A2 and Bcl-2
protein levels were significantly higher, whereas the Bax protein
level was significantly lower, in AGS cells transfected with
miR-181a mimic compared with these in cells transfected with the
negative control (all P<0.01; Fig.
5A-D). P21 and cyclin D1 levels were not significantly
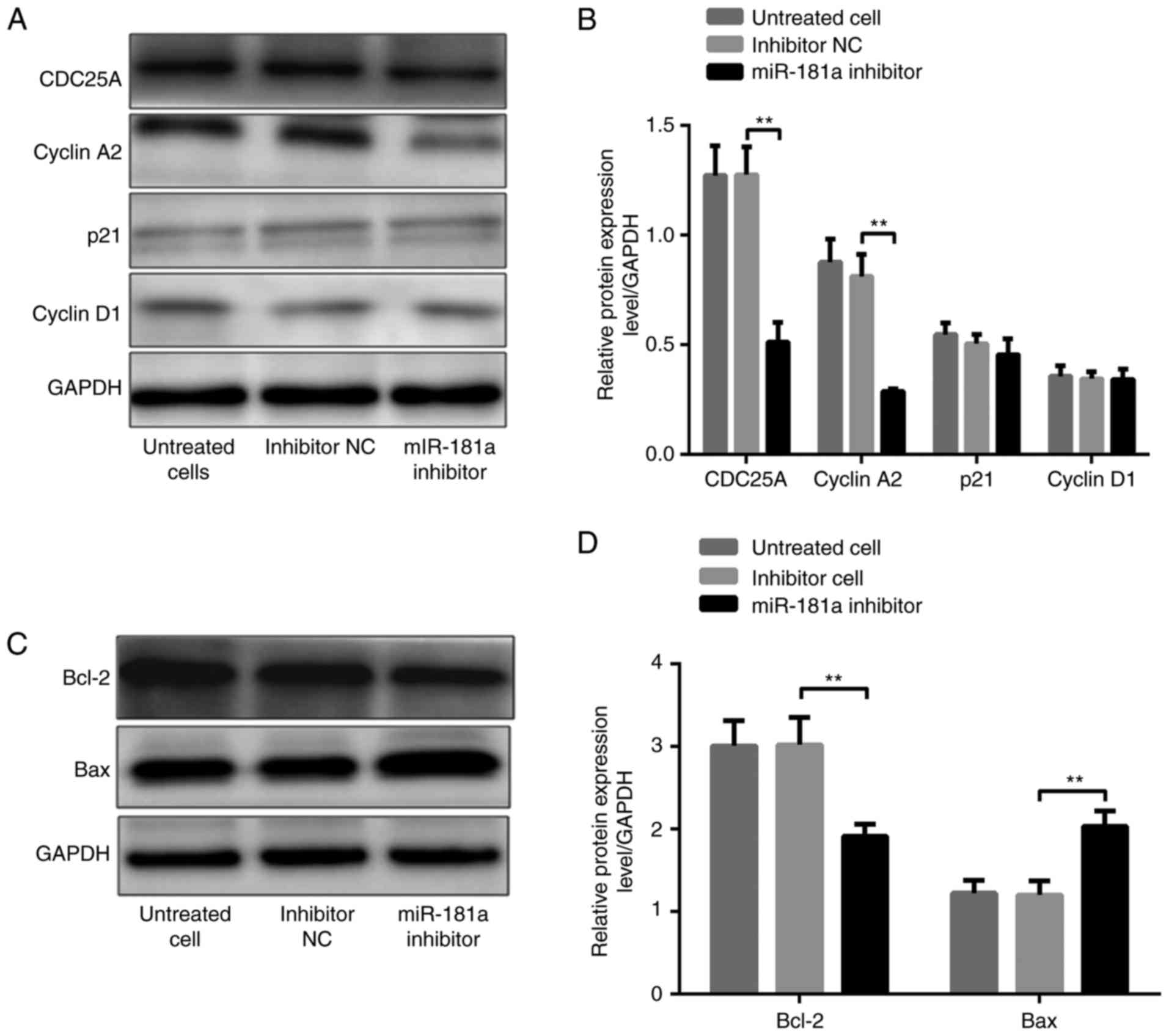
different in the three groups (both P>0.05; Fig. 5A and B). In contrast, CDC25A, cyclin
A2 and Bcl-2 protein levels were significantly lower, whereas the
Bax protein level was significantly higher, in SGC-7901 cells
transfected with miR-181a inhibitors compared with these in cells
transfected with inhibitor NC (all P<0.01; Fig. 6A-D). Similarly, p21 and cyclin D1
levels were not significantly different among the three groups
(both P>0.05; Fig. 6A and
B).
These results indicated the complexity of the
miR-181a signaling pathway, and that miR-181a targeted multiple
downstream effectors, which in turn serve essential roles in
tumorigenesis.
Discussion
In previous years, abnormalities in miR-181a
expression were reported to serve an essential role in the
pathogenesis of numerous types of cancer (8,9,21–23).
However, the exact role of miR-181a in tumorigenesis has remained
controversial due to inconsistent results in different tumor types.
miR-181a is reported to be an oncogene in head and neck cancer
(21), breast cancer (9), and hepatocellular carcinoma (8). In contrast, several studies indicated
miR-181a acts as a tumor-suppressor in primary glioblastoma
(22) and aggressive chronic
lymphocytic leukemia (23). These
findings demonstrate that the role of miR-181a in tumorigenesis is
tumor-specific. The result of the present study revealed elevated
miR-181a expression in gastric cancer tissues, indicating that
miR-181a may be an oncogene in gastric cancer.
Identifying cancer-specific miRNA targets is an
important step in clarifying the roles of miRNAs in tumorigenesis
and progression. miR-181a has been reported to inhibit tumor growth
via the downregulation of oncogene K-ras in oral squamous cell
carcinoma (21). Liu et al
(24) reported that miR-181a
promotes the transition of G0/G1 to S and cell growth via targeting
tumor suppressor ataxia telangiectasia mutated in pediatric acute
myeloid leukemia. In the present study, RASSF1A was identified as a
direct target of miR-181a, and the luciferase activity was lower in
293 cells co-transfected with wild-type constructs and miR-181a
mimics compared with cells co-transfected with wild-type constructs
and NC. Mutation of the putative binding sites in the 3′-UTR of
RASSF1A abolished these effects, suggesting that miR-181a directly
binds with the 3′-UTR of RASSF1A, thereby suppressing its
translation. In addition, in the gastric cancer cell lines, an
increase in miR-181a level was associated with reduced RASSF1A
protein levels, whereas a decrease in miR-181a level was associated
with an increased RASSF1A protein level. Notably, miR-181a did not
significantly affect RASSF1A mRNA expression in the AGS and
SGC-7901 cell lines, indicating that miR-181a directly suppresses
RASSF1A expression at the translational level. Furthermore, in
tumor tissues from patients with gastric cancer, a negative
correlation between the miR-181a level and RASSF1A protein level
was identified.
miRNAs have emerged as an essential modulators of
cell proliferation, apoptosis and cell cycle dysregulation
(25). RASSF1A was observed to
suppress gastric cancer cell proliferation and G1/S transition, and
promote apoptosis in the present study. Thus, the downregulation of
RASSF1A by miR-181a would expect to cause opposing effects. Indeed,
the results demonstrated that miR-181a promoted cell growth and
G1/S transition, and suppressed apoptosis in gastric cancer cell
lines. Furthermore, RASSF1A knockdown attenuated the effects of
downregulation of miR-181a on proliferation and apoptosis in
SGC-7901 cells.
Increasing evidence suggests that RASSF1A acts as a
tumor suppressor in numerous types of cancer through multiple
mechanisms (26,27). The results of the present study
suggest that RASSF1A promoted apoptosis, and suppressed cell growth
and proliferation, which are consistent with a previous study
whereby RASSF1A was demonstrated to suppress cell cycle progression
at the G1/S transition by preventing cyclin D1 accumulation
(28). Oh et al (29) reported that RASSF1A is required for
full activation of macrophage stimulating 1 (Mst1) and enhanced
Mst1-mediated apoptosis in vivo. Furthermore, in a previous
study, we reported that RASSF1A inhibits SGC-7901 cell invasion
under hypoxic conditions, which is associated with matrix
metalloproteinase-2 inhibition (30). Inactivation of RASSF1A may result
from multiple mechanisms in tumorigenesis, including
transcriptional silencing through promoter hypermethylation, loss
of heterozygosity and chromosome deficiency (31). The present study reported that
miR-181a suppressed RASSF1A by directly interacting with its 3′-UTR
region, resulting in downregulation of RASSF1A at the translational
level in gastric cancer cell lines. Therefore, it is likely that
miR-181a promotes gastric cancer progression by suppressing
RASSF1A.
In addition, it was demonstrated that miR-181a was
negatively associated with the Bax protein level, and positively
associated CDC25A, cyclin A2 and Bcl-2 protein levels in gastric
cancer cell lines. CDC25A, a member of the CDC25 family, serves an
important role in regulating the G1/S checkpoint and the G2/M
checkpoint (32). The increase in
the G1/S transition in the presence of miR-181a may be mediated by
upregulating CDC25A. In addition, CDC25A has been reported to be
involved in the hyperactivation of cyclin A2-cyclin-dependent
kinase 1 in the G0/G1 to S transition. The present data revealed
that cyclin A2 protein expression increased in cells overexpressing
miR-181a, suggesting that upregulation of CDC25A and cyclin A2
mediated by miR-181a results in the promotion of gastric cancer
cell proliferation and the G1/S transition. Similarly, it was
demonstrated that the expression of anti-apoptosis protein Bcl-2
and proapoptosis protein Bax expression was regulated by miR-181a,
consistent with a study by Xu et al (33) reporting similar effects of miR-181a
in cervical cancer. However, the exact mechanisms of miR-181a on
gastric cancer cell growth and apoptosis remain to be elucidated.
It is likely that miR-181a promotes gastric cancer progression via
multiple signaling pathways.
miR-181a as a predictor of prognosis in other
malignancies has been reported. Pichler et al (34) reported that the level of miR-181a
expression level is associated with poor survival of patients with
colorectal cancer. Xiang et al (35) reported that higher miR-181a
expression was associated with shorter recurrence-free survival and
shorter overall survival times in esophageal cancer. However, no
significant association was identified between the miR-181a
expression and clinical features of gastric cancer in the present
study. This inconsistency may be due to the TNM stage of patients
included in the current study, whereby the majority of patients
were at stage of TNM III and IV. Furthermore, the limited available
tissue samples did not allow for reliable correlation analyses to
be performed. miR-181a as the prognosis predictor for gastric
cancer remains to be validated, as follow-up visits have not
reached the universal standard of 3 or 5 years.
Several targets of miR-181a that are associated with
cell growth, apoptosis and invasion in a variety of human tumors
have been identified, including K-ras (21), caudal type homeobox 2 (36), GATA binding protein 6 (36) and nemo-like kinase (36). The present study results suggested
that RASSF1A is a direct target of miR-181a in gastric cancer, and
that RASSF1A is highly likely a tumor suppressor in gastric cancer.
Previous studies have reported that protein tyrosine phosphatase
MEG2 (37) and autophagy-related 5
(38) are directly targeted by
miR-181a in gastric cancer. In conclusion, the results of the
present study suggest that the upregulation of miR-181a promoted
gastric cancer cell growth and the G1/S transition, and inhibited
apoptosis, possibly through upregulation of CDC25A, cyclin A2 and
Bcl-2, and downregulation of Bax. Taken together, miR-181a
functions as an oncogene in gastric cancer that possibly promotes
cancer progression by suppressing RASSF1A and may represent a
potential molecular target for gastric cancer therapy. However,
further studies are required to fully understand the involvement of
miR-181a/RASSF1A signaling in gastric cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81101874 and
81172362), The Science and Technology Project of Shaanxi Province
(grant nos. 2016SF-015 and 2016SF-157), and The Coordinative and
Innovative Plan Projects of the Science and Technology Program in
Shaanxi Province (grant no. 2013KTCQ03-08).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JHY and JQ performed the experiments, acquired the
data and drafted the manuscript. WW and GBW collected gastric
cancer samples and clinicopathological characteristics, and
assisted with the experiments. YHW and QG analyzed and interpreted
data. XJS and JBZ substantially contributed to the study conception
and design. All authors read and approved the final manuscript and
agreed to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of The First Affiliated Hospital of Xi'an Jiaotong
University (Xi'an, China) and written informed consent was obtained
from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Global Burden of Disease Cancer
Collaboration, . Fitzmaurice C, Dicker D, Pain A, Hamavid H,
Moradi-Lakeh M, MacIntyre MF, Allen C, Hansen G, Woodbrook R, Wolfe
C, et al: The Global Burden of Cancer 2013. JAMA Oncol. 1:505–527.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Deng JY and Liang H: Clinical significance
of lymph node metastasis in gastric cancer. World J Gastroenterol.
20:3967–3975. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ono H: Early gastric cancer: Diagnosis,
pathology, treatment techniques and treatment outcomes. Eur J
Gastroenterol Hepatol. 18:863–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bayoumi AS, Sayed A, Broskova Z, Teoh JP,
Wilson J, Su H, Tang YL and Kim IM: Crosstalk between long
noncoding RNAs and microRNAs in health and disease. Int J Mol Sci.
17:3562016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jansson MD and Lund AH: MicroRNA and
cancer. Mol Oncol. 6:590–610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pichiorri F, Suh SS, Ladetto M, Kuehl M,
Palumbo T, Drandi D, Taccioli C, Zanesi N, Alder H, Hagan JP, et
al: MicroRNAs regulate critical genes associated with multiple
myeloma pathogenesis. Proc Natl Acad Sci USA. 105:12885–12890.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Li Z, He C, Wang D, Yuan X, Chen J
and Jin J: MicroRNAs expression signatures are associated with
lineage and survival in acute leukemias. Blood Cells Mol Dis.
44:191–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meng F, Glaser SS, Francis H, DeMorrow S,
Han Y, Passarini JD, Stokes A, Cleary JP, Liu X, Venter J, et al:
Functional analysis of microRNAs in human hepatocellular cancer
stem cells. J Cell Mol Med. 16:160–173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Taylor MA, Sossey-Alaoui K, Thompson CL,
Danielpour D and Schiemann WP: TGF-β upregulates miR-181a
expression to promote breast cancer metastasis. J Clin Invest.
123:150–163. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim CH, Kim HK, Rettig RL, Kim J, Lee ET,
Aprelikova O, Choi IJ, Munroe DJ and Green JE: miRNA signature
associated with outcome of gastric cancer patients following
chemotherapy. BMC Med Genomics. 4:792011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin F, Li Y, Yan S, Liu S, Qian W, Shen D,
Lin Q and Mao W: MicroRNA-181a inhibits tumor proliferation,
invasiveness, and metastasis and is downregulated in gastric
cancer. Oncol Res. 22:75–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dammann R, Li C, Yoon JH, Chin PL, Bates S
and Pfeifer GP: Epigenetic inactivation of a RAS association domain
family protein from the lung tumour suppressor locus 3p21.3. Nat
Genet. 25:315–319. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dubois F, Keller M, Calvayrac O, Soncin F,
Hoa L, Hergovich A, Parrini MC, Mazières J, Vaisse-Lesteven M,
Camonis J, et al: RASSF1A suppresses the invasion and metastatic
potential of human non-small cell lung cancer cells by inhibiting
YAP activation through the GEF-H1/RhoB pathway. Cancer Res.
76:1627–1640. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liao A, Tan G, Chen L, Zhou W and Hu H:
RASSF1A inhibits gastric cancer cell proliferation by
miR-711-mediated downregulation of CDK4 expression. Oncotarget.
7:5842–5851. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bräuer-Hartmann D, Hartmann JU, Wurm AA,
Gerloff D, Katzerke C, Falzacappa Verga MV, Pelicci PG,
Müller-Tidow C, Tenen DG, Niederwieser D, et al: PML/RARα-regulated
miR-181a/b cluster targets the tumor suppressor RASSF1A in acute
promyelocytic leukemia. Cancer Res. 75:3411–3424. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo Q, Wang HB, Li YH, Li HF, Li TT, Zhang
WX, Xiang SS and Sun ZQ: Correlations of promoter methylation in
WIF-1, RASSF1A, and CDH13 genes with the risk and prognosis of
esophageal cancer. Med Sci Monit. 22:2816–2824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Du Z, Ma K, Sun X, Li A, Wang H, Zhang L,
Lin F, Feng X and Song J: Methylation of RASSF1A gene promoter and
the correlation with DNMT1 expression that may contribute to
esophageal squamous cell carcinoma. World J Surg Oncol. 13:1412015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang L, Ma Z, Wang D, Zhao W, Chen L and
Wang G: MicroRNA-602 regulating tumor suppressive gene RASSF1A is
overexpressed in hepatitis B virus-infected liver and
hepatocellular carcinoma. Cancer Biol Ther. 9:803–808. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shin KH, Bae SD, Hong HS, Kim RH, Kang MK
and Park NH: miR-181a shows tumor suppressive effect against oral
squamous cell carcinoma cells by downregulating K-ras. Biochem
Biophys Res Commun. 404:896–902. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi L, Cheng Z, Zhang J, Li R, Zhao P, Fu
Z and You Y: hsa-mir-181a and hsa-mir-181b function as tumor
suppressors in human glioma cells. Brain Res. 1236:185–193. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marton S, Garcia MR, Robello C, Persson H,
Trajtenberg F, Pritsch O, Rovira C, Naya H, Dighiero G and Cayota
A: Small RNAs analysis in CLL reveals a deregulation of miRNA
expression and novel miRNA candidates of putative relevance in CLL
pathogenesis. Leukemia. 22:330–338. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu X, Liao W, Peng H, Luo X, Luo Z, Jiang
H and Xu L: miR-181a promotes G1/S transition and cell
proliferation in pediatric acute myeloid leukemia by targeting ATM.
J Cancer Res Clin Oncol. 142:77–87. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Santarpia L, Nicoloso M and Calin GA:
MicroRNAs: A complex regulatory network drives the acquisition of
malignant cell phenotype. Endocr Relat Cancer. 17:F51–F75. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Grawenda AM and O'Neill E: Clinical
utility of RASSF1A methylation in human malignancies. Br J
Cancer. 113:372–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fernandes MS, Carneiro F, Oliveira C and
Seruca R: Colorectal cancer and RASSF family - a special emphasis
on RASSF1A. Int J Cancer. 132:251–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shivakumar L, Minna J, Sakamaki T, Pestell
R and White MA: The RASSF1A tumor suppressor blocks cell cycle
progression and inhibits cyclin D1 accumulation. Mol Cell Biol.
22:4309–4318. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oh HJ, Lee KK, Song SJ, Jin MS, Song MS,
Lee JH, Im CR, Lee JO, Yonehara S and Lim DS: Role of the tumor
suppressor RASSF1A in Mst1-mediated apoptosis. Cancer Res.
66:2562–2569. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou PH, Zheng JB, Wei GB, Wang XL, Wang
W, Chen NZ, Yu JH, Yao JF, Wang H, Lu ST, et al:
Lentivirus-mediated RASSF1A expression suppresses aggressive
phenotypes of gastric cancer cells in vitro and in vivo. Gene Ther.
22:793–801. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Agathanggelou A, Cooper WN and Latif F:
Role of the Ras-association domain family 1 tumor suppressor gene
in human cancers. Cancer Res. 65:3497–3508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Verduzco D, Dovey JS, Shukla AA, Kodym E,
Skaug BA and Amatruda JF: Multiple isoforms of CDC25 oppose ATM
activity to maintain cell proliferation during vertebrate
development. Mol Cancer Res. 10:1451–1461. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu H, Zhu J, Hu C, Song H and Li Y:
Inhibition of microRNA-181a may suppress proliferation and invasion
and promote apoptosis of cervical cancer cells through the
PTEN/Akt/FOXO1 pathway. J Physiol Biochem. 72:721–732. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pichler M, Winter E, Ress AL, Bauernhofer
T, Gerger A, Kiesslich T, Lax S, Samonigg H and Hoefler G: miR-181a
is associated with poor clinical outcome in patients with
colorectal cancer treated with EGFR inhibitor. J Clin Pathol.
67:198–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiang Z, Dong X, Sun Q, Li X and Yan B:
Clinical significance of up-regulated miR-181a in prognosis and
progression of esophageal cancer. Acta Biochim Biophys Sin.
46:1007–1010. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ji J, Yamashita T, Budhu A, Forgues M, Jia
HL, Li C, Deng C, Wauthier E, Reid LM, Ye QH, et al: Identification
of microRNA-181 by genome-wide screening as a critical player in
EpCAM-positive hepatic cancer stem cells. Hepatology. 50:472–480.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Z, Sun F, Hong Y, Liu Y, Fen M, Yin K,
Ge X, Wang F, Chen X and Guan W: MEG2 is regulated by miR-181a-5p
and functions as a tumour suppressor gene to suppress the
proliferation and migration of gastric cancer cells. Mol Cancer.
16:1332017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao J, Nie Y, Wang H and Lin Y: MiR-181a
suppresses autophagy and sensitizes gastric cancer cells to
cisplatin. Gene. 576:828–833. 2016. View Article : Google Scholar : PubMed/NCBI
|