Lung cancer, recognized as the leading cause of
cancer-associated mortality worldwide, is classified into small
cell lung cancer (SCLC) and non-SCLC (NSCLC). SCLC constitutes ~15%
of all confirmed cases of lung cancer worldwide (1–3).
Distinct from NSCLC, SCLC is unique in its inclination for quick
metastasis and sensitivity to initial systemic cytotoxic
chemotherapy. Systemic chemotherapy is the solid foundation of
treatment for the limited and extensive stages of this disease.
Nevertheless, the commonly adopted management standard of
platinum-oriented chemotherapy has reached an efficacy bottleneck,
mainly due to chemoresistance and relapse in SCLC patients
(4). Despite plentiful clinical
trials in the past four decades, systematic treatment for SCLC
patients has not changed markedly. As a result, the majority of
patients live for only 1 year or less following diagnosis, with the
overall 5-year survival rate staying low at <7% (5). The widely employed technique to
diagnose SCLC from a tiny amount of malignant cells, in combination
with a lack of proved predictive biomarkers that would require
tissue biopsies and relatively rare surgical resection, has cut
down the source of SCLC tissue for more profound studies (5). In NSCLC, a growing number of gene
fusions or mutations instruct treatment selections for specific
patient subgroups, particularly those with anaplastic lymphoma
kinase or epidermal growth factor receptor (EGFR) (6) mutations. In marked contrast, numerous
experimental and targeted agents regarding SCLC have failed to
yield convincing clinical benefits (7). Novel and effective therapies for SCLC
patients are urgently required. Regarding the molecular mechanisms
of carcinogenesis, medical communities have mostly concentrated on
genes with protein-coding capacity. Unexpectedly, the ENCODE
project identified that up to three-quarters of the human genome
could be transcribed, though <3% of it encodes protein. This
unexpected fact indicated that non-coding RNAs (ncRNAs) make up the
vast majority of the human transcriptome (8). Long ncRNAs (lncRNAs) are >200
nucleotides in size and possess no or a very low protein-coding
ability. Bioinformatics platforms and high-throughput sequencing
emerging in recent years have facilitated uncovering the mystique
of lncRNAs, which function as key molecules in wide-ranging
cellular processes, including cell growth, adhesion, proliferation
and apoptosis (9,10). lncRNA deregulation is involved in
numerous human diseases, and there is also increasing evidence
suggesting that lncRNAs are involved in SCLC pathogenesis and
clinical outcomes (11–15). Digging deeper into the biological
functions and molecular mechanisms of lncRNAs will enable
researchers to further understand the biology of SCLC and develop
lncRNA-oriented therapeutics.
lncRNAs are a set of ubiquitous genes participating
in various biological mechanisms. There are four archetypes in
which lncRNAs execute their molecular functions, namely as signals,
decoys, guides and scaffolds (25).
The signal archetype of lncRNAs may serve as markers of
functionally significant biological events, as their expression
exhibits cell type, time and space specificity. For example, lncRNA
homeobox (HOX) transcript antisense intergenic RNA (HOTAIR) located
in the HOXC locus exists in posterior and distal cells, whereas
another HOXC lncRNA, Frigidair, is expressed in an anterior
pattern. Conversely, lncRNA HOXA transcript at the distal tip
(HOTTIP), located in the far end of the human HOXA cluster, is
expressed in distal cells (26,27).
The decoys archetype is a type of lncRNA that regulates
transcription through binding to and then carrying away protein
targets, yet it does not exert extra functions. Decoys display as
‘molecular sinks’ for chromatin modifiers, transcription factors or
other regulatory factors, all of which are RNA-binding proteins
(25). For instance, by directly
binding to and sequestering nuclear transcription factor Y subunit
that drives a DNA damage-induced apoptotic program, lncRNA
p21-associated ncRNA DNA damage-activated suppresses apoptotic gene
expression to facilitate cell cycle arrest, leading to the
promotion of cell survival (28).
Knockdown of lncRNAs of this archetype may imitate the
gain-of-function of the target proteins, while a rescue phenotype
could be induced by loss-of-function of the lncRNA and its effector
(25). The guides archetype of
lncRNA can bind chromatin modifying proteins and direct the
localization of ribonucleoprotein complexes to specific targets in
a cis or trans manner. The well-known cis
mechanism, mammalian X inactivation center, specifies a set of
ncRNAs, X-inactive specific transcript (Xist) included (29,30). A
1.6-kb lncRNA, RepA RNA, stemming from the 5′ end of Xist, produces
polycomb repressive complex 2 (PRC2) in cis. PRC2 is
involved in extra X-chromosome inactivation (31). In contrast to cis-regulatory
lncRNAs, certain lncRNAs serve their chromosome-wide
transcriptional roles in trans, such as lncRNA HOTAIR, which
is capable of directing PRC2 to target genes in trans
(32–34). The scaffolds archetype of lncRNA can
act a platform where components are assembled, precisely regulating
the sophisticated molecular interactions and signaling
transductions involved in diverse biological signaling processes
(35). For example, telomerase
catalytic activity necessitates the combination of two common
telomerase units, the telomerase RNA (TERC) and the telomerase
reverse transcriptase (TERT). TERC is an essential lncRNA unit that
offers the template for repeat synthesis, and it also possesses
domains that promote TERT binding, catalytic activity and stability
of the complex (36). Certain
morbid states, including dyskeratosis congenital, presumably result
from mutations altering the equilibrium between different
conformations of TERC, more specifically, through destruction of
the RNA scaffold structure where modular biding sites for telomeric
regulatory proteins are located (37).
lncRNAs exert functions in an enormous range of
biological processes by promoting or inhibiting the transcription
and translation of protein-coding genes. Unlike highly conserved
small ncRNAs that participate in gene silencing transcriptionally
and post-transcriptionally (38–40),
lncRNAs are poorly conserved and can modulate target gene
expression via various mechanisms at different levels.
At the transcriptional level, lncRNAs have the
following roles: i) Functioning as decoys for RNA polymerase II or
transcription factors (TFs) to inhibit their binding to enhancers
or promoters of target genes, therefore specifically promoting or
repressing target gene expression (26); ii) alteration of TF localization or
modification to promote or inhibit gene transcription (40); iii) interaction with DNA to form a
triple helix structure, thereby affecting target gene transcription
(41); and iv) presenting as
competitive endogenous RNAs (ceRNAs) to inhibit the transcription
of target genes (42).
At the post-transcriptional level, lncRNAs have the
following roles: i) Providing different transcripts by regulating
pre-mRNA alternative splicing (43); ii) combining with mRNAs to
synthesize double-stranded RNA complexes, thereby effectively
enhancing the stability of mRNAs (44); and iii) interaction with miRNAs to
regulate signaling events (45).
At the epigenetic level, lncRNAs have the following
roles: i) The regulation of histone modifications, including
acetylation, methylation and ubiquitination, among others (46); ii) participating in chromatin
remodeling and conformational alterations by combining with
chromatin modification complexes that are crucial for gene
transcription (47); and iii)
participating in gene silencing via modulating DNA methylation in
the promoter region of target genes (48).
To summarize, lncRNAs participate in diverse
transcriptional, post-transcriptional and epigenetic molecular
mechanisms, covering regulation of chromatin structure or
modification, transcription, splicing and translation, therefore
regulating a multitude of physiological and pathological courses,
including cell proliferation, differentiation, apoptosis, the heat
shock response, cancer development and chemoresistance (49–51).
Amongst these functions, the regulation of gene expression is of
paramount significance in elucidating how lncRNAs promote or
suppress tumorigenesis. Genome-wide studies of tumor samples have
verified plentiful lncRNAs that are linked to distinct types of
cancer. Dysregulated expression of lncRNAs can stimulate
carcinogenesis and metastasis. However, from an overall
perspective, the function of lncRNAs may not be one-sided, and
could be tumor-promoting or tumor-suppressing.
Representing one of the largest classes of
transcripts, lncRNAs possess highly diverse characteristics and
functions. Progress in high-throughput sequencing technology has
accelerated the identification of lncRNAs as key regulatory
molecules participating in various cellular processes and their
dysregulation in human diseases. Although only a few lncRNAs have
been well described thus far, accumulating evidence suggests that
lncRNAs contribute to tumor biology. Given the aforementioned
difficulties, breakthroughs in SCLC research remain stagnant
compared with those in other types of cancer. However, it remains
worthwhile to investigate the research status of SCLC from an
lncRNA point of view, as this field may open novel and optimistic
windows to elucidate SCLC molecular mechanisms. In the following
text, previous findings in the expression of lncRNAs in SCLC are
reviewed. The roles that HOTTIP, HOTAIR, taurine upregulated gene 1
(TUG1), colon cancer-associated transcript 2 (CCAT2) and
plasmacytoma variant translocation 1 (PVT1) serve in SCLC are
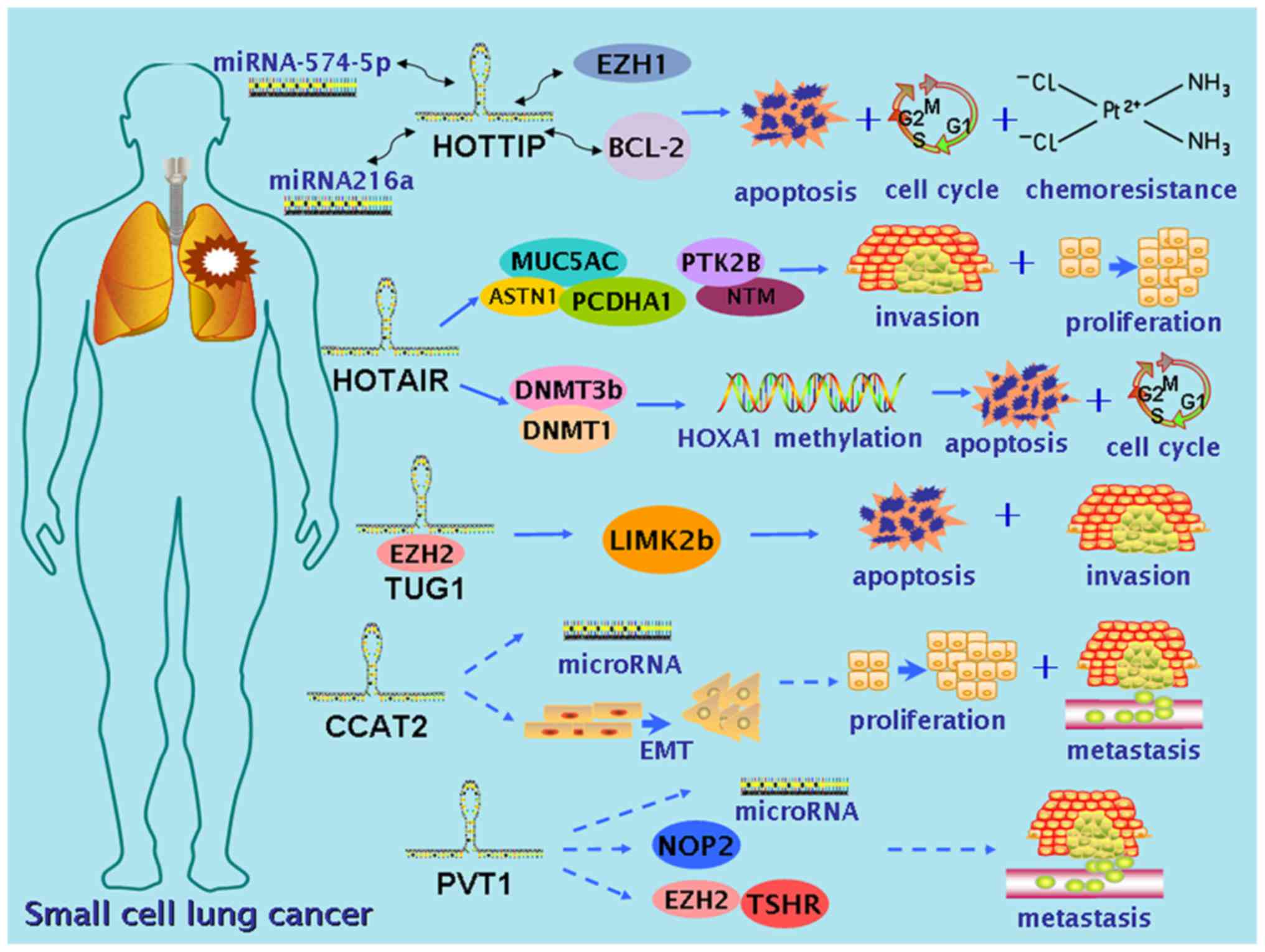
discussed and also briefly presented in Table I. A schematic diagram of these genes
in SCLC and their mechanisms is shown in Fig. 1. The dysregulations and functions of
these five lncRNAs in other malignancies are also summarized, as a
contrast and enlightenment to their roles in SCLC (Table II).
HOTTIP, as the lncRNA encoded by the HOTTIP gene
that is located at the HOXA locus, was initially identified in
human fibroblasts distributed in anatomically distal regions of the
body (52). Wang et al
(53) verified the direct coupling
of HOTTIP and the adaptor protein, WD repeat domain 5 (WDR5) to
target WDR5/lysine methyltransferase 2A complexes across HOXA,
thereby impelling histone lysine 4 trimethylation and the
transcription of various 5′ HOXA genes. Multiple studies confirmed
the positive correlation between the expression level of HOTTIP and
HOXA genes in a variety of malignancies (52,54–56).
In brief, HOTTIP could activate HOX genes by recruiting
histone-modifying enzymes to suppress tumor-suppressor genes. Sun
et al (57,58) completed pioneering studies unveiling
the underlying molecular mechanism of HOTTIP in SCLC utilizing a
series of experiments conducted in vitro and in vivo.
At first, gene expression array analysis revealed the
overexpression of HOTTIP in H69 and H69R cell lines, and the result
was further supported by the significant overexpression of HOTTIP,
as detected by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) in 50 clinical SCLC tissues prior to
chemotherapy, compared with their non-cancerous counterparts. In
addition, higher HOTTIP expression was significantly associated
with a poorer prognosis. Manipulation of HOTTIP loss- and gain- of
function experiments in SCLC cell lines also demonstrated that
HOTTIP overexpression contributed to cell proliferation, as it led
to a decreased number of G2-phase cells and an increased number of
S-phase cells. In vivo, HOTTIP loss and gain of function
experiments conducted in xenograft nude mice showed that mice with
knockdown of HOTTIP had a smaller mean tumor volume in comparison
to those in the negative control group. Afterwards, by employing
web-based bioinformatics platform RNA22-seq (https://cm.jefferson.edu/), miR-574-5p and enhancer of
zeste homolog 1 (EZH1) were predicted to possess targeted binding
sites for HOTTIP, and this association was later verified by
RT-qPCR. Therefore, it was assumed that HOTTIP may exert its effect
on SCLC through a regulatory network of miRNA-574-5p-HOTTIP-EZH1
(57). Notably, the hypothesis was
verified by a subsequent co-transfection dual luciferase reporter
assay, indicating that HOTTIP acts as an oncogene by sponging
miR-574-5p to abrogate the expression of polycomb group protein
EZH1 induced by miR-574-5p, thereby promoting the progression of
SCLC (57). In another study by Sun
et al (58), a similar
experimental design was applied to investigate the role of HOTTIP
in SCLC, and the association of HOTTIP with SCLC chemoresistance
was also investigated, which enriched the clinical value of the
study. The expression of HOTTIP and HOXA13 was markedly upregulated
in SCLC cell lines and biopsy samples. Overexpression of HOTTIP
impaired the anti-chemoresistance effects of etoposide, irinotecan
and cisplatin toward SCLC cells in vitro and in vivo,
whereas knockdown of HOTTIP exhibited a reversed effect. In
addition, the finding that knockdown of HOTTIP suppressed HOXA13
expression, combined with the result of a rescue experiment by
HOXA13 overexpression implied that HOTTIP exerts its function in
SCLC chemoresistance and progression partly via manipulating
HOXA13. Likewise, the online bioinformatics tool RNA22-seq
excavated miR-216a as possessing targeted binding sites with
HOTTIP, and unexpectedly, an atoptosis-related gene, B-cell
leukemia/lymphoma-2 (BCL-2). Subsequent experiments confirmed that
HOTTIP could function as a competing ‘sponge’ through binding
miR-216a, thereby diminishing its silencing effect toward BCL-2,
contributing to the chemoresistance and progression of SCLC
(58). Although the aforementioned
findings may only be the tip of the iceberg, they widen the
landscape of research into the molecular mechanism of SCLC, as a
novel network composed of lncRNA, miRNA and specific cancer-related
genes is put forward, providing inspiration for developing
prognostic and therapeutic agents.
HOTAIR is one of the most well-characterized lncRNAs
and is overexpressed in certain malignancies, including breast,
colorectal, hepatocellular, gastrointestinal and non-small cell
lung cancer (59). First identified
in 2007, HOTAIR resides in the HOXC locus. Previous reports
revealed that the molecular mechanism of HOTAIR is its
transcription from the HOXC gene as an antisense transcript and
then binding to PRC2 (composed of EZH2, polycomb protein SUZ12 and
polycomb protein EED) and lysine-specific demethylase/CoREST/REST
complex as a scaffold, leading to catalyzation of trimethylation of
histone H3 on lysine 27 (H3K27) and spontaneous demethylation of
H3K4, and repression of the transcription of HOXD genes (27,60).
With regard to DNA methylation, EZH2, a compartment of PRC2,
directly interacts with DNA methyltransferases. This interaction
assists in maintaining DNA methylation and stabilizing the
repression of certain genes, including various tumor suppressors
(61). As targets of HOTAIR, the
homeobox-containing genes are a set of regulators that
transcriptionally encode DNA-binding homeodomains that participate
in controlling normal development (51,62).
In addition, abnormal expression of HOX genes is associated with
oncogenesis and paramorphia (61,63).
Additionally, by inducing epithelial-to-mesenchymal transition
(EMT), HOTAIR associates with tumorigenesis (64). HOTAIR also triggers
ubiquitin-mediated proteolysis via interaction with RNA-binding
protein MEX3B and E3 ubiquitin-protein ligase DZIP3 (65). Ono et al (66) studied the association of HOTAIR with
SCLC cellular processes and clinical characteristics. The study
assessed HOTAIR expression in 35 surgically resected SCLC tissues
and 10 SCLC cell lines, and observed that expression of HOTAIR in
pure SCLC was markedly overexpressed compared with that in those
combined with lung adenocarcinoma (LUAD), large cell carcinoma or
large cell neuroendocrine carcinoma, and that HOTAIR overexpression
was clearly associated with relapse and lymphatic invasion. In
vitro experiments indicated that the expression of HOTAIR in
half of the SCLC cell lines was elevated compared with that in
normal cells. Knockdown of HOTAIR reduced cellular invasiveness and
proliferative activity of SBC-3 cells. Gene expression analysis
revealed that a reduction in HOTAIR led to upregulated expression
of mucin production-related genes, including mucin 5AC, and cell
adhesion-related genes, including astrotactin 1 and protocadherin
α1, and downregulated expression of genes such as neurotrimin and
protein tyrosine kinase 2β, participating in neuronal growth and
signal transduction, respectively (66). Recently, Fang et al (67) investigated the role of HOTAIR
expression in the chemoresistance of SCLC and its underlying
mechanism. The study assessed the impact of HOTAIR on SCLC
chemoresistance in vitro and observed that HOTAIR expression
was also markedly upregulated in drug-resistant cell lines compared
with that in the parental cell lines. The study showed that
downregulated expression of HOTAIR promoted cell cycle arrest and
apoptosis to increase sensitivity to antitumor drugs, while
repressing tumor growth in vivo. In addition, increased
HOXA1 methylation was observed in the drug-resistant cells. An
enzyme-linked immunosorbent assay revealed that a reduction of
HOTAIR lessened HOXA1 methylation via reducing the expression of
DNA (cytosine-5)-methyltransferase (DNMT)3b and DNMT1. RNA
immunoprecipitation validated the interaction between HOXA1 and
HOTAIR. Together, these findings indicated that HOTAIR mediates
chemoresistance by increasing HOXA1 methylation. Hence, HOTAIR
could serve as a possible target for novel therapeutics to combat
chemoresistance. Based on these previous findings, lncRNA HOTAIR is
involved in the relapse and lymphatic invasion in SCLC patients,
and it could also act as a biomarker for prognosis and chemotherapy
response, and as a therapeutic target to overcome the
chemoresistance of SCLC.
TUG1 was initially described as a spliced and
polyadenylated lncRNA necessary for the development of
photoreceptors in mice retina (68). Increasing evidence has demonstrated
that TUG1 serves a crucial role in a number of human tumors,
including hepatocellular carcinoma, osteosarcoma, glioma,
esophageal, gastric and bladder cancer (69–74).
Aberrant expression of PRC2-related lncRNAs is involved in
tumorigenesis and progression. In a previous study, TUG1 was found
to be induced by p53, prior to binding to PCR2 and influencing
certain genes involved in the modulation of mitosis, spindle
construction and cell-cycle phasing (34). Yang et al (75) revealed that a combination of
methylated PRC2 and TUG1 manipulates the relocation of
growth-control genes between interchromatin granules and polycomb
bodies in response to growth signals, therefore portraying a role
that TUG1 serves in the relocation of transcription units to
coordinate gene expression (75).
Niu et al (76) investigated
the functions of TUG1 in the cell proliferation and chemoresistance
of SCLC, and its underlying molecular mechanism (76). The study analyzed TUG1 expression in
tissue samples from SCLC patients (n=33) who had undergone biopsy
or bronchofiberscopy, and elevated TUG1 expression was found in
cancerous tissues compared with that in adjacent non-cancerous
tissues. Statistical analysis showed that higher expression of TUG1
was associated with shorter survival time, advanced clinical stage
and cigarette smoking. In vitro Cell Counting Kit-8 and
colony formation assays indicated that silencing TUG1 markedly
reduced cell growth. The results from flow cytometry analysis
conducted to assess the effect of TUG1 on cell apoptosis suggested
that knockdown of TUG1 promoted apoptosis and led to a significant
accumulation of G1-phase cells, and that downregulated TUG1
expression increased apoptosis in H44DDP and H69AR cell lines
exposed to anticancer drugs. The chemoresistance-inducing ability
of TUG1 in vivo was further investigated using a mouse
xenograft model, and the result was consistent with that of the
in vitro experiment. Moreover, TUG1 could modulate LIM
domain kinase 2 expression through binding with EZH2, and
subsequently led to increased cell growth and chemoresistance in
SCLC. Outcomes of this study could be guidance to the development
of innovative TUG1-directed prognostic and therapeutic
strategies.
CCAT2 was first introduced in 2013 as an lncRNA
located in the 8q24 gene desert region, and it possesses a
tumor-related single nucleotide polymorphism rs6983276.
Additionally, overexpression of CCAT2 in colon cancer was observed,
and it was considered to serve an oncogenic role, promoting
colorectal cancer cell proliferation and motility, metastasis and
chromosomal instability by regulating myc and Wnt pathways
(77). A CCAT2 genetic
polymorphism, rs6983267, is associated with platinum-based
chemotherapy sensitivity in lung cancer patients (78). Since its discovery, the oncogenic
role of CCAT 2 has been increasingly demonstrated in different
tumors, including gastric, breast, lung, liver, colon, cervical,
ovarian, bladder, prostate and esophageal cancer (79–84).
The stimulatory effects on the Wnt/β-catenin signaling pathway,
cancer metabolism and EMT may underlie its oncogenic action
(79). CCAT2 is upregulated in an
estimated two-thirds of breast cancer patients (80). High CCAT2 expression was associated
with a poor curative effect from
cyclophosphamide/methotrexate/fluorouracil-containing adjuvant
chemotherapy in breast cancer patients with lymph node metastasis
(80). Chen et al (85) detected CCAT2 expression in 102 human
SCLC tissues, 15 paired non-tumor tissues, SCLC cell lines (DMS-53
and H446) and a normal bronchial epithelial cell line (16HBE). The
association between clinicopathological factors and CCAT2
expression was subsequently analyzed. The study reported that CCAT2
level was significantly overexpressed in SCLC tissue and cell lines
compared with that in normal lung tissues. Subgroup analyses also
indicated that higher expression of CCAT2 was correlated with
malignant status and poor prognosis in SCLC patients. Moreover,
knockdown of CCAT2 to inhibit SCLC cell growth and metastasis in
vitro was observed. To conclude, CCAT2 may serve as an oncogene
and a negative prognostic indicator in SCLC.
PVT1 is an lncRNA homologous to the mouse
plasmacytoma variant translocation gene (Pvt1), which was first
identified as being frequently involved in a variant translocation
in plasmacytoma in the mid-80s in mice (86,87).
Soon after, the PVT1 locus emerged as a site of variant
translocations in Burkitt lymphoma. Subsequent studies support the
role of PVT1 as a cancer risk locus in relation to the well-known
myc oncogene (88). PVT1 presents
the capacity to facilitate cell growth and suppress cell apoptosis
in the tumorigenesis of various types of cancer, including gastric
(89), liver (90), thyroid (91) and pancreatic (92) cancer, and non-small cell lung cancer
(93). The expression of PVT1 in
tumor samples of these types of cancer is elevated. In vitro
and in vivo experiments conducted by Wang et al
(90) demonstrated that PVT1
promotes cell proliferation, cell cycling and the acquisition of
stem cell-like properties in hepatocellular carcinoma cell by
stabilizing NOP2 nucleolar protein (90). However, the underlying mechanisms of
the functional exertion of PVT1 and its interaction with downstream
targets remain largely unknown. Partially known molecular functions
of PVT1 can be categorized into three key pathways: Partaking in
DNA rearrangement, encoding microRNAs and intercommunicating with
myc (94). Recently, Huang et
al (95) first identified the
role of PVT1 in SCLC. In the study, PVT1 expression was detected in
SCLC tissues, paired normal gastric tissues and two SCLC cell
lines. Meanwhile, the association of PVT1 expression levels with
clinical features of 120 enrolled SCLC patients was analyzed.
RT-PCR analysis showed that PVT1 expression was significantly
higher in SCLC tissues and cell lines than in their normal
counterparts, and positive correlations between PVT1 overexpression
and the status of clinical stage, lymph node metastasis and distal
metastasis in SCLC were noted. Furthermore, multivariate analysis
revealed that PVT overexpression could be an independent prognostic
biomarker for the survival of SCLC patients. Cell migration and
invasion were significantly suppressed in vitro by silencing
of PVT1 in SCLC. To conclude, PVT1 possesses the potency to be a
novel marker and a prospect to develop targeted therapy for SCLC.
However, further investigations are required for thorough
elucidation of the molecular mechanism of PVT1 in SCLC.
Thus far, the present review has summarized and
discussed five lncRNAs (HOTTIP, HOTAIR, TUG1, CCAT2 and PVT1)
involved in SCLC. However, these lncRNAs are also involved in
NSCLC, which accounts for ~85% of all lung cancer cases. It is of
merit to include information on the roles that these lncRNAs serve
in NSCLC, as a contrast and possibly, enlightenment. HOTTIP was
reported to be significantly upregulated in NSCLC and to function
as an oncogene by regulating HOXA13, which coincides with the
findings observed in other malignancies, implying that HOXA13 is a
key element through which HOTTIP promotes carcinogenesis (96). Moreover, overexpression of HOTTIP
was found to motivate LUAD cell proliferation and chemoresistance
via regulating the protein kinase B (AKT) signaling pathway
(97). As for HOTAIR, multiple
studies also demonstrated its overexpression in NSCLC, and it is
involved in the initiation and development of NSCLC through
interacting with unc-51-like autophagy-activating kinase 1 to
suppress autophagy (98), targeting
caveolin 1 (99) and miR-613
(100). Based on the available
literature, there is controversy with regard to the expression of
TUG1 in NSCLC. Studies by Lin et al (101) and Zhang et al (102) revealed that TUG1 was downregulated
in NSCLC, while a study by Liu et al (103) showed that it was upregulated.
According to the study by Liu et al, TUG1 could inhibit
apoptosis by silencing BCL-2 associated X via interacting with EZH2
(103). TUG1 RNA could target PRC2
in the promotor region of CUGBP Elav-like family member 1 (CELF1)
and CELF1 expression was therefore negatively regulated (101). Zhang et al (102) suggested that TUG1 acted as a
growth regulator in NSCLC partly through controlling HOXB7. CCAT2
exerted overexpression in NSCLC (104,105), and could promote oncogenesis via
overexpression of zinc finger and BTB domain containing 7A
(104). With regard to PVT1, it is
of note that accumulating recent studies (106–109) investigated the roles of
upregulated PVT1 in NSCLC via the ceRNA-regulated network, which is
a trending hotspot in the research field. PVT1 was demonstrated to
exert its oncogenic functions by sponging miR199a5p (106), miR-126 (107), miR-497 (108) and miR-195 (109).
SCLC is a fatal disease with an aggressive and
brutal nature; it comprises ~15% of all lung cancer cases. The
management of SCLC remains challenging, while disease outcome has
remained poor, mainly due to limited options for effective
treatment. The majority of the cases are at an irreversible
advanced stage when diagnosed and rapidly develop treatment
resistance despite a high success rate of initial chemotherapy and
radiation. The pathogenesis of SCLC has been investigated by
researchers across the world; nevertheless, the implicit molecular
mechanism remains mostly unidentified. In light of next-generation
sequencing techniques and bioinformatics tools, lncRNAs have been
shown to exert distinguishable functions in a broad range of human
diseases, including the most concerning types of cancer. Although
great discoveries and advances in cancer pathogenesis and
therapeutics have been made over the decade, the elucidation of the
SCLC molecular mechanism and its frontline treatment have developed
slowly due to obstacles from various aspects, including difficulty
in sample collection, research funding, late diagnosis, rapid
progression and chemoresistance. Investigation into the role of
lncRNAs in SCLC is underway, yet no lncRNAs have been extensively
investigated, let alone clinically utilized for prognosis,
diagnosis or therapeutic design. According to the available
published literature, the current research state of SCLC is
relatively superficial compared with that of NSCLC. Thus, more
research is urgently required.
Despite the aforementioned challenges, there have
been certain notable novel findings that have the potential to
advance the field. Given the scarcity of SCLC tissues, a
multidisciplinary, interoperable, cross-institutional approach is
required to collect adequate SCLC tissues for more translational
research projects. Emerging techniques, including next-generation
sequencing and bioinformatics, have created opportunities to
conduct larger scale and deeper studies on SCLC. The past decade
has witnessed the emergence of lncRNAs involved in various types of
cancer. For example, lncRNA prostate cancer antigen 3 and lncRNA
highly upregulated in liver cancer can be detected in prostate and
liver cancer, respectively, and serve as sensitive diagnostic
markers (110,111). As an intensively studied lncRNA,
lncRNA metastasis-associated lung adenocarcinoma transcript 1 is
found to be involved in multiple malignancies, including lung,
colon, breast and liver cancer, indicating its general
participation in cancer cell proliferation (112). Overexpression of lncRNA antisense
noncoding RNA in the INK4 locus is observed in a number of types of
cancer and is associated with a poor prognosis in gastric and
prostate cancer (113). lncRNA
CCAT1 could be used as a clinically detectable marker to predict
the therapeutic responsiveness of bromodomain and extraterminal
inhibitors in patients with colorectal cancer (114). lncRNA maternally expressed gene 3,
which acts as a tumor-suppressor via promoting p53 accumulation and
recruiting PRC2, is downregulated in multiple primary human tumors
(115). As these lncRNAs have been
verified as promising predictive markers for diagnosis, prognosis
and chemotherapy sensitivity in cancer patients, they also have the
potential to lead to a greater understanding of SCLC tumorigenesis
and chemoresistance, and could serve as efficient therapeutic
targets. The diagnostic sensitivity and specificity may be enhanced
by joint detection of disparate lncRNAs, and this may become
particularly useful in non-invasive screening for early-stage SCLC
patients. The functional roles of lncRNAs involve diverse signaling
pathways and investigation into these pathways may yield crucial
signaling targets that could be blocked to impede tumor
progression. Signaling pathways frequently altered in cancer
include phosphoinositide 3-kinase/AKT, Kirsten rat sarcoma viral
oncogene homolog/V-raf murine sarcoma b-viral oncogene homolog B1,
retrovirus-associated DNA sequences/mitogen-activated protein
kinase, EGFR, fibroblast growth factor receptor, Wnt and myc, among
others (116). Currently, among
nearly 20,000 identified lncRNAs, only five have been investigated
in SCLC. Therefore, more efforts should be put into this field of
great potential. As a large genetic information treasury and a
potential opening to combat diseases, lncRNAs will play no small
part in identifying SCLC mechanisms.
Not applicable.
The present study was supported by funds from the
National Natural Science Foundation of China (grant no.
NSFC81560469 and NSFC81360327), the Natural Science Foundation of
Guangxi, China (grant no. 2015GXNSFCA139009, 2016GXNSFAA380255 and
2017GXNSFAA198016) and the Guangxi Medical University Training
Program for Distinguished Young Scholars (grant no. 2017).
Not applicable.
TTL contributed to the literature retrieval and
manuscript preparation. RQH and JM contributed to the manuscript
modification and commented on multiple aspects of the manuscript.
ZYL, XHH and GC jointly supervised the construction of the study,
and contributed to the design and approval of the final version of
the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
van Meerbeeck JP, Fennell DA and De
Ruysscher DK: Small-cell lung cancer. Lancet. 378:1741–1755. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barnes H, See K, Barnett S and Manser R:
Surgery for limited-stage small-cell lung cancer. Cochrane Database
Syst Rev. 4:CD0119172017.PubMed/NCBI
|
|
3
|
Sabari JK, Lok BH, Laird JH, Poirier JT
and Rudin CM: Unravelling the biology of SCLC: Implications for
therapy. Nat Rev Clin Oncol. 14:549–561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pillai RN and Owonikoko TK: Small cell
lung cancer: Therapies and targets. Semin Oncol. 41:133–142. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Byers LA and Rudin CM: Small cell lung
cancer: Where do we go from here? Cancer. 121:664–672. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dolly SO, Collins DC, Sundar R, Popat S
and Yap TA: Advances in the development of molecularly targeted
agents in non-small-cell lung cancer. Drugs. 77:813–827. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seeber A, Leitner C, Philipp-Abbrederis K,
Spizzo G and Kocher F: What's new in small cell lung
cancer-extensive disease? An overview on advances of systemic
treatment in 2016. Future Oncol. 13:1427–1435. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
George J, Lim JS, Jang SJ, Cun Y, Ozretić
L, Kong G, Leenders F, Lu X, Fernández-Cuesta L, Bosco G, et al:
Comprehensive genomic profiles of small cell lung cancer. Nature.
524:47–53. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pleasance ED, Stephens PJ, O'Meara S,
McBride DJ, Meynert A, Jones D, Lin ML, Beare D, Lau KW, Greenman
C, et al: A small-cell lung cancer genome with complex signatures
of tobacco exposure. Nature. 463:184–190. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ulitsky I and Bartel DP: LincRNAs:
Genomics, evolution, and mechanisms. Cell. 154:26–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang J, Peng J and Guo L: Non-coding RNA:
A new tool for the diagnosis, prognosis, and therapy of small cell
lung cancer. J Thorac Oncol. 10:28–37. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kwok ZH and Tay Y: Long noncoding RNAs:
Lincs between human health and disease. Biochem Soc Trans.
45:805–812. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ohno S: So much ‘junk’ DNA in our genome.
Brookhaven Symp Biol. 23:366–370. 1972.PubMed/NCBI
|
|
17
|
Spornraft M, Kirchner B, Pfaffl MW and
Riedmaier I: Comparison of the miRNome and piRNome of bovine blood
and plasma by small RNA sequencing. Biotechnol Lett. 37:1165–1176.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Busch H, Reddy R, Rothblum L and Choi YC:
SnRNAs, SnRNPs, and RNA processing. Annu Rev Biochem. 51:617–654.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ota T, Suzuki Y, Nishikawa T, Otsuki T,
Sugiyama T, Irie R, Wakamatsu A, Hayashi K, Sato H, Nagai K, et al:
Complete sequencing and characterization of 21,243 full-length
human cDNAs. Nat Genet. 36:40–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bertone P, Stolc V, Royce TE, Rozowsky JS,
Urban AE, Zhu X, Rinn JL, Tongprasit W, Samanta M, Weissman S, et
al: Global identification of human transcribed sequences with
genome tiling arrays. Science. 306:2242–2246. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Okazaki Y, Furuno M, Kasukawa T, Adachi J,
Bono H, Kondo S, Nikaido I, Osato N, Saito R, Suzuki H, et al:
Analysis of the mouse transcriptome based on functional annotation
of 60,770 full-length cDNAs. Nature. 420:563–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hon CC, Ramilowski JA, Harshbarger J,
Bertin N, Rackham OJ, Gough J, Denisenko E, Schmeier S, Poulsen TM,
Severin J, et al: An atlas of human long non-coding RNAs with
accurate 5′ ends. Nature. 543:199–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hombach S and Kretz M: Non-coding RNAs:
Classification, biology and functioning. Adv Exp Med Biol.
937:3–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang KC, Helms JA and Chang HY:
Regeneration, repair and remembering identity: The three Rs of
Hox gene expression. Trends Cell Biol. 19:268–275. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E, et
al: Functional demarcation of active and silent chromatin domains
in human HOX loci by noncoding RNAs. Cell. 129:1311–1323.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hung T, Wang Y, Lin MF, Koegel AK, Kotake
Y, Grant GD, Horlings HM, Shah N, Umbricht C, Wang P, et al:
Extensive and coordinated transcription of noncoding RNAs within
cell-cycle promoters. Nat Genet. 43:621–629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Plath K, Mlynarczyk-Evans S, Nusinow DA
and Panning B: Xist RNA and the mechanism of X chromosome
inactivation. Annu Rev Genet. 36:233–278. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee JT: The X as model for RNA's niche in
epigenomic regulation. Cold Spring Harb Perspect Biol.
2:a0037492010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wutz A, Rasmussen TP and Jaenisch R:
Chromosomal silencing and localization are mediated by different
domains of Xist RNA. Nat Genet. 30:167–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao J, Ohsumi TK, Kung JT, Ogawa Y, Grau
DJ, Sarma K, Song JJ, Kingston RE, Borowsky M and Lee JT:
Genome-wide identification of polycomb-associated RNAs by RIP-seq.
Mol Cell. 40:939–953. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Khalil AM, Guttman M, Huarte M, Garber M,
Raj A, Morales Rivea D, Thomas K, Presser A, Bernstein BE, van
Oudenaarden A, et al: Many human large intergenic noncoding RNAs
associate with chromatin-modifying complexes and affect gene
expression. Proc Natl Acad Sci USA. 106:11667–11672. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Spitale RC, Tsai MC and Chang HY: RNA
templating the epigenome: Long noncoding RNAs as molecular
scaffolds. Epigenetics. 6:539–543. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Collins K: Physiological assembly and
activity of human telomerase complexes. Mech Ageing Dev. 129:91–98.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen JL and Greider CW: Telomerase RNA
structure and function: Implications for dyskeratosis congenita.
Trends Biochem Sci. 29:183–192. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Moretti F, Thermann R and Hentze MW:
Mechanism of translational regulation by miR-2 from sites in the 5′
untranslated region or the open reading frame. RNA. 16:2493–2502.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ørom UA, Nielsen FC and Lund AH:
MicroRNA-10a binds the 5′UTR of ribosomal protein mRNAs and
enhances their translation. Mol Cell. 30:460–471. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hu X, Feng Y, Zhang D, Zhao SD, Hu Z,
Greshock J, Zhang Y, Yang L, Zhong X, Wang LP, et al: A functional
genomic approach identifies FAL1 as an oncogenic long
noncoding RNA that associates with BMI1 and represses p21
expression in cancer. Cancer Cell. 26:344–357. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yin Y, Yan P, Lu J, Song G, Zhu Y, Li Z,
Zhao Y, Shen B, Huang X, Zhu H, et al: Opposing roles for the
lncRNA Haunt and its genomic locus in regulating HOXA
gene activation during embryonic stem cell differentiation. Cell
Stem Cell. 16:504–516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tripathi V, Ellis JD, Shen Z, Song DY, Pan
Q, Watt AT, Freier SM, Bennett CF, Sharma A, Bubulya PA, et al: The
nuclear-retained noncoding RNA MALAT1 regulates alternative
splicing by modulating SR splicing factor phosphorylation. Mol
Cell. 39:925–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-beta promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Steck E, Boeuf S, Gabler J, Werth N,
Schnatzer P, Diederichs S and Richter W: Regulation of H19 and its
encoded microRNA-675 in osteoarthritis and under anabolic and
catabolic in vitro conditions. J Mol Med. 90:1185–1195. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Houseley J, Rubbi L, Grunstein M,
Tollervey D and Vogelauer M: A ncRNA modulates histone modification
and mRNA induction in the yeast GAL gene cluster. Mol Cell.
32:685–695. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hainer SJ, Gu W, Carone BR, Landry BD,
Rando OJ, Mello CC and Fazzio TG: Suppression of pervasive
noncoding transcription in embryonic stem cells by esBAF. Genes
Dev. 29:362–378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Berghoff EG, Clark MF, Chen S, Cajigas I,
Leib DE and Kohtz JD: Evf2 (Dlx6as) lncRNA regulates
ultraconserved enhancer methylation and the differential
transcriptional control of adjacent genes. Development.
140:4407–4416. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen G, Wang Z, Wang D, Qiu C, Liu M, Chen
X, Zhang Q, Yan G and Cui Q: LncRNADisease: A database for
long-non-coding RNA-associated diseases. Nucleic Acids Res.
41:D983–D986. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lian Y, Cai Z, Gong H, Xue S, Wu D and
Wang K: HOTTIP: A critical oncogenic long non-coding RNA in human
cancers. Mol biosyst. 12:3247–3253. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wang KC, Yang YW, Liu B, Sanyal A,
Corces-Zimmerman R, Chen Y, Lajoie BR, Protacio A, Flynn RA, Gupta
RA, et al: A long noncoding RNA maintains active chromatin to
coordinate homeotic gene expression. Nature. 472:120–124. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chang S, Liu J, Guo S, He S, Qiu G, Lu J,
Wang J, Fan L, Zhao W and Che X: HOTTIP and HOXA13 are oncogenes
associated with gastric cancer progression. Oncol Rep.
35:3577–3585. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Quagliata L, Matter MS, Piscuoglio S,
Arabi L, Ruiz C, Procino A, Kovac M, Moretti F, Makowska Z,
Boldanova T, et al: Long noncoding RNA HOTTIP/HOXA13 expression is
associated with disease progression and predicts outcome in
hepatocellular carcinoma patients. Hepatology. 59:911–923. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li Z, Zhao X, Zhou Y, Liu Y, Zhou Q, Ye H,
Wang Y, Zeng J, Song Y, Gao W, et al: The long non-coding RNA
HOTTIP promotes progression and gemcitabine resistance by
regulating HOXA13 in pancreatic cancer. J Transl Med. 13:842015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun Y, Zhou Y, Bai Y, Wang Q, Bao J, Luo
Y, Guo Y and Guo L: A long non-coding RNA HOTTIP expression is
associated with disease progression and predicts outcome in small
cell lung cancer patients. Mol Cancer. 16:1622017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Sun Y, Hu B, Wang Q, Ye M, Qiu Q, Zhou Y,
Zeng F, Zhang X, Guo Y and Guo L: Long non-coding RNA HOTTIP
promotes BCL-2 expression and induces chemoresistance in small cell
lung cancer by sponging miR-216a. Cell Death Dis. 9:852018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sun G, Alzayady K, Stewart R, Ye P, Yang
S, Li W and Shi Y: Histone demethylase LSD1 regulates neural stem
cell proliferation. Mol Cell Biol. 30:1997–2005. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tsai MC, Spitale RC and Chang HY: Long
intergenic noncoding RNAs: New links in cancer progression. Cancer
Res. 71:3–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Alves Pádua C, Fonseca AS, Muys BR, de
Barros E Lima, Bueno R, Bürger MC, de Souza JE, Valente V, Zago MA
and Silva WA Jr: Brief report: The lincRNA Hotair is required for
epithelial-to-mesenchymal transition and stemness maintenance of
cancer cell lines. Stem Cells. 31:2827–2832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yoon JH, Abdelmohsen K, Kim J, Yang X,
Martindale JL, Tominaga-Yamanaka K, White EJ, Orjalo AV, Rinn JL,
Kreft SG, et al: Scaffold function of long non-coding RNA
HOTAIR in protein ubiquitination. Nat Commun. 4:29392013.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Ono H, Motoi N, Nagano H, Miyauchi E,
Ushijima M, Matsuura M, Okumura S, Nishio M, Hirose T, Inase N, et
al: Long noncoding RNA HOTAIR is relevant to cellular
proliferation, invasiveness, and clinical relapse in small-cell
lung cancer. Cancer Med. 3:632–642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Fang S, Gao H, Tong Y, Yang J, Tang R, Niu
Y, Li M and Guo L: Long noncoding RNA-HOTAIR affects
chemoresistance by regulating HOXA1 methylation in small cell lung
cancer cells. Lab Invest. 96:60–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Young TL, Matsuda T and Cepko CL: The
noncoding RNA taurine upregulated gene 1 is required for
differentiation of the murine retina. Curr Biol. 15:501–512. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Huang MD, Chen WM, Qi FZ, Sun M, Xu TP, Ma
P and Shu YQ: Long non-coding RNA TUG1 is up-regulated in
hepatocellular carcinoma and promotes cell growth and apoptosis by
epigenetically silencing of KLF2. Mol Cancer. 14:1652015.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ma B, Li M, Zhang L, Huang M, Lei JB, Fu
GH, Liu CX, Lai QW, Chen QQ and Wang YL: Upregulation of long
non-coding RNA TUG1 correlates with poor prognosis and disease
status in osteosarcoma. Tumour Biol. 37:4445–4455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liu Q, Sun S, Yu W, Jiang J, Zhuo F, Qiu
G, Xu S and Jiang X: Altered expression of long non-coding RNAs
during genotoxic stress-induced cell death in human glioma cells. J
Neurooncol. 122:283–292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Han Y, Liu Y, Gui Y and Cai Z: Long
intergenic non-coding RNA TUG1 is overexpressed in urothelial
carcinoma of the bladder. J Surg Oncol. 107:555–559. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Xu Y, Wang J, Qiu M and Xu L, Li M, Jiang
F, Yin R and Xu L: Upregulation of the long noncoding RNA TUG1
promotes proliferation and migration of esophageal squamous cell
carcinoma. Tumour Biol. 36:1643–1651. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhang E, He X, Yin D, Han L, Qiu M, Xu T,
Xia R, Xu L, Yin R and De W: Increased expression of long noncoding
RNA TUG1 predicts a poor prognosis of gastric cancer and regulates
cell proliferation by epigenetically silencing of p57. Cell Death
Dis. 7:e21092016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yang L, Lin C, Liu W, Zhang J, Ohgi KA,
Grinstein JD, Dorrestein PC and Rosenfeld MG: ncRNA- and Pc2
methylation-dependent gene relocation between nuclear structures
mediates gene activation programs. Cell. 147:773–788. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Niu Y, Ma F, Huang W, Fang S, Li M, Wei T
and Guo L: Long non-coding RNA TUG1 is involved in cell growth and
chemoresistance of small cell lung cancer by regulating LIMK2b via
EZH2. Mol Cancer. 16:52017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ling H, Spizzo R, Atlasi Y, Nicoloso M,
Shimizu M, Redis RS, Nishida N, Gafà R, Song J, Guo Z, et al:
CCAT2, a novel noncoding RNA mapping to 8q24, underlies
metastatic progression and chromosomal instability in colon cancer.
Genome Res. 23:1446–1461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Gong WJ, Yin JY, Li XP, Fang C, Xiao D,
Zhang W, Zhou HH, Li X and Liu ZQ: Association of
well-characterized lung cancer lncRNA polymorphisms with lung
cancer susceptibility and platinum-based chemotherapy response.
Tumour Biol. 37:8349–8358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Xin Y, Li Z, Zheng H, Chan MTV and Ka Kei
Wu W: CCAT2: A novel oncogenic long non-coding RNA in human
cancers. Cell Prolif. 50:2017. View Article : Google Scholar
|
|
80
|
Redis RS, Sieuwerts AM, Look MP, Tudoran
O, Ivan C, Spizzo R, Zhang X, de Weerd V, Shimizu M, Ling H, et al:
CCAT2, a novel long non-coding RNA in breast cancer: Expression
study and clinical correlations. Oncotarget. 4:1748–1762. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Cai Y, He J and Zhang D: Long noncoding
RNA CCAT2 promotes breast tumor growth by regulating the Wnt
signaling pathway. Onco Targets Ther. 8:2657–2664. 2015.PubMed/NCBI
|
|
82
|
Huang S, Qing C, Huang Z and Zhu Y: The
long non-coding RNA CCAT2 is up-regulated in ovarian cancer and
associated with poor prognosis. Diagn Pathol. 11:492016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zheng J, Zhao S, He X, Zheng Z, Bai W,
Duan Y, Cheng S, Wang J, Liu X and Zhang G: The up-regulation of
long non-coding RNA CCAT2 indicates a poor prognosis for
prostate cancer and promotes metastasis by affecting
epithelial-mesenchymal transition. Biochem Biophys Res Commun.
480:508–514. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wang CY, Hua L, Yao KH, Chen JT, Zhang JJ
and Hu JH: Long non-coding RNA CCAT2 is up-regulated in gastric
cancer and associated with poor prognosis. Int J Clin Exp Pathol.
8:779–785. 2015.PubMed/NCBI
|
|
85
|
Chen S, Wu H, Lv N, Wang H, Wang Y, Tang
Q, Shao H and Sun C: LncRNA CCAT2 predicts poor prognosis
and regulates growth and metastasis in small cell lung cancer.
Biomed Pharmacother. 82:583–588. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Webb E, Adams JM and Cory S: Variant
(6;15) translocation in a murine plasmacytoma occurs near an
immunoglobulin kappa gene but far from the myc oncogene. Nature.
312:777–779. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Cory S, Graham M, Webb E, Corcoran L and
Adams JM: Variant (6;15) translocations in murine plasmacytomas
involve a chromosome 15 locus at least 72 kb from the c-myc
oncogene. EMBO J. 4:675–681. 1985.PubMed/NCBI
|
|
88
|
Colombo T, Farina L, Macino G and Paci P:
PVT1: A rising star among oncogenic long noncoding RNAs. Biomed Res
Int. 2015:3042082015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ding J, Li D, Gong M, Wang J, Huang X, Wu
T and Wang C: Expression and clinical significance of the long
non-coding RNA PVT1 in human gastric cancer. Onco Targets Ther.
7:1625–1630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang F, Yuan JH, Wang SB, Yang F, Yuan SX,
Ye C, Yang N, Zhou WP, Li WL, Li W, et al: Oncofetal long noncoding
RNA PVT1 promotes proliferation and stem cell-like property of
hepatocellular carcinoma cells by stabilizing NOP2. Hepatology.
60:1278–1290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhou Q, Chen J, Feng J and Wang J: Long
noncoding RNA PVT1 modulates thyroid cancer cell proliferation by
recruiting EZH2 and regulating thyroid-stimulating hormone receptor
(TSHR). Tumour Biol. 37:3105–3113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Huang C, Yu W, Wang Q, Cui H, Wang Y,
Zhang L, Han F and Huang T: Increased expression of the lncRNA PVT1
is associated with poor prognosis in pancreatic cancer patients.
Minerva Med. 106:143–149. 2015.PubMed/NCBI
|
|
93
|
Cui D, Yu CH, Liu M, Xia QQ, Zhang YF and
Jiang WL: Long non-coding RNA PVT1 as a novel biomarker for
diagnosis and prognosis of non-small cell lung cancer. Tumour Biol.
37:4127–4134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cui M, You L, Ren X, Zhao W, Liao Q and
Zhao Y: Long non-coding RNA PVT1 and cancer. Biochem Biophys Res
Commun. 471:10–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Huang C, Liu S, Wang H, Zhang Z, Yang Q
and Gao F: LncRNA PVT1 overexpression is a poor prognostic
biomarker and regulates migration and invasion in small cell lung
cancer. Am J Transl Res. 8:5025–5034. 2016.PubMed/NCBI
|
|
96
|
Sang Y, Zhou F, Wang D, Bi X, Liu X, Hao
Z, Li Q and Zhang W: Up-regulation of long non-coding HOTTIP
functions as an oncogene by regulating HOXA13 in non-small cell
lung cancer. Am J Transl Res. 8:2022–2032. 2016.PubMed/NCBI
|
|
97
|
Zhang GJ, Song W and Song Y:
Overexpression of HOTTIP promotes proliferation and drug resistance
of lung adenocarcinoma by regulating AKT signaling pathway. Eur Rev
Med Pharmacol Sci. 21:5683–5690. 2017.PubMed/NCBI
|
|
98
|
Yang Y, Jiang C, Yang Y, Guo L, Huang J,
Liu X, Wu C and Zou J: Silencing of LncRNA-HOTAIR decreases drug
resistance of non-small cell lung cancer cells by inactivating
autophagy via suppressing the phosphorylation of ULK1. Biochem
Biophys Res Commun. 497:1003–1010. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Liu W, Yin NC, Liu H and Nan KJ: Cav-1
promote lung cancer cell proliferation and invasion through lncRNA
HOTAIR. Gene. 641:335–340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Jiang C, Yang Y, Yang Y, Guo L, Huang J,
Liu X, Wu C and Zou J: Long noncoding RNA (lncRNA) HOTAIR affects
tumorigenesis and metastasis of non-small cell lung cancer by
up-regulating miR-613. Oncol Res. 26:725–734. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lin PC, Huang HD, Chang CC, Chang YS, Yen
JC, Lee CC, Chang WH, Liu TC and Chang JG: Long noncoding RNA
TUG1 is downregulated in non-small cell lung cancer and can
regulate CELF1 on binding to PRC2. BMC Cancer. 16:5832016.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhang EB, Yin DD, Sun M, Kong R, Liu XH,
You LH, Han L, Xia R, Wang KM, Yang JS, et al: P53-regulated long
non-coding RNA TUG1 affects cell proliferation in human non-small
cell lung cancer, partly through epigenetically regulating HOXB7
expression. Cell Death Dis. 5:e12432014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Liu H, Zhou G, Fu X, Cui H, Pu G, Xiao Y,
Sun W, Dong X, Zhang L, Cao S, et al: Long noncoding RNA TUG1 is a
diagnostic factor in lung adenocarcinoma and suppresses apoptosis
via epigenetic silencing of BAX. Oncotarget. 8:101899–101910.
2017.PubMed/NCBI
|
|
104
|
Zhao Z, Wang J, Wang S, Chang H, Zhang T
and Qu J: LncRNA CCAT2 promotes tumorigenesis by over-expressed
Pokemon in non-small cell lung cancer. Biomed Pharmacother.
87:692–697. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Qiu M, Xu Y, Yang X, Wang J, Hu J, Xu L
and Yin R: CCAT2 is a lung adenocarcinoma-specific long non-coding
RNA and promotes invasion of non-small cell lung cancer. Tumour
Biol. 35:5375–5380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wang C, Han C, Zhang Y and Liu F: LncRNA
PVT1 regulate expression of HIF1α via functioning as ceRNA for
miR199a5p in nonsmall cell lung cancer under hypoxia. Mol Med Rep.
17:1105–1110. 2018.PubMed/NCBI
|
|
107
|
Li H, Chen S, Liu J, Guo X, Xiang X, Dong
T, Ran P, Li Q, Zhu B, Zhang X, et al: Long non-coding RNA PVT1-5
promotes cell proliferation by regulating miR-126/SLC7A5 axis in
lung cancer. Biochem Biophys Res Commun. 495:2350–2355. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Guo D, Wang Y, Ren K and Han X: Knockdown
of LncRNA PVT1 inhibits tumorigenesis in non-small-cell lung cancer
by regulating miR-497 expression. Exp Cell Res. 362:172–179. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wu D, Li Y, Zhang H and Hu X: Knockdown of
lncrna PVT1 enhances radiosensitivity in non-small cell lung cancer
by sponging Mir-195. Cell Physiol Biochem. 42:2453–2466. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Smolle MA, Bauernhofer T, Pummer K, Calin
GA and Pichler M: Current insights into long non-coding RNAs
(lncRNAs) in prostate cancer. Int J Mol Sci. 18:E4732017.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yu X, Zheng H, Chan MT and Wu WK: HULC: An
oncogenic long non-coding RNA in human cancer. J Cell Mol Med.
21:410–417. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Gutschner T, Hämmerle M and Diederichs S:
MALAT1-a paradigm for long noncoding RNA function in cancer. J Mol
Med. 91:791–801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
McCleland ML, Mesh K, Lorenzana E, Chopra
VS, Segal E, Watanabe C, Haley B, Mayba O, Yaylaoglu M, Gnad F, et
al: CCAT1 is an enhancer-templated RNA that predicts BET
sensitivity in colorectal cancer. J Clin Invest. 126:639–652. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhou Y, Zhang X and Klibanski A:
MEG3 noncoding RNA: A tumor suppressor. J Mol Endocrinol.
48:R45–R53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Yap TA, Omlin A and de Bono JS:
Development of therapeutic combinations targeting major cancer
signaling pathways. J Clin Oncol. 31:1592–1605. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zhang SR, Yang JK, Xie JK and Zhao LC:
Long noncoding RNA HOTTIP contributes to the progression of
prostate cancer by regulating HOXA13. Cell Mol Biology. 62:84–88.
2016.
|
|
119
|
Sørensen KP, Thomassen M, Tan Q, Bak M,
Cold S, Burton M, Larsen MJ and Kruse TA: Long non-coding RNA
HOTAIR is an independent prognostic marker of metastasis in
estrogen receptor-positive primary breast cancer. Breast Cancer Res
Treat. 142:529–536. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Wu ZH, Wang XL, Tang HM, Jiang T, Chen J,
Lu S, Qiu GQ, Peng ZH and Yan DW: Long non-coding RNA HOTAIR is a
powerful predictor of metastasis and poor prognosis and is
associated with epithelial-mesenchymal transition Rep. 32:395–402.
2014.
|
|
121
|
Svoboda M, Slyskova J, Schneiderova M,
Makovicky P, Bielik L, Levy M, Lipska L, Hemmelova B, Kala Z,
Protivankova M, et al: HOTAIR long non-coding RNA is a negative
prognostic factor not only in primary tumors, but also in the blood
of colorectal cancer patients. Carcinogenesis. 35:1510–1515. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Huang L, Liao LM, Liu AW, Wu JB, Cheng XL,
Lin JX and Zheng M: Overexpression of long noncoding RNA HOTAIR
predicts a poor prognosis in patients with cervical cancer. Arch
Gynecol Obstet. 290:717–723. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Ji F, Wuerkenbieke D, He Y and Ding Y:
Long noncoding RNA HOTAIR: An oncogene in human cervical cancer
interacting with MicroRNA-17-5p. Oncol Res. 2017.Doi:
10.3727/096504017X15002869385155. View Article : Google Scholar :
|
|
125
|
Wu X, Cao X and Chen F: LncRNA-HOTAIR
activates tumor cell proliferation and migration by suppressing
MiR-326 in cervical cancer. Oncol Res. 2017.Doi:
10.3727/096504017X15037515496840. View Article : Google Scholar :
|
|
126
|
Endo H, Shiroki T, Nakagawa T, Yokoyama M,
Tamai K, Yamanami H, Fujiya T, Sato I, Yamaguchi K, Tanaka N, et
al: Enhanced expression of long non-coding RNA HOTAIR is associated
with the development of gastric cancer. PLoS One. 8:e770702013.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Hajjari M, Behmanesh M, Sadeghizadeh M and
Zeinoddini M: Up-regulation of HOTAIR long non-coding RNA in human
gastric adenocarcinoma tissues. Med Oncol. 30:6702013. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Chen WM, Chen WD, Jiang XM, Jia XF, Wang
HM, Zhang QJ, Shu YQ and Zhao HB: HOX transcript antisense
intergenic RNA represses E-cadherin expression by binding to EZH2
in gastric cancer. World J Gastroenterol. 23:6100–6110. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yan J, Dang Y, Liu S, Zhang Y and Zhang G:
LncRNA HOTAIR promotes cisplatin resistance in gastric cancer by
targeting miR-126 to activate the PI3K/AKT/MRP1 genes. Tumour Biol.
Nov 30–2016.(Epub ahead of print). View Article : Google Scholar :
|
|
130
|
Lin YH, Wu MH, Huang YH, Yeh CT, Cheng ML,
Chi HC, Tsai CY, Chung IH, Chen CY and Lin KH: Taurine upregulated
gene 1 functions as a master regulator to coordinate glycolysis and
metastasis in hepatocellular carcinoma. Hepatology. 67:188–203.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhao L, Sun H, Kong H, Chen Z, Chen B and
Zhou M: The lncrna-TUG1/EZH2 axis promotes pancreatic cancer cell
proliferation, migration and EMT phenotype formation through
sponging mir-382. Cell Physiol Biochem. 42:2145–2158. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Liu Q, Liu H, Cheng H, Li Y, Li X and Zhu
C: Downregulation of long noncoding RNA TUG1 inhibits proliferation
and induces apoptosis through the TUG1/miR-142/ZEB2 axis in bladder
cancer cells. Onco Targets Ther. 10:2461–2471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Yu Y, Nangia-Makker P, Farhana L and
Majumdar AP: A novel mechanism of lncRNA and miRNA interaction:
CCAT2 regulates miR-145 expression by suppressing its maturation
process in colon cancer cells. Mol Cancer. 16:1552017. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Hua F, Li CH, Chen XG and Liu XP: Long
noncoding RNA CCAT2 Knockdown suppresses tumorous progression by
sponging miR-424 in epithelial ovarian cancer. Oncol Res.
26:241–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Li T, Meng XL and Yang WQ: Long noncoding
RNA PVT1 acts as a ‘Sponge’ to inhibit microRNA-152 in gastric
cancer cells. Dig Dis Sci. 62:3021–3028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Huang T, Liu HW, Chen JQ, Wang SH, Hao LQ,
Liu M and Wang B: The long noncoding RNA PVT1 functions as a
competing endogenous RNA by sponging miR-186 in gastric cancer.
Biomed Pharmacother. 88:302–308. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Zhao L, Kong H, Sun H, Chen Z, Chen B and
Zhou M: LncRNA-PVT1 promotes pancreatic cancer cells proliferation
and migration through acting as a molecular sponge to regulate
miR-448. J Cell Physiol. 233:4044–4055. 2018. View Article : Google Scholar : PubMed/NCBI
|