Introduction
Colorectal cancer (CRC) is a malignancy derived from
the colorectal epithelium and is the third most commonly diagnosed
cancer type worldwide (1). Although
the mortality rates of CRC have been decreasing due to screening,
reduced risk factor prevalence and/or improved therapies (2,3), CRC
remains a global health burden in terms of morbidity and mortality,
with ~700,000 estimated mortalities annually (1). It has been reported that the
complicated and complex pathogenetic mechanisms of CRC involve
genomic rearrangements, chromatin remodeling, genetic mutations and
epigenetic changes (4,5).
The sphingolipid rheostat is a proposed concept that
may regulate cell fate decisions (6). The two major components of the
sphingolipid rheostat are ceramide and sphinogosine-1 phosphate,
which are interconvertible sphingolipid metabolites that regulate
cell growth and survival by modulating sphingolipid
rheostat-related signaling (6,7).
Ceramide has tumor suppressive anticancer properties, including
potentiating signaling networks that drive apoptosis, autophagy and
cell cycle arrest (8). Ceramide
synthases (CerSs) are integral membrane proteins of the endoplasmic
reticulum that synthesize ceramides of different acyl chain
lengths. To date, six CerS families have been identified in mammals
(9). Dysregulation of CerS activity
has been reported to be associated with tumor cell invasion
(10), proliferation (11), apoptosis (12) and epithelial-mesenchymal transition
(13), as well as with the
prognosis of patients with cancer (14). For example, in head and neck
squamous cell carcinoma, downregulation of CERS1 leads to
apoptotic resistance (15), while
CERS1 overexpression enhances growth-inhibitory effects
(16). Additionally, CERS2,
CERS4 and CERS6 mRNA expression levels are increased in
breast cancer (17), and the
upregulation of CERS4 and CERS6 leads to reduced cell
proliferation and the induction of apoptosis (18). Given these results and the
association of altered CerS expression with malignant
transformation, the present study aimed to characterize the mRNA
expression of various CerS genes in CRC and non-neoplastic adjacent
tissues (NST).
The present study investigated the mRNA expression
levels of various CerS genes using mRNA expression data from six
independent CRC cohorts and a Korean CRC dataset. Furthermore, the
clinical significance of altered CerS genes expression was
evaluated in the Korean CRC dataset.
Materials and methods
Gene expression databases and cluster
analysis
Gene expression RNAseq dataset (Level 3) and
clinical data for The Cancer Genome Atlas Colon and Rectal Cancer
(TCGA-COADREAD) cohort (19) were
downloaded from the UCSC Xena (https://xena.ucsc.edu). CRC gene expression microarray
data used in this study were downloaded from the publicly available
GEO databases (http://www.ncbi.nlm.nih.gov/geo/): GSE21815 (20), GSE44076 (21), GSE44861 (22), GSE41258 (23) and GSE33113 (24). The GEO datasets used in this study
include 562 CRC tissues and 222 NST from respective same patient
groups. The downloaded raw data of GEO databases were normalized at
the transcript and gene level using the Robust Multichip Average
method (25). Cluster analysis was
performed using Cluster 3.0 to classify the samples into
statistically similar groups, and the resulting heatmaps were
visualized in TreeView 1.6 (www.eisenlab.org/eisen). The four CerS genes present
in the TCGA COADREAD, GSE44076 and GSE44861 cohorts were LASS2,
LASS4, LASS5 and LASS6. The present study meets the publication
guidelines provided by TCGA.
Patients and tissues
A total of 59 patients (mean age, 64.83±9.48; age
range, 38–83; 34 males and 25 females) diagnosed with CRC were
included in the present study. CRC and NSTs were obtained from
patients undergoing surgery in Keimyung University Dongsan Medical
Center (Daegu, Korea) between April 2008 and January 2010. Enrolled
patients with CRC were classified according to the AJCC Tumor-Node
Metastasis (TNM) staging criteria (26). Tissue samples were immediately
frozen in liquid nitrogen and stored at −196°C until RNA isolation.
Tissue samples were provided by Keimyung Human Bio-Resource Bank
(Daegu, Korea). Written informed consent was obtained from each
study participant and the protocols were approved by the
Institutional Review Board of Keimyung University Dongsan Medical
Center (approval no. 2015-11-059-001).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total cellular RNA was extracted from tissues using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). RNA was quantified using NanoDrop 1000 (Thermo
Fisher Scientific, Inc.). Each cDNA was synthesized from 2 µg total
RNA using MMLV reverse transcriptase (Promega Corporation, Madison,
WI, USA), according to the manufacturer's protocol. qPCR was
performed on the LightCycler® 480 Real-Time PCR system
(Roche Diagnostics GmbH, Mannheim, Germany) using the specific
primer pairs presented in Table I
and SYBR-Green Premix (Toyobo Life Science, Osaka, Japan). The qPCR
was performed using the following thermocycling conditions: 95°C
for 10 min; followed by 45 cycles of 95°C for 10 sec, 60°C for 10
sec, and 72°C for 12 sec. Melting curve was analyzed to determine
primer specificity. b-actin was used as a housekeeping gene for
normalization, and a no-template sample was used as a negative
control. qPCR data were analyzed using the 2−∆∆Cq method
(27). Each experiment was
performed three times.
 | Table I.Primer sequences used in quantitative
polymerase chain reaction. |
Table I.
Primer sequences used in quantitative
polymerase chain reaction.
| Primer | Sequence |
|---|
| CERS2 sense |
5′-ATCGTCTTCGCCATTGTTTT-3′ |
| CERS2
antisense |
5′-GGCAGGATAGAGCTCCAGTG-3′ |
| CERS4 sense |
5′-GGAGGCCTGTAAGATGGTCA-3′ |
| CERS4
antisense |
5′-GAGGACCAGTCGGGTGTAGA-3′ |
| CERS5 sense |
5′-TGGAATTGGCCTTCTATTGG-3′ |
| CERS5
antisense |
5′-CAATGGTGACCAAGTGATGC-3′ |
| CERS6 sense |
5′-TGCCATTCTGGAAAAGGTCT-3′ |
| CERS6
antisense |
5′-ATGCTTCGAACATCCCAGTC-3′ |
| β-actin sense |
5′-CAGCCATGTACGTTGCTATCCAGG-3′ |
| β-actin
antisense |
5′-AGGTCCAGACGCAGGATGGCATG-3′ |
Statistical analysis
Statistical analysis was performed using SPSS 22.0
(IBM Corp., Armonk, NY, USA). The cell viability data were analyzed
using one-way analysis of variance and the Student-Newman-Keuls
post hoc test. Differences between the groups were analyzed
statistically using Student's t-test or Mann Whitney U test. The
co-expression of the mRNAs of various CerS genes in TCGA-COADREAD
cohort were searched using cBioPortal (http://cbioportal.org) (28). The association between
inter-individual mRNA expression levels of CerS genes in Korean
patients with CRC was assessed using Pearson's correlation
coefficient analysis for continuous variables. Clinicopathological
associations with the mRNA expression levels of various CerS genes
in Korean CRC were analyzed using the Linear by linear association,
the Pearson's Chi-square test and the Fisher's exact test for
categorical variables. The mean value was used as the cut-off value
(low and high) for categorical variables. P<0.05 was considered
to indicate a statistically significant difference.
Transient transfection
Various human colorectal adenocarcinoma cell lines,
HCT116, HT29, SW403 and SW480 cells, were plated onto 6-well plates
at a density 7×105 cells/well and cultured overnight.
pcDNA3.1-empty vector was used for plasmid constructs, including
HA-tagged form of CERS2 (HA-CERS2) and HA-tagged form of CERS6
(HA-CERS6) constructs. All plasmids, including pcDNA3.1-empty
vector, HA-CERS2 and HA-CERS6 were provided by Professor Anthony H.
Futerman (Weizmann Institute of Science, Rehovot, Israel). The CRC
cells were transfected with pcDNA3.1-empty vector, 2 µg HA-CERS2
and HA-CERS6 plasmid in 6-well plates using Lipofectamine reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. At 24 h after plasmid transfection, the
subsequent experiments were conducted.
Western blot analysis
The transient transfected CRC cells were collected
and washed twice with cold PBS, and cell pellets were prepared by
suspending in modified radioimmunoprecipitation assay buffer (50 mM
Tris-HCl pH 7.4, 1% NP-40, 0.25% Na-deoxycholate, 150 mM NaCl, 1 mM
Na3VO4 and 1 mM NaF) containing protease
inhibitors (100 µM phenylmethylsulfonyl fluoride, 10 µg/ml
leupeptin, 10 µg/ml pepstatin and 2 mM EDTA). The lysates were
centrifuged at 10,000 × g for 10 min at 4°C, and the supernatant
fractions were collected. The total protein concentration was
measured using Micro BCA™ Protein assay kit (Thermo Fisher
Scientific, Inc.), according to the manufacturers protocol.
Cellular proteins (60 mg) were mixed with protein 5X sample buffer
(Elpis Biotech., Inc., Daejeon, Korea) and heated at 95°C for 5
min. The proteins were separated by 10% SDS-PAGE and then
electrotransferred to Immobilon-P membranes (EMD Millipore,
Billerica, MA, USA). The membranes were then blocked at room
temperature with 5% skimmed dried milk in PBS/0.1% Tween-20 for 1
h, and incubated overnight at 4°C with anti-HA (1:2,000; mouse
monoclonal; cat. no. SAB1411737) and anti-β-actin (1:2,000; mouse
monoclonal; cat. no. A5441; both Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). The membranes were then washed six times with
PBS/0.1% Tween-20 (30 min each) and incubated with the
corresponding secondary antibodies (horseradish
peroxidase-conjugated, horse antibodies to mouse IgG; 1:2,000; cat.
no. 7076; Cell Signaling Technology, Inc.) for 1 h at room
temperature. Following washing six times in PBS/0.1% Tween-20, the
specific protein bands were detected using an enhanced
chemiluminescence western blotting kit (EMD Millipore), according
to the manufacturer's protocol.
Results
Altered expression levels of
sphingolipid metabolism-related genes in six independent CRC
cohorts
To investigate whether the sphingolipid
metabolism-related genes (29) are
dysregulated in CRC tissues, the present study re-analyzed the raw
data of six independent CRC cohorts. To begin with, the cancer gene
expression RNAseq datasets of 380 CRC patients were taken from the
TCGA-COADREAD cohort through UCSC Xena. Next, CRC gene expression
microarray data were downloaded from the publicly available Gene
Expression Omnibus databases. The CRC gene expression microarrays,
GSE21815, GSE33113, GSE41258, GSE44076 and GSE44861, were analyzed
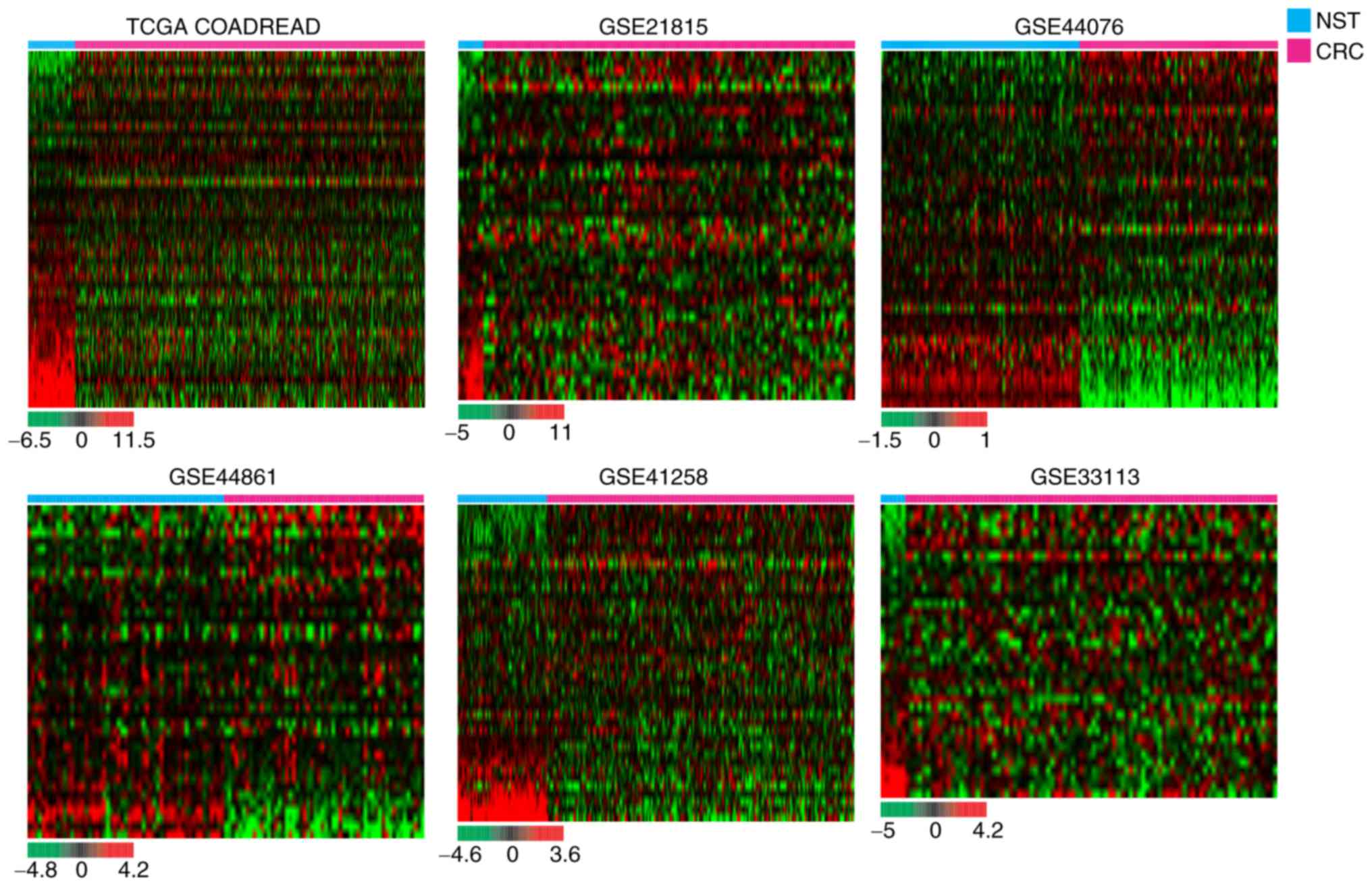
for potential transcriptome changes. Hierarchical clustering
revealed that various sphingolipid metabolism-related genes were
dysregulated in carcinomatous tissues compared with NST of patients
with CRC (Fig. 1). To identify the
significance of altered mRNA expression levels between CRC and NST,
Student's t-test or Mann Whitney U test were performed (P<0.05).
As demonstrated in Fig. 2,
hierarchical clustering revealed that various sphingolipid
metabolism-related genes were significantly dysregulated in CRC
tissues compared with NST from the same patient groups. The list of
analyzed sphingolipid metabolism-related genes is presented in
Table II. Sphingosine kinase 1
(SPHK1) and UDP-glucose glycoprotein glucosyltransferase 2
(UGGT2) were significantly upregulated in the CRC tissues of
all cohorts, while 15-hydroxyprostaglandin dehydrogenase
(HPGD), lysophosphatidic acid receptor 1 (LPAR1),
N-acylethanolamine acid amidase (NAAA), sphingomyelin
phosphodiesterase 1 (SMPD1) and sphingomyelin
phosphodiesterase acid-like 3A (SMPDL3A) were significantly
downregulated in the CRC tissues of all cohorts (Fig. 2 and Table II).
| Table II.List of the analyzed genes involved
in sphingolipid metabolism (Student's t-test, Mann Whitney U test;
P<0.05). |
Table II.
List of the analyzed genes involved
in sphingolipid metabolism (Student's t-test, Mann Whitney U test;
P<0.05).
| Dataset |
TCGA-COADREADa |
GSE21815b |
GSE44076a |
GSE44861a |
GSE41258a |
GSE33113b |
|---|
| No. analyzed
genes | 36 | 20 | 35 | 19 | 24 | 15 |
| Gene symbol | A4GALT | B4GALT6 | ASAH1 | ASAHL (NAAA) | ASAH1 | CERS5 |
|
| ASAH1 | BECN1 | B4GALT6 | B4GALT6 | BECN1 | CERS6 |
|
| B4GALNT1 | CERK | BECN1 | BNIP3 | CERK | HPGD |
|
| B4GALT6 | CERS2 | CERK | CERK | CERS2 | LPAR1 |
|
| BECN1 | CERS5 | DEGS1 | DEGS1 | CERS6 | NAAA |
|
| CERK | DEGS1 | GAL3ST1 | EDG2 (LPAR1) | GAL3ST1 | NSMAF |
|
| GAL3ST1 | GBA | GBA | EDG5 (S1PR2) | HPGD | SFTPB |
|
| GALC | HPGD | HPGD | HPGD | LCT | SMPD1 |
|
| GBA | LPAR1 | LASS1 | CERS2 | LPAR1 | SMPDL3A |
|
| HPGD | NAAA | LASS2 | NSMAF | LPAR2 | SMPDL3B |
|
| LASS1 | S1PR4 | LASS5 | SMPD1 | NAAA | SPHK1 |
|
| LASS2 | SLC26A10 | LASS6 | SMPD2 | NSMAF | SPTLC1 |
|
| LASS3 | SMPD1 | LCT | SMPDL3A | S1PR1 | SPTLC2 |
|
| LASS4 | SMPDL3A | LPAR1 | SMPDL3B | SLC26A10 | ST6GALNAC5 |
|
| LASS5 | SMPDL3B | LPAR2 | SPHK1 | SMPD1 | UGGT2 |
|
| LASS6 | SPHK1 | NAAA | SPHK2 | SMPDL3A |
|
|
| LPAR1 | ST3GAL5 | NSMAF | SPTLC1 | SMPDL3B |
|
|
| LPAR2 | ST8SIA1 | S1PR1 | ST6GALNAC5 | SPHK1 |
|
|
| NAAA | UGGT1 | S1PR4 | UGCGL2 (UGGT2) | SPHK2 |
|
|
| NSMAF | UGGT2 | SFTPB |
| ST3GAL5 |
|
|
| S1PR1 |
| SLC26A10 |
| ST8SIA1 |
|
|
| S1PR4 |
| SMPD1 |
| UGCG |
|
|
| SLC26A10 |
| SMPD2 |
| UGGT1 |
|
|
| SMPD1 |
| SMPDL3A |
| UGGT2 |
|
|
| SMPDL3A |
| SMPDL3B |
|
|
|
|
| SMPLL3B |
| SPHK1 |
|
|
|
|
| SPHK1 |
| SPHK2 |
|
|
|
|
| SPHK2 |
| SPTLC1 |
|
|
|
|
| SPTLC2 |
| SPTLC2 |
|
|
|
|
| ST3GAL5 |
| ST3GAL5 |
|
|
|
|
| ST6GALNAC5 |
| ST8SIA1 |
|
|
|
|
| ST8SIA1 |
| ST8SIA3 |
|
| ST8SIA3 |
| UGCG |
|
|
|
|
| UGCG |
| UGGT1 |
|
|
|
|
| UGGT1 |
| UGGT2 |
|
|
|
|
| UGGT2 |
|
|
|
|
|
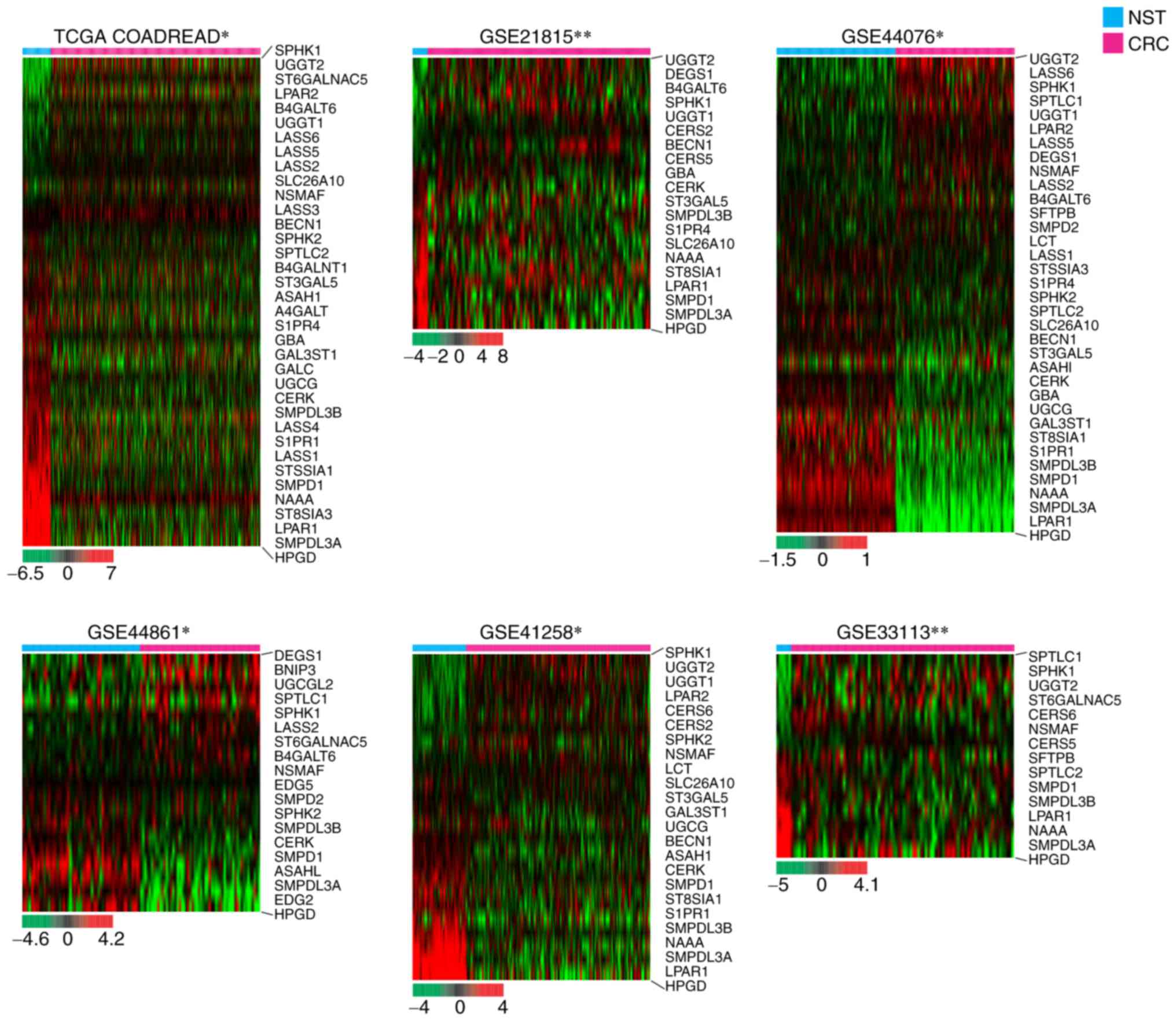
Dysregulation of various CerSs in six
independent CRC cohorts
Next, the present study evaluated whether the mRNA
expression levels of the four CerS genes, which are abundant in
colorectal tissues (30), are
dysregulated in human CRC specimens with respect to NST. As
demonstrated in Fig. 3, among six
cohorts, CERS2 mRNA levels were significantly increased in
five independent cohorts, while CERS5 and CERS6 were
significantly upregulated in four independent cohorts. The specific
platforms of each cohort and their associated studies are listed in
Table III.
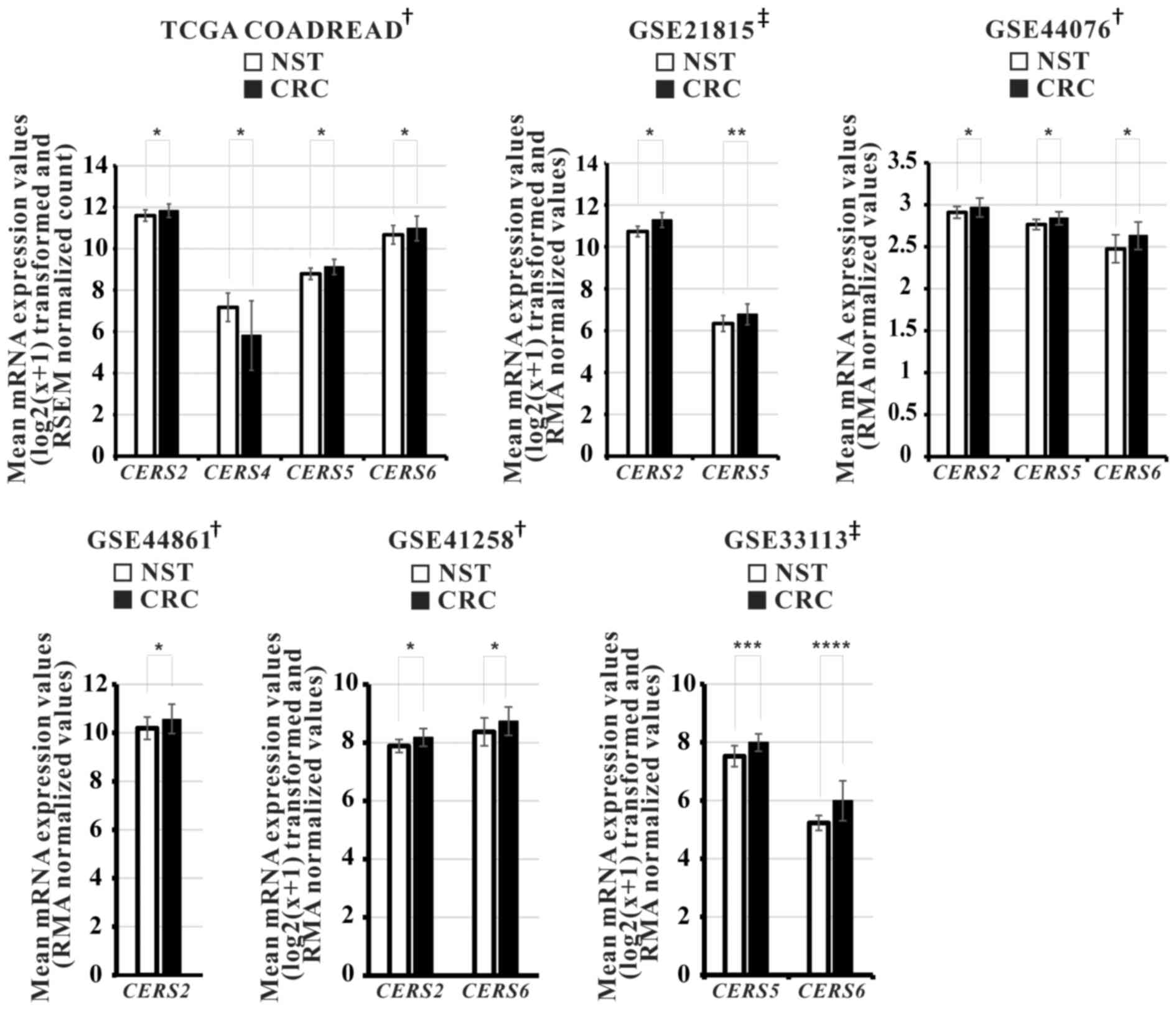 | Figure 3.Relative mRNA expression levels of
various ceramide synthases in the 6 independent CRC cohorts. Gene
expression profiling datasets TCGA-COADREAD (NST, n=51; CRC,
n=380), GSE21815 (NST, n=9; CRC, n=132), GSE44076 (NST, n=98; CRC,
n=98), GSE44861 (NST, n=55; CRC, n=56), GSE41258 (NST, n=54; CRC,
n=186) and GSE33113 (NST, n=6; CRC, n=90). P-values were calculated
using †Student's t-test and ‡Mann Whitney U
test, *P<0.001, **P=0.005, ***P=0.002, ****P=0.006. CRC,
colorectal cancer; NST, non-neoplastic surrounding colon tissues;
TCGA-COADREAD, The Cancer Genome Atlas Colon and Rectal Cancer. |
| Table III.mRNA expression levels of CerS gene
in colorectal cancer tissues of patients from various datasets used
in the present study. |
Table III.
mRNA expression levels of CerS gene
in colorectal cancer tissues of patients from various datasets used
in the present study.
| Dataset | Platform | CERS2 | CERS4 | CERS5 | CERS6 | (Refs.) |
|---|
| TCGA
(COADREAD) | RNA sequencing | Up | Down | Up | Up | (19) |
| GSE21815 | Human Whole Genome
Microarray 4×4K G4112F | Up | N/A | Up | N/A | (20) |
| GSE44076 | Affymetrix Human
Genome U219 Array | Up | N/A | Up | Up | (21) |
| GSE44861 | Affymetrix HT Human
Genome U133A Array | Up | N/A | N/A | N/A | (22) |
| GSE41258 | Affymetrix U133A
Array | Up | N/A | N/A | Up | (23) |
| GSE33113 | Affymetrix Human
Genome U133 Plus 2.0 Array | N/A | N/A | Up | Up | (24) |
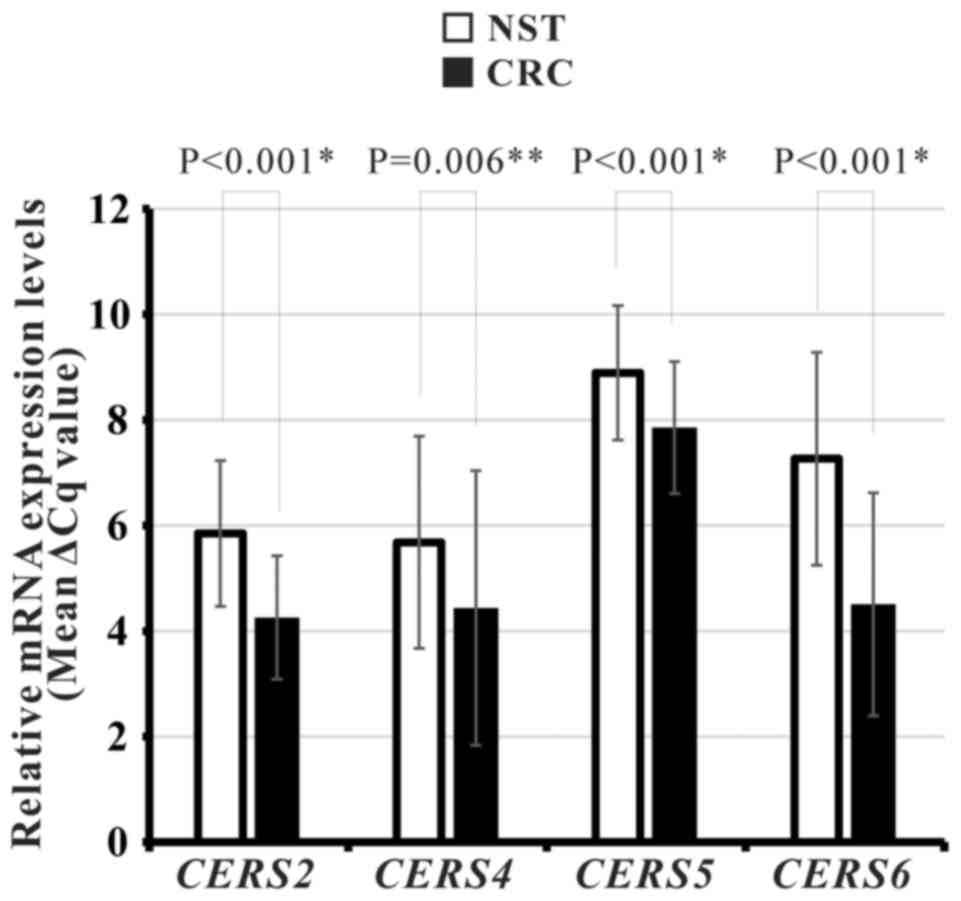
Altered CerS genes mRNA expression in
Korean patients with CRC
To determine whether there is altered CERS2,
CERS4, CERS5 and/or CERS6 mRNA expression in Korean
patients with CRC, the expression levels of these four CerSs were
measured using qPCR in 59 paired CRC and NST specimens from Korean
patients. Following exclusion of unqualified results, the qPCR data
were analyzed. The present study revealed that mRNA expression
levels of all four CerS genes were significantly upregulated in CRC
tissues compared with corresponding NSTs (CERS2, P<0.001;
CERS4, P=0.006; CERS5, P<0.001; CERS6,
P<0.001; Fig. 4; Table IV).
| Table IV.mRNA expression levels of CerS gene
in CRC tissues as compared with NST of Korean patients with
CRC. |
Table IV.
mRNA expression levels of CerS gene
in CRC tissues as compared with NST of Korean patients with
CRC.
| CERS
family |
| CERS2 |
|
| CERS4 |
|
| CERS5 |
|
| CERS6 |
|
|---|
| No. patients |
| 59 |
|
| 55 |
|
| 49 |
|
| 55 |
|
| Type of tissue | NST |
| CRC | NST |
| CRC | NST |
| CRC | NST |
| CRC |
| Mean (ΔCq
value) | 5.85 |
| 4.26 | 5.68 |
| 4.43 | 8.89 |
| 7.85 | 7.26 |
| 4.51 |
| Regulation |
| Up |
|
| Up |
|
| Up |
|
| Up |
|
| P-value |
| <0.001 |
|
| 0.06 |
|
| <0.001 |
|
| <0.001 |
|
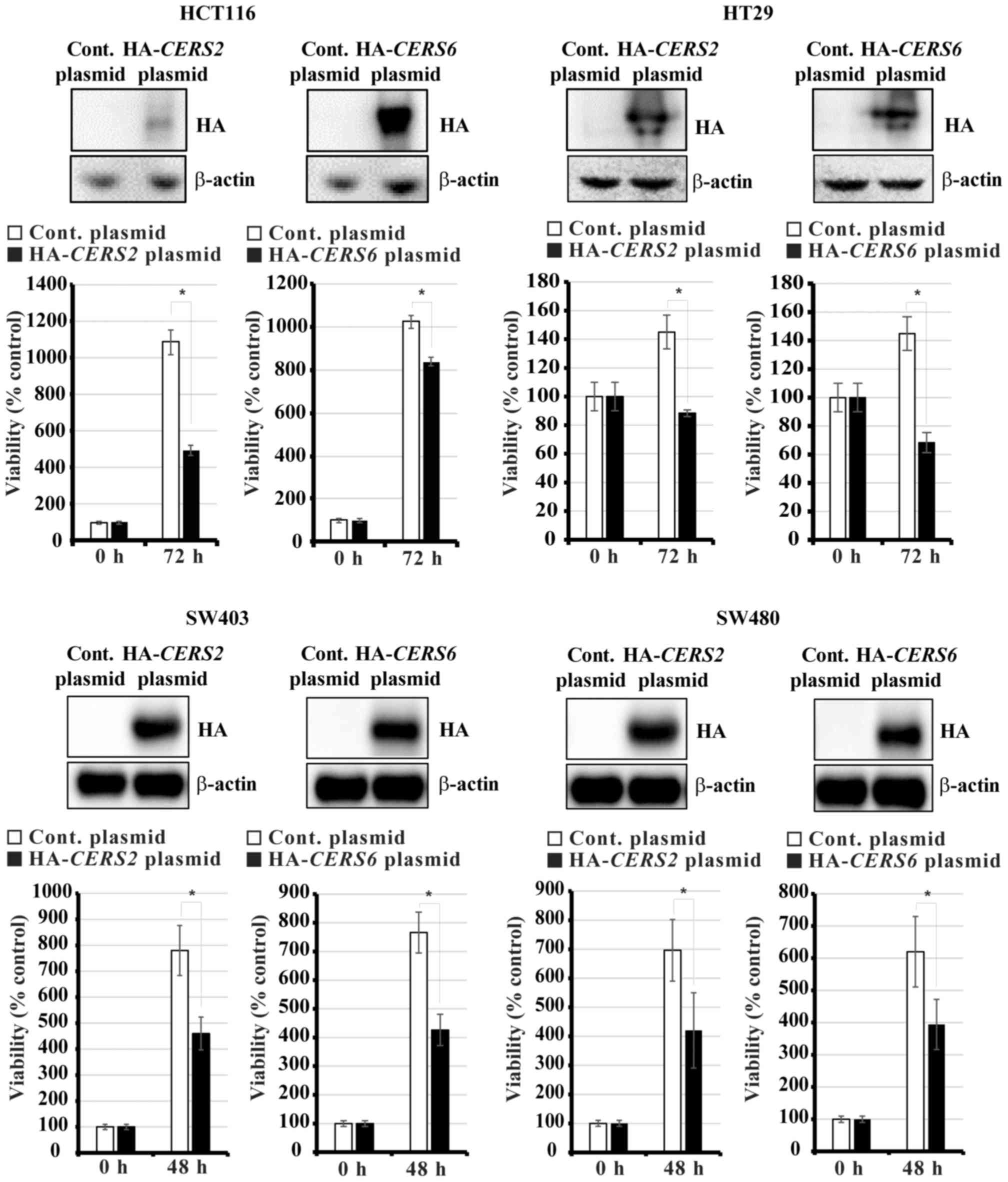
Exogenous CERS2 and CERS6 expression
decreases the viability of human CRC cells
It has previously been observed that
CERS6-overexpression reduces the proliferation of CRC cells
and induces apoptosis, whereas CERS2-overexpression
increases the proliferation of CRC cells (18). To confirm the effect of
overexpressing CerSs in CRC cells, HCT116, HT29, SW403 and SW480
cells were transiently transfected with constructs to overexpress
HA-CERS2 and HA-CERS6, respectively. After 48 and 72
h, the numbers of viable cells were counted using a hemocytometer.
As demonstrated in Fig. 5,
overexpression of CERS2 and CERS6 decreased the
viability of this panel of CRC cell lines.
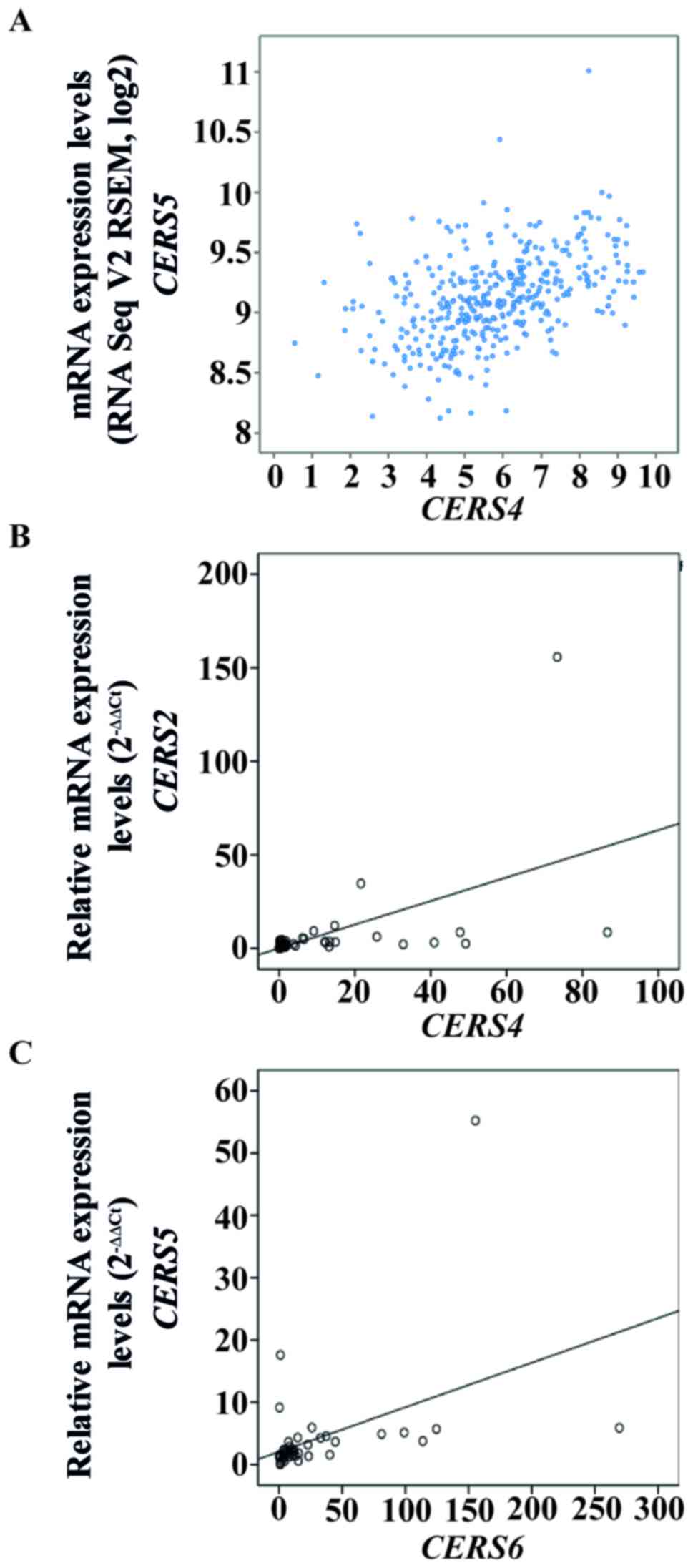
Inter-individual associations between
mRNA expression levels of CerS genes in patients with CRC
Combinational patterns of CerS gene expression,
including CerS hetero-complexes and co-expression of CerS genes,
serve important roles in sphingolipid metabolism (31,32).
Therefore, associations between the mRNA levels of each CerS gene
were identified in the TCGA-COADREAD cohort and in the Korean CRC
cohort. Using cBioPortal to analyze the TCGA-COADREAD cohort,
co-expression analysis revealed that CERS4 and CERS5
had high correlation coefficients (Pearson's correlation=0.36;
Spearman's correlation=0.48; Fig.
6A). Next, these correlations were assessed using Pearson's
correlation coefficient analysis in the 59 Korean patients with
CRC. There were significant correlations between CERS2 and
CERS4, and also between CERS5 and CERS6, with
a Pearson's correlation coefficient value of 0.532 (P<0.001;
Fig. 6B) and 0.439 (P=0.003;
Fig. 6C), respectively.
Furthermore, significant correlations between the mRNA expression
levels of CERS2 and CERS4 (P=0.009) and of
CERS5 and CERS6 (P<0.001) were identified using
Fisher's exact test (Table V).
| Table V.Association between mRNA expression
levels of various CerS genes and clinicopathological parameters in
Korean patients with colorectal cancer. |
Table V.
Association between mRNA expression
levels of various CerS genes and clinicopathological parameters in
Korean patients with colorectal cancer.
|
| CERS2
expression | CERS4
expression | CERS5
expression | CERS6
expression |
|---|
|
|
|
|
|
|
|---|
| Parameter | Low | High | P-value | Low | High | P-value | Low | High | P-value | Low | High | P-value |
|---|
| Sex |
|
| 1.000b |
|
| 0.195b |
|
| 0.719b |
|
| 1.000b |
|
Male | 24 | 4 |
| 17 | 11 |
| 20 | 8 |
| 22 | 6 |
|
|
Female | 14 | 2 |
| 13 | 3 |
| 13 | 3 |
| 13 | 3 |
|
| Age, years |
|
| 0.606b |
|
| 1.000b |
|
| 0.408b |
|
| 0.659b |
|
≤50 | 8 | 2 |
| 7 |
| 3 | 9 | 1 |
| 9 | 1 |
|
|
>50 | 30 | 4 |
| 23 | 11 |
| 24 | 10 |
| 26 | 8 |
|
| T stage |
|
| 0.609c |
|
| 0.675c |
|
| 0.457c |
|
| 0.140c |
| T1 | 2 | 0 |
| 2 | 0 |
| 0 | 2 |
| 0 | 2 |
|
| T2 | 7 | 1 |
| 4 | 4 |
| 8 | 0 |
| 7 | 1 |
|
| T3 | 24 | 4 |
| 21 | 7 |
| 20 | 8 |
| 23 | 5 |
|
| T4 | 5 | 1 |
| 3 | 3 |
| 5 | 1 |
| 5 | 1 |
|
| N stage |
|
| 0.063c |
|
| 0.061c |
|
| 0.055c |
|
| 0.288c |
| N0 | 25 | 2 |
| 21 | 6 |
| 18 | 9 |
| 21 | 6 |
|
| N1 | 7 | 1 |
| 5 | 3 |
| 6 | 2 |
| 5 | 3 |
|
| N2 | 6 | 3 |
| 4 | 5 |
| 9 | 0 |
| 9 | 0 |
|
| N3 | 0 | 0 |
| 0 | 0 |
| 0 | 0 |
| 0 | 0 |
|
| M stage |
|
| 0.456b |
|
| 0.581b |
|
| 0.558b |
|
| 0.566b |
|
Negative | 35 | 5 |
| 28 | 12 |
| 29 | 11 |
| 31 | 9 |
|
|
Positive | 3 | 1 |
| 2 | 2 |
| 4 | 0 |
| 4 | 0 |
|
| BMI |
|
| 0.653c |
|
| 0.320c |
|
| 0.593c |
|
| 0.566c |
|
≤18.5 | 0 | 0 |
| 0 | 0 |
| 0 | 0 |
| 0 | 0 |
|
|
18.5–24.9 | 1 | 0 |
| 1 | 0 |
| 1 | 0 |
| 1 | 0 |
|
|
25-29.9 | 27 | 4 |
| 19 | 12 |
| 22 | 9 |
| 25 | 6 |
|
|
>30 | 10 | 2 |
| 10 | 2 |
| 10 | 2 |
| 9 | 3 |
|
| CEA |
|
| 1.000b |
|
| 0.540a |
|
| 0.706b |
|
| 0.703b |
|
>5 | 11 | 2 |
| 8 | 5 |
| 9 | 4 |
| 11 | 2 |
|
| ≤5 | 27 | 4 |
| 22 | 9 |
| 24 | 7 |
| 24 | 7 |
|
| CERS2 |
|
|
|
|
| 0.009b |
|
| 0.630b |
|
| 1.000b |
|
Low |
|
|
| 29 | 9 |
| 29 | 9 |
| 30 | 8 |
|
|
High |
|
|
| 1 | 5 |
| 4 | 2 |
| 5 | 1 |
|
| CERS4 |
|
| 0.009b |
|
|
|
|
| 0.456b |
|
| 0.233b |
|
Low | 29 | 1 |
|
|
|
| 21 | 9 |
| 22 | 8 |
|
|
High | 9 | 5 |
|
|
|
| 12 | 2 |
| 13 | 1 |
|
| CERS5 |
|
| 0.630b |
|
| 0.456b |
|
|
|
|
|
<0.001b |
|
Low | 29 | 4 |
| 21 | 12 |
|
|
|
| 31 | 2 |
|
|
High | 9 | 2 |
| 9 | 2 |
|
|
|
| 4 | 7 |
|
| CERS6 |
|
| 1.000b |
|
| 0.233b |
|
|
<0.001b |
|
|
|
|
Low | 30 | 5 |
| 22 | 13 |
| 31 | 4 |
|
|
|
|
|
High | 8 | 1 |
| 8 | 1 |
| 2 | 7 |
|
|
|
|
Association between mRNA expression
levels of CerS genes and clinicopathological parameters of Korean
patients with CRC
To determine the clinicopathological implications of
dysregulated expression of specific CerS genes in CRC, the
association between CerS gene mRNA level and clinicopathological
characteristics, which are used to represent progression and
aggressiveness, were evaluated. Prior to the statistical analysis,
the 44 patients, whose clinical data were available, were
classified according to each clinicopathological characteristic
(Table V). The results obtained
from the statistical analysis of the Korean cohort revealed that
altered mRNA expression levels of CerS genes were not significantly
associated with any clinical parameters, including sex, age,
Tumor-Node-Metastasis stage, body mass index or carcinoembryonic
antigen titer.
Discussion
Sphingolipid metabolism serves a critical role in
mammalian cell growth arrest and survival (33). Accumulating evidence have
demonstrated that CerS, a major component in sphingolipid
metabolism (7), regulates various
biological phenomenon, including apoptosis (34), cancer (17,35),
ER stress (36), hepatopathy
(37), hypoxia/re-oxygenation
injury (38), lipid metabolism
(39), neurodegeneration (40), and sensitivity to chemotherapeutic
drugs and radiation (30). Although
aberrant CerS expression is correlated with cell death and
proliferation (10–14) in various types of cancer, much
uncertainty remains regarding the dysregulated mRNA levels of CerS
gene in CRC and the clinical implications of this.
The aims of the present study were to investigate
the mRNA expression levels and functions of CerS genes, which are
primarily expressed in the intestine (30,41),
and analyze their clinicopathological implications in patients with
CRC. To begin with, significantly dysregulated sphingolipid
metabolism-related genes were identified in the heat-maps of 6
independent CRC cohorts (Fig. 2).
The hierarchical clustering results demonstrated considerable
dysregulation of sphingolipid metabolism-related genes in CRC
tissues compared with corresponding NST of independent CRC cohorts.
Among the considerably altered genes, certain genes were
overlapping over 6 independent cohorts. SPHK1 and
UGGT2 were significantly upregulated in CRC tissues. This
result is in accordance with those of recent studies that indicated
that SPHK1 is overexpressed and serves an important role in
tumorigenesis, proliferation, invasiveness and metastasis in CRC
(42,43). On the other hand, HPGD, LPAR1,
NAAA, SMPD1 and SMPDL3A were all significantly
downregulated in CRC tissues. HPGD is a cytoplasmic enzyme
responsible for degrading PGE2 in colorectal tissue (44), and functions as a tumor suppressor
gene in various types of cancer (45–48).
The present study observed that HPGD was downregulated in 6
independent CRC cohorts (Fig. 2).
However, little is known regarding the cellular functions and
clinicopathological implications of LPAR1, NAAA, SMPD1,
SMPDL3A and UGGT2 in CRC. Therefore, further studies
investigating the functional role of these genes in CRC are
required as these transcripts may be diagnostic markers or
promising therapeutic candidates.
Additionally, the differential mRNA levels of
CERS2, CERS4, CERS5 and CERS6 in CRC and NST were
analyzed in 1,001 patients with CRC from 6 independent
publicly-available CRC cohorts and a cohort of Korean patients with
CRC. The results of the present study should be interpreted with
caution as qPCR, RNA-Seq and microarray are different experimental
platforms with different sensitivities, principles and dynamic
ranges. Nonetheless, the results revealed that CERS2 was
significantly upregulated in the majority of cohorts (Fig. 3; Table
III) and in the cohort of Korean patients with CRC (Fig. 4; Table
IV). It was recently demonstrated that
CERS2-overexpression had no effect on the viability of
HCT116 cells, whereas overexpressing CERS2 plus the addition
of very-long chain acyl-CoAs significantly enhanced colony
formation in HCT116 cells (18).
Unlike these previous results, the present study revealed that
CERS2-overexpression reduced the viability of various human
CRC cells, including HCT116 cells (Fig.
5). The primary technical differences between these two
experiments are the culture time following transfection and the use
of different expression plasmids. Additionally, knockdown
experiments were performed using shRNAs against CERS2 mRNA
and CERS6 mRNA, knockdown of CerS2 or CerS6 did not affect
the proliferation of CRC SW403 and SW480 cells (data not shown).
Although the exact mechanism that underlies the effect on cell
viability was not elucidated in the present study, it is possible
that sustained cell culture time following transfection may affect
the synthesis of ceramides of various chain lengths.
It has been reported that increased expression of
CerS6 and C16:0-Ceramide resulted in a sensitization of SW620 cells
to TRAIL-induced apoptosis (49),
and CERS6-overexpression significantly inhibited the colony
formation capacity and increased the apoptosis of HCT116 cells
(18). In accordance with these
previous results, the present study demonstrated that CERS6
is significantly upregulated in CRC tissues, compared with NST
(Figs. 3 and 4; Table
IV) and CERS6-overexpression led to inhibition of cell
viability in various human CRC cells (Fig. 5). Notably, controversial results
have demonstrated that CerS6 and C16:0-Ceramide protected cells
against ER-stress in human head and neck squamous cell carcinomas
(36). Although oncogenes are
usually upregulated in cancer tissues compared with non-neoplastic
tissues, previous studies and the results presented in the present
study indicated that the roles of CerSs and ceramides of specific
chain lengths are complicated and cell type-dependent. Notably, it
was demonstrated that CERS4 was significantly upregulated,
but only in the cohort of Korean patients with CRC (Fig. 4), while it was downregulated in the
TCGA-COADREAD cohort (Fig. 3).
Future studies specifically focused on CERS4 in different
CRC populations are required in order to understand this
phenomena.
Additionally, the present study evaluated
correlations between inter-individual mRNA expression levels of
CerS genes and their clinicopathological implications in patients
with CRC. A recent study revealed that inter-individual differences
in the mRNA expression levels of CerS genes are significantly
correlated with each other in cancer tissues (17). Furthermore, Combinational patterns
of CerS expression are involved in sphingolipid metabolism
(31,32). The results of analyzing correlations
between inter-individual CerS genes mRNA expression levels revealed
a correlation between CERS4 and CERS5 in
TCGA-COADREAD, between CERS2 and CERS4, and between
CERS5 and CERS6 in the cohort of Korean patients with
CRC. However, combinational patterns of CerS expression may be
associated with sphingolipid metabolism. Therefore, it will be
important to determine which components serve critical roles in
sphingolipid metabolism in different disease and tissue settings.
To the best of our knowledge, the present study was the first to
investigate the clinicopathological implications of dysregulated
CerS genes mRNA expression in CRC. However, no correlation was
observed between mRNA expression levels of specific CerS genes and
the investigated clinicopathological parameters.
In conclusion, the present study revealed that the
mRNA expression levels of CERS2, CERS4, CERS5 and
CERS6 were significantly upregulated or downregulated in
various independent CRC cohorts, suggesting that dysregulated CerS
gene expression may serve a role in CRC development.
Acknowledgements
The authors would like to thank all members of their
research group for providing enthusiastic participation in the
present study. The biospecimens for the present study were provided
by the Keimyung Human Bio-Resource Bank, a member of the National
Biobank of Korea, which is supported by the Ministry of Health and
Welfare. All samples derived from the National Biobank of Korea
were obtained following receipt of written informed consent and
Institutional Review Board approval.
Funding
The present study was supported by the National
Research Foundation of Korea Grant funded by the Korean Government
(Ministry of Science, ICT & Future Planning; grant nos.
2017R1C1B5016670 and 2014R1A5A2010008).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable request.
The primary and processed data used to generate the analyses
presented here can be downloaded by registered users from The
Cancer Genome Atlas at http://tcga-data.nci.nih.gov/tcga/tcgaDownload.jsp.
Authors' contributions
SWJ, WJP and SK contributed to the conception and
design of the study, analysis of the data, interpretation of
results and the writing of the manuscript. SWJ, WJP, HM and SK
contributed to the acquisition of data. SWJ, WJP, HM, SKB, IH and
SK performed the experiments. TKK contributed to the conception and
design of the study. JWP and IH contributed to the conception and
design of the study and provided guidance regarding the clinical
implications of the study. WJP and SK reviewed and edited the
manuscript. All authors read and approved the manuscript, and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The experimental study was approved by the
Institutional Review Board of Keimyung University Dongsan Medical
Center (approval no. 2015-11-059-001). Written informed consent was
obtained from each study participant.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
CerS
|
ceramide synthase
|
|
TCGA-COADREAD
|
The Cancer Genome Atlas Colon and
Rectal Cancer
|
|
qPCR
|
quantitative polymerase chain
reaction
|
|
NST
|
non-neoplastic surrounding colon
tissues
|
|
LPAR1
|
lysophosphatidic acid receptor 1
|
|
NAAA
|
N-acylethanolamine acid amidase
|
|
SPHK1
|
sphingosine kinase 1
|
|
HPGD
|
15-hydroxyprostaglandin
dehydrogenase
|
|
SMPD1
|
sphingomyelin phosphodiesterase 1
|
|
SMPDL3A
|
sphingomyelin phosphodiesterase
acid-like 3A
|
|
UGGT2
|
UDP-glucose glycoprotein
glucosyltransferase 2
|
|
HA-CERS2
|
HA-tagged form of CERS2
|
|
HA-CERS6
|
HA-tagged form of CERS6
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Edwards BK, Ward E, Kohler BA, Eheman C,
Zauber AG, Anderson RN, Jemal A, Schymura MJ, Lansdorp-Vogelaar I,
Seeff LC, et al: Annual report to the nation on the status of
cancer, 1975–2006, featuring colorectal cancer trends and impact of
interventions (risk factors, screening, and treatment) to reduce
future rates. Cancer. 116:544–573. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bosetti C, Levi F, Rosato V, Bertuccio P,
Lucchini F, Negri E and La Vecchia C: Recent trends in colorectal
cancer mortality in Europe. Int J Cancer. 129:180–191. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fearon ER and Vogelstein B: A genetic
model for colorectal tumorigenesis. Cell. 61:759–767. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fearon ER: Molecular genetics of
colorectal cancer. Ann Rev Pathol. 6:479–507. 2011. View Article : Google Scholar
|
|
6
|
Cuvillier O, Pirianov G, Kleuser B, Vanek
PG, Coso OA, Gutkind S and Spiegel S: Suppression of
ceramide-mediated programmed cell death by sphingosine-1-phosphate.
Nature. 381:800–803. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Newton J, Lima S, Maceyka M and Spiegel S:
Revisiting the sphingolipid rheostat: Evolving concepts in cancer
therapy. Exp Cell Res. 333:195–200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Morad SA and Cabot MC:
Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer.
13:51–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stiban J, Tidhar R and Futerman AH:
Ceramide synthases: Roles in cell physiology and signaling. Adv Exp
Med Biol. 688:60–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fan SH, Wang YY, Lu J, Zheng YL, Wu DM,
Zhang ZF, Shan Q, Hu B, Li MQ and Cheng W: CERS2 suppresses tumor
cell invasion and is associated with decreased V-ATPase and
MMP-2/MMP-9 activities in breast cancer. J Cell Biochem.
116:502–513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen J, Li X, Ma D, Liu T, Tian P and Wu
C: Ceramide synthase-4 orchestrates the cell proliferation and
tumor growth of liver cancer in vitro and in vivo through the
nuclear factor-κB signaling pathway. Oncol Lett. 14:1477–1483.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suzuki M, Cao K, Kato S, Komizu Y,
Mizutani N, Tanaka K, Arima C, Tai MC, Yanagisawa K, Togawa N, et
al: Targeting ceramide synthase 6-dependent metastasis-prone
phenotype in lung cancer cells. J Clin Invest. 126:254–265. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Edmond V, Dufour F, Poiroux G, Shoji K,
Malleter M, Fouqué A, Tauzin S, Rimokh R, Sergent O, Penna A, et
al: Downregulation of ceramide synthase-6 during
epithelial-to-mesenchymal transition reduces plasma membrane
fluidity and cancer cell motility. Oncogene. 34:996–1005. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fitzgerald S, Sheehan KM, Espina V,
O'Grady A, Cummins R, Kenny D, Liotta L, O'Kennedy R, Kay EW and
Kijanka GS: High CerS5 expression levels associate with reduced
patient survival and transition from apoptotic to autophagy
signalling pathways in colorectal cancer. J Pathol Clin Res.
1:54–65. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Separovic D, Breen P, Joseph N, Bielawski
J, Pierce JS, VAN Buren E and Gudz TI: siRNA-mediated
down-regulation of ceramide synthase 1 leads to apoptotic
resistance in human head and neck squamous carcinoma cells after
photodynamic therapy. Anticancer Res. 32:2479–2485. 2012.PubMed/NCBI
|
|
16
|
Senkal CE, Ponnusamy S, Rossi MJ,
Bialewski J, Sinha D, Jiang JC, Jazwinski SM, Hannun YA and
Ogretmen B: Role of human longevity assurance gene 1 and
C18-ceramide in chemotherapy-induced cell death in human
head and neck squamous cell carcinomas. Mol Cancer Ther. 6:712–722.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Erez-Roman R, Pienik R and Futerman AH:
Increased ceramide synthase 2 and 6 mRNA levels in breast cancer
tissues and correlation with sphingosine kinase expression. Biochem
Biophys Res Commun. 391:219–223. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hartmann D, Lucks J, Fuchs S, Schiffmann
S, Schreiber Y, Ferreirós N, Merkens J, Marschalek R, Geisslinger G
and Grösch S: Long chain ceramides and very long chain ceramides
have opposite effects on human breast and colon cancer cell growth.
Int J Biochem Cell Biol. 44:620–628. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cancer Genome Atlas Network: Comprehensive
molecular characterization of human colon and rectal cancer.
Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sanz-Pamplona R, Berenguer A, Cordero D,
Molleví DG, Crous-Bou M, Sole X, Paré-Brunet L, Guino E, Salazar R,
Santos C, et al: Aberrant gene expression in mucosa adjacent to
tumor reveals a molecular crosstalk in colon cancer. Mol Cancer.
13:462014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ryan BM, Zanetti KA, Robles AI, Schetter
AJ, Goodman J, Hayes RB, Huang WY, Gunter MJ, Yeager M, Burdette L,
et al: Germline variation in NCF4, an innate immunity gene, is
associated with an increased risk of colorectal cancer. Int J
Cancer. 134:1399–1407. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sheffer M, Bacolod MD, Zuk O, Giardina SF,
Pincas H, Barany F, Paty PB, Gerald WL, Notterman DA and Domany E:
Association of survival and disease progression with chromosomal
instability: A genomic exploration of colorectal cancer. Proc Natl
Acad Sci USA. 106:7131–7136. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
de Sousa E, Melo F, Colak S, Buikhuisen J,
Koster J, Cameron K, de Jong JH, Tuynman JB, Prasetyanti PR,
Fessler E, van den Bergh SP, et al: Methylation of
cancer-stem-cell-associated Wnt target genes predicts poor
prognosis in colorectal cancer patients. Cell Stem Cell. 9:476–485.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edge SB and Cancer AJCo: AJCC Cancer
Staging Handbook: Form the AJCC Cancer Staging Manual. Springer;
New York: 2010
|
|
27
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ruckhäberle E, Rody A, Engels K, Gaetje R,
von Minckwitz G, Schiffmann S, Grösch S, Geisslinger G, Holtrich U,
Karn T, et al: Microarray analysis of altered sphingolipid
metabolism reveals prognostic significance of sphingosine kinase 1
in breast cancer. Breast Cancer Res Treat. 112:41–52. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mullen TD, Hannun YA and Obeid LM:
Ceramide synthases at the centre of sphingolipid metabolism and
biology. Biochem J. 441:789–802. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Park JW, Park WJ and Futerman AH: Ceramide
synthases as potential targets for therapeutic intervention in
human diseases. Biochim Biophys Acta. 1841:671–681. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mesicek J, Lee H, Feldman T, Jiang X,
Skobeleva A, Berdyshev EV, Haimovitz-Friedman A, Fuks Z and
Kolesnick R: Ceramide synthases 2, 5, and 6 confer distinct roles
in radiation-induced apoptosis in HeLa cells. Cell Signal.
22:1300–1307. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mandala SM, Thornton R, Tu Z, Kurtz MB,
Nickels J, Broach J, Menzeleev R and Spiegel S: Sphingoid base
1-phosphate phosphatase: A key regulator of sphingolipid metabolism
and stress response. Proc Natl Acad Sci USA. 95:150–155. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Panjarian S, Kozhaya L, Arayssi S, Yehia
M, Bielawski J, Bielawska A, Usta J, Hannun YA, Obeid LM and Dbaibo
GS: De novo N-palmitoylsphingosine synthesis is the major
biochemical mechanism of ceramide accumulation following p53
up-regulation. Prostaglandins Other Lipid Mediat. 86:41–48. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Karahatay S, Thomas K, Koybasi S, Senkal
CE, Elojeimy S, Liu X, Bielawski J, Day TA, Gillespie MB, Sinha D,
et al: Clinical relevance of ceramide metabolism in the
pathogenesis of human head and neck squamous cell carcinoma
(HNSCC): Attenuation of C18-ceramide in HNSCC tumors
correlates with lymphovascular invasion and nodal metastasis.
Cancer Lett. 256:101–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Senkal CE, Ponnusamy S, Bielawski J,
Hannun YA and Ogretmen B: Antiapoptotic roles of
ceramide-synthase-6-generated C16-ceramide via selective
regulation of the ATF6/CHOP arm of ER-stress-response pathways.
FASEB J. 24:296–308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pewzner-Jung Y, Brenner O, Braun S, Laviad
EL, Ben-Dor S, Feldmesser E, Horn-Saban S, Amann-Zalcenstein D,
Raanan C, Berkutzki T, et al: A critical role for ceramide synthase
2 in liver homeostasis: II. insights into molecular changes leading
to hepatopathy. J Biol Chem. 285:10911–10923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jin J, Hou Q, Mullen TD, Zeidan YH,
Bielawski J, Kraveka JM, Bielawska A, Obeid LM, Hannun YA and Hsu
YT: Ceramide generated by sphingomyelin hydrolysis and the salvage
pathway is involved in hypoxia/reoxygenation-induced Bax
redistribution to mitochondria in NT-2 cells. J Biol Chem.
283:26509–26517. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pewzner-Jung Y, Park H, Laviad EL, Silva
LC, Lahiri S, Stiban J, Erez-Roman R, Brügger B, Sachsenheimer T,
Wieland F, et al: A critical role for ceramide synthase 2 in liver
homeostasis: I. alterations in lipid metabolic pathways. J Biol
Chem. 285:10902–10910. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Spassieva SD, Ji X, Liu Y, Gable K,
Bielawski J, Dunn TM, Bieberich E and Zhao L: Ectopic expression of
ceramide synthase 2 in neurons suppresses neurodegeneration induced
by ceramide synthase 1 deficiency. Proc Natl Acad Sci USA.
113:5928–5933. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Laviad EL, Albee L, Pankova-Kholmyansky I,
Epstein S, Park H, Merrill AH Jr and Futerman AH: Characterization
of ceramide synthase 2: Tissue distribution, substrate specificity,
and inhibition by sphingosine 1-phosphate. J Biol Chem.
283:5677–5684. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bao Y, Guo Y, Zhang C, Fan F and Yang W:
Sphingosine kinase 1 and sphingosine-1-phosphate signaling in
colorectal cancer. Int J Mol Sci. 18:E21092017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Long J, Xie Y, Yin J, Lu W and Fang S:
SphK1 promotes tumor cell migration and invasion in colorectal
cancer. Tumour Biol. 37:6831–6836. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Smartt HJ, Greenhough A, Ordóñez-Morán P,
Talero E, Cherry CA, Wallam CA, Parry L, Al Kharusi M, Roberts HR,
Mariadason JM, et al: β-catenin represses expression of the tumour
suppressor 15-prostaglandin dehydrogenase in the normal intestinal
epithelium and colorectal tumour cells. Gut. 61:1306–1314. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wolf I, O'Kelly J, Rubinek T, Tong M,
Nguyen A, Lin BT, Tai HH, Karlan BY and Koeffler HP:
15-hydroxyprostaglandin dehydrogenase is a tumor suppressor of
human breast cancer. Cancer Res. 66:7818–7823. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ding Y, Tong M, Liu S, Moscow JA and Tai
HH: NAD+-linked 15-hydroxyprostaglandin dehydrogenase
(15-PGDH) behaves as a tumor suppressor in lung cancer.
Carcinogenesis. 26:65–72. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Song HJ, Myung SJ, Kim IW, Jeong JY, Park
YS, Lee SM, Nam WH, Ryu YM, Fink SP, Yang DH, et al:
15-hydroxyprostaglandin dehydrogenase is downregulated and exhibits
tumor suppressor activity in gastric cancer. Cancer Invest.
29:257–265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Myung SJ, Rerko RM, Yan M, Platzer P, Guda
K, Dotson A, Lawrence E, Dannenberg AJ, Lovgren AK, Luo G, et al:
15-Hydroxyprostaglandin dehydrogenase is an in vivo suppressor of
colon tumorigenesis. Proc Natl Acad Sci USA. 103:12098–12102. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
White-Gilbertson S, Mullen T, Senkal C, Lu
P, Ogretmen B, Obeid L and Voelkel-Johnson C: Ceramide synthase 6
modulates TRAIL sensitivity and nuclear translocation of active
caspase-3 in colon cancer cells. Oncogene. 28:1132–1141. 2009.
View Article : Google Scholar : PubMed/NCBI
|