Introduction
Hepatocellular carcinoma (HCC) is one of the leading
causes of cancer-related mortality worldwide (1). Due to its insidious onset, lack of
apparent symptoms in the early stage, and quick progression, HCC
usually is advanced when diagnosed. The efficacy of traditional
treatments, such as chemotherapy and radiotherapy, is not
satisfactory, and emerging therapeutics, including targeted therapy
and immunotherapy, have achieved limited success in the treatment
of advanced HCC thus far (2).
Cancer metastasis is the major cause of treatment failure (2), exerting a marked negative effect on
the cure and survival of patients with HCC. Numerous studies have
investigated metastasis-associated markers as potential prognostic
indicators and candidate therapeutic targets. As one of the most
well-studied microRNAs (miRNAs/miRs) in HCC, the overexpression of
miR-21 was previously reported to promote cell proliferation,
metastasis and invasion, and to be an indicator of poor prognosis
(3). Knockdown of miR-21 was also
demonstrated to significantly inhibit cancer cell migration in
vitro (4).
Exosomes are small membrane vesicles (50–150 nm
diameter) that are released by most cell types and are present in
various body fluids, including plasma, urine, saliva, breast milk
and malignant effusions. Exosomes are able to envelop proteins and
miRNAs within their double-membrane structure and transfer contents
from donor cells to recipient cells (5,6). The
release of exosomes from primary tumors into the circulatory system
has been demonstrated in various models (7), and numerous studies have reported that
exosomal miRNAs can contribute to cancer progression and metastasis
(8). Accumulating research is
investigating the use of exosome contents as biomarkers for patient
diagnosis, treatment and drug resistance.
Exosomes are also present in the supernatant of
cultured cells, including tumor cells (9). Despite the ubiquitous use of
commercial cancer cell lines, it is often questioned whether these
cell lines are able to model the biological processes of tumors
effectively, as commercial cell lines tend to lose their original
tumor characteristics due to repeated passaging (10). Additionally, it has been previously
demonstrated that patient-derived cells (PDCs) inherit the
complexity and genetic diversity of original tumors, as they are
directly derived from fresh tumor tissues (11). Therefore, PDCs were selected for use
in the present study, as they may be of a superior representative
value compared with conventional cell line models. To the best of
our knowledge, this is the first use of PDCs for investigation of
exosomal miRNAs in HCC.
miRNA expression profiling has proven useful in
diagnosing and monitoring the development and progression of
tumors. Reverse transcription-quantitative polymerase chain
reaction (PCR) and microarrays are the classical methods for miRNA
expression analysis; however, these only detect a limited number of
known miRNAs. In recent years, with the rapid development of
next-generation sequencing, miRNA sequencing (miRNA-Seq) has
offered increased specificity and sensitivity in miRNA profiling
(12); in particular, it is able to
identify novel miRNAs, which enables rapid profiling and further
investigation of miRNAs.
Given the current limited efficacy of treatments for
advanced HCC, biomarkers for liver metastasis as potential
prognostic indicators are worth extensive investigation. Although
current research is investigating the applications of exosomes as
biomarkers in the detection, diagnosis and treatment monitoring in
various cancers, little has been done to evaluate
metastasis-associated exosomal biomarkers in HCC. Therefore, the
aim of the present study was to identify differentially expressed
exosomal miRNAs in PDCs grouped by the migration rate, explore the
pathway enrichment of miRNA-targeted genes and verify the
association between the expression level of miRNAs and patient
survival using data from The Cancer Genome Atlas (TCGA). To the
best of our knowledge, the present study is the first to
investigate metastasis-related exosomal miRNA biomarkers using HCC
PDC models.
Materials and methods
PDC culture
HCC tissues were collected from the Eastern
Hepatobiliary Surgery Hospital (Shanghai, China) with informed
consent obtained from the 36 patients for PDC culture and further
research from August 2013 to June 2015. These patients with a
median age of 49 years (range, 37–74) consisted of 30 males and 6
females (Table I). PDCs were
established and cultured by 3D Medicine Inc. (Shanghai, China)
using standard procedures. In brief, fresh tissues from partial
tumor of HCC patients were washed with Dulbeccos modified Eagles
medium (DMEM)/F12 medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) to remove excess blood. The tissues were minced
into 2-mm pieces and incubated with DMEM/F12 medium supplemented
with 5% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.). When the cells reached 80% confluence, they were trypsinized
and prepared for subculture. Subsequently, the medium was changed
every 3 days.
 | Table I.Clinical information of the patients
with hepatocellular carcinoma. |
Table I.
Clinical information of the patients
with hepatocellular carcinoma.
| Clinical
information | All patients
(n=36) | Fast-migrating
group (n=10) | Low-migrating group
(n=10) | P-value |
|---|
| Sex |
|
Male | 30 | 8 | 9 | 1.000 |
|
Female | 6 | 2 | 1 |
|
| Age (years) |
|
<40 | 5 | 3 | 1 | 0.582 |
|
>40 | 31 | 7 | 9 |
|
| Hepatitis B surface
antigen |
|
Positive | 32 | 9 | 9 | 1.000 |
|
Negative | 4 | 1 | 1 |
|
| Histological
grade |
|
Well/moderately
differentiated | 0 | 0 | 0 | 1.000 |
| Poorly
differentiated | 35 | 10 | 10 |
|
|
Undifferentiated | 1 | 0 | 0 |
|
| Number of
lesions |
|
Solitary | 12 | 5 | 1 | 0.266 |
|
Multiple | 9 | 3 | 4 |
|
| NA | 15 | 2 | 5 |
|
| Microvascular
invasion |
| M0 | 5 | 1 | 2 | 0.915 |
| M1 | 8 | 1 | 3 |
|
| M2 | 8 | 1 | 4 |
|
| NA | 15 | 7 | 1 |
|
Wound healing assay
Cells in the logarithmic growth phase were seeded on
6-well plates (5×105 cells/well) and incubated for 24 h
to obtain a 90% confluent monolayer. The wound healing assay was
performed by creating scratches using 200-µl pipette tips, followed
by gentle washing three times with phosphate-buffered saline (PBS).
Cells were cultivated in serum-free medium for 24 h, and the images
of the wound gaps were captured at 0, 6 and 24 h after the wound
was made. Wound healing ability was determined by measuring the
change in the scraped area using Image-Pro Plus 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA). Each experiment was
repeated three times.
Exosome isolation
Exosomes were collected from PDCs during passages
10–15. Briefly, when the cells reached 80% confluence, the culture
medium was replaced with fresh serum-free DMEM/F12, and the cells
were cultured for an additional 48 h. Subsequently, the
supernatants were centrifuged at 1,000 × g for 5 min and 4,000 × g
for 5 min to remove cellular debris, and then collected in 50-ml
tubes for storage at −80°C. Exosomes were isolated using a 3D
Medicine exosome isolation kit (CFDA license no. Hu min xie bei
20170019). Briefly, supernatants were brought to room temperature,
passed through a 0.45-µm filter, and then a 0.22-µm filter. 3D-TC
reagent was added to the supernatants at a 1:2 ratio and mixed by
inverting the tubes several times. The mixture was incubated
overnight at 4°C, and centrifuged at 4,700 × g for 30 min at 4°C to
obtain the precipitated exosomes. The isolated exosomes were
resuspended in 200 µl PBS.
Transmission electron microscopy
Exosomes were fixed with 2% glutaraldehyde
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). One drop of the
sample was loaded onto Formvar® coated copper grids
(Agar Scientific, Ltd., Stansted, UK), and was dried at room
temperature for 10 min. The grids were washed with ultrapure water
three times and negatively stained with 2.5% uranyl acetate (pH
7.0; SPI-Chem; Structure Probe, Inc., West Chester, PA, USA),
followed by methyl cellulose/uranyl acetate (pH 4.0; Sigma-Aldrich;
Merck KGaA). Exosome particles were visualized using an H-600
electron microscope (Hitachi, Ltd., Tokyo, Japan) at 100 kV
accelerating voltage.
Western blot analysis
Cellular protein was extracted from PDCs using
radioimmunoprecipitation assay lysis buffer (Thermo Fisher
Scientific, Inc.). Exosomal protein was extracted from exosomes
using 3D Medicine protein lysis buffer. The protein lysate was then
centrifuged at 4°C for 15 min, and the supernatant was quantified
using a BCA kit (Pierce; Thermo Fisher Scientific, Inc.). Protein
(70 µg/well) was electrophoresed by SDS-PAGE on 10% gels and
transferred to a polyvinylidene difluoride membrane (EMD Millipore,
Billerica, MA, USA). The membranes were blocked using 3% bovine
serum albumin (BSA; Thermo Fisher Scientific, Inc.) at 4°C for 12 h
and then incubated with CD9 antibody (diluted at 1:1,000; cat. no.
sc-13118; Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Horseradish peroxidase-conjugated anti-mouse IgG was used as the
secondary antibody (diluted at 1:5,000; cat. no. sc-2380; Santa
Cruz Biotechnology, Inc.). Bands were visualized using enhanced
chemiluminescence (Thermo Fisher Scientific, Inc.). Comparison of
the grey signal intensity bands on the western blots were performed
using software Adobe Photoshop CS6. Each experiment was repeated
three times.
Exosomal and cellular RNA
isolation
RNA from exosomes was isolated using miRNeasy
Serum/Plasma kit (Qiagen, Inc., Valencia, CA, USA) according to the
manufacturers protocol. Briefly, 700 µl QIAzol was added to 200 µl
exosomes, vortexed and incubated. Then 90 µl chloroform was added
to the mixture for further vortexing and incubation, and the
mixture was centrifuged at 4°C for 15 min. The upper aqueous phase
was transferred to a new tube, and two volumes of 100% ethanol were
added. Following vortexing, the solution was loaded to a spin
column and centrifuged at 8,000 × g for 15 sec. The flow-through
was discarded, and the spin column was rinsed twice with wash
buffer, air-dried and the recovered exosomal RNA was eluted using
15 µl RNase-free water. Total RNA from PDCs was isolated using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturers protocol. RNA quality and concentration were
analyzed using the NanoDrop ND-100 Spectrophotometer (Thermo Fisher
Scientific, Inc., Wilmington, DE, USA).
Small RNA library construction and
sequencing
Small RNA library preparation for high throughput
sequencing was performed using the protocols and reagents from the
NEBNext Small RNA Library Prep Set for Illumina (New England
BioLabs, Inc., Ipswich, MA, USA). The small RNA library prepared
from PDCs was fractionated by DNA length on a 2% TAE agarose gel
for the excision of the 150-bp band. The small RNA library was
purified using NucleoSpin Gel and PCR Clean-Up kit (Qiagen,
Shanghai, China). The purified library was analyzed for purity
using an Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.,
Santa Clara, CA, USA) and then subjected to single-ended
strand-specific sequencing on the Illumina HiSeq X10 platform
(Illumina, Inc., San Diego, CA, USA).
Sequence mapping and small RNA
identification
Following trimming of adaptor sequences, reads were
mapped to hg19 using the Burrows-Wheeler Aligner 0.7.12-r1039 and a
number of reads mapped to miRNAs from miRBase v21 were calculated.
The expression levels were normalized using DESeq2 version 1.20.0.
The principal component analysis (PCA) and the contribution of each
gene to the dominant direction were performed as described by Huang
et al (13). miRNAs that met
all the following conditions were considered differentially
expressed: i) P<0.05 between two groups; ii) the absolute value
of contribution was >0.03; and iii) the average expression level
of groups with higher mean expression was >100 reads per
kilobase per million mapped reads (RPKM).
Statistical analysis
The data were presented as the mean ± SD of three
independent experiments. The different migration rates between the
fast and slow groups were assessed by Students t-test. For
differentially expressed miRNAs, the Students t-test was performed
to calculate the P-value. Pearsons correlation analysis of miRNA
expression between PDCs and their exosomes was performed. The
target genes of differentially expressed miRNAs were analyzed using
miRTarBase (http://mirtarbase.mbc.nctu.edu.tw) instead of
frequently used computational tools, such as TargetScan, miRanda,
PicTar and DIANA-microT. The overall survival (OS) in patients with
HCC was analyzed using the Kaplan-Meier method and log-rank test.
Fishers test and Chi-square test were used to analyze the clinical
characteristic differences between the fast and slow groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Characterization of PDCs
HCC tissues obtained from surgical resection were
cultured to establish PDCs. All cell lines were free of
contamination by bacteria or mycoplasma. Short tandem repeat (STR)
analysis revealed that all PDCs were derived from the corresponding
tissue samples and the STR loci profiles were all unique when
compared with each other (data not shown). The growth rate was
estimated by the passage time, and the PDCs with a passage time of
>20 days were excluded. Considering the potential similarity of
PDCs derived from the same patient, all selected PDCs were derived
from different patients. In total, 36 PDC cultures were selected
for further use in migration assays.
The clinical information of the patients is
presented in Table I. The 21 PDCs
with a passage time of ≤7 days were classified as having a fast
growth rate, and the 15 PDCs with a passage time of >7 days were
classified as having a slow growth rate. Hepatitis B virus (HBV)
DNA integration in PDCs was examined by polymerase chain reaction
(PCR) amplification of the virus gene, and 20 PDCs tested positive,
while 5 tested negative for HBV. Serological analysis for HBV
antigen and antibody of the derived patient was also performed. HBV
surface antigen was detected in 32 patients, indicating a chronic
HBV infection in 89% of patients. Edmondson-Steiner grading
indicated poor differentiation in the majority of the patients.
Quantity and distribution of microvascular invasion (MVI) was
evaluated and classified as M0-MVI for no visible MVI, M1-MVI for
low-risk MVI, and M2-MVI for high-risk MVI. Other clinical
information, including TNM stage and prognosis, was
unavailable.
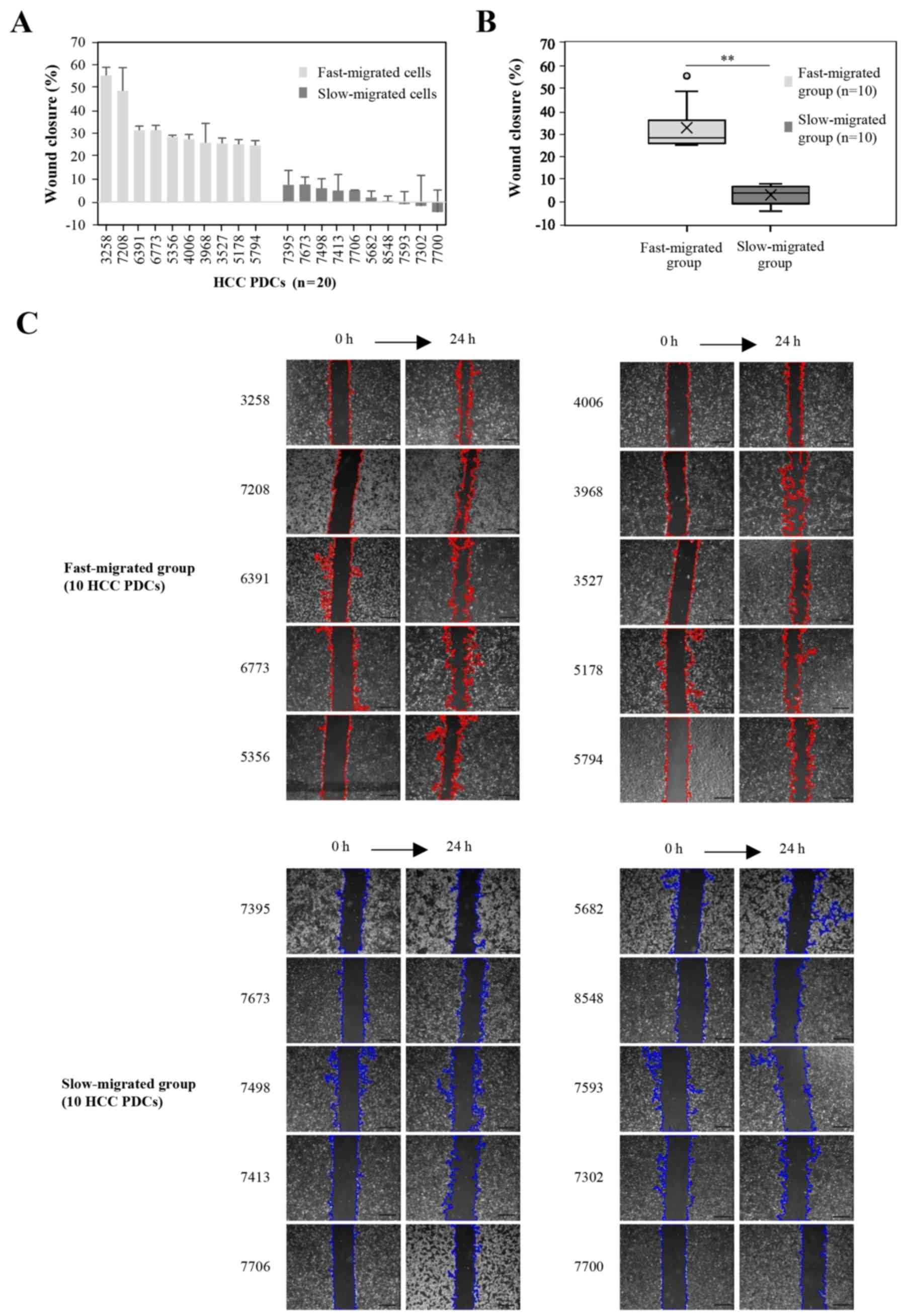
Group enrollment of PDCs by migration
rate
To identify miRNAs that may play a role in the
metastasis of liver tumors, cell migration rates were determined
using an in vitro wound healing assay. The migration rate
was quantified as the wound closure percentage at 24 h after the
scratch wound was made. Among the 36 PDCs, the top 10 and bottom 10
PDCs according to the migration rate were designated as the fast-
and slow-migrated groups, respectively. No association was observed
between PDC migration rate and patient clinical characteristics,
such as the growth rate and HBV infection status in the patients.
The wound closure percentages were all ≥24.8% in the fast-migrated
group, and ≤7.5% in the slow-migrated group. Statistically, the
mean and median values were 32.5 and 28.0% in the fast-migrated
group, and 2.7 and 3.6% in the slow-migrated group (Fig. 1), respectively, with a significant
difference in wound closure between the groups
(P=1.8×10−7). In order to exclude the influence of the
proliferation differences of HCC PDCs on the cellular migration,
the 20 HCC PDCs in the fast-migrated group and the slow-migrated
group were resuscitated and detected in serum-free medium. When
cells were cultivated in serum-free medium for 24 h, the results
revealed that the proliferation rates in the fast-migrated group
and the slow-migrated group were not significant (data not
shown).
Identification of PDC-derived
exosomes
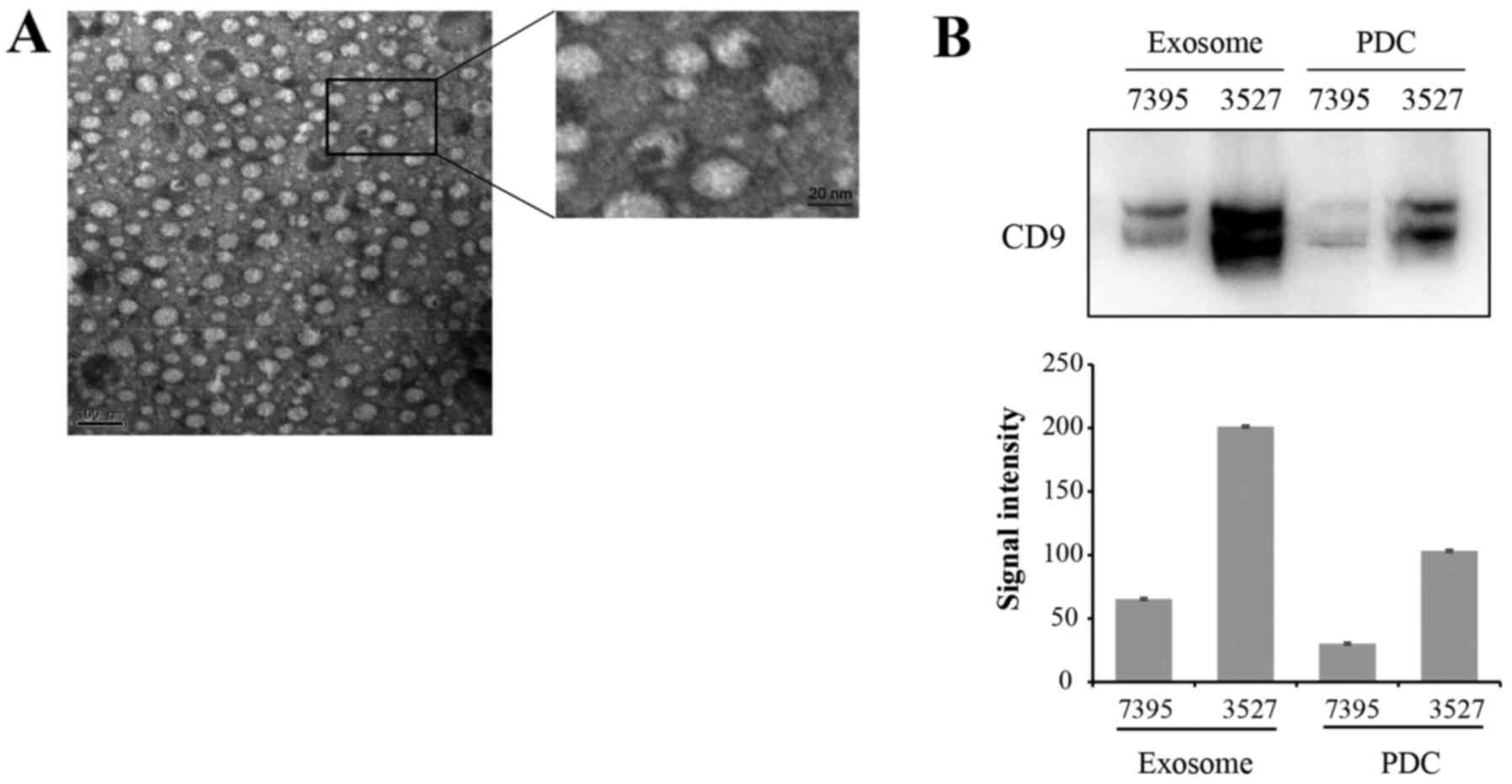
Exosomes were isolated from the supernatant of PDCs.
Fig. 2A presents PDC-derived
exosomes visualized by electron microscopy. Exosomes appeared as
typical spherical vesicles ranging from 50–150 nm in diameter.
Protein expression of known exosome marker CD9 was confirmed by
western blot analysis in PDC-derived exosomes and in the PDCs
(Fig. 2B upper image). The gray
signal intensities of CD9 were presented in Fig. 2B (lower image). The CD9 blot
produced two protein bands, with the upper band presumably due to
glycosylation modification.
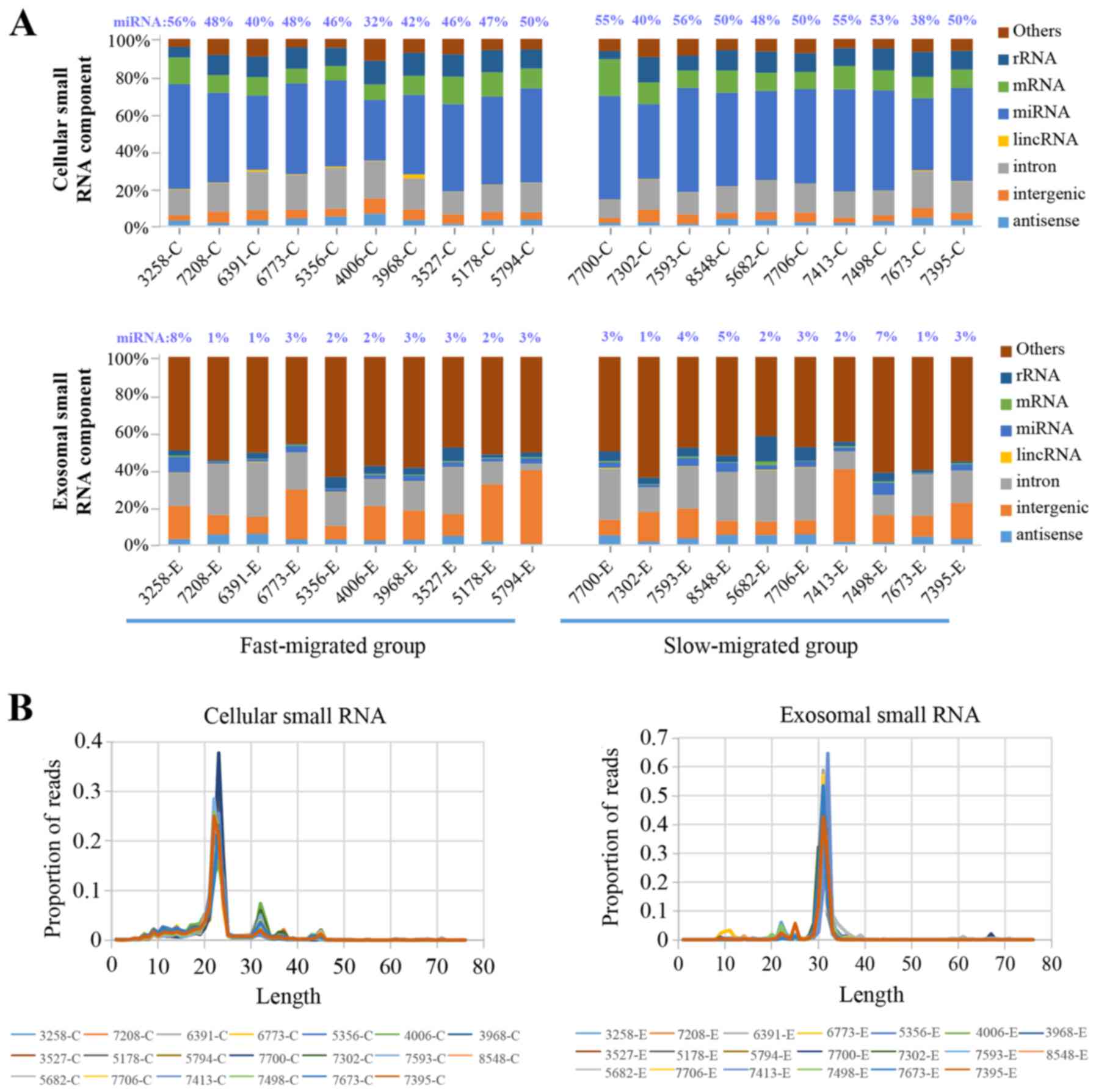
Composition of cellular and exosomal
small RNAs
Deep sequencing of small RNA generated an average of
13 million and 9 million total reads for PDC and PDC-derived
exosome samples, respectively. Of the total reads, >97% were
successfully mapped to the reference genome and the sequencing data
was adequate for further analysis.
In PDC samples, miRNA was the most abundant class of
small RNAs, representing 47.5% of the entire cellular small RNA
component. The percentage of miRNAs among all small RNAs varied
from 32.3 to 55.9% from sample to sample, with no significant
difference between the fast- and slow-migrated groups (Fig. 3A). By contrast, in exosome samples
miRNAs accounted for only 3.0% of all small RNA molecules.
Similarly, no significant difference in the exosome miRNA
proportion was observed between the two groups. The most abundant
class of small RNAs present in the exosomes was unannotated small
RNA (Fig. 3A).
Considering the marked difference in the composition
of the small RNA population between cellular and exosomal samples,
the read length distribution of the samples was investigated. The
PDC samples produced a predominant peak at 23 nucleotides and an
additional peak at 32 nucleotides, while exosomes had a major peak
at 32 nucleotides, and tiny peaks from 22–25 nucleotides (Fig. 3B). miRNAs are 21–25 nucleotides,
which corresponded to the major peak in cellular small RNAs and the
minor peaks in the exosomal components. The distribution of read
length corresponded to the percentile of miRNAs present in cellular
and exosomal RNA samples.
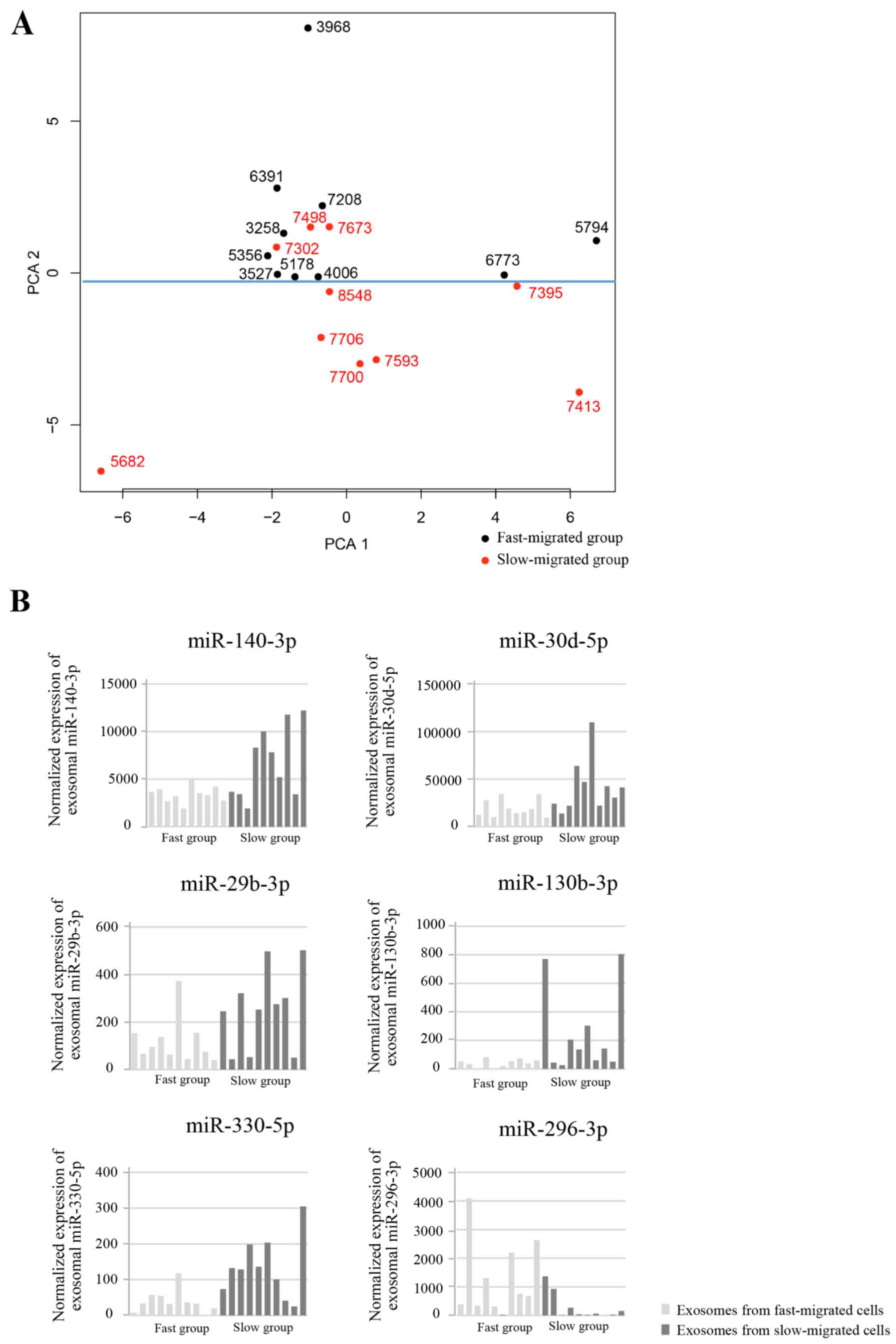
Differentially expressed exosomal
miRNAs are associated with migration rate
Next it was also investigated whether specific
miRNAs were upregulated or downregulated in the fast-migrated group
compared with the slow-migrated group. PCA was performed and the
contribution of each gene to the dominant direction was calculated.
The unsupervised PCA clustered the samples into two groups
(separated by the blue line in Fig.
4A), which generally accorded with the migration rate of each
sample. To focus on the miRNAs with relatively high expression,
miRNAs with mean expressions of <100 RPKM were removed. Each
miRNA with a Students t-test result of P<0.05 between groups,
and an absolute value of contribution of >0.03 was considered to
be differentially expressed. Six exosomal miRNAs were
differentially expressed between the fast- and slow-migrated
groups. In the fast-migrated group, five miRNAs (miR-140-3p,
miR-30d-5p, miR-29b-3p, miR-130b-3p and miR-330-5p) were
downregulated, and one miRNA (miR-296-3p) was upregulated (Fig. 4B). The detailed parameters for the
six miRNAs are listed in Table
II.
| Table II.Screened parameters for the six
differentially expressed exosomal microRNAs. |
Table II.
Screened parameters for the six
differentially expressed exosomal microRNAs.
| miRNA | PC2 value | Average fast | Average slow | P-value
(t-test) |
|---|
| hsa-miR-140-3p | −0.058 | 3413 | 6781 | 0.02 |
| hsa-miR-30d-5p | −0.042 | 19531 | 41768 | 0.037 |
| hsa-miR-29b-3p | −0.037 | 121 | 254 | 0.047 |
|
hsa-miR-130b-3p | −0.033 | 41 | 254 | 0.048 |
| hsa-miR-330-5p | −0.032 | 39 | 135 | 0.006 |
| hsa-miR-296-3p | 0.044 | 1276 | 287 | 0.046 |
Pathway analysis
To reveal the potential roles of the differentially
expressed miRNAs in HCC, their target genes were analyzed using
miRTarBase (http://mirtarbase.mbc.nctu.edu.tw). The prediction
principle for computational tools predominantly depends on the
complementarity between miRNA seed region and the 3-untranslated
region of their target genes. The majority of miRNAs imperfectly
bind to the complementary sites, thus various computational methods
have been generated for miRNA target prediction, and the resulting
lists of candidate target genes from different algorithms often do
not overlap. However, miRTarBase only includes experimentally
validated miRNA-target interactions, which provides more reliable
data for subsequent pathway analysis. To further reveal the
potential roles of these miRNAs, the miRNAs and their target genes
were analyzed using the miRTarBase database. In summary, 1652
non-repetitive genes were potential targets of the six
differentially expressed miRNAs. Pathway enrichment analysis was
performed using DAVID 6.8 (https://david.ncifcrf.gov/) for Kyoto Encyclopedia of
Genes and Genomes (KEGG) and Reactome analysis.
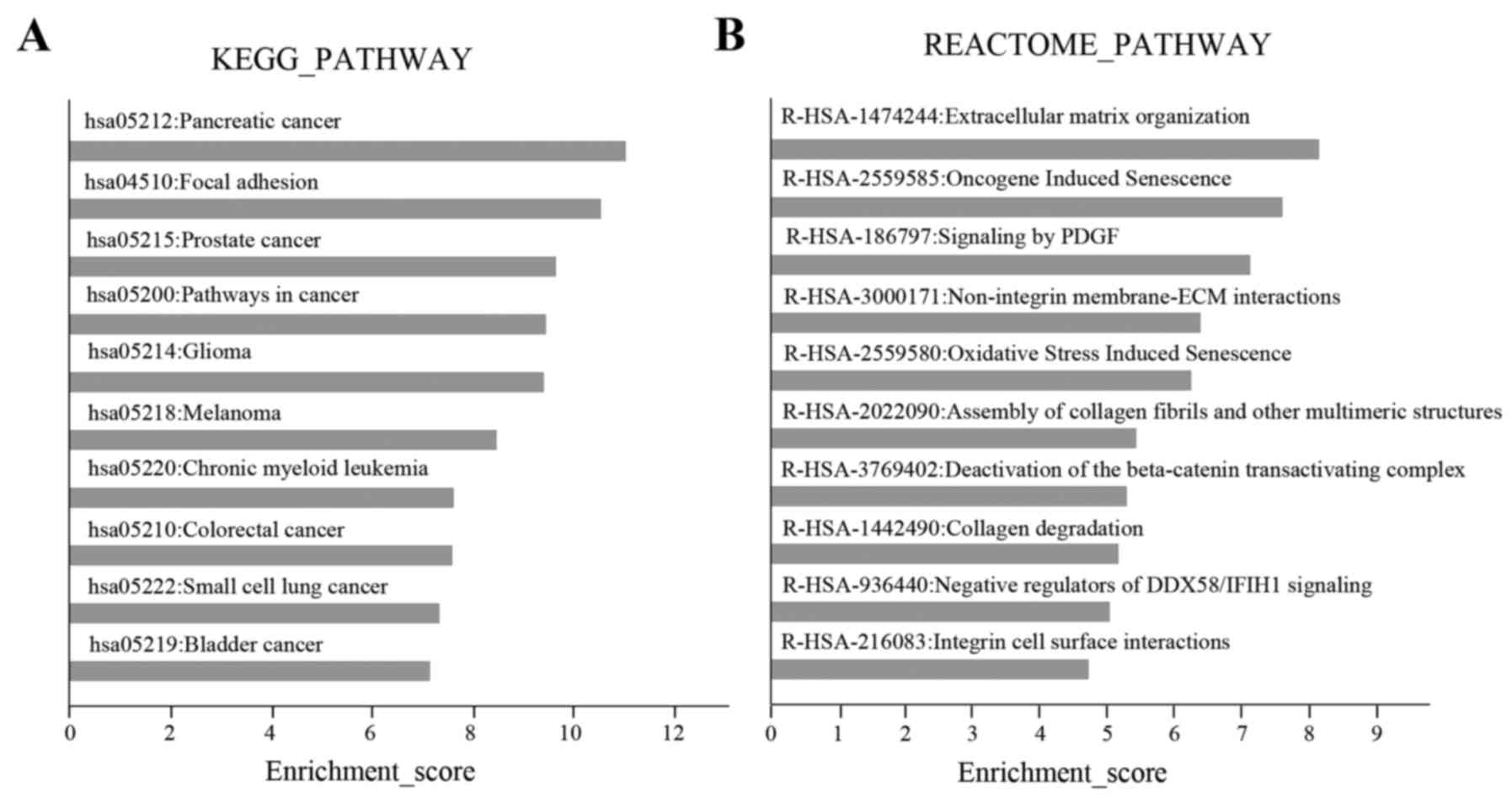
In KEGG pathway analysis, the top 10 pathways were
predominantly cancer-associated pathways, including ‘pancreatic
cancer’, ‘prostate cancer’ and ‘pathways in cancer’. The second
highest scoring pathway was ‘focal adhesion’ (Fig. 5A), which is involved in the
regulation of cell migration (14).
In the Reactome analysis, the highest scoring pathway was
‘extracellular matrix (ECM) organization’, which was consistent
with the ‘focal adhesion’ pathway identified by KEGG analysis.
Other Reactome pathways, including ‘signaling by PDGF’ and
‘non-integrin membrane-ECM interactions’ were also associated with
cell migration (Fig. 5B). The genes
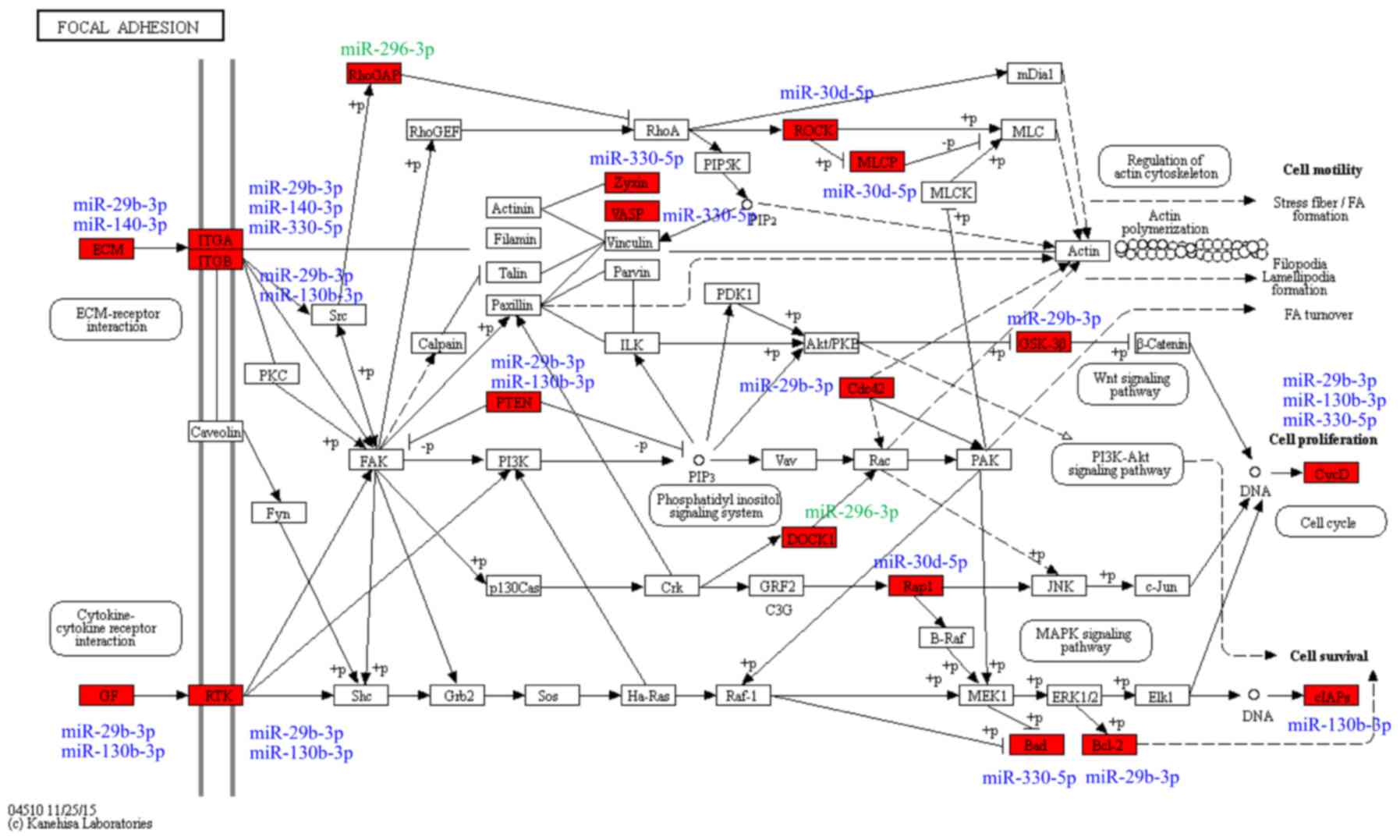
associated with the ‘focal adhesion’ pathway that were predicted as
targets of the six exosomal miRNAs were comprised of upstream
cytokine-receptor interaction molecules, including ECM components
(COL1A1, COL5A2, LAMA and LAMC1), growth factors
(PDGFC, VEGFA and IGF1), receptor tyrosine kinases
(ERBB2 and PDGFRA) and integrins (ITGA5 and
ITGB1), and downstream factors (PTEN and
PIK3CA), cell proliferation genes (CCND1 and
CCND2), cell survival genes (BCL2 and BAD) The
targets of the six differentially expressed miRNAs in the ‘focal
adhesion’ pathway and their associated miRNAs are presented in
Fig. 6.
Validation of migration-associated
miRNAs using TCGA data
The expression of individual miRNAs was compared
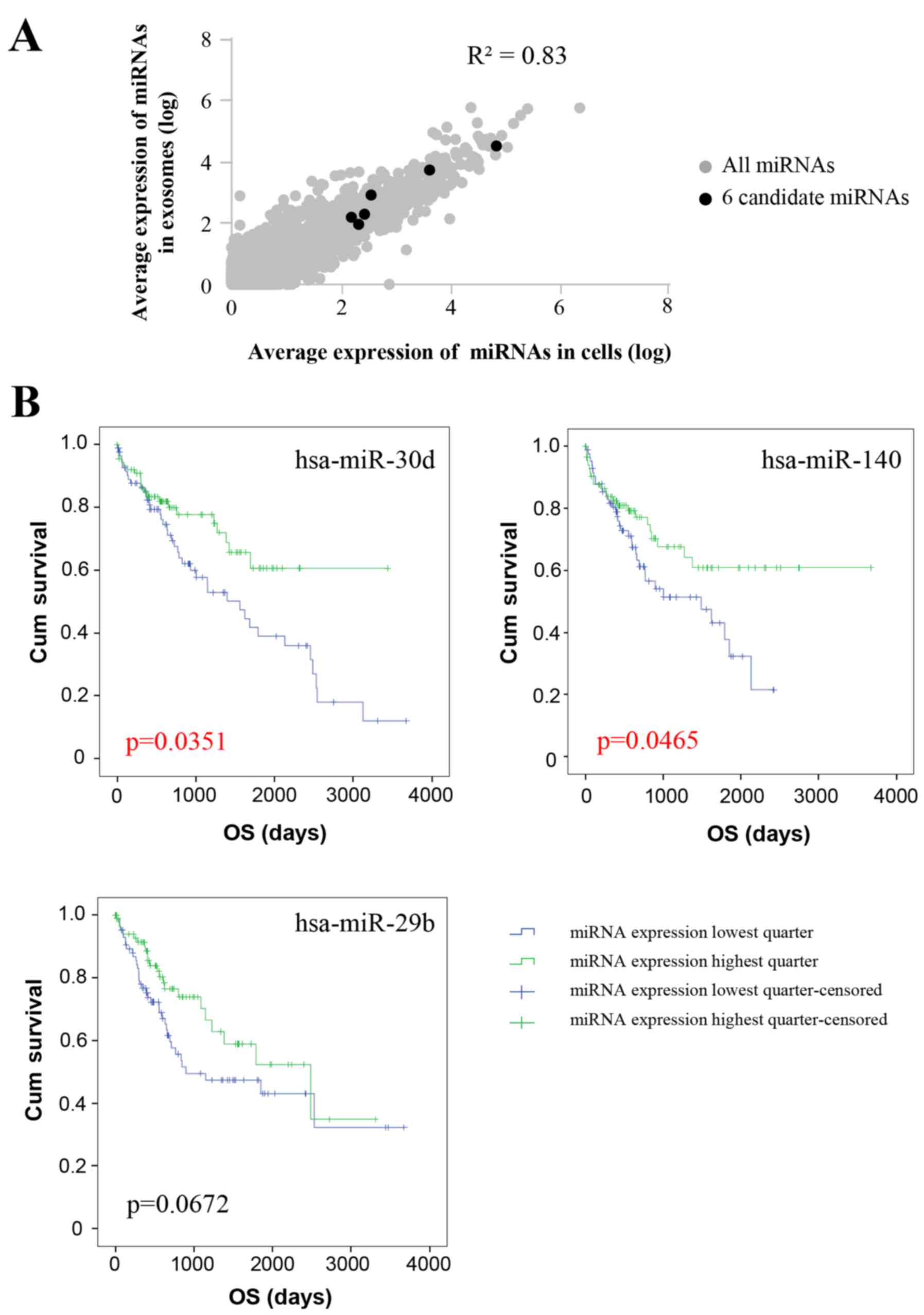
between exosomes and their corresponding PDCs. Generally, the
association analysis highlighted a strong correlation
(R2=0.83) between normalized miRNA expression in PDCs
and exosomes, which indicated that exosomes reflect the miRNA
composition of the derived cells. As for the six differentially
expressed miRNAs, normalized exosomal miRNA levels were also
correlated with the miRNA levels of PDCs as well (Fig. 7A). Considering the average miRNA
expression levels were evaluated above, the expression levels of
the 6 miRNAs in individual PDC samples and the paired exosome
samples were further analyzed, and a similar trend of high
correlation of miRNA expression between PDCs and their
corresponding exosomes was observed (data not shown).
In line with the finding that exosome miRNAs may
reflect the miRNA composition of the PDCs that the exosomes were
derived from, tumor tissue-derived miRNA levels and the clinical
information of liver tumors were obtained from TCGA for validation
analysis. Among the 372 available records, only 4 patients were
categorized as M1 for definitive distal metastasis; therefore, it
was impossible to study the association between miRNA levels and
metastasis directly. Considering distal metastasis was positively
correlated with poor clinical prognosis, the association between OS
and miRNA levels were analyzed. The patients were divided by the
expression level of a certain differentially expressed miRNA, with
the top quartile and bottom quartile patients set as the low and
high miRNA expression groups, respectively. The miRNA expression
level and corresponding OS was analyzed using the Kaplan-Meier
method (Fig. 7B). Patients with
high expression of the three differentially expressed miRNAs,
miR-30d, miR-140 and miR-29b, exhibited improved OS (P=0.0351,
P=0.0465 and P=0.0672, respectively). Analysis of the clinical
characteristics, including tumor stage, pathological T, N and M
stage, detected no differences between the low and high miRNA
expression groups (Tables
III–V), suggesting that the
result is not a secondary effect caused by the differences in the
clinical characteristics between the two groups. Moreover, we
analyzed the difference between the cirrhosis of these patients and
survival from TCGA database. The following result revealed that
there was no difference between cirrhosis and cum survival
(P=0.773), indicating that our result concerning miRNAs and
survival is not an epiphenomenon from their differences in
underlying cirrhosis (data not shown).
| Table III.Clinical characteristics of HCC
patients in the hsa-mir-140 lowest and highest quarters. |
Table III.
Clinical characteristics of HCC
patients in the hsa-mir-140 lowest and highest quarters.
|
|
| hsa-mir-140 |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | No. of
patients | Lowest quarter | Highest
quarter | P-value |
|---|
| Stage |
| I | 73 | 32 | 41 | 0.240 |
| II | 47 | 23 | 24 |
|
|
III | 48 | 29 | 19 |
|
| IV | 1 | 0 | 1 |
|
| NA | 17 | 9 | 8 |
|
| Pathologic_T |
| T1 | 75 | 33 | 42 | 0.215 |
| T2 | 49 | 24 | 25 |
|
| T3 | 41 | 25 | 16 |
|
| NA | 13 | 7 | 6 |
|
| Pathologic_N |
| N0 | 125 | 62 | 63 | 0.363 |
| N1 | 2 | 2 | 0 |
|
| NX | 47 | 23 | 24 |
|
| NA | 12 | 6 | 6 |
|
| Pathologic_M |
| M0 | 129 | 64 | 65 | 0.577 |
| M1 | 1 | 0 | 1 |
|
| MX | 44 | 23 | 21 |
|
| NA | 12 | 6 | 6 |
|
| Table V.Clinical characteristics of HCC
patients in the hsa-mir-29b lowest and highest quarters. |
Table V.
Clinical characteristics of HCC
patients in the hsa-mir-29b lowest and highest quarters.
|
|
| hsa-mir-29b |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | No. of
patients | Lowest quarter | Highest
quarter | P-value |
|---|
| Stage |
| I | 80 | 36 | 44 | 0.536 |
| II | 42 | 20 | 22 |
|
|
III | 42 | 23 | 19 |
|
| IV | 4 | 3 | 1 |
|
| NA | 18 | 11 | 7 |
|
| Pathologic_T |
| T1 | 83 | 37 | 46 | 0.433 |
| T2 | 46 | 23 | 23 |
|
| T3 | 42 | 24 | 18 |
|
| TX | 1 | 0 | 1 |
|
| NA | 9 | 5 | 4 |
|
| Pathologic_N |
| N0 | 121 | 63 | 58 | 0.546 |
| N1 | 3 | 1 | 2 |
|
| NX | 52 | 23 | 29 |
|
| NA | 10 | 6 | 4 |
|
| Pathologic_M |
| M0 | 127 | 63 | 64 | 0.203 |
| M1 | 3 | 3 | 0 |
|
| MX | 47 | 22 | 25 |
|
| NA | 9 | 5 | 4 |
|
Discussion
In PDC samples, miRNA was the most abundant class of
small RNAs, accounting for 47.5% of the entire cellular small RNA
population, which is comparable to the proportion in cells and
plasma/serum reported in previous studies (15). However, in PDC-derived exosome
samples, miRNAs accounted for only 3.0% of the small RNA
repertoire, and the length distribution pattern differed from that
in PDCs. Notably, similar phenomena were also observed in other
studies (15). It was reported
miRNAs represented 30 and 2–5% of the total small RNAome in
mesenchymal stem cells and cell-derived exosomes, respectively
(16). In exosomes collected from
cell culture medium, miRNAs accounted for 2–7% of all small RNAs
obtained by different isolation methods (17).
To identify potential miRNAs that may be involved in
the metastasis of liver tumors, a wound healing assay was used to
obtain two groups of PDCs with fast or slow cell migration rates.
Six exosomal miRNAs were differentially expressed between the fast
and slow-migrated groups. Five miRNAs (miR-140-3p, miR-30d-5p,
miR-29b-3p, miR-130b-3p and miR-330-5p) were downregulated, and one
miRNA (miR-296-3p) was upregulated in the fast-migrated group
compared with the slow-migrated group. Although few studies have
explored the association between these exosomal miRNAs and tumor
metastasis directly, accumulating evidence indicates that these
miRNAs may have vital roles in tumor metastasis. For example,
exosomal miR-140 was identified as a negative regulator of cell
migration in breast cancer via targeting of SRY-box 9 (18). This indicated that miR-140 acts as a
potential tumor-suppressor by targeting oncogenes, and this may
explain why it was downregulated in the fast-migrated group.
Similarly, miR-30d-5p was reported to inhibit tumor cell
proliferation and migration by directly targeting cyclin E2 in
non-small cell lung cancer (19).
Tumor-suppressive miRNA-29s directly regulated lysyl oxidase like 2
expression and inhibited cancer cell migration and invasion in
renal cell carcinoma (20).
miR-130b-3p was reported to inhibit cell invasion and migration by
targeting the Notch ligand gene delta-like 1 in breast carcinoma
(21). miR-330-5p regulated
tyrosinase and protein disulfide isomerase family A member 3
expression, and suppressed cell proliferation, migration and
invasion in cutaneous malignant melanoma (22). As an oncogenic miRNA, the level of
miR-296-3p was markedly higher in highly metastatic human prostate
cancer cells than in non-metastatic cells (23). However, the roles of miRNAs in tumor
metastasis may vary among different studies, in different tumor
types, under different physiological and pathological conditions,
and through different mechanisms. miR-30d was also demonstrated to
promote the metastatic behavior of melanoma cells by directly
suppressing the GalNac transferase polypeptide
N-acetylgalactosaminyltransferase 7 (24), while miR-296 was reported to inhibit
the metastasis and epithelial-mesenchymal transition of colorectal
cancer by targeting S100 calcium binding protein A4 (25).
In HCC, specific target genes could not be
identified for these six differentially expressed miRNAs. However,
the potential targeted genes were enriched in the ‘focal adhesion’
pathway. Focal adhesions are formed of integrin and other adapter
proteins, and the dynamic assembly and disassembly of focal
adhesion-ECM has a central role in cell dissemination and migration
(14), resulting in the ability of
cancer cells to metastasize. Focal adhesion kinase was revealed to
be associated with the development and progression of HCC (26). Accumulating evidence suggests that
miRNAs may regulate tumor migration and metastasis by affecting
signal-mediated cytoskeletal and cell matrix adhesion remodeling
(23), and the analysis in the
present study demonstrated the involvement of ‘focal adhesion’
associated pathways in tumor cell migration and metastasis.
In summary, PDCs, which may be of superior
representative value compared with commercial cancer cell line
models, were used to investigate exosome miRNAs as biomarkers in
the present study. miRNA-Seq was applied for comprehensive
screening of differentially expressed exosomal miRNAs, rather than
using RT-q PCR or microarray methods. Consequently, six
differentially expressed exosomal miRNAs were identified as
potential biomarkers for metastasis in HCC, and miRNA-targeted
genes were enriched in the ‘focal adhesion’ pathway, supporting the
role of miRNA regulation in cell migration and tumor metastasis.
Finally, the six miRNA expression levels and corresponding patient
survival profiles from TCGA were explored, and the association
between the expression levels of miRNAs and patient survival was
validated, indicating the potential role of these miRNAs in
prognosis. To the best of our knowledge, this is the first study to
investigate migration-associated exosomal miRNA biomarkers in HCC
PDCs.
Acknowledgements
Not applicable.
Funding
This present study was supported by the National Key
Basic Research Program of China (grant no. 2014CB542102) and
Science Fund for Creative Research Groups, and the National Natural
Science Foundation of China (grant no. 81521091).
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors contributions
LXY designed the migration rate study, performed the
wound healing assay, contributed to the data interpretation and
wrote the manuscript; BLZ performed the pathway analysis and wrote
the manuscript; YY conceived the study, designed the electron
microscopy identification of the exosomes, interpreted the data and
wrote the manuscript; MCW analyzed the miRNA data from TCGA
database, contributed to the data interpretation and wrote the
manuscript; GLL isolated the exosomal and cellular RNA and
contributed to the pathway analysis; YG performed the western blot
identification of the exosomes and prepared the exosomal samples
for transmission electron microscopy; HL collected clinical
information of the PDCs and contributed to the data interpretation;
CHX conducted the electron microscopy identification of the
exosomes and wrote the manuscript; JJX constructed small RNA
library, contributed to the quality control of small RNA library
and performed miRNA-seq; HQ performed the bioinformatics analysis
of the miRNA-seq data, contributed to the data interpretation and
wrote the manuscript; XYX isolated the PDC-derived exosomes and
performed the migration rate study; ZSC cultured and established
the PDCs, performed the wound healing assay and wrote the
manuscript; DDZ contributed to the revised manuscript and data
interpretation; FGL discussed the hypothesis and contributed to the
data interpretation; SGZ conceived the study and led the project;
RL conceived the study and participated in its design and
coordination. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
HCC tissues were collected with ethics approval
obtained from the Eastern Hepatobiliary Surgery Hospital, and
informed consent was obtained from the patients for PDC culture and
further research.
Patient consent for publication
Not applicable.
Competing interests
All authors affiliated to 3D Medicine Inc., are
current or former employees. No potential conflicts of interest
were disclosed by the other authors.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
PDCs
|
patient-derived cells
|
|
miRNA-seq
|
microRNA sequencing
|
|
miRNA
|
microRNA
|
|
RPKM
|
reads per kilobase per million mapped
reads
|
|
OS
|
overall survival
|
|
STR
|
short tandem repeat
|
|
MVI
|
microvascular invasion
|
|
PCA
|
principal component analysis
|
|
ECM
|
extracellular matrix
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steeg PS: Targeting metastasis. Nat Rev
Cancer. 16:201–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu G, Zhang Y, Wei J, Jia W, Ge Z, Zhang Z
and Liu X: MicroRNA-21 promotes hepatocellular carcinoma HepG2 cell
proliferation through repression of mitogen-activated protein
kinase-kinase 3. BMC Cancer. 13:4692013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan LX, Wu QN, Zhang Y, Li YY, Liao DZ,
Hou JH, Fu J, Zeng MS, Yun JP, Wu QL, et al: Knockdown of miR-21 in
human breast cancer cell lines inhibits proliferation, in vitro
migration and in vivo tumor growth. Breast Cancer Res. 13:R22011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li J, Liu K, Liu Y, Xu Y, Zhang F, Yang H,
Liu J, Pan T, Chen J, Wu M, et al: Exosomes mediate the
cell-to-cell transmission of IFN-α-induced antiviral activity. Nat
Immunol. 14:793–803. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lässer C, Alikhani VS, Ekström K, Eldh M,
Paredes PT, Bossios A, Sjöstrand M, Gabrielsson S, Lötvall J and
Valadi H: Human saliva, plasma and breast milk exosomes contain
RNA: Uptake by macrophages. J Transl Med. 9:92011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suetsugu A, Honma K, Saji S, Moriwaki H,
Ochiya T and Hoffman RM: Imaging exosome transfer from breast
cancer cells to stroma at metastatic sites in orthotopic nude-mouse
models. Adv Drug Deliv Rev. 65:383–390. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Peinado H, Alečković M, Lavotshkin S,
Matei I, Costa-Silva B, Moreno-Bueno G, Hergueta-Redondo M,
Williams C, García-Santos G, Ghajar C, et al: Melanoma exosomes
educate bone marrow progenitor cells toward a pro-metastatic
phenotype through MET. Nat Med. 18:883–891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kogure T and Patel T: Isolation of
extracellular nanovesicle microRNA from liver cancer cells in
culture. Methods Mol Biol. 1024:11–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
van Staveren WC, Solís DY, Hébrant A,
Detours V, Dumont JE and Maenhaut C: Human cancer cell lines:
Experimental models for cancer cells in situ? For cancer stem
cells? Biochim Biophys Acta. 1795:92–103. 2009.PubMed/NCBI
|
|
11
|
Byrne AT, Alférez DG, Amant F, Annibali D,
Arribas J, Biankin AV, Bruna A, Budinská E, Caldas C, Chang DK, et
al: Interrogating open issues in cancer precision medicine with
patient-derived xenografts. Nat Rev Cancer. 17:254–268. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Buitrago DH, Patnaik SK, Kadota K,
Kannisto E, Jones DR and Adusumilli PS: Small RNA sequencing for
profiling microRNAs in long-term preserved formalin-fixed and
paraffin-embedded non-small cell lung cancer tumor specimens. PLoS
One. 10:e01215212015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang J, Qin H, Yang Y, Chen X, Zhang J,
Laird S, Wang CC, Chan TF and Li TC: A comparison of transcriptomic
profiles in endometrium during window of implantation between women
with unexplained recurrent implantation failure and recurrent
miscarriage. Reproduction. 153:749–758. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van Roosmalen W, Le Dévédec SE, Golani O,
Smid M, Pulyakhina I, Timmermans AM, Look MP, Zi D, Pont C, de
Graauw M, et al: Tumor cell migration screen identifies SRPK1 as
breast cancer metastasis determinant. J Clin Invest. 125:1648–1664.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rodríguez M, Bajo-Santos C, Hessvik NP,
Lorenz S, Fromm B, Berge V, Sandvig K, Linē A and Llorente A:
Identification of non-invasive miRNAs biomarkers for prostate
cancer by deep sequencing analysis of urinary exosomes. Mol Cancer.
16:1562017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baglio SR, Rooijers K, Koppers-Lalic D,
Verweij FJ, Pérez Lanzón M, Zini N, Naaijkens B, Perut F, Niessen
HW, Baldini N, et al: Human bone marrow- and adipose-mesenchymal
stem cells secrete exosomes enriched in distinctive miRNA and tRNA
species. Stem Cell Res Ther. 6:1272015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang YT, Huang YY, Zheng L, Qin SH, Xu XP,
An TX, Xu Y, Wu YS, Hu XM, Ping BH, et al: Comparison of isolation
methods of exosomes and exosomal RNA from cell culture medium and
serum. Int J Mol Med. 40:834–844. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gernapudi R, Yao Y, Zhang Y, Wolfson B,
Roy S, Duru N, Eades G, Yang P and Zhou Q: Targeting exosomes from
preadipocytes inhibits preadipocyte to cancer stem cell signaling
in early-stage breast cancer. Breast Cancer Res Treat. 150:685–695.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen D, Guo W, Qiu Z, Wang Q, Li Y, Liang
L, Liu L, Huang S, Zhao Y and He X: MicroRNA-30d-5p inhibits tumour
cell proliferation and motility by directly targeting CCNE2 in
non-small cell lung cancer. Cancer Lett. 362:208–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nishikawa R, Chiyomaru T, Enokida H,
Inoguchi S, Ishihara T, Matsushita R, Goto Y, Fukumoto I, Nakagawa
M and Seki N: Tumour-suppressive microRNA-29s directly regulate
LOXL2 expression and inhibit cancer cell migration and invasion in
renal cell carcinoma. FEBS Lett. 589:2136–2145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shui Y, Yu X, Duan R, Bao Q, Wu J, Yuan H
and Ma C: miR-130b-3p inhibits cell invasion and migration by
targeting the Notch ligand Delta-like 1 in breast carcinoma. Gene.
609:80–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su BB, Zhou SW, Gan CB and Zhang XN:
MiR-330-5p regulates tyrosinase and PDIA3 expression and suppresses
cell proliferation and invasion in cutaneous malignant melanoma. J
Surg Res. 203:434–440. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou L, Liu F, Wang X and Ouyang G: The
roles of microRNAs in the regulation of tumor metastasis. Cell
Biosci. 5:322015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gaziel-Sovran A, Segura MF, Di Micco R,
Collins MK, Hanniford D, Vega-Saenz de Miera E, Rakus JF, Dankert
JF, Shang S, Kerbel RS, et al: miR-30b/30d regulation of GalNAc
transferases enhances invasion and immunosuppression during
metastasis. Cancer Cell. 20:104–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He Z, Yu L, Luo S, Li M, Li J, Li Q, Sun Y
and Wang C: miR-296 inhibits the metastasis and
epithelial-mesenchymal transition of colorectal cancer by targeting
S100A4. BMC Cancer. 17:1402017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Panera N, Crudele A, Romito I, Gnani D and
Alisi A: Focal adhesion kinase: Insight into molecular roles and
functions in hepatocellular carcinoma. Int J Mol Sci.
18:992017.doi: 10.3390/ijms18010099. View Article : Google Scholar :
|