Introduction
Esophageal carcinoma (EC) constitutes one of the
most common types of cancer with high mortality and incidence
worldwide (1). It is ranked fourth
in mortality in China. Esophageal squamous cell carcinoma (ESCC) is
the predominant histological type and represents approximately 90%
of all cases in China (2). ESCC
patients always have a low survival rate since they are diagnosed
at an advanced stage or even with distant metastasis (3,4).
Metastasis has become the main cause of death in these patients.
Therefore, elucidating the molecular mechanism concerning its
biological behaviour, such as metastasis, is essential for EC
treatment.
MicroRNAs (miRNAs) are approximately 20–22
nucleotides molecules and have been reported to play an important
role in tumorigenesis and development in certain cancers including
ESCC (5–9). MircroRNA-21 (miR-21) has been revealed
to be highly expressed in many solid tumors such as ESCC, oral
squamous cell carcinoma (OSCC), and breast, lung and gastric cancer
(10–17). It may therefore be considered as a
new biomarker for cancer diagnosis and a new target for cancer
treatment. A recent study revealed that serum miR-21 was related to
the progression of ESCC (12).
Studies on miR-21 revealed that it regulated the invasion,
migration and metastasis of cells (such as A549 and PC-3) (9,18) and
epithelial-mesenchymal transition (EMT), which is considered as the
first step to metastasis (19–27).
The biological behaviors of cells such as proliferation, migration
and invasion have been regarded as precursors to metastasis. A
previous study revealed that the expression of miR-21 was
significantly altered in patients with ESCC as determined by
microarray (28). miRNAs have been
demonstrated to play a regulatory role by interacting with their
target genes. The results from accurate prediction software
(TargetScan, miRwalk, miRanda and PITA) revealed that RAS p21
protein activator 1 (RASA1) may be the target gene of miR-21.
However, the role of miR-21 and its target gene in regulating the
biological behavior of ESCC has yet to be elucidated.
In the present study, cell colonies assay, wound
healing and Transwell assay were used to detect the role of miR-21
and its target gene in cell proliferation, migration and invasion
of ESCC. We examined the interaction between miR-21 and RASA1 using
a luciferase reporter assay. The relative gene expression levels of
RASA1 and miR-21 were detected by quantitative polymerase chain
reaction (q-PCR). Finally, the effect of miR-21 on ESCC growth
in vivo was examined in a nude mouse model. Generally, in
the present study, we aimed to identify the role of miR-21 in the
regulation of ESCC cells and to reveal some new targets for cancer
treatment.
Materials and methods
Microarray data
Gene expression profiles of GSE13937 were downloaded
from the GEO repository. These data were based on the GPL8835
platform. The data of 44 patients with ESCC (including 44 samples
of normal adjacent esophageal tissues and 44 samples of tumor
tissues) were chosen to perform further analysis and create the
heat maps with Morpheus online tool (https://software.broadinstitute.org/morpheus/). Then,
the top 20 of differentially expressed (DE) miRNAs were screened.
This study was published by Mathé et al (28). Total RNA was extracted and detected
by miRNA microarray chips. Version 3. R (BioConductor) (http://www.bioconductor.org/install/)
was used for background correction and normalization of the
data.
Sample collection
All samples were collected at the Panyu Central
Hospital and the Third Affiliated Hospital of Southern Medical
University from February 27, 2010 to May 2, 2017. One patient was
female and 19 patients were male, with a mean age of 62.4±5.8
years, and diagnosed with clinicopathological characteristics of
ESCC. These samples were used only for the detection of the gene
expression of miRNA and 10 patient samples among the 20 were
utilized for target gene detection. All samples were stored at
−80°C after collection.
Ethics statement
This study was approved by the Ethics Committee of
Panyu Central Hospital (Guangzhou, China). All patients agreed to
participate provided informed consent for this study.
Cell culture and reagents
The cell line Eca-109 (human esophageal squamous
carcinoma cell) with metastatic ability and 293T cell were obtained
from the State Laboratory of Oncology in South China, Sun Yat-Sen
University Cancer Center (Guangzhou, China). The cell line KYSE510
(well differentiated squamous cell carcinoma cell line) was
obtained from the Central Laboratory, Nanfang Hospital, Southern
Medical University (Guangzhou, China). The Eca-109 and KYSE510
cells were cultured in RPMI-1640 media, supplemented with 10% fetal
calf serum (FCS; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and were maintained under 5% CO2 at 37°C. The
293T cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.), supplemented with
10% FCS and 1% GlutaMAX and 1% penicillin with streptomycin.
Animals
Ten BALB/C nude mice aged 4–6 weeks old with a male
to female ratio of 1:1 weighing 20–25 g were used in the present
study. These mice were obtained from Chase Reward Ltd. (Guangzhou,
China) and kept under the specific pathogen-free conditions
(temperature, 20–26°C; 12-h light-dark cycle; aseptic food and
water). The animal experiments were approved by the Ethics
Committee of Panyu Central Hospital (no. K20170002).
Cell transfection
miR-21 mimics (overexpression), miR-21 inhibitor
(downregulation) and negative control (NC including mimic NC,
inhibitor NC) were purchased from Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). Transfections were performed on Eca-109 cells
with Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's protocol. Briefly,
5×105 cells were cultured in 6-well plates for 24 h
prior to transfection. The miR-21 inhibitor, mimics and their NC
nucleotides were added to the cell with transfection reagent to a
final concentration of 50 and 100 nm/l, respectively. The cells
were treated for 48 h and harvested for analysis.
Design and construction of eukaryotic
expression vector for RASA1
miRWalk (miR Walk1.0) (http://zmf.umm.uniheidelberg.de/apps/zmf/mirwalk/micrornapredictedtarget.html)
was utilized to predict the target gene and RASA1 was predicted as
the target gene of miR-21. Thus, the vector for RASA1 was
constructed. The 3′ untranslated regions (3′UTR) of RASA1 mRNA were
amplified using the PCR instrument (GeneAmp PCR System 2400;
Applied Biosystems; Thermo Fisher Scientific, Inc.), which was
bound to the mature miR-21 (MIMAT0000076). To prevent formation of
a termination signal, ‘taagcta’ was selected as the region in a
mutational expression vector template and was amplified
(amplification primers are presented in Table I). Then, the aforementioned template
was transfected with psi-CHECK-2 luciferase plasmid and used for
dual-luciferase reporter assay.
 | Table I.The primer sequence of the genes. |
Table I.
The primer sequence of the genes.
| Gene | Primer (5′-3′) |
|---|
| RASA13′ | UTR-F:
ccgctcgagCAGCCTTCGCCCCAGTGTTCTG |
| RASA13′ | UTR-R:
ataagaatgcggccgcTAATCAATTATGCAAGATATCCC |
| mutation
RASA1: |
|
| RASA13′ | UTR-mF:
GTGAATAACTATGCCAGCAACCTTGATTCGATTCTGTGCAGGATATTTGCACTATTT |
| RASA13′ | UTR-mR:
ataagaatgcggccgcTAATCAATTATGCAAGATATCCC |
| RASA1-F: |
GAACTTGGGAATGTACCTGAAC |
| RASA1-R: |
TGTGCACCACGCTCATTAC |
| miR-21-F: |
ACACTCCAGCTGGGTAGCTTATCAGACTGATG |
| miR-21-RT: |
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCAACATC |
| miRNA-R: |
CTCAACTGGTGTCGTGGA |
| U6-F: |
CTCGCTTCGGCAGCACA |
| U6-R: |
AACGCTTCACGAATTTGCGT |
| K-ras-F: |
GAGGCCTGCTGAAAATGAC |
| K-ras-R: |
GCTGTGTCGAGAATATCCAA |
| AKT-F: |
ATCGCTTCTTTGCCGGTATC |
| AKT-R: |
CTTGGTCAGGTGGTGTGATG |
| PI3K-F: |
AGGGAGCGAGTGCCTTTTAT |
| PI3K-R: |
AAGCCCTGCAGTCAACATCA |
| E-caherin-F: |
GTACTCAAAGCCCAGAATCC |
| E-caherin-R: |
CCCTCAACTAACCCCCTTTA |
| Vimentin-F: |
CGCCAGATGCGTGAAATGG |
| Vimentin-R: |
ACCAGAGGGAGTGAATCCAGA |
| Snail-F: |
TGGTTGCTTCAAGGACACAT |
| Snail-R: |
GTTGCAGTGAGGGCAAGAA |
| 18s rRNA-F: |
CCTGGATACCGCAGCTAGGA |
| 18s rRNA-R: |
GCGGCGCAATACGAATGCCCC |
| β-actin-F: |
ACTCTTCCAGCCTTCCTTCC |
| β-actin-R: |
GTACTTGCGCTCAGGAGGAG |
Dual-luciferase reporter assay
The 3′UTR fragments of the RASA1 gene were amplified
and then transfected and cloned into the psi-CHECK-2 luciferase
miRNA expression reporter vector. The 293T cells (2×104)
were cultured in 24-well plates and transfected with 50 nM miR-21
mimics, 100 nM inhibitor or NC, 0.5 µg of psi-CHECK-2 luciferase
reporter vector that contained the wild-type or mutant 3′UTR of
RASA1 or empty plasmid. Transfections were performed using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Forty-eight hours
after transfection, the luciferase activity was assessed by the
Dual-Luciferase Reporter Assay System (GloMax; Promega Corporation,
Madison, WI, USA). The experiments were repeated in 3 independent
experiments in duplicate.
Cell colonies assay
Eca-109 cells (100) were cultured in 6-well plates
until colonies could be observed. Then the cells were fixed by 4%
of triformol and stained with Giemsa for 15 min at room temperature
(Nanjing Jiancheng Taihao Biotechnology Co., Ltd., Nanjing, China).
Finally, the clones were counted under a light microscope
(magnification, ×100). The clone which contained >10 cells was
regarded as one colony formation, and the colony rate was
calculated using the following formula: colony formation rate =
(colony formation numbers/100) × 100%.
Scratch assay
Eca-109 cells (8.0×105) were cultured in
6-well plates for 24 h and confluence (>90%) without any vacant
space. Subsequently the cells were scratched using 10-µl tips with
a wound midline of the culture well, and then replaced in 1% FBS
DMEM. Finally, the difference in width of the wounds was measured
at 0, 24 and 48 h, respectively to evaluate the migration of cells
and the healing rate was calculated.
Cell migration and invasion assay
Cell migration activities were detected using
Transwell migration. After the transfected Eca-109 or KYSE510 cells
were cultured for 24 h, the cells (1×105) were
resuspended in serum-free media and cultured in the inserts (8-µm
diameter pore size; Corning Inc., Corning, NY, USA), which were
placed in the 6-wells plates with 10% FBS serum media. The cells
migrated from the upper surface to the lower surface of the
membrane after 48-h incubation. Subsequently, the migrated cells
were fixed with 100% methanol and stained with 1% toluidine.
Finally, these stained cells were counted in 5 random optical
fields under a light microscope (magnification, ×200). The cell
invasion assay was similar to the cell migration assay except the
Transwell membrane was pre-coated with 24 mg/ml Matrigel (Corning
Inc.).
Quantitative real-time PCR
Transfected Eca-109 cells and tissues were processed
in TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and
total RNA was extracted according to the manufacturer's
instructions. First-strand cDNA was synthesized with 1 µg total RNA
per sample using the PrimeScript™ RT Synthesis system (Takara Bio,
Inc., Shiga, Japan). Subsequently, the cDNA sample was amplified
using QuantiTect reagents (GeneCopoeia, Inc., Rockville, MD, USA)
in a final volume of 20 µl under the LightCycler 480II detector
(Roche Diagnostics, Basel, Switzerland). The amplifications were
performed as follows: predestination for 2 min at 50°C,
denaturation for 30 sec at 95°C, followed by 40 cycles of 95°C for
5 sec, and 60°C for 34 sec. The melting curve analysis was
performed to detect the specificity of amplification products. The
experiments were performed in triplicate. U6 and 18S rRNA genes
were used as the miRNA-21 and RASA1 endogenous reference controls,
respectively. β-actin was used for the rest of the genes as an
endogenous reference control. The relative gene expression level
was calculated using the 2−ΔΔCq method (29). Specific sense primers are presented
in Table I.
Construction of a tumor-burdened mouse
model and in vivo treatment
The Eca-109 (2×106) cells were
subcutaneously injected in the posterior right gluteal area. When
the tumor volume reached ~0.1 mm3, the mice were
randomly assigned into 2 groups. One group received an injection
with antagomir NC (10 nmol, 50 µl) and the other antagomir (10
nmol, 50 µl) every 3 days with 5 mice in each group. The tumor
volume was measured before and every 3 days after treatment. The
volume was calculated with the following formula: V = 1/2 (a ×
b2), in which a was the long diameter and b was the
short diameter of the tumor. Twenty-one days after treatment, the
mice were euthanized with CO2 and the tumor tissues were
dissected.
Data analysis
The data was calculated and analyzed with Excel and
SPSS 13.0 packages (SPSS, Inc., Chicago, IL, USA). Data was
expressed as the mean ± standard deviation (SD) of separate
experiments. Statistical analysis was presented by the repeated
measure, independent-samples t-test and one-way analysis of
variance (ANOVA) followed by Dunnett's post hoc test. P<0.05 was
regarded as a significant difference.
Results
Identification DE miRNAs
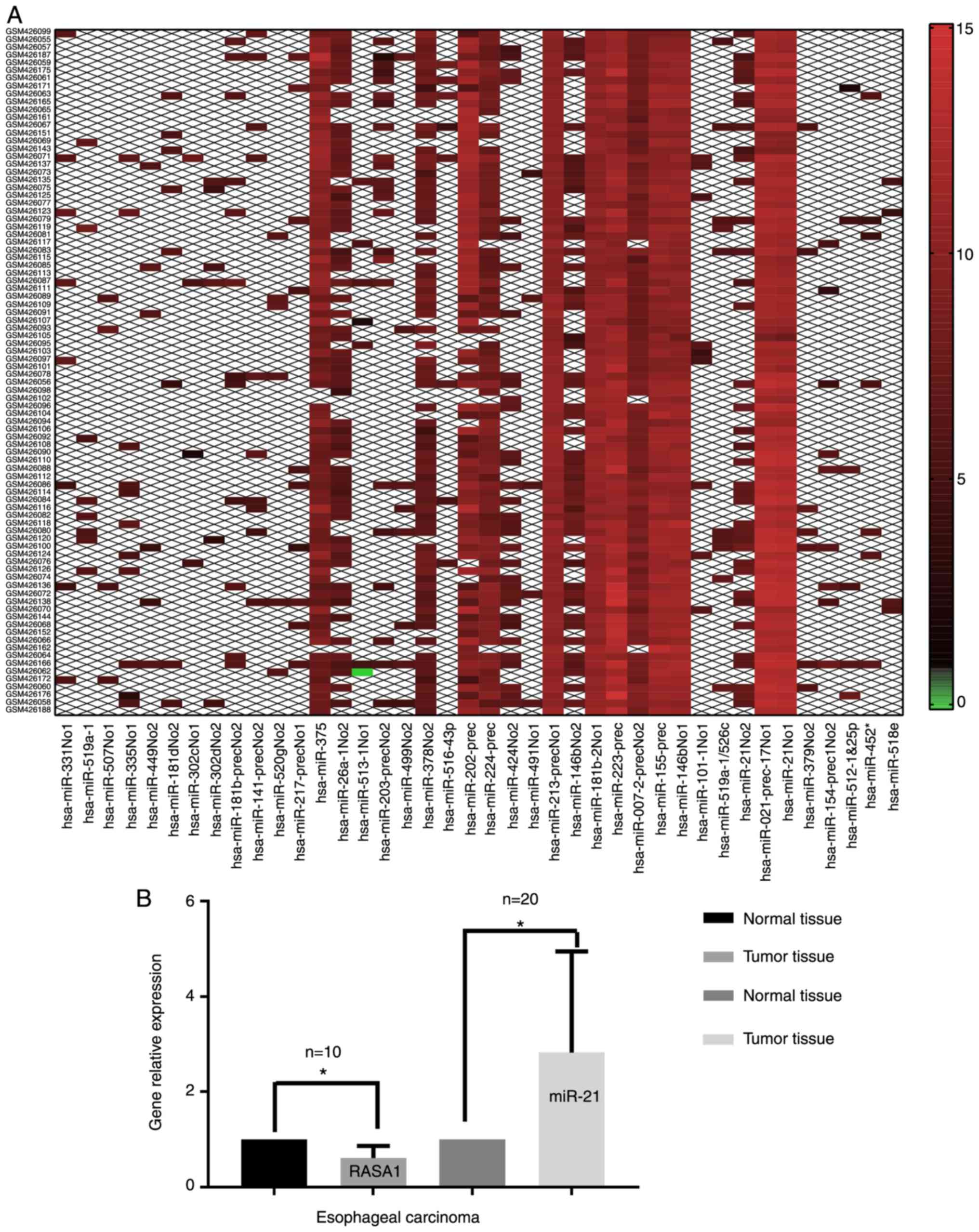
A total of 627 DE miRNAs were identified. The top 20
DE miRNAs (top 20 upregulated and 20 downregulated miRNAs) was
analyzed with the Morpheus online tool to form a heat map, which
was screened by SNR value. The heat map revealed that miR-21 was
one of the principal genes and was selected for further research
(Fig. 1).
Overexpressed miR-21 enhances the cell
colony formation rate and cell healing rate
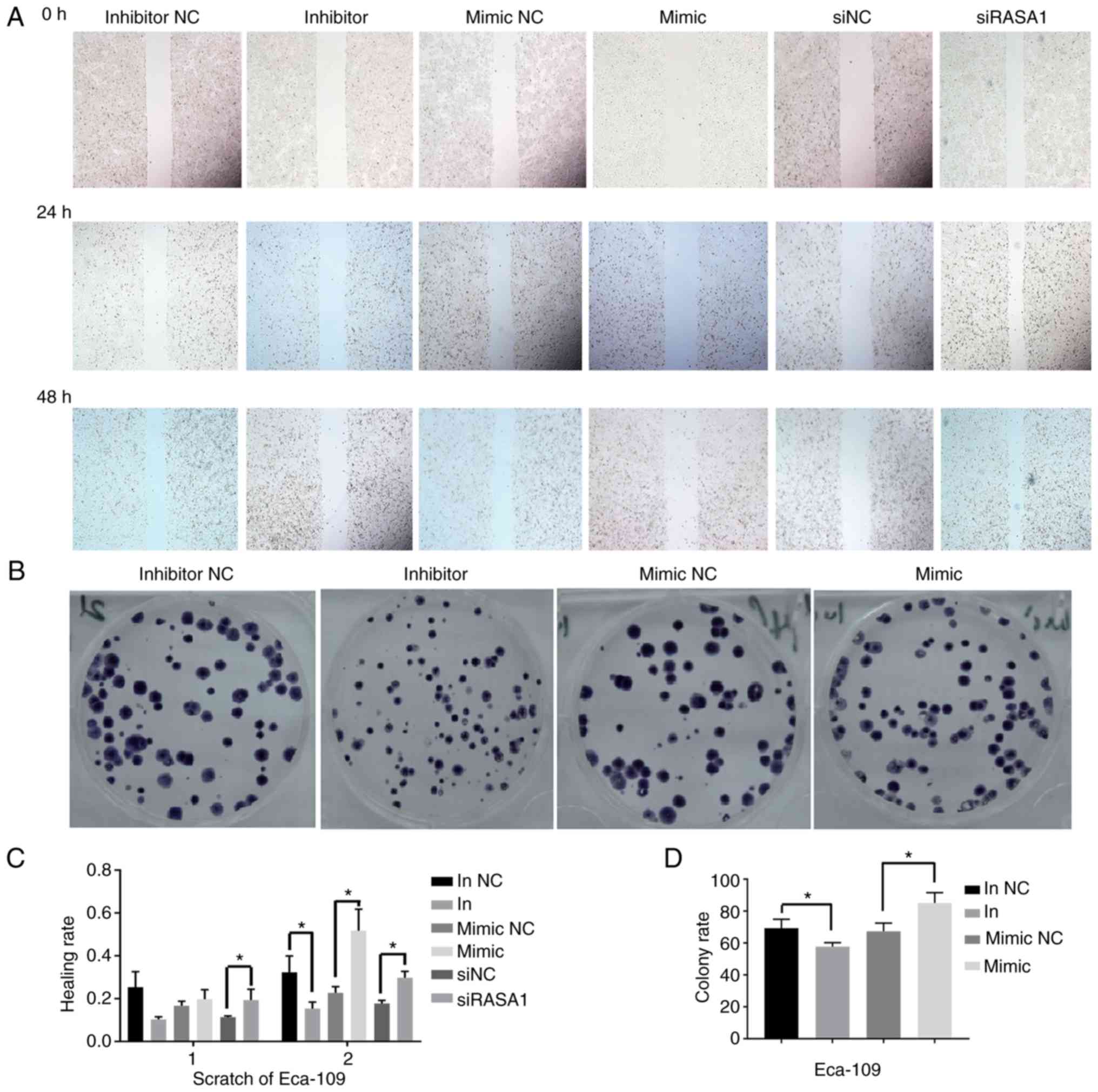
The cell colonies assay revealed that the cell
colony formation rate was significantly decreased in the miR-21
inhibitor group compared with the inhibitor NC group, while the
rate was significantly increased in the miR-21 mimic group compared
with the mimic NC group (P<0.05) (Fig. 2). The results of the scratch assay
revealed that, the cell healing rate was significantly inhibited in
the miR-21 inhibitor group compared with the inhibitor NC group
after the cells were transfected for 48 h. Conversely, the healing
rate was significantly enhanced in the miR-21 mimic group and the
siRAS1 group compared with the mimic NC and siNC group,
respectively (all P<0.05) (Fig.
2).
Cell migration and invasion assay
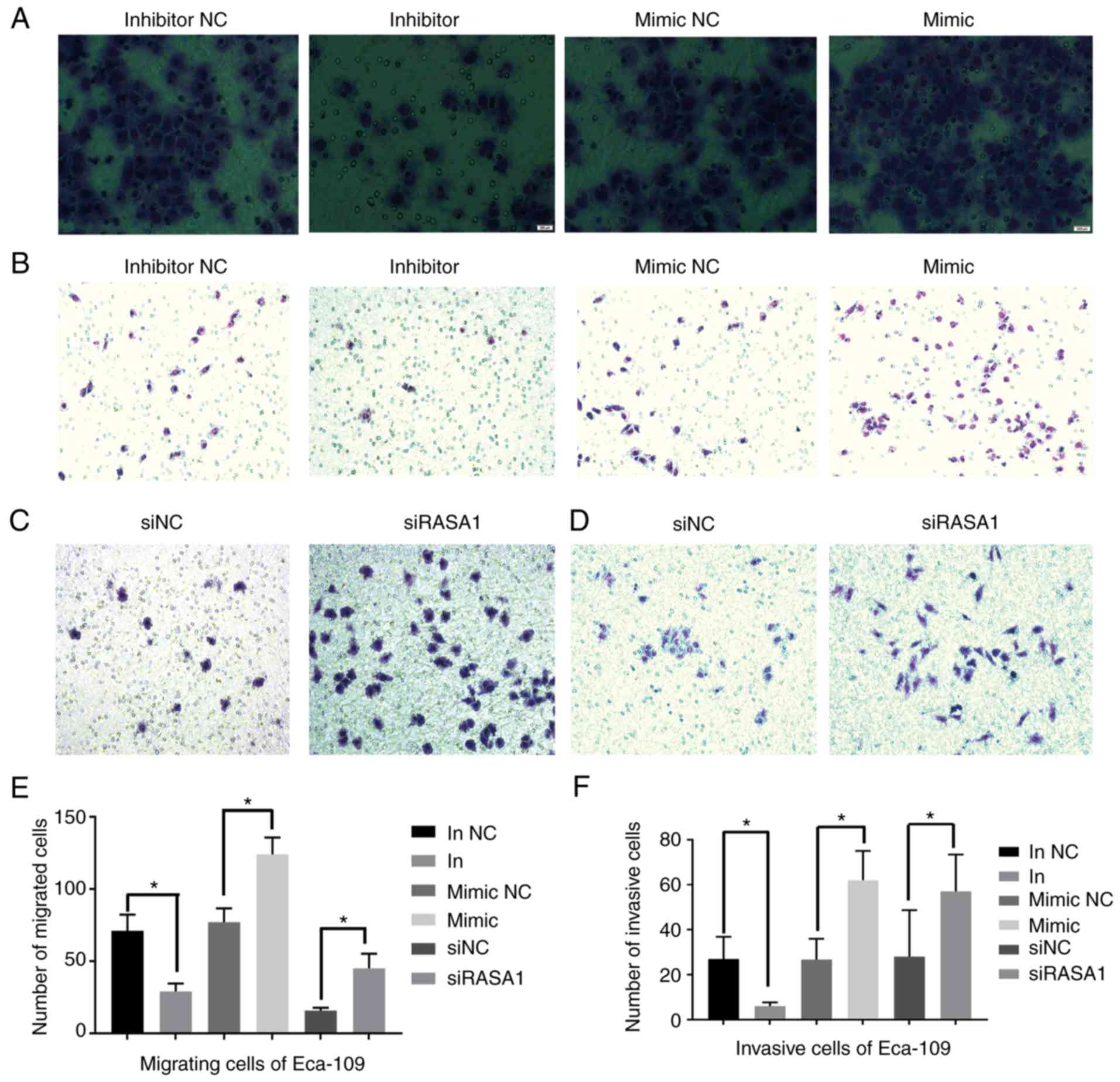
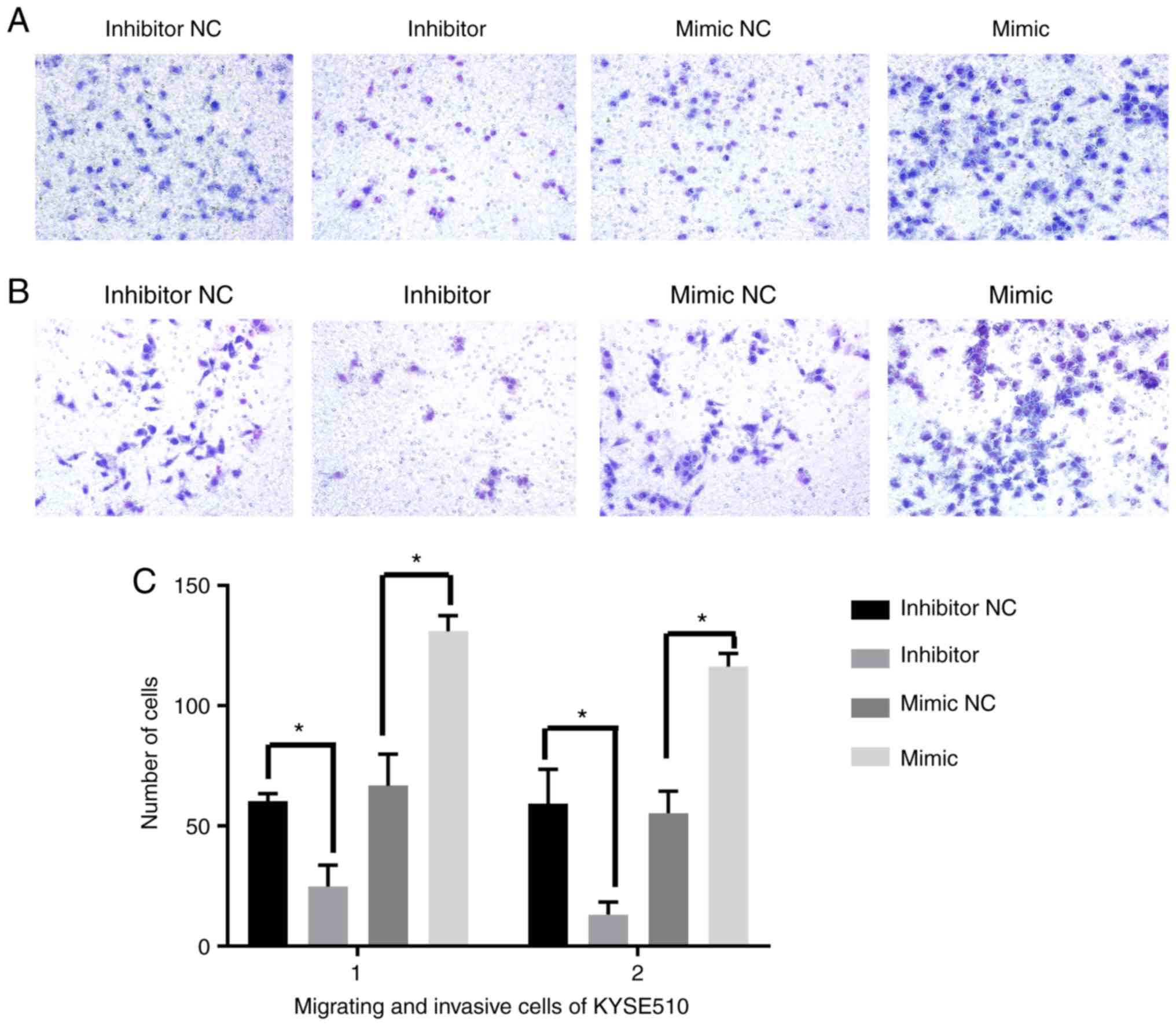
The number of migrated and invasive cells was
significantly decreased in the miR-21 inhibitor group compared with
the inhibitor NC group (all P<0.05). Conversely, the cells were
significantly increased in the miR-21 mimic group compared with the
mimic NC group (all P<0.05). Notably, we observed that the
number of migrated and invasive Eca-109 cells was also
significantly increased in the siRASA1 group compared with the siNC
group (all P<0.05). Similar results were also obtained with the
KYSE510 cells (Figs. 3 and 4).
Construction of a vector for
RASA1
The sequence of RASA1 3′UTR which was combined to
miR-21 (mature sequence: UAGCUUAUCAGACUGAUGUUGA) was amplified and
detected at 1043 bp using electrophoresis. The vector with mutated
or wild-type RASA1 was constructed and used for the luciferase
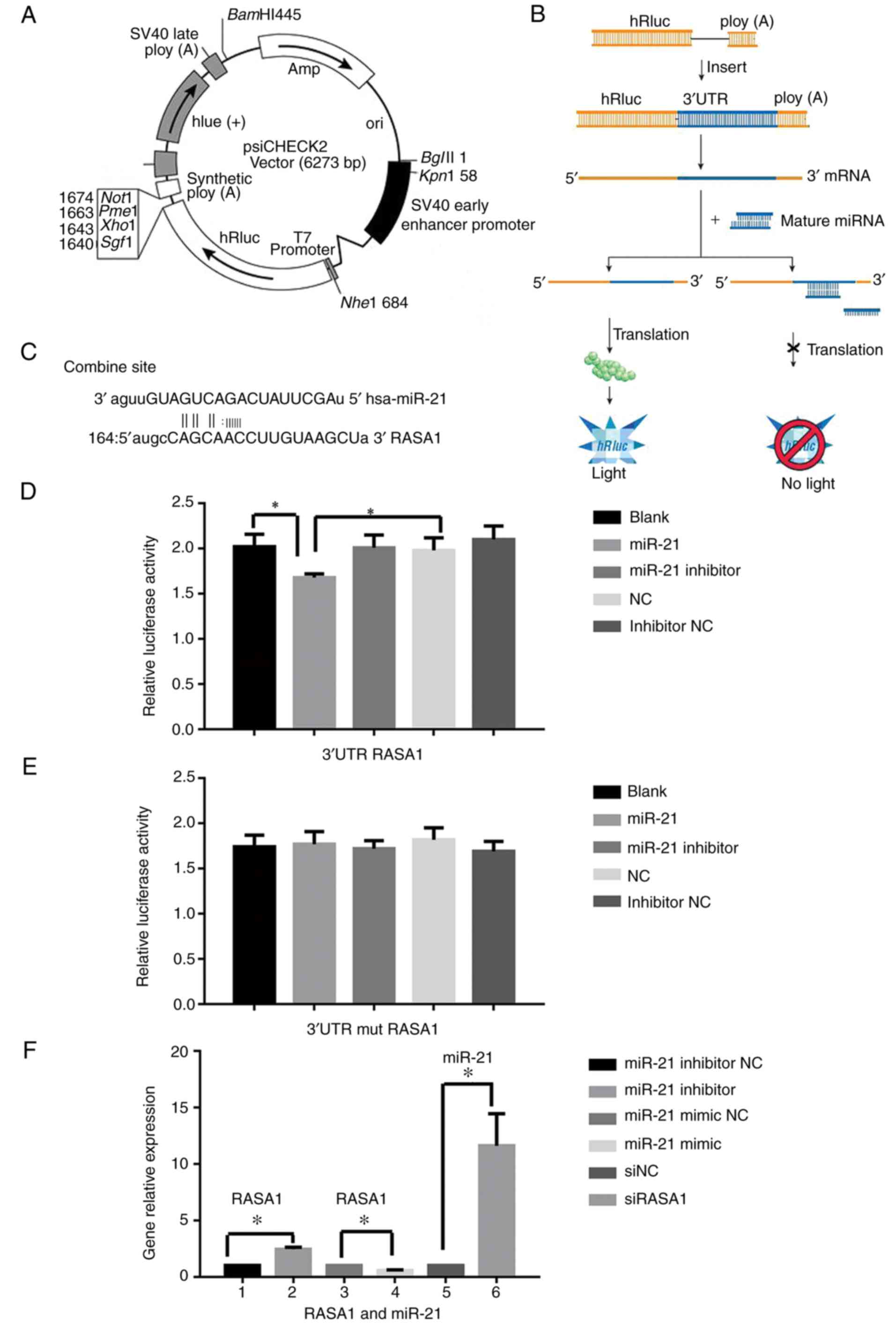
reporter assay (Fig. 5A-C).
Dual-luciferase reporter assay
The result of the dual-luciferase reporter assay
revealed that the luciferase activity in the miR-21 group
transfected with the RASA1 vector was significantly inhibited
compared with the NC and the blank group (P<0.05) (Fig. 5D), while the activity in the miR-21
group transfected with the RASA1 mutated vector was not
significantly altered compared with the NC and the blank group
(P>0.05) (Fig. 5E).
q-PCR results
It predicted that RASA1 may be the target gene of
miR-21 by miRWalk. Their relative expression by PCR revealed that
miR-21 was significantly increased and RASA1 was decreased in the
tumor tissues compared to the normal adjacent tissues in patients
with ESCC (Fig. 1B). The gene level
of RASA1 was significantly increased when the Eca-109 cells were
transfected with the miR-21 inhibitor compared with the inhibitor
NC (P<0.05). Conversely, the expression was significantly
reduced in the miR-21 mimic group when compared with the mimic NC
group (P<0.05). The expression of miR-21 was significantly
increased in the group of cells which were transfected with siRASA1
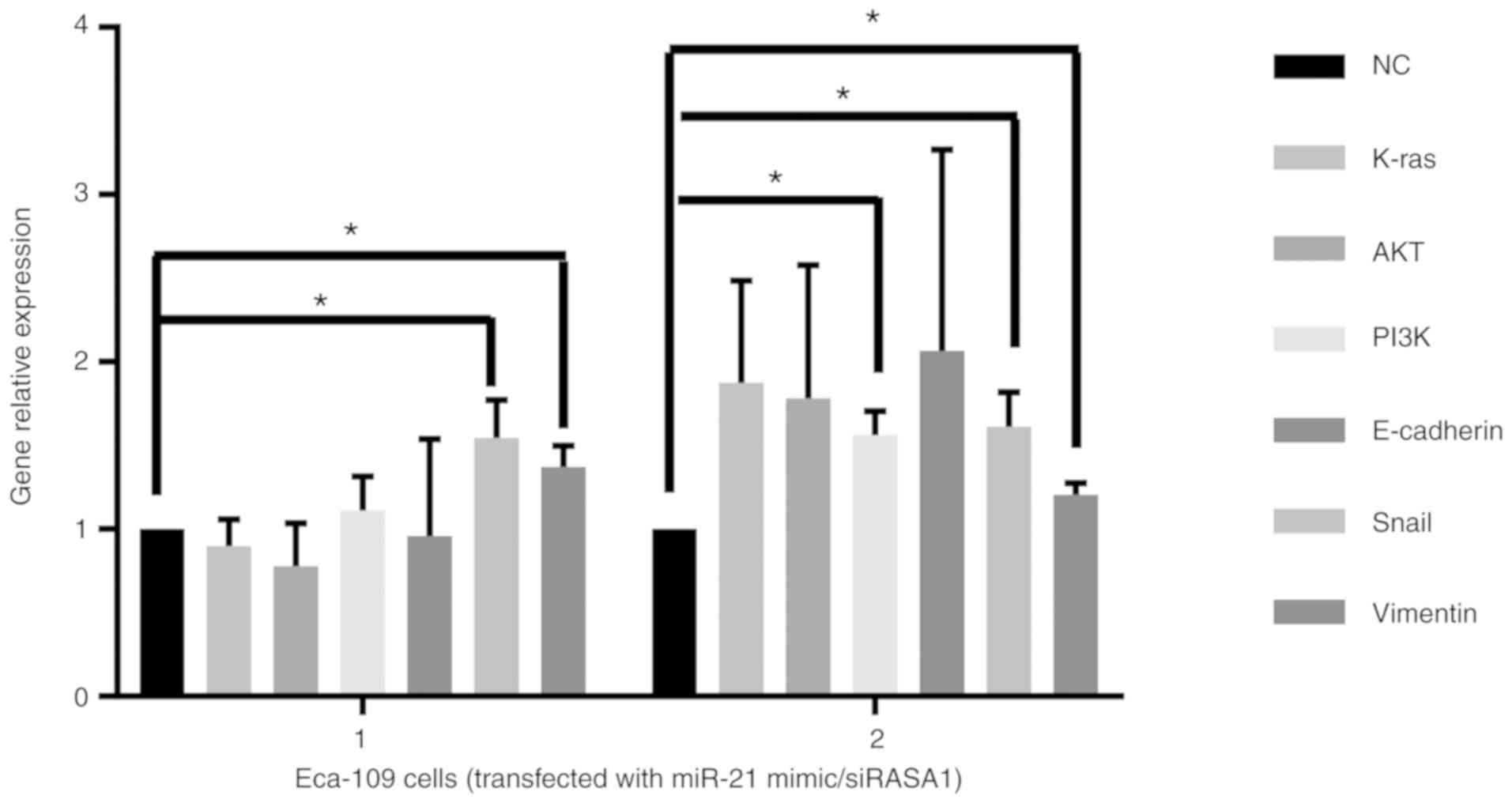
when compared with the siNC group (P<0.05) (Fig. 5F). The relative gene expression of
Snail and vimentin was significantly increased by upregulated
miR-21 or downregulated RASA1 (Fig.
6).
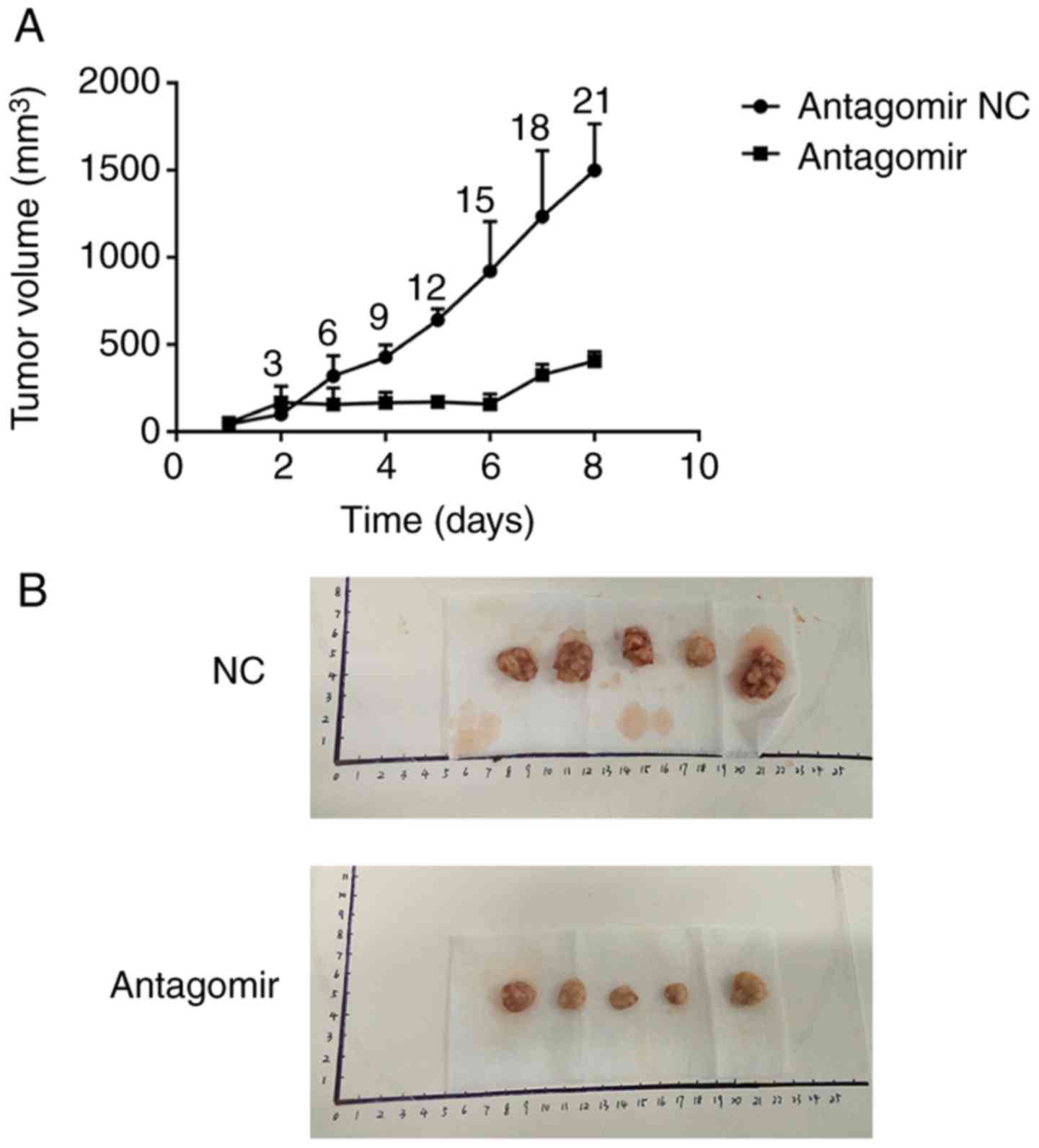
The effect of miR-21 in tumor
xenografts
Nude mice bearing tumors were established to
evaluate the effects of miR-21 inhibitor (antagomir) on esophageal
carcinoma growth in vivo. Compared to the NC group,
application of antagomir significantly inhibited the growth of the
tumor 6 days after treatment (P<0.05). The mice volume in the 2
groups was not significantly different before treatment (P>0.05)
(Fig. 7). No diarrhea or body
weight loss was observed in any of the animals.
Discussion
Recent studies have revealed that miRNAs play an
important role in tumorigenesis and progression. They are involved
in multiple biological processes, including tumor invasion,
migration, proliferation and metastasis. miR-21 has been reported
to be overexpressed in certain cancers including ESCC patients and
may become a new biomarker for tumor diagnosis. A study by Winther
et al (30) revealed that
miR-21 could be identified as an independent prognostic biomarker
for disease-specific survival (DSS) in esophagogastric
adenocarcinomas (EAC) patients. Lv et al (31) revealed that miR-21 was overexpressed
in Kazak or Uighur EC patients and was an independent factor for
prognosis. Considering its pivotal role for diagnosis, researchers
drew attention to the role of miR-21 in tumor processes such as
invasion, migration and metastasis. Shen et al (9) found that solasodine could downregulate
the expression of miR-21 and increase its target gene RECK to
inhibit the invasion of A549 cells. Yang et al (18) studied the biological behaviors of
miR-21 in prostate cancer cell lines PC-3 and found that miR-21
could promote cell proliferation and invasion by overexpression of
miR-21 to target PTEN. As aforementioned, miR-21 is expected to
become a new target in cancer treatment (4,32,33–35).
However, the role that miR-21 plays in the biological behavior of
ESCC has yet to be elucidated.
In the present study, we concentrated on the role of
miR-21 and its target gene in the biological behavior of ESCC cell
lines. From the data of microarray analysis, miR-21 was found to be
one of the 20 top DE miRNAs (upregulated) in patients with ESCC.
Our results also revealed that the relative expression of miR-21
was significantly increased in tumor tissues compared to normal
adjacent tissues. Furthermore, the results revealed that RASA1 may
be a target gene of miR-21 and that miR-21 and its target gene may
play an important role in regulating ESCC. Thus, a functional
experiment of miR-21 and RASA1 was performed on ESCC cell lines.
Notably, we observed that overexpressed miR-21 could significantly
enhance the healing rate and increase the number of invasive and
migrated cells as well as the cell colony formation rate.
Conversely, downregulation of the expression of miR-21 could
significantly inhibit the healing rate and decrease the number of
invasive and migrated cells as well as the cell colony formation
rate. Futhermore, downregulated RASA1 (siRASA1), significantly
increased the number of invasive and migrated cells. These results
indicated that miR-21 and its target gene RASA1 regulated cell
invasion, migration and proliferation of ESCC cells.
It has been reported that miRNAs negatively regulate
their target gene via binding to the 3′UTR of target mRNAs, which
causes mRNA degradation suppressing translation (31). In the present study, we observed
that overexpression of miR-21 significantly decreased the
expression of RASA1. Conversely, downregulated miR-21 significantly
increased the RASA1 gene expression. These results revealed that
miR-21 negatively regulated RASA1. In order to further elucidate
the relationship between RASA1 and miR-21, we constructed an mRNA
vector of RASA1 combined to mature miR-21 and performed a
luciferase reporter assay. The assay revealed that RASA1 was
combined with miR-21. With the PCR results, the relative gene
expression of Snail and vimentin was significantly increased by
upregulated miR-21 or downregulated RASA1. These results indicated
that miR-21 directly targeted RASA1, and that the regulatory role
may be via Snail and vimentin. RASA1 was firstly discovered as a
RAS guanosine triphosphate enzyme-activated protein, which
inactivated RAS to inhibit the RAS downstream pathway, thereby
resulting in the proliferation, migration, invasion and metastasis
of cancer cells. Snail is a pivotal gene which is related to EMT.
Thus, we hypothesized that miR-21 regulated migration and invasion
by directly targeting RASA1 which may be via regulation of Snail.
However, the precise mechanism in the downstream pathway requires
further research.
In order to investigate the effect of miR-21 in
vivo, we set up a nude mouse tumor-bearing model. We observed
that application of miR-21 inhibitor significantly inhibited the
growth of the tumor compared to the NC group at days 3 and 6 after
treatment, respectively. This indicated that anti-miR-21 had an
antitumor effect and may be considered as a promising target for
ESCC therapy.
In conclusion, miR-21 regulated cell proliferation,
migration, invasion and tumor growth of ESCC by directly targeting
RASA1, possibly via regulation of Snail. miR-21 and RASA1 may be
considered as possible targets for ESCC therapy.
Acknowledgements
The authors thank Professor Edward I. Wong (Milton
International Education Group, Hong Kong) for revising the
manuscript.
Funding
The present study was supported by grants from the
Technical New Star of Zhujiang, Panyu District, Guangzhou
(2013-special-15-6.10), the Science and Technology Program of Panyu
(2015-Z03-09) and the Science and Technology Program of Guangzhou
(grant no. 201804010012).
Availability of data and materials
The data used during the study are available from
the corresponding author upon reasonable request.
Authors' contributions
XCh and SC performed the analysis of the data and
wrote the paper. XCa and XZ performed the data analysis of GEO. LW
and HL collected the samples and the experimental data. SC, BL, LW,
WL and ZW performed the experiments. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Panyu Central Hospital (Guangzhou, China). All patients provided
informed consent to participate in this study. The animal
experiments were approved by the Ethics Committee of Panyu Central
Hospital (no. K20170002).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tran GD, Sun XD, Abnet CC, Fan JH, Dawsey
SM, Dong ZW, Mark SD, Qiao YL and Taylor PR: Prospective study of
risk factors for esophageal and gastric cancers in the Linxian
general population trial cohort in China. Int J Cancer.
113:456–463. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ando N, Kato H, Igaki H, Shinoda M, Ozawa
S, Shimizu H, Nakamura T, Yabusaki H, Aoyama N, Kurita A, et al: A
randomized trial comparing postoperative adjuvant chemotherapy with
cisplatin and 5-fluorouracil versus preoperative chemotherapy for
localized advanced squamous cell carcinoma of the thoracic
esophagus (JCOG9907). Ann Surg Oncol. 19:68–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu EH, Tu HF, Wu CH, Yang CC and Chang KW:
MicroRNA-21 promotes perineural invasion and impacts survival in
patients with oral carcinoma. J Chin Med Assoc. 80:383–388. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Teoh SL and Das S: The role of microRNAs
in diagnosis, prognosis, metastasis and resistant cases in breast
cancer. Curr Pharm Des. 23:1845–1859. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tseng HH, Tseng YK, You JJ, Kang BH, Wang
TH, Yang CM, Chen HC, Liou HH, Liu PF, Ger LP, et al:
Next-generation sequencing for microRNA profiling: MicroRNA-21-3p
promotes oral cancer metastasis. Anticancer Res. 37:1059–1066.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cao J, Liu J, Xu R, Zhu X, Liu L and Zhao
X: MicroRNA-21 stimulates epithelial-to-mesenchymal transition and
tumorigenesis in clear cell renal cells. Mol Med Rep. 13:75–82.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li P, Mao WM, Zheng ZG, Dong ZM and Ling
ZQ: Down-regulation of PTEN expression modulated by dysregulated
miR-21 contributes to the progression of esophageal cancer. Dig Dis
Sci. 58:3483–3493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen KH, Hung JH, Chang CW, Weng YT, Wu MJ
and Chen PS: Solasodine inhibits invasion of human lung cancer cell
through downregulation of miR-21 and MMPs expression. Chem Biol
Interact. 268:129–135. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Z, Yu T, Cabay RJ, Jin Y, Mahjabeen
I, Luan X, Huang L, Dai Y and Zhou X: miR-486-3p, miR-139-5p, and
miR-21 as biomarkers for the detection of oral tongue squamous cell
carcinoma. Biomark Cancer. 9:1–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mei LL, Qiu YT, Zhang B and Shi ZZ:
MicroRNAs in esophageal squamous cell carcinoma: Potential
biomarkers and therapeutic targets. Cancer Biomark. 19:1–9. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
He Y, Jin J, Wang L, Hu Y, Liang D, Yang
H, Liu Y and Shan B: Evaluation of miR-21 and miR-375 as prognostic
biomarkers in oesophageal cancer in high-risk areas in China. Clin
Exp Metastasis. 34:73–84. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han JG, Jiang YD, Zhang CH, Yang YM, Pang
D, Song YN and Zhang GQ: A novel panel of serum
miR-21/miR-155/miR-365 as a potential diagnostic biomarker for
breast cancer. Ann Surg Treat Res. 92:55–66. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Markou A, Zavridou M and Lianidou ES:
miRNA-21 as a novel therapeutic target in lung cancer. Lung Cancer.
7:19–27. 2016.PubMed/NCBI
|
|
15
|
Dong J, Zhang Z, Gu T, Xu SF, Dong LX, Li
X, Fu BH and Fu ZZ: The role of microRNA-21 in predicting brain
metastases from non-small cell lung cancer. Onco Targets Ther.
10:185–194. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren J, Kuang TH, Chen J, Yang JW and Liu
YX: The diagnostic and prognostic values of microRNA-21 in patients
with gastric cancer: A meta-analysis. Eur Rev Med Pharmacol Sci.
21:120–130. 2017.PubMed/NCBI
|
|
17
|
Arantes LM, Laus AC, Melendez ME, de
Carvalho AC, Sorroche BP, De Marchi PR, Evangelista AF,
Scapulatempo-Neto C, de Souza Viana L and Carvalho AL: MiR-21 as
prognostic biomarker in head and neck squamous cell carcinoma
patients undergoing an organ preservation protocol. Oncotarget.
8:9911–9921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Guo JX and Shao ZQ: miR-21 targets
and inhibits tumor suppressor gene PTEN to promote prostate cancer
cell proliferation and invasion: An experimental study. Asian Pac J
Trop Med. 10:87–91. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huo W, Zhao G, Yin J, Ouyang X, Wang Y,
Yang C, Wang B, Dong P, Wang Z, Watari H, et al: Lentiviral
CRISPR/Cas9 vector mediated miR-21 gene editing inhibits the
epithelial to mesenchymal transition in ovarian cancer cells. J
Cancer. 8:57–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu CH, Huang Q, Jin ZY, Zhu CL and Liu
Zand Wang C: miR-21 and KLF4 jointly augment epithelial-mesenchymal
transition via the Akt/ERK1/2 pathway. Int J Oncol. 50:1109–1115.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li C, Song L, Zhang Z, Bai XX, Cui MF and
Ma LJ: MicroRNA-21 promotes TGF-β1-induced epithelial-mesenchymal
transition in gastric cancer through up-regulating PTEN expression.
Oncotarget. 7:66989–67003. 2016.PubMed/NCBI
|
|
22
|
Yan L, Cao R, Liu Y, Wang L, Pan B, Lv X,
Jiao H, Zhuang Q, Sun X and Xiao R: MiR-21-5p links
epithelial-mesenchymal transition phenotype with stem-like cell
signatures via AKT signaling in keloid keratinocytes. Sci Rep.
6:282812016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun SS, Zhou X, Huang YY, Kong LP, Mei M,
Guo WY, Zhao MH, Ren Y, Shen Q and Zhang L: Targeting STAT3/miR-21
axis inhibits epithelial-mesenchymal transition via regulating CDK5
in head and neck squamous cell carcinoma. Mol Cancer. 14:2132015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu ZH, Tao ZH, Zhang J, Li T, Ni C, Xie J,
Zhang JF and Hu XC: MiRNA-21 induces epithelial to mesenchymal
transition and gemcitabine resistance via the PTEN/AKT pathway in
breast cancer. Tumour Biol. 37:7245–7254. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Z, Jin ZY, Liu CH, Xie F, Lin XS and
Huang Q: MicroRNA-21 regulates biological behavior by inducing EMT
in human cholangiocarcinoma. Int J Clin Exp Pathol. 8:4684–4694.
2015.PubMed/NCBI
|
|
26
|
Bornachea O, Santos M, Martínez-Cruz AB,
García-Escudero R, Dueñas M, Costa C, Segrelles C, Lorz C, Buitrago
A, Saiz-Ladera C, et al: EMT and induction of miR-21 mediate
metastasis development in Trp53-deficient tumours. Sci Rep.
2:4342012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han M, Wang Y, Liu M, Bi X, Bao J, Zeng N,
Zhu Z, Mo Z, Wu C and Chen X: MiR-21 regulates
epithelial-mesenchymal transition phenotype and hypoxia-inducible
factor-1alpha expression in third-sphere forming breast cancer stem
cell-like cells. Cancer Sci. 103:1058–1064. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mathé EA, Nguyen GH, Bowman ED, Zhao Y,
Budhu A, Schetter AJ, Braun R, Reimers M, Kumamoto K, Hughes D, et
al: MicroRNA expression in squamous cell carcinoma and
adenocarcinoma of the esophagus: Associations with survival. Clin
Cancer Res. 15:6192–6200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Winther M, Alsner J, Tramm T, Baeksgaard
L, Holtved E and Nordsmark M: Evaluation of miR-21 and miR-375 as
prognostic biomarkers in esophageal cancer. Acta Oncol.
54:1582–1591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lv H, He Z, Wang H, Du T and Pang Z:
Differential expression of miR-21 and miR-75 in esophageal
carcinoma patients and its clinical implication. Am J Transl Res.
8:3288–3298. 2016.PubMed/NCBI
|
|
32
|
Sheikh AM, Small HY, Currie G and Delles
C: Systematic review of Micro-RNA expression in pre-eclampsia
identifies a number of common pathways associated with the disease.
PLoS One. 11:e01608082016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan LX, Liu YH, Xiang JW, Wu QN, Xu LB,
Luo XL, Zhu XL, Liu C, Xu FP, Luo DL, et al: PIK3R1 targeting by
miR-21 suppresses tumor cell migration and invasion by reducing
PI3K/AKT signaling and reversing EMT, and predicts clinical outcome
of breast cancer. Int J Oncol. 48:471–484. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cufi S, Bonavia R, Vazquez-Martin A,
Oliveras-Ferraros C, Corominas-Faja B, Cuyàs E, Martin-Castillo B,
Barrajón- Catalán E, Visa J, Segura-Carretero A, et al: Silibinin
suppresses EMT-driven erlotinib resistance by reversing the high
miR-21/low miR-200c signature in vivo. Sci Rep.
3:24592013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han M, Liu M, Wang Y, Chen X, Xu J, Sun Y,
Zhao L, Qu H, Fan Y and Wu C: Antagonism of miR-21 reverses
epithelial-mesenchymal transition and cancer stem cell phenotype
through AKT/ERK1/2 inactivation by targeting PTEN. PLoS One.
7:e395202012. View Article : Google Scholar : PubMed/NCBI
|