Introduction
Glioblastomas are malignant tumors with a high
incidence and high mortality rate among all brain tumors with an
absence of precise clinical classification biomarkers and effective
therapeutic methods. Glioblastomas are associated with poor
prognosis and a median patient survival of approximately 15 months
(1,2). Therefore, finding effective target
genes and specific biomarkers for glioma diagnosis and
proliferation block is becoming a ‘hotspot’ of research.
Long non-coding RNAs (lncRNAs) are defined as a
novel class of RNAs which have been a ‘hotspot’ for research
(3,4). Although they lack transcript protein
potential, a growing number of lncRNAs have been reported to play a
vital role in physiological progresses such as imprinting control,
cell differentiation and tumorigenesis (5,6) and
lncRNAs are expressed in embryonic stem cells (7), brain tissues (8) and differentiated neurons (9). More significantly, lncRNAs are
involved in several diseases and some are the key factors for the
evaluation of prognosis (10).
However, currently, little is known concerning the related
mechanisms of lncRNAs in gliomas.
Polycomb group (PcG) proteins were first found in
Drosophila melanogaster and these proteins which implement
transcriptional silencing are divided into two main family
complexes, called polycomb repressive complex 1 (PRC1) and polycomb
repressive complex 2 (PRC2) (11),
taking crucial part in embryonic stem cell stage, determination of
cell fate and tumorigenesis (12).
Nervous system polycomb 1 (NSPc1), a key member of mouse PRC1,
shares high homology with PcG protein Bmi-1. Human NSPc1 gene
encodes a 29-kDa nuclear localized protein containing an N-terminal
RING finger domain. Previous research pointed out that NSPc1 is
highly expressed in the early developing nervous system and is
involved in the differentiation of neural crest cells (13,14).
The expression of NSPc1 is significantly altered in different grade
malignant gliomas and is related with the maintenance of the
stemness of cancer stem cells (15). NSPc1 downregulates the
cyclin-dependent kinase (CDK) inhibitor p21Waf1/Cip1 via
the retinoid acid response element (RARE element) resulting in
promotion of tumor cell proliferation and cell cycle transition
(16). In addition, NSPc1 directed
by EZH2, a member of PRC2, mediated histone 2A (H2A) ubiquitination
and DNA methylation (17), which
means that they are interdependent in specific gene silencing in
cancer cells.
Markedly, most PcGs lack the capability of combining
proteins due to their own structure. Thus, they must target
corresponding genomic loci by some cofactors. As reported, lncRNAs
recruit several PRC2 members to specific genes (18), thus, regulating relevant signaling
pathways (19). For instance,
metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is
among the earliest identified lncRNAs and is also named as
nuclear-enriched abundant transcript 2 (NEAT2) (20). MALAT1 has been considered to be
correlated with growth and metastasis of various types of cancer
cells. In renal cancer, MALAT1 was found to regulate downstream
efforts via EZH2-promoting methylation of histone H3 lysine 27
(H3K27), leading to cancer progression and invasion (21). Additionally, MALAT1 was found to
recruit EZH2 to suppress the tumor suppressor PCDH10 contributing
to gastric cancer proliferation and metastasis (22). Nevertheless, the relationship
between PRC1 members and lncRNAs has rarely been reported to date.
Therefore, the main focus of our research was the relationship
between PRC1 member NSPc1 and lncRNAs.
In the present study, we screened possible lncRNAs
which may bind to PRC1 member NSPc1 among numerous candidates and
examined potential correlations between NSPc1 and MALAT1/SOX2OT,
indicating a cross-talk and functional interaction between the
NSPc1 protein complex and MALAT1/SOX2OT in glioma cells. These
findings may provide novel biomarker complexes to the clinical
treatment of glioma.
Materials and methods
Bioinformatic analysis
The scores of NSPc1-lncRNA interactions were
evaluated by bioinformatic analysis through the bioinformatics tool
lncPro (online server: http://bioinfo.bjmu.edu.cn/lncpro) (23). We selected the lncRNAs closely
related in carcinogenesis or multipotency of glioma. Complexes were
downloaded from the Protein DataBank (PDB) database (http://www.pdb.org) by encoding RNA and protein
sequences into numeric vectors. Then the scores for each
lncRNA-NSPc1 pair was used to measure the interactions between
them.
Cell cultures
Glioma H4 cells were purchased from the Cell Culture
Center of the Chinese Academy of Medical Science (Beijing, China).
HK-2 cells were obtained from the American Type Cell Culture
Collection (ATCC; Manassas, VA, USA; http://www.atcc.org/). The cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10%
heat-inactivated fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in 5% CO2.
RNA binding protein
immunoprecipitation (RIP)
RIP was performed to characterize the NSPc1 protein
complex-associated RNAs following the manufacturer's instructions
(EMD Millipore, Billerica, MA, USA) with some modifications.
Briefly, typically one nuclear RIP reaction using one antibody
required 50 µl of chromatin from~1.0×106 cells. The
optimization for shearing cross-linked RNA condition was 24 sec on,
30 sec off for 14 cycles at 4°C. lncRNA-NSPc1 immunoprecipitation
was performed with anti-NSPc1 monoclonal antibody (m-NSPc1) and
anti-NSPc1 polyclonal antibody (poly-NSPc1) both obtained from our
laboratory, anti-EZH2 antibody (cat. no. 07-689; EMD Millipore) as
non-anti-NSPc1 control and normal mouse IgG (cat. no. 17-371; EMD
Millipore) as RIP efficiency control. m-NSPc1 (2 µg), poly-NSPc1 (2
µg), anti-EZH2 (1 µg) and normal mouse IgG (1 µg) were added to
each reaction. The co-precipitated lncRNAs were detected by
qRT-PCR. Total RNAs (input controls), non-anti-NSPc1 control were
assayed to demonstrate endogenous expression of 8 lncRNAs,
efficiency of the reaction and specificity of RNA binding to
NSPc1.
Isolation of RNA and real-time
quantitative reverse transcription polymerase chain reaction
(qRT-PCR)
Total RNA isolated from cell cultures were routinely
used for qRT-PCR. Total RNA was prepared by TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). TRIzol extractions
were conducted according to the manufacturer's protocol
(Invitrogen; Thermo Fisher Scientific, Inc.). For qRT-PCR, the
cDNAs were synthesized using PrimeScript™ RT Master Mix (DRR820A;
Takara Bio Inc., Shiga, Japan) with total RNA as templates. All the
primers of interest were designed and verified through PubMed (data
not shown). qRT-PCR was performed using an StepOnePlus™ System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and SYBR
Premix Ex Taq™ II (DRR820A; Takara Bio Inc.) according to the
manufacturer's instructions. The amplification conditions were 95°C
for 30 sec, as well as 40 cycles at 95°C for 5 sec and 60°C for 60
sec. The fold relative enrichment was quantified together with
normalization by the largest CT level for RIP, and by the GAPDH
level for transfection. Results were analyzed by 2−∆∆Cq
method (24). All qRT-PCR analyses
were performed in triplicate.
Cell transfection
H4 cells were transfected using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). The plasmid
pCDEF-NSPc1 and small interfering RNAs (siRNAs) targeting NSPc1
were constructed by our team as in a previous study (15). One pair of siRNAs against MALAT1 and
SOX2OT and the scramble were constructed by Shanghai GenePharma
Co., Ltd. (Shanghai, China) as siRNA1-MALAT1 (sense,
GAGGUGUAAAGGGAUUU AUTT and antisense, AUAAAUCCCUUUACACCUCTT);
siRNA1-SOX2OT (sense, GGAGUCCAGUCAACUUCAUTT and antisense,
AUGAAGUUGACUGGACUCCTT) and negative control (sense,
UUCUCCGAACGUGUCACGUTT and antisense, ACGUGACACGUUCGGAGAATT). The
other pairs for each were from articles reported and chemically
synthesized by Shanghai GenePharma Co., Ltd., as siRNA2-MALAT1
(sense, GGGCUUCUCUUAACAUUU AUU and antisense,
UAAAUGUUAAGAGAAGCCCUU) and siRNA2-SOX2OT (sense, GGAGAUUGUGACCUGGCU
UTT and antisense, AAGCCAGGUCACAAUCUCCTT).
Western blot analysis
H4 cells were transfected separately with
siRNA-MALAT1, siRNA-SOX2OT and the negative control. The cells were
collected using RIPA Lysis Buffer (cat. no. P0013B; Beyotime
Institute of Biotechnology, Nanjing, China) for western blot
analysis after 72 h of transfection. Protein concentrations were
determined using a BCA Protein Assay reagents (cat. no. P0009;
Beyotime Institute of Biotechnology), equal amounts of protein (20
µg) were loaded per lane and resolved on 10% SDS-PAGE gels, and
then transferred to polyvinylidene difluoride (PVDF) membranes (ΕΜD
Millipore). The membranes were blocked with 5% skimmed milk in 1%
Tween-20 in Tris-based saline (TBST) and then incubated with
primary antibodies (NSPc1, diluted at 1:5,000 was laboratory
prepared and β-actin diluted at 1:6,000 was from Bioworld, Inc.,
St. Louis Park, MN, USA), respectively at 4°C overnight. After
washing with TBST, the membranes were incubated with HRP-conjugated
secondary antibody diluted at 1:6,000 (cat. no. A0216; Bomeike,
Tianjin, China) for 1 h at room temperature and the immuno-reactive
bands were visualized using ECL Western Blot Detection reagents
(EMD Millipore), normalized to β-actin and quantified by ImageJ
system (version 1.47v; NIH; National Institutes of Health,
Bethesda, MD, USA).
Cell proliferation and apoptosis
Briefly, the MTT assay (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was applied to evaluate the cell growth state.
H4 cells were transfected with negative control, siRNA-NSPc1,
siRNA-MALAT1 and siRNA-SOX2OT and co-transfected with siRNA-NSPc1
and siRNA-MALAT1, siRNA-NSPc1 and siRNA-SOX2OT in a 96-well plate.
The MTT assay was carried out following the manufacturer's
protocol. Absorbance values (OD) were determined with an
enzyme-linked immunosorbent detector after 24, 48, 72 and 96 h of
transfection, respectively.
Apoptosis of H4 cells was determined by dual
staining with FITC-Annexin V (BD Biosciences, Franklin Lakes, NJ,
USA) and propidium iodide (PI; Sigma-Aldrich; Merck KGaA). H4 cells
cultured in a 6-well plate were transfected as mentioned in the MTT
assay. According to the manufacturer's protocol, after a 24-h
transfection, H4 cells were analyzed with a flow cytometry system
(BD FACSCalibur™ flow cytometer; BD Biosciences) equipped with
CellQuest software (BD Biosciences).
Statistical analysis
Results are expressed as the mean ± SEM. Statistical
analyses were performed using SPSS software (version 13.0; SPSS,
Inc., Chicago, IL, USA). The significance of the differences
between various groups was analyzed using one-way analysis of
variance (ANOVA). Multiple comparison between the groups was
performed using Student-Newman-Keuls (SNK) method. P<0.05 was
considered to indicate a statistically significant result.
Results
The prediction of in vivo NSPc1-lncRNA
interactions by bioinformatic analysis
To determine the possible lncRNAs that bind to NSPc1
by bioinformatics, the top 7 high scoring lncRNAs were chosen as
candidates to be investigated (Table
I). Moreover, ADAMTS9-AS2 (25)
reported currently to be highly expressed in gliomas was also
selected.
 | Table I.The interaction scores evaluated by
bioinformatic analysis. |
Table I.
The interaction scores evaluated by
bioinformatic analysis.
| Protein | lncRNA | Interaction
score |
|---|
| NSPc1/PCGF1 | SOX2OT | 82. 3497 |
| gi⎪13436326⎪ | MALAT1 | 76. 9802 |
| gb⎪AAH04952.
1⎪ | APTR | 75. 095 |
|
| ANRIL | 71. 9274 |
|
| MEG3 | 68. 4526 |
|
| HOTAIR | 61. 5625 |
|
| H19 | 58. 8719 |
|
| HCG4 | 55. 2284 |
|
| FAL1 | 44. 1196 |
|
| CRNDE | 28. 8089 |
Differential expression of endogenous
lncRNAs and NSPc1 between glioma and non-cancerous cells
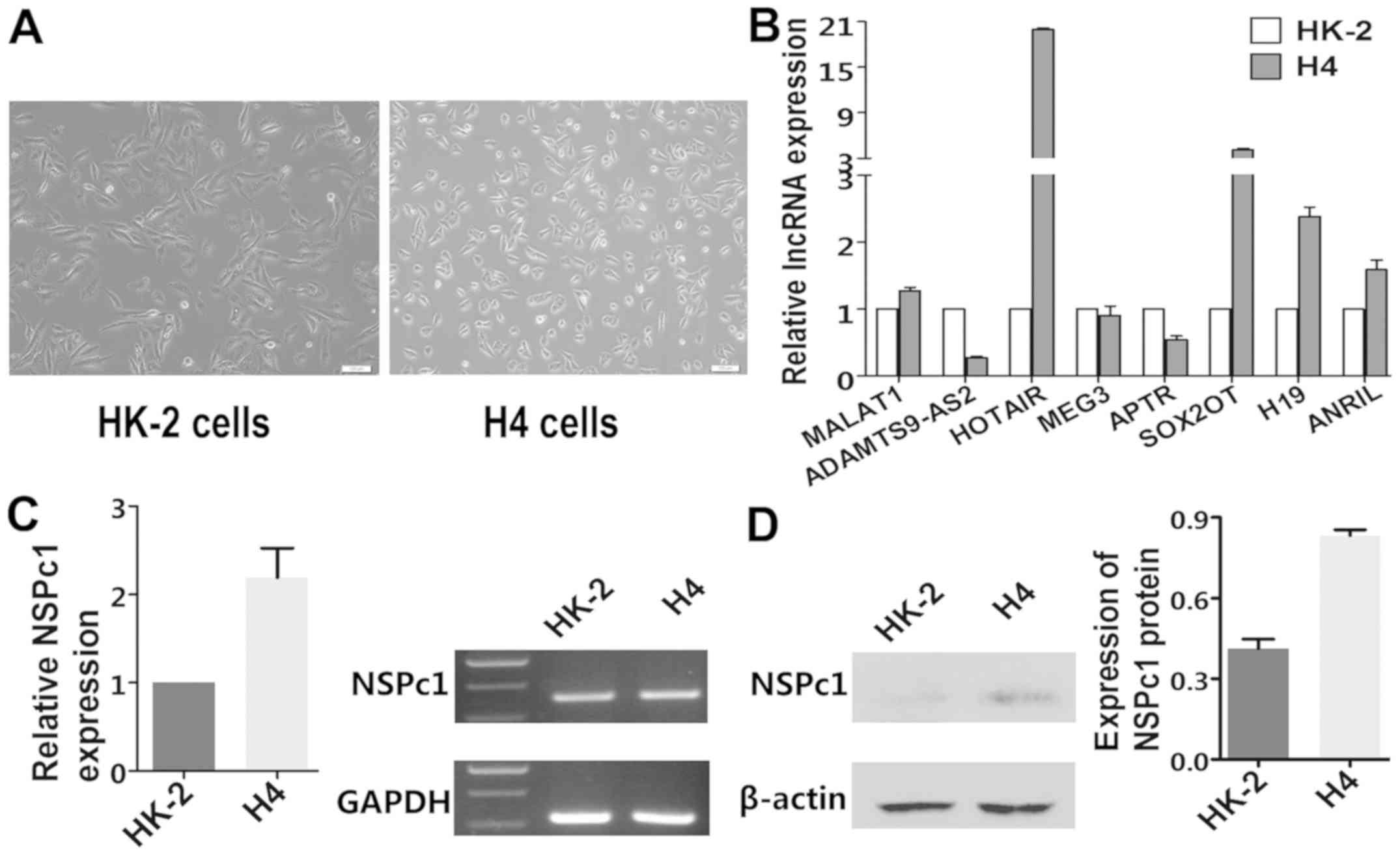
Images of normal cell morphology of HK-2 cells and
H4 cells in the same culture condition are shown in Fig. 1A. Expression levels of MALAT1,
HOTAIR, SOX2OT, H19, ANRIL and NSPc1 were higher in the H4 cells
than levels in the HK-2 cells (Fig.
1B-D), while expression levels of ADASTM9-AS2 and APTR were
lower in the H4 cells compared to that in the HK-2 cells (Fig. 1B).
Identification of lncRNAs which bind
to the NSPc1 protein complex in H4 cells
To investigate the potential interaction between
endogenous NSPc1 protein complex and candidate lncRNAs, RIP-NSPc1
assays were applied in H4 cells. SOX2OT and ADAMTS9-AS2 in the
anti-NSPc1 monoclonal antibody (m-NSPc1) group and MEG3 in the
anti-NSPc1 polyclonal antibody (poly-NSPc1) group were
preferentially enriched compared with anti-EZH2 (non-anti-NSPc1
control) (Fig. 2A). These results
indicated that SOX2OT, ADAMTS9-AS2 and MEG3 may have an in
vivo interaction with NSPc1. In addition, although the
enrichment of MALAT1 in the m-NSPc1 group was lower than that in
the anti-EZH2 control, in poly-NSPc1 group was higher than
anti-EZH2 (Fig. 2A), which also
indicated that MALAT1 has the possibility to bind to NSPc1.
Furthermore, the enrichment of MEG3 and SOX2OT was higher in both
the poly-NSPc1 and m-NSPc1 groups. Remarkably, the endogenous
expression of MALAT1 was prominently high in 10% input (Fig. 2B).
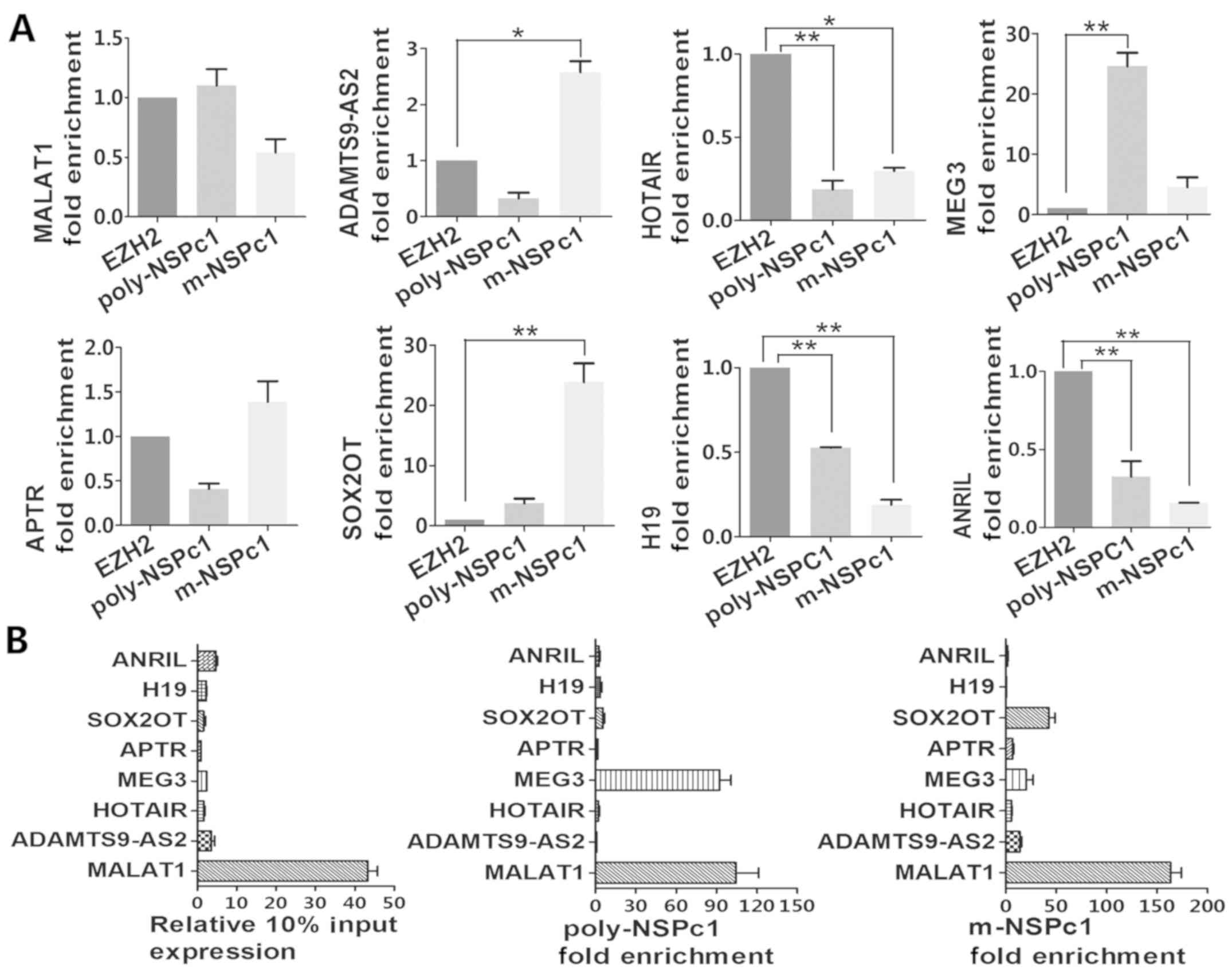 | Figure 2.RIP approach for lncRNA-NSPc1 binding
profile analysis. (A) Purified RNA immunoprecipitated by the
antibodies was then analyzed by qRT-PCR. Verification of RIP
enrichment was performed applying the comparative Ct (∆∆Cq) method
with m-NSPc1 binding to lncRNA compared to positive control binding
lncRNA and poly-NSPc1 binding to lncRNA compared to positive
control binding lncRNA. (B) qRT-PCR was performed with eight pairs
of primers targeting 10% input total lncRNA, m-NSPc1 combining
lncRNA and poly-NSPc1 combining lncRNA. Then, the relative
expression of the eight candidates or enrichment folds were
calculated by Ct (∆∆Cq) method in each group. All data are
presented as the mean ± SD from three independent experiment.
One-way ANOVA, *P<0.05, **P<0.01. EZH2, anti-EZH2 antibody;
m-NSPc1, anti-NSPc1 monoclonal antibody; poly-NSPc1, anti-NSPc1
polyclonal antibody; RIP, RNA binding protein immunoprecipitation;
lncRNAs, long non-coding RNAs; NSPc1, nervous system polycomb
1. |
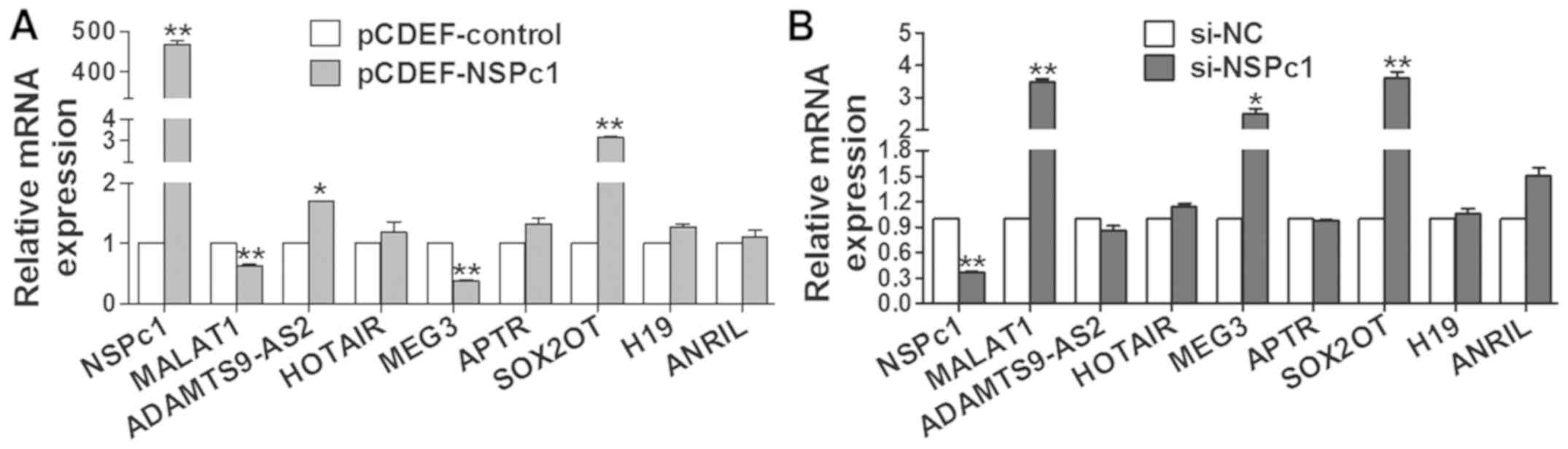
Exogenous expression levels of NSPc1
affect the expression levels of MALAT1, SOX2OT and MEG3
To investigate whether the variability of NSPc1
expression influenced the expression of those 8 lncRNAs, the
overexpression vector pCDEF-NSPc1 and siRNA-NSPc1 were used. We
found that the expression of MALAT1 and MEG3 (Fig. 3A) was decreased by elevated
exogenous expression of NSPc1 in H4 cells, and vice versa (Fig. 3B). Expression of SOX2OT was always
significantly increased no matter how the expression of NSPc1 was
altered while the other 5 lncRNAs exhibited no significant changes
(Fig. 3).
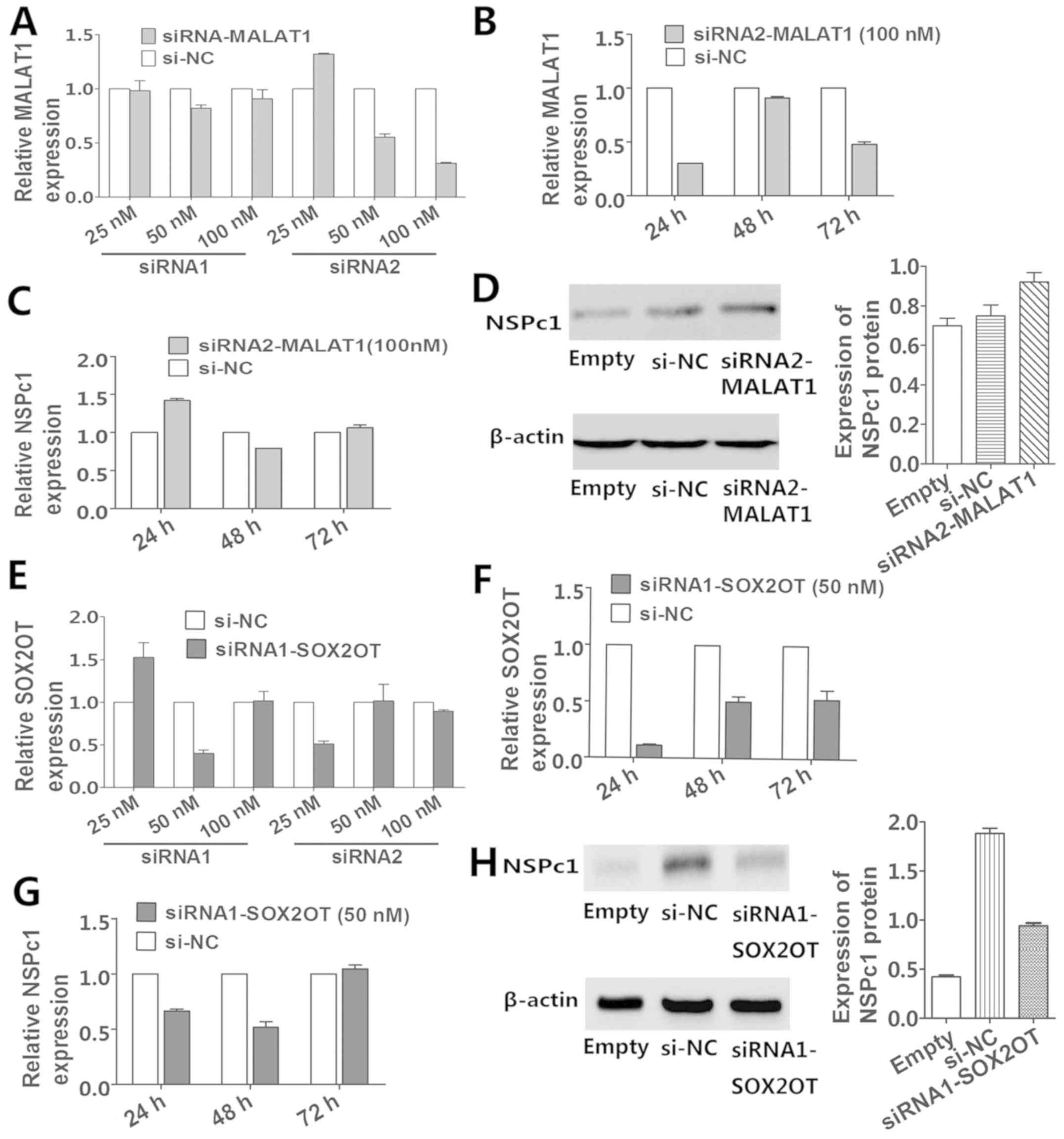
Knockdown of MALAT1 and SOX2OT affects
the expression of NSPc1
MALAT1 and SOX2OT were chosen for subsequent
research as the P-value of their expression alterations with NSPc1
variation were much less than the other candidates. The optimum
transfection efficiency of the silencing of siRNA2-MALAT1 (Fig. 4A and B) and siRNA1-SOX2OT (Fig. 4E and F) were optimum at
downregulating their target genes 24 h post-DNA addition (Fig. 4B and F). Above all, the RNA level of
NSPc1 increased after 24 and 72 h of transfection and the protein
level of NSPc1 increased after 72 h of transfection in response to
the decrease in MALAT1 (Fig. 4C and
D), whereas both the RNA and protein levels of NSPc1 were
increased following the decrease in SOX2OT (Fig. 4G and H) after 72 h of
transfection.
Decrease in the co-expression of NSPc1
and MALAT1/SOX2OT inhibits H4 cell growth and promotes apoptosis
more significantly than the other single transfections in
vitro
The proliferation and apoptosis of H4 cells after
transfection are shown in Fig. 5.
MTT assay showed that the knockdown of expression of NSPc1, MALAT1,
SOX2OT and knockdown of the co-expression of NSPc1 and MALAT1,
NSPc1 and SOX2OT decreased the cell proliferation compared with the
negative control. Above all, the silenced co-expression of NSPc1
and MALAT1, NSPc1 and SOX2OT blocked the growth of H4 cells more
significantly than the single downregulation of the expression of
NSPc1, MALAT1 and SOX2OT (Fig. 5A and
B). Furthermore, the cell apoptotic rate following the
simultaneous knockdown of the expression of NSPc1 and SOX2OT
(65.05%) was higher than the sum of those following single
downregulation of NSPc1 and SOX2OT (38.23±22.55%). Whereas the cell
apoptotic rate following the decrease in the co-expression of NSPc1
and MALAT1 (49.17%) was not higher than the sum of those following
the respectively descending expression of NSPc1 and MALAT1
(38.23±16.84%). In addition, flow cytometric analysis indicated
that the decreased expression of NSPc1, MALAT1, SOX2OT and
co-expression of NSPc1 and MALAT1, NSPc1 and SOX2OT induced more
cell apoptosis than the negative control. The coordinate knockdown
of NSPc1 and MALAT1, NSPc1 and SOX2OT with siRNAs promoted more H4
cell death compared with the other single transfections (Fig. 5C-H).
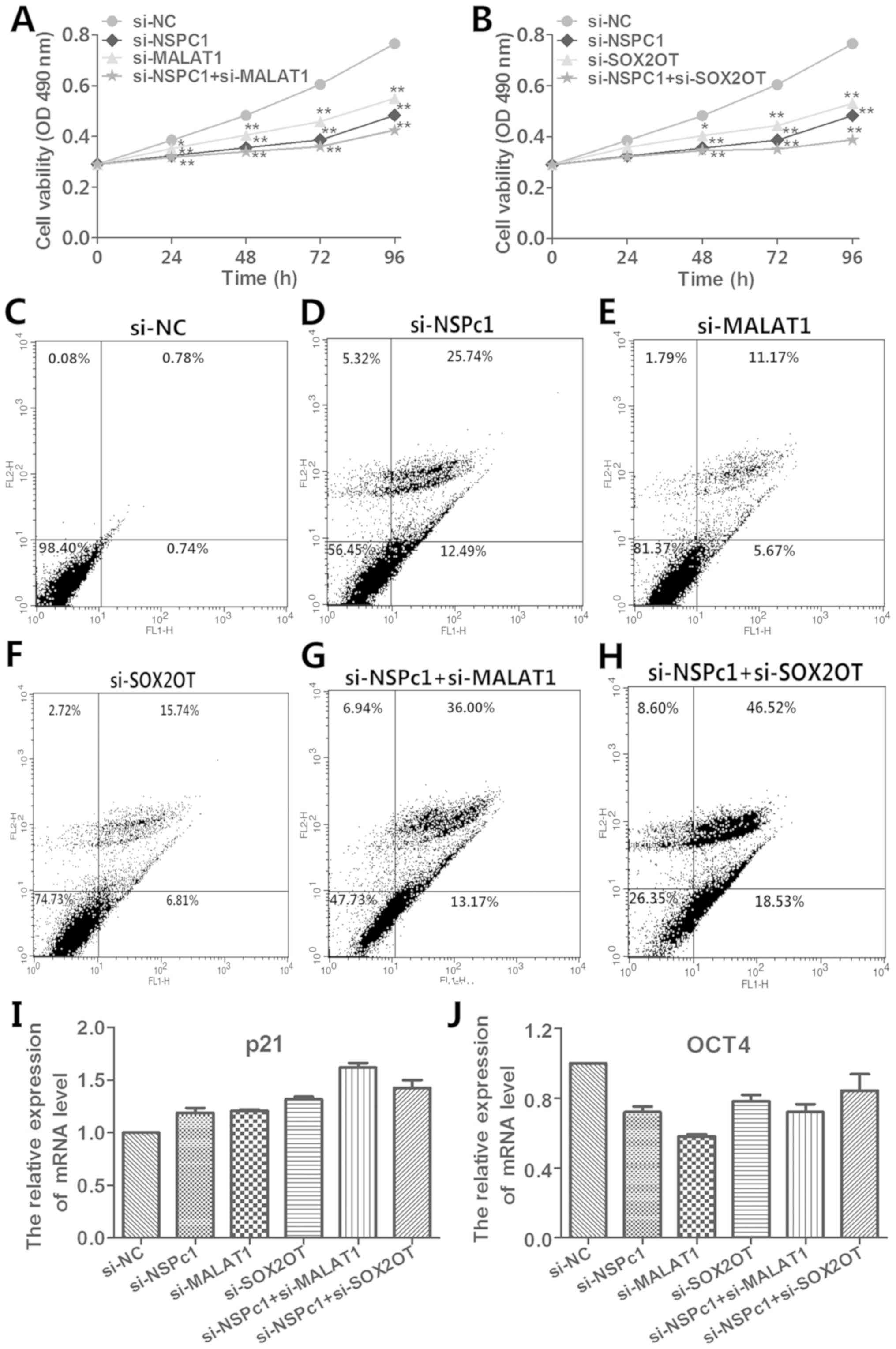 | Figure 5.Effects of the knockdown of NSPc1,
MALAT1, SOX2OT, NSPc1 plus MALAT1, NSPc1 plus SOX2OT on H4 cell
proliferation and apoptosis in vivo and the effects of the
knockdown of the expression of NSPc1, MALAT1, SOX2OT and
co-suppression of NSPc1 and MALAT1, NSPc1 and SOX2OT on the
expression of p21 and OCT4. (A and B) MTT assay was applied to
determine H4 cell viability in the si-NC, si-NSPc1, si-MALAT1,
si-SOX2OT, si-NSPc1+si-MALAT1 and si-NSPc1+si-SOX2OT transfected
cells. One-way ANOVA, *P<0.05, **P<0. 01. (C-H)
Representative flow cytometric analysis were performed to evaluate
the H4 cell apoptosis 24 h following transfection. (I and J) The
knockdown transfection was used to explore the influence on the
expression levels of p21 and OCT4. MALAT1, metastasis-associated
lung adenocarcinoma transcript 1; NSPc1, nervous system polycomb
1. |
Decreased expression of NSPc1, MALAT1,
SOX2OT and co-expression of NSPc1 and MALAT1, NSPc1 and SOX2OT
affect the expression of p21 and OCT4
As shown in Fig. 5I and
J, the expression of p21 was increased and the expression of
OCT4 was decreased following the decreased expression of NSPc1,
MALAT1, SOX2OT, NSPc1+MALAT1 and NSPc1+SOX2OT (P<0.01).
Moreover, the decrease in co-expression of NSPc1 and MALAT1 caused
the expression of p21 to be more highly and significantly increased
when compared to the other knockdown groups (Fig. 5I). Also, silencing of the expression
of MALAT1 significantly decreased the expression of OCT4 to a
greater degree than the other knockdown groups (Fig. 5J).
Discussion
Accumulating evidence indicates that long non-coding
RNAs (lncRNAs) are engaged in different pathologic processes
including gliomagenesis, and the polycomb group (PcG) members play
a vital role in cellular life span and stimulate the growth and
promotion of many cancer types (26). Links between PcG complexes and
lncRNAs have long been proposed. Several lncRNAs which have
probable interaction with PcG members have been suggested. CBX7 of
polycomb repressive complex 1 (PRC1) (27) and SUZ1 of polycomb repressive
complex 2 (PRC2) (28) associate
with lncRNA ANRIL to repress the tumor suppressor INK4b/ARF/INK4a
locus and control senescence in cancer tissues. lncRNA FAL1
combines with PRC1 core protein, BMI1 to regulate the transcription
of various genes such as CDKN1A (29). HOTAIR recruits EZH2, SU12 and EED,
which is a PRC2 member, to enhance methylation of histone H3 lysine
27 (H3K27) and accelerated metastasis of breast cancer by silencing
metastasis-suppressing genes (30).
We used the latest RNA binding protein immunoprecipitation (RIP)
technique to identify the most possible lncRNAs which may interplay
with nervous system polycomb 1 (NSPc1) protein complex. Among eight
candidates, we calculated a high score by bioinformatics. Yet, one
limitation included the lack of direct binding data from the RNA
pull-down assay which should further be researched in future
studies. In summary, we identified the probable interactions with
NSPc1 protein complex in the aspect of expression and function in
H4 cells.
In addition to metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1)-EZH2 and NSPc1-EZH2
interaction mentioned above (16,19,20),
NSPc1 recruitment is the downstream event of EZH2-driven H3K27
methylation during tumorigenesis (16) and MALAT1 regulates downstream
factors via EZH2-mediated H3K27 in renal cancer tissue (19). This research suggested that
MALAT1-EZH2 interaction and NSPc1-EZH2 combination, which indicates
a novel correlation of MALAT1-NSPc1. As expected, among the 8
candidates, our RNA immunoprecipitation results indicated the
possible binding relationship between MALAT1 and NSPc1 protein
complex in H4 cells. Next, we identified that the expression of
MALAT1 as measured by qRT-PCR was contrary to the variation of
NSPc1 expression regulated by pCDEF-NSPc1 and siRNA-NSPc1
separately in H4 cells. Furthermore, the decreased expression of
MALAT1 increased gene and protein levels of NSPc1 in H4 cells.
Thus, MALAT1 is negatively regulated by NSPc1 in H4 cells. However,
more research should be conducted to explore the regulatory
mechanism of the NSPc1/PRC1/MALAT1 complex model in the future.
The overexpression of MALAT1 has been shown to act
as a tumor-promotor factor in colorectal (31), lung (32) and gastric cancer (33). The expression of NSPc1 is
significantly increased in high grade malignant gliomas (15). We identified that expression levels
of MALAT1 and NSPc1 were higher in glioma cells than levels in
non-cancerous cells by qRT-PCR. Furthermore, it was observed that
inhibition of the respective expression levels of NSPc1 and MALAT1
or the suppression of the co-expression of these was able to retard
H4 cell growth and promote their apoptosis. Noticeably, the
decreased co-expression of NSPc1 and MALAT1 had a more significant
impact on survival and apoptosis than separately silenced
expression. However, the cell apoptosis rate caused by the
downregulation of the co-expression of NSPc1 and MALAT1 was not
higher than the total cell apoptosis caused by a separate decrease
in expression of MALAT1 and NSPc1. We speculated that the primary
cause of this was that in the situation of the co-transfection of
NSPc1 and MALAT1, upon the silencing of the expression of MALAT1,
the endogenous expression of NSPc1 increased due to the negative
regulation of MALAT1. At the same time, the expression of NSPc1 was
knocked down by siRNA based on the basic condition of
MALAT1-mediated increase of the endogenous expression of NSPc1. As
a result, the downregulation efficiency of NSPc1 in the single
NSPc1 transfection group was much higher than that in the
co-transfection group, leading to the phenomenon that the decreased
co-expression effect on promoting cell apoptosis was partly
neutralized by the negative cross-talk between them. In summary,
these data demonstrated that the NSPc1/PRC1/MALAT1 functional
interaction may regulate glioma cell proliferation and apoptosis to
a certain extent, which may play important roles as biomarkers for
glioma H4 cells. Moreover, it is inappropriate to apply
co-downregulation of NSPc1 and MALAT1 strategy to the gene therapy
in glioma H4 cells. However, the conclusion, to some degree, lacks
generalized applicability for the target therapy for all types of
glioma cells because the only one type of glioma cells was used.
Therefore, additional glioma cell lines and animal models will be
utilized for further research in order to confirm the biomarkers
for brain tumors. Furthermore, the definite molecular mechanisms
need further investigation.
According to a previous study, Nspc1 is highly
expressed in differentiated pluripotency P19 cells and maintains
the pluripotency of P19 cells (34). Furthermore, Nspc1 activates the key
pluripotent Oct4/Nanog/Sox2 axis in P19 cells and Oct4, Nanog and
Sox2 are positively regulated by Nspc1 in P19 cells. SOX2, a master
regulator of pluripotency, is embedded within the third intron of
an lncRNA known as SOX2 overlapping transcript (SOX2OT) (35). SOX2OT has been suggested to
participate in the transcriptional regulation of SOX2 (36). However, the presence of any physical
interaction between SOX2OT and SOX2 has been alleged in recent
studies that SOX2OT has a positive effect on SOX2 expression via
knockdown and overexpression assays (37,38).
Also, SOX2OT plays a vital role in regulating and mediating
pluripotency and tumorigenesis events, probably by the expression
of SOX2 (39). Therefore, in the
present study, we confirmed the in vivo cooperation between
SOX2OT and the NSPc1 protein complex by RIP method. Furthermore, we
identified that the expression of SOX2OT was consistently increased
no matter how NSPc1 expression was altered whereas the gene and
protein expression of NSPc1 was decreased by the downregulation of
SOX2OT in H4 cells. Moreover, whether the regulation between NSPc1
and SOX2OT affects transcription of target genes or expression of
each will be further studied as a next research step.
In addition, recent studies reported that patients
with low expression of SOX2OT had prolonged survival compared with
those with a high level of SOX2OT in hepatocellular carcinoma
(40) and gastric cancer (41). As in this study, the expression of
SOX2OT was much higher in glioma cells than that noted in
non-cancerous cells. We also studied the interaction between the
NSPc1 protein complex and SOX2OT at the functional level. Our
results showed that downregulation of NSPc1 and SOX2OT
co-expression reduced the cell growth and increased cell death more
obviously compared with that of NSPc1 and SOX2OT alone. Moreover,
the cell apoptosis rate following decreased co-expression of NSPc1
and SOX2OT was higher than the sum of that in the respective
downregulation of the expression of NSPc1 and SOX2OT. The main
reason that we speculated was that in the co-downregulation of
SOX2OT and NSPc1 conditions, the endogenous expression of NSPc1 was
decreased by the downregulation of SOX2OT. Besides this
circumstance, the expression of NSPc1 was knocked down by exogenous
siRNA simultaneously. Hence, the downregulation efficiency of NSPc1
in the co-expression group was higher than that in the single NSPc1
silenced group, resulting in the more significant influence on
accelerating cell apoptosis. It is meaningful that this
collaborative cross-talk between NSPc1 and SOX2OT could act as a
more effective therapeutic complex target for glioma H4 cells.
Nevertheless, the data of our research do not represent universal
therapeutic complex targets for all glioma cell lines. In addition,
the exact biological function of the interaction between the NSPc1
protein complex and SOX2OT has yet remained unknown. Hence, we will
conduct in-depth study on this unsolved issue.
It has been reported that p21 is the direct target
gene regulated by NSPc1 in tumor cell growth (15) and the pluripotency stem cell marker
OCT4 was positively regulated by NSPc1 in P19 cells (33). Thus, we explored the expression
influence of p21 and OCT4 following knockdown of the expression of
NSPc1, MALAT1, SOX2OT, NSPc1+MALAT1 and NSPc1+SOX2OT. We found that
co-knockdown of the expression of NSPc1 and MALAT1 increased the
expression of p21 most obviously in glioma H4 cells. Furthermore,
silencing of the expression of MALAT1 decreased the expression of
OCT4 most significantly in glioma H4 cells. This may provide a new
concept in the influence of the lncRNA-NSPc1 complex on regulating
target cancer marker genes.
In conclusion, our data provide an initial insight
into the potential function of the MALAT1-NSPc1 protein complex and
SOX2OT-NSPc1 protein complex binding and crosstalk relationship in
H4 glioma cells. In addition, it highlights the negative
correlation between MALTA1 and NSPc1 in H4 cells and that NSPc1 is
positively regulated by SOX2OT in H4 cells. However, the altered
expression of NSPc1 promotes SOX2OT expression increase. In
addition, decreased co-expression of NSPc1 and MALAT1, NSPc1 and
SOX2OT suppressed the growth of glioma H4 cells and accelerated
apoptosis more obviously compared with the respective decline in
expression. Furthermore, the co-expression of NSPc1 and SOX2OT
induced a higher cell apoptosis compared with the sum of the
decreased expression levels of NSPc1 and SOX2OT alone, which may
provide a novel complex in gene strategy. Nevertheless, this effect
was partially counteracted in the silenced co-expression of MALAT1
and NSPc1 due to the negative correlation between them.
In brief, these findings may provide potential
therapeutic gene targets and novel biomarkers for malignant glioma.
Moreover, further research, particularly different animal models,
must be conducted to explore the detail mechanism of the functional
complexes of lncRNAs-PcGs and their molecular mechanisms in
glioma.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Sciences Foundation of China (no. 31070929).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YW, ZL and HL performed the experiments. HL, JT, YS
and YG analyzed and interpreted the data. HL and YG conceived and
designed the study. YW and YG were major contributors in writing
the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnson DR and O'Neill BP: Glioblastoma
survival in the United States before and during the temozolomide
era. J Neurooncol. 107:359–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnsson P, Lipovich L, Grandér D and
Morris KV: Evolutionary conservation of long non-coding RNAs;
sequence, structure, function. Biochim Biophys Acta.
1840:1063–1071. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tripathi V, Ellis JD, Shen Z, Song DY, Pan
Q, Watt AT, Freier SM, Bennett CF, Sharma A, Bubulya PA, et al: The
nuclear-retained noncoding RNA MALAT1 regulates alternative
splicing by modulating SR splicing factor phosphorylation. Mol
Cell. 39:925–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wilusz JE: Long noncoding RNAs: Re-writing
dogmas of RNA processing and stability. Biochim Biophys Acta.
1859:128–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dinger ME, Amaral PP, Mercer TR, Pang KC,
Bruce SJ, Gardiner BB, Askarian-Amiri ME, Ru K, Soldà G, Simons C,
et al: Long noncoding RNAs in mouse embryonic stem cell
pluripotency and differentiation. Genome Res. 18:1433–1445. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mercer TR, Dinger ME, Sunkin SM, Mehler MF
and Mattick JS: Specific expression of long noncoding RNAs in the
mouse brain. Proc Natl Acad Sci USA. 105:716–721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mercer TR, Qureshi IA, Gokhan S, Dinger
ME, Li G, Mattick JS and Mehler MF: Long noncoding RNAs in
neuronal-glial fate specification and oligodendrocyte lineage
maturation. BMC Neurosci. 11:142010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang X, Sun S, Pu JK, Tsang AC, Lee D,
Man VO, Lui WM, Wong ST and Leung GK: Long non-coding RNA
expression profiles predict clinical phenotypes in glioma.
Neurobiol Dis. 48:1–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Morey L and Helin K: Polycomb group
protein-mediated repression of transcription. Trends Biochem Sci.
35:323–332. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sparmann A and van Lohuizen M: Polycomb
silencers control cell fate, development and cancer. Nat Rev
Cancer. 6:846–856. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nunes M, Blanc I, Maes J, Fellous M,
Robert B and McElreavey K: NSPc1, a novel mammalian Polycomb gene,
is expressed in neural crest-derived structures of the peripheral
nervous system. Mech Dev. 102:219–222. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong Y, Wang X, Liu J, Shi L, Yin B, Peng
X, Qiang B and Yuan J: NSPc1, a mainly nuclear localized protein of
novel PcG family members, has a transcription repression activity
related to its PKC phosphorylation site at S183. FEBS Lett.
579:115–121. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu PS, Xia QS, Wu F, Li DK, Qi YJ, Hu Y,
Wei ZZ, Li SS, Tian NY, Wei QF, et al: NSPc1 promotes cancer stem
cell self-renewal by repressing the synthesis of all-trans retinoic
acid via targeting RDH16 in malignant glioma. Oncogene.
36:4706–4718. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gong Y, Yue J, Wu X, Wang X, Wen J, Lu L,
Peng X, Qiang B and Yuan J: NSPc1 is a cell growth regulator that
acts as a transcriptional repressor of p21Waf1/Cip1 via the RARE
element. Nucleic Acids Res. 34:6158–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu X, Gong Y, Yue J, Qiang B, Yuan J and
Peng X: Cooperation between EZH2, NSPc1-mediated histone H2A
ubiquitination and Dnmt1 in HOX gene silencing. Nucleic Acids Res.
36:3590–3599. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Simon JA and Kingston RE: Mechanisms of
polycomb gene silencing: Knowns and unknowns. Nat Rev Mol Cell
Biol. 10:697–708. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Davidovich C, Zheng L, Goodrich KJ and
Cech TR: Promiscuous RNA binding by Polycomb repressive complex 2.
Nat Struct Mol Biol. 20:1250–1257. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gutschner T, Hämmerle M and Diederichs S:
MALAT1 - a paradigm for long noncoding RNA function in cancer. J
Mol Med (Berl). 91:791–801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hirata H, Hinoda Y, Shahryari V, Deng G,
Nakajima K, Tabatabai ZL, Ishii N and Dahiya R: Long noncoding RNA
MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and
interacts with miR-205. Cancer Res. 75:1322–1331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi Y, Ooi HS, Wu J, Chen J, Zhang X, Tan
S, Yu Q, Li YY, Kang Y, Li H, et al: MALAT1 long ncRNA promotes
gastric cancer metastasis by suppressing PCDH10. Oncotarget.
7:12693–12703. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu Q, Ren S, Lu M, Zhang Y, Zhu D, Zhang X
and Li T: Computational prediction of associations between long
non-coding RNAs and proteins. BMC Genomics. 14:6512013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Δ Δ C(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yao J, Zhou B, Zhang J, Geng P, Liu K, Zhu
Y and Zhu W: A new tumor suppressor lncRNA ADAMTS9-AS2 is regulated
by DNMT1 and inhibits migration of glioma cells. Tumour Biol.
35:7935–7944. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Laugesen A and Helin K: Chromatin
repressive complexes in stem cells, development, and cancer. Cell
Stem Cell. 14:735–751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yap KL, Li S, Muñoz-Cabello AM, Raguz S,
Zeng L, Mujtaba S, Gil J, Walsh MJ and Zhou MM: Molecular interplay
of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by
polycomb CBX7 in transcriptional silencing of INK4a. Mol Cell.
38:662–674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kotake Y, Nakagawa T, Kitagawa K, Suzuki
S, Liu N, Kitagawa M and Xiong Y: Long non-coding RNA ANRIL is
required for the PRC2 recruitment to and silencing of p15(INK4B)
tumor suppressor gene. Oncogene. 30:1956–1962. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu X, Feng Y, Zhang D, Zhao SD, Hu Z,
Greshock J, Zhang Y, Yang L, Zhong X, Wang LP, et al: A functional
genomic approach identifies FAL1 as an oncogenic long noncoding RNA
that associates with BMI1 and represses p21 expression in cancer.
Cancer Cell. 26:344–357. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ji Q, Liu X, Fu X, Zhang L, Sui H, Zhou L,
Sun J, Cai J, Qin J, Ren J, et al: Resveratrol inhibits invasion
and metastasis of colorectal cancer cells via MALAT1 mediated
Wnt/β-catenin signal pathway. PLoS One. 8:e787002013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shen L, Chen L, Wang Y, Jiang X, Xia H and
Zhuang Z: Long noncoding RNA MALAT1 promotes brain metastasis by
inducing epithelial-mesenchymal transition in lung cancer. J
Neurooncol. 121:101–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang J, Su L, Chen X, Li P, Cai Q, Yu B,
Liu B, Wu W and Zhu Z: MALAT1 promotes cell proliferation in
gastric cancer by recruiting SF2/ASF. Biomed Pharmacother.
68:557–564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li H, Fan R, Sun M, Jiang T and Gong Y:
Nspc1 regulates the key pluripotent Oct4-Nanog-Sox2 axis in P19
embryonal carcinoma cells via directly activating Oct4. Biochem
Biophys Res Commun. 440:527–532. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fantes J, Ragge NK, Lynch SA, McGill NI,
Collin JR, Howard-Peebles PN, Hayward C, Vivian AJ, Williamson K,
van Heyningen V, et al: Mutations in SOX2 cause anophthalmia. Nat
Genet. 33:461–463. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Amaral PP, Neyt C, Wilkins SJ,
Askarian-Amiri ME, Sunkin SM, Perkins AC and Mattick JS: Complex
architecture and regulated expression of the Sox2ot locus during
vertebrate development. RNA. 15:2013–2027. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Askarian-Amiri ME, Seyfoddin V, Smart CE,
Wang J, Kim JE, Hansji H, Baguley BC, Finlay GJ and Leung EY:
Emerging role of long non-coding RNA SOX2OT in SOX2 regulation in
breast cancer. PLoS One. 9:e1021402014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shahryari A, Rafiee MR, Fouani Y, Oliae
NA, Samaei NM, Shafiee M, Semnani S, Vasei M and Mowla SJ: Two
novel splice variants of SOX2OT, SOX2OT-S1, and SOX2OT-S2 are
coupregulated with SOX2 and OCT4 in esophageal squamous cell
carcinoma. Stem Cells. 32:126–134. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shahryari A, Jazi MS, Samaei NM and Mowla
SJ: Long non- coding RNA SOX2OT: Expression signature, splicing
patterns, and emerging roles in pluripotency and tumorigenesis.
Front Genet. 6:1962015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shi XM and Teng F: Up-regulation of long
non-coding RNA Sox2ot promotes hepatocellular carcinoma cell
metastasis and correlates with poor prognosis. Int J Clin Exp
Pathol. 8:4008–4014. 2015.PubMed/NCBI
|
|
41
|
Zhang Y, Yang R, Lian J and Xu H: lncRNA
Sox2ot overexpression serves as a poor prognostic biomarker in
gastric cancer. Am J Transl Res. 8:5035–5043. 2016.PubMed/NCBI
|