Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common cancer in the world and the third leading cause of
cancer-associated deaths in China (1,2). The
pathogenesis of HCC is closely associated with chronic
inflammation, including inflammation initiated by contamination
with bacteria, chemical materials and oxidation or metabolic
stress. The liver is the organ where foreign antigens from the
gastrointestinal tract encounter the immune system (3). The immune defense of the liver primarily
depends on natural immunity.
The liver develops a variety of mechanisms against
liver cancer gene mutations which result from foreign antigens
(4). Toll-like receptors (TLRs) are
important pattern recognition receptors required to maintain the
balance of inflammatory and fibrogenic signaling in the liver.
Stimulation of TLRs to activate the innate immune system has been a
valid therapeutic strategy for treating patients with HCC for a
number of years (5). TLR2 is one of
the most well-studied TLRs and recognizes a variety of pathogens
and damage-associated molecules, including myeloid differentiation
factor 88 (MyD88) and the apoptosis signal regulating kinase 1
(ASK1)/p38 mitogen-activated protein kinase (p38 MAPK)/nuclear
factor κB (NF-κB) signaling pathway (6). Systemic targeting of TLR2 was
demonstrated to be an efficacious option for treating HCC in mice
(7). TLR2-deficient mice present a
significantly increased rate of HCC carcinogenesis and progression.
Animal data have shown that a new class of modified TLR2 agonists
which were used for the pretreatment of liver sinusoidal
endothelial cells (LSECs) can be administered systemically as a
potential therapeutic intervention for improving liver cell
immunity (8).
A number of studies have demonstrated that aberrant
overexpression of TLR2 is present in liver samples of patients with
different stages of liver disease (9), that TLR2 affects the bioactivity of
HepG2 and BEL-7402 cell lines (10),
and that Tlr2-null mice exhibit increased aggressive HCC
development (11).
However, the mechanism through which TLR2 activation
underlies tumor proliferation remains unknown. Confusingly, it has
been reported that the role of TLR2 differs significantly between
tumor tissues of the liver and peritumoral tissues (9). TLR2 in normal tissues directly attacks
tumor cells, and additionally performs other indirect guard-like
functions such as promoting the secretion of a number of cytokines.
In contrast, TLR2 expression in the tumor site is considerably
lower, and thus it may be used as an indicator for immune escape of
tumor cells (11).
In the present study, TLR2 expression was found to
be increased in liver carcinoma tissues at both the gene and
protein levels, and the expression and downstream factors of TLR2
signaling were determined in B76/Huh7 cells cultured in the serum
of HCC patients after stimulation of Pam3CSK4, a potent activator
of the proinflammatory transcription factor NF-κB1 in vitro.
The B76/Huh7 cells were treated with a variety of activating or
inhibitory stimuli. The GFP-expressing plasmid pcDH-TLR2-GFP-PMV
and pcDNA-MYD88-GFP were transfected into B76/Huh7 cells to
determine the effects on HCC progression. The downstream endogenous
cytokines interleukin (IL)-6, IL-8, tumor necrosis factor-α
(TNF-α), and interferon-β (IFN-β), which ultimately converge on the
NF-κB signaling pathway, were auxiliary factors. Overexpression of
either TLR2 or MyD88 with Pam3CSK4 stimulation in B76/Huh7 cells
decreased the growth of cells by inhibiting the activation of the
downstream pathway. Transfection with siRNAs targeting TLR2 or
MyD88 significantly increased the downstream signaling cascade,
which exacerbated the progression of hepatocarcinogenesis.
Expression of the downstream cytokines IL-6, IL-8, TNF-α and IFN-β
was altered correspondingly. The results of the present study
demonstrate a novel TLR2 signaling pathway involved in
proliferation in HCC. Additional research is required to further
illustrate the function of TLR2 in HCC.
Materials and methods
Patients and specimens
To investigate the relationship between HCC and the
signaling pathway molecules of TLR2, a total of 20 paired fresh
liver carcinoma tissues and pericarcinoma tissues were collected
from patients undergoing curative resection without consideration
of recurrence and/or metastasis at the Mengchao Hepatobiliary
Hospital of Fujian Medical University (Fujian, China) between June
2010 and July 2016. Clinical and pathological diagnosis of HCC met
the criteria of The American Association for the Study of Liver
Diseases (https://www.aasld.org). Tumor stage was
determined according to Barcelona Clinic Liver Cancer (BCLC)
staging classification (12). Primary
HCC and matched peritumoral tissues were defined by pathological
sampling specification; all tumor tissues excised by hepatectomy
were trimmed by an experienced pathologist to remove as much normal
liver tissue as possible. Generally the pericarcinoma tissues were
located 2 cm beyond the cancer tissues. The final obtained tumor
tissues had at least 80% HCC cells as confirmed by H&E
staining. Subsequently, the specimens were dissected and then
frozen in liquid nitrogen for DNA extraction and subsequent
experiments. Serum from patients with HCC was additionally
preserved prior to undergoing curative resection. Additionally, the
serum of normal healthy individuals was obtained from donors. The
serum samples were centrifuged for 5 min at 300 × g at 4°C and were
frozen at −80°C. The study was approved by the Ethics Committee of
the Mengchao Hepatobiliary Hospital of Fujian Medical University
(study license no. 2016-007-01). Informed consent was obtained from
patients and health recruits.
Cell lines and cell culture
The human hepatocellular carcinoma cell line,
B76/Huh7, was purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). The B76/Huh7 cells were cultured in DMEM
medium (Hyclone; Thermo, Fisher Scientific, Inc., Waltham, MA, USA)
with 10% fetal bovine serum (Gibco; Thermo, Fisher Scientific,
Inc.) in a humidified incubator with 5% CO2 at 37°C.
Flow cytometry (FCM) detection of TLR2
expression
To determine the association between HCC and TLR2 as
well as TLR2 downstream factors, the HCC tumor and peritumoral
tissues were dissociated and 2×106 cells were analyzed
by FACS Verse (FCM; BD Biosciences, San Jose, CA, USA). FlowJo
7.6.1(BD Biosciences, San Jose, CA, USA) was used for flow
cytometric analysis (Engine 2.79000OS version: Windows 7 Java
Version: 14.1-b02). Suspensions of 2×106 B76/Huh7 cells
cultured in DMEM, in serum from patients with HCC, in serum from
healthy individuals or in 10% FBS were also used to assess TLR2
protein expression by FCM. The cells were collected and washed
twice with PBS, stained with 5 µl anti-human TLR2-phycoerythrin
(PE) antibody (Product #12-9922-41; 1:1,000; eBioscience™; Thermo
Fisher Scientific, Inc.) at 4°C for 30 min away from light. After
washing twice with PBS, the cells were collected by low speed
centrifugation (1,000 × g; 5 min) at 4°C, suspended in 500 µl PBS
and analyzed by FCM. As a negative control an immunoglobulin G2a-PE
antibody was used.
Pam3CSK4 activation and plasmid
transfection
The B76/Huh7 cells were seeded at 2×105
cells/well into 6-well dishes and cultured overnight until they
reached 70–80% confluence. The cells were treated with 1 µg/µl
Pam3CSK4 (Invitrogen; Thermo Fisher Scientific, Inc.) for 72 h to
stimulate the cells. Total cellular protein was extracted for
western blotting 24 h after stimulation. Another set of B76/Huh7
cells were transfected with pcDH-TLR2-GFP-PMV which was designed
and constructed by United Innovation of Mengchao Hepatobiliary
Technology Key Laboratory of Fujian Province. To create this
plasmid, human TLR2 cDNA was PCR amplified and ligated into a
Xho1 and BamH digested pcDH-CMV-EF-PURD-GFP vector,
and insertion was confirmed by a sequencing company. Additionally,
another set of cells was transfected with a GFP-expressing
pcDNA-MyD88 plasmid which was provided by another laboratory. The
transfected B76/Huh7 cells were seeded at 2×105
cells/well into 6-well dishes and cultured overnight until they
reached 70–80% confluence. Lipofectamine™ 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) was used for transfection of the
B76/Huh7 cells according to the manufacturer's protocol. A plasmid
DNA (pcDH-TLR2-GFP-PMV, pcDNA-MyD88-GFP or scrambled plasmid) was
added to reach a total DNA of 2 µg per transfection. The
transfected cells were split equally into two 6-well dishes, and
cells were incubated for 48 h. In the second 6-well dishes, 24 h
after transfection, the cells were treated with Pam3CSK4 for 5 h in
3 wells each. Green fluorescence was emitted after successful
transfection. Images were observed and captured using a
fluorescence microscope (Axiocam 506; Carl Zeiss) 48 h after
transfection. Images of the cells were composed from white brand
images and green fluorescence images.
siRNA transfections
The B76/Huh7 cells were seeded at 2×105
cells/well into 6-well dishes and cultured overnight until they
reached 70–80% confluence. Three groups: TLR2-siRNA-transfected
groups; scramble control group; and the blank group were treated in
parallel. The sequences for the siRNAs are shown in Table I. Transfections were also performed
with Lipofectamine™ 3000 reagent. Transfected cells were incubated
for 24 h and harvested for reverse transcription-quantitative
(RT-q)PCR. In regards to the other cells, 48 h after transfection,
these cells were collected for western blotting.
 | Table I.Primer pairs used for the siRNA
sequences. |
Table I.
Primer pairs used for the siRNA
sequences.
| siRNA name | Sequences
(5′-3′) |
|---|
| TLR2-homo-1375
F |
GCCCUCUCUACAAACUUUATT |
| TLR2-homo-1375
R |
UAAAGUUUGUAGAGAGGGCTT |
| TLR2-homo-1648
F |
GCAACUCAAAGAACUUUAUTT |
| TLR2-homo-1648
R |
AUAAAGUUCUUUGAGUUGCTT |
| TLR2-homo-950
R |
GGUGAAACAAAUUCAUUGATT |
| TLR2-homo-950
F |
UCAAUGAAUUUGUUUCACCTT |
| MYD88-homo-987
F |
CCCAUCAGAAGCGACUGAUTT |
| MYD88-homo-987
R |
AUCAGUCGCUUCUGAUGGGTT |
| MYD88-homo-760
F |
GGCAACUGGAACAGACAAATT |
| MYD88-homo-760
R |
UUUGUCUGUUCCAGUUGCCTT |
| MYD88-homo-316
F |
GCCUGUCUCUGUUCUUGAATT |
| MYD88-homo-316
R |
UUCAAGAACAGAGACAGGCTT |
| Negative control
F |
GAGGCCAAGCCCTGGTATG |
| Negative control
R |
CGGGCCGATTGATCTCAGC |
| NCFAM F |
UUCUCCGAACGUGUCACGUTT |
| NCFAM R |
ACGUGACACGUUCGGAGAATT |
| GAPDH F |
UGACCUCAACUACAUGGUUTT |
| GAPDH R |
AACCAUGUAGUUGAGGUCATT |
RT-qPCR experiments
Total RNA was extracted from fresh liver HCC tissues
and cell lines using Trizol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). Subsequently, 1 µg of total RNA was
reverse-transcribed using GoScript™ Reverse Transcription mix,
Oligo(dT) (Promega Corporation, Madison, WI, USA). For qPCR
analysis, single-stranded cDNA was synthesized using oligo(dT)
primer in a 20 µl reaction mixture. The primer sequences used are
presented in Table II. The PCR
thermocycling conditions were: 95°C for 2 min; and 40 cycles of
95°C for 10 sec, 60°C for 30 sec and 72°C for 30 sec. 18S rRNA was
amplified as a control to normalize expression levels. The level of
each mRNA was expressed as a ratio relative to the housekeeper gene
using the 2−ΔΔCq method (13). Experiments were repeated at least
three times for each sample to ensure the reproducibility of the
results.
| Table II.Primer pairs used for TLR2 pathway
molecules and downstream factors. |
Table II.
Primer pairs used for TLR2 pathway
molecules and downstream factors.
| Oligo name | Primer sequences
(5′-3′) |
|---|
| TLR2 F |
AACTTACTGGGAAATCCTTAC |
| TLR2 R |
AAAAATCTCCAGCAGTAAAAT |
| MYD88 F |
CAGCGACATCCAGTTTGTGC |
| MYD88 R |
GGCCTTCTAGCCAACCTCTT |
| NF-κB F |
AACAGCAGATGGCCCATACCT |
| NF-κB R |
ACGCTGAGGTCCATCTCCTTG |
| 18S F |
AGAAACGGCTACCACATCCA |
| 18S R |
CACCAGACTTGCCCTCCA |
| IL-6 F |
ACTCACCTCTTCAGAACGAATTG |
| IL-6 R |
CCATCTTTGGAAGGTTCAGGTTG |
| IL-8 F |
ACTGAGAGTGATTGAGAGTGGAC |
| IL-8 R |
AACCCTCTGCACCCAGTTTTC |
| TNF-α F |
GAGGCCAAGCCCTGGTATG |
| TNF-α R |
CGGGCCGATTGATCTCAGC |
| IFN-β F |
GCTTGGATTCCTACAAAGAAGCA |
| IFN-β R |
ATAGATGGTCAATGCGGCGTC |
Western blot analysis
Cells were harvested and lysed in RIPA buffer
(Beyotime Institute of Biotechnology, Haimen, China) for 20 min.
The lysate was centrifugated at 17,000 × g at 4°C for 30 min and
protein concentration was determined using a bicinchoninic acid
assay. Cell lysates were resolved using a 12% gel using SDS-PAGE,
and transferred to a PVDF membrane (EMD Millipore, Billerica, MA,
USA). Membranes were blocked in 5% non-fat milk and then blots were
probed with antibodies against TLR2 (cat. no. 12276; dilution
1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), MyD88
(cat. no. 50010; dilution 1:1,000; Cell Signaling Technology),
NF-κB (cat. no. 8242; dilution 1:1,000; Cell Signaling Technology)
or GAPDH (cat. no. sc-47724; dilution 1:8,000; Santa Cruz
Biotechnology, Inc.) overnight at 4°C. GAPDH (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) was used as a loading control
with the same operation. After washing the membrane three times
with PBST for 10 min each wash, the membrane was incubated with the
appropriate horseradish peroxidase-conjugated secondary antibody
(cat. no. sc-2357; 1:5,000; Santa Cruz Biotechnology, Inc. or cat.
no. sc-516102; dilution 1:5,000; Santa Cruz Biotechnology, Inc.).
After washing three times with PBST for 10 min each wash, the
protein bands were visualized using enhanced chemiluminescence.
Densitometry analysis (Bio-Rad Laboratories) was performed using
GAPDH as a control.
Cell proliferation assay and
ELISA
Proliferation of B76/Huh7 cells was measured using
Cell Counting Kit-8 (CCK-8) assay (Phygene, Shanghai, China) at 0,
24, 48 and 72 h following treatment with 5 µg/ml TLR2 agonist
Pam3CSK4. Cells were cultured in 96-well cell plates
(1.5×105/well; 6 wells per condition) overnight, after
which the cells were transfected or treated as described above.
After transfection, a sample of the transfected cells was collected
as the 0 h sample, while the other cells were further cultured for
24, 48 or 72 h. At the end of each treatment period, CCK-8 reagent
was added to the culture medium at a concentration of 5 mg/ml and
incubated for 4 h at 37°C. The supernatant was then removed, and
cells were mixed with 100 µl/well CCK-8 solution. The absorption
was measured using a microplate reader (Spectra Max M5; Molecular
Devices, LLC, Sunnyvale, CA, USA) at 450 nm. The supernatant was
collected, and ELISA kits were used to measure IL-6, IL-8, IFN-β,
TNF-α production (R&D Systems China Co., Ltd., Shanghai, China)
24, 48 and 72 h after transfection.
Flow cytometry for determining
apoptosis
A total of 48 h after transfection, the cells were
trypsinized, collected, washed twice with cold PBS and
1×106 cells were resuspended in 100 µl of Annexin V
binding buffer. A total of 5 µl APC Annexin V and 5 µl 7-AAD (BD
Biosciences) was added into the cell suspension. The cells were
gently vortexed and incubated for 15 min at room temperature in the
dark. A total of 400 µl binding buffer was added to each tube and
analyzed by flow cytometry.
Statistical analysis
Statistical analysis was performed using GraphPad
6.0 (GraphPad Software Inc, La Jolla, CA, USA). Data from at least
three experiments are expressed as mean value ± standard deviation
and were interpreted using one-way ANOVA followed by Bonferroni or
Dunnett test where relevant. All comparisons were determined using
a repeated measures one-way ANOVA between the experimental group
and the control group. P<0.05 was considered to indicate a
statistically significant difference.
Results
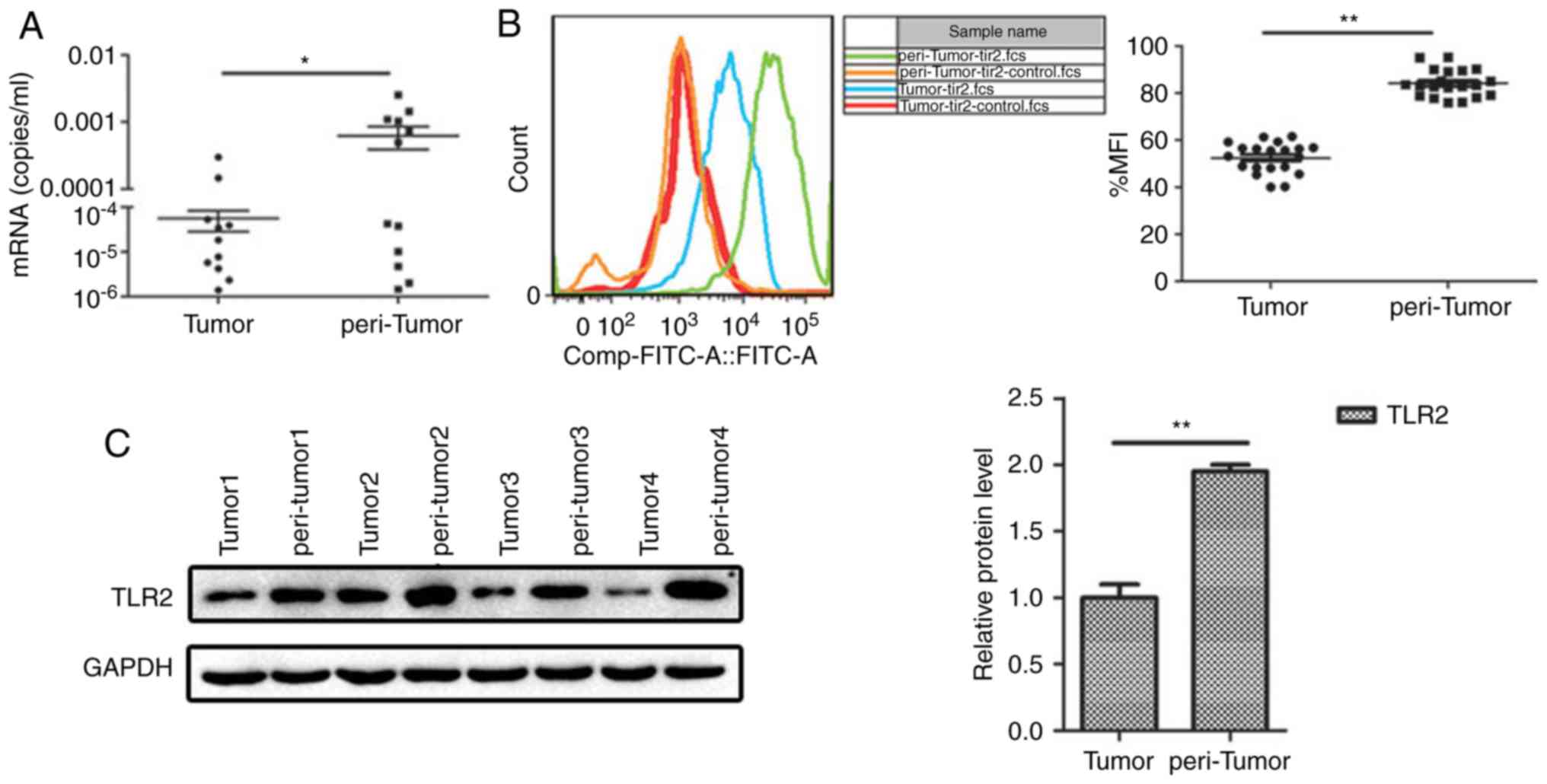
TLR2 expression is significantly
downregulated in liver tumor tissue of patients with HCC
To examine the role of TLR2 in HCC progression,
semi-quantitative RT-PCR and FCM were used. The results showed that
TLR2 expression in the peritumoral tissues was significantly higher
compared with that noted in the tumor tissues (Fig. 1A and B). The protein expression level
of TLR2 was determined using western blotting, and similar to the
PCR results, protein expression of TLR2 was significantly higher in
the peritumoral tissue compared with that noted in the tumor tissue
(Fig. 1C). Our complementary study of
141 samples showed the expression levels of TLR2 in the tissues of
liver and no differences were found for different stage and
prognosis (Table SI).
TLR2 signaling pathway and HCC
proliferation
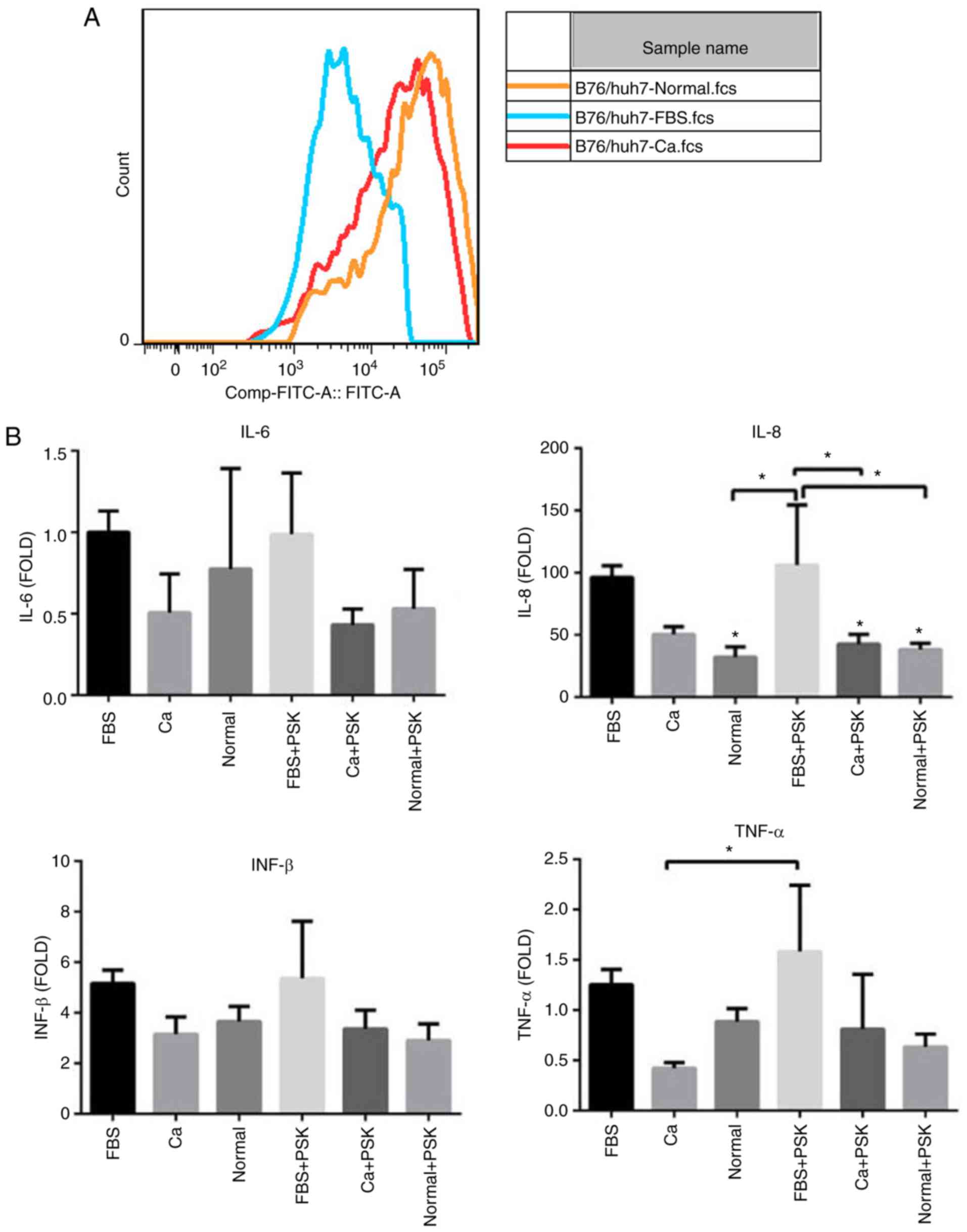
To distinguish whether the serum of patients with
HCC was able to interfere with the TLR2 signaling pathway in HCC
cells through the activation of Pam3CSK4, the TLR2 expression in
B76/Huh7 cells was detected by FCM after 48 h of culturing with the
serum of HCC patients (Ca), with the serum of normal healthy
individuals (Normal) or FBS. The results showed that the expression
of TLR2 decreased in the cells treated with serum from HCC patients
when compared with cells treated with the serum from healthy
individuals (Fig. 2A). Transcription
of various inflammatory cytokines such as IL-8 and TNF-α was
altered, as determined by qPCR. Compared with the cells treated
with FBS, expression of inflammatory cytokines was decreased in
cells treated with serum from patients with HCC (Ca) and in cells
treated with serum from healthy individuals (Normal). Inflammatory
cytokine levels of TLR2-mediated activation of cells treated with
the serum of HCC patients and serum from healthy individuals were
lower than in the cells treated with FBS especially following the
activation of Pam3CSK4 (PSK), confirming efficient inverse feedback
(Fig. 2B). This suggests that some
component of the serum from patients with HCC interfered with the
TLR2 signaling pathway through Pam3CSK4 activation in liver cells
and reduced the TLR2-mediated immune inflammation. Yet, this
substance remains unknown and further experiments are
warranted.
To confirm the findings with TLR2 in vitro, a
TLR2 agonist, Pam3CSK4, was used to stimulate B76/Huh7 cells.
Following treatment with the agonist, the cells were assayed by
qPCR, western blotting and FCM (Fig.
S1). The protein expression levels of signaling pathway
proteins, including MyD88, IRAK1, TRAF6 and NF-κB were
significantly increased 48 h after stimulation with Pam3CSK4
(Fig. S2A). The relative protein
levels of the signaling molecules were also increased (Fig. S2B).
Overexpression of TLR2 and Myd88
combined with treatment with the agonist Pam3CSK4 inhibits the
proliferation and accelerates apoptosis
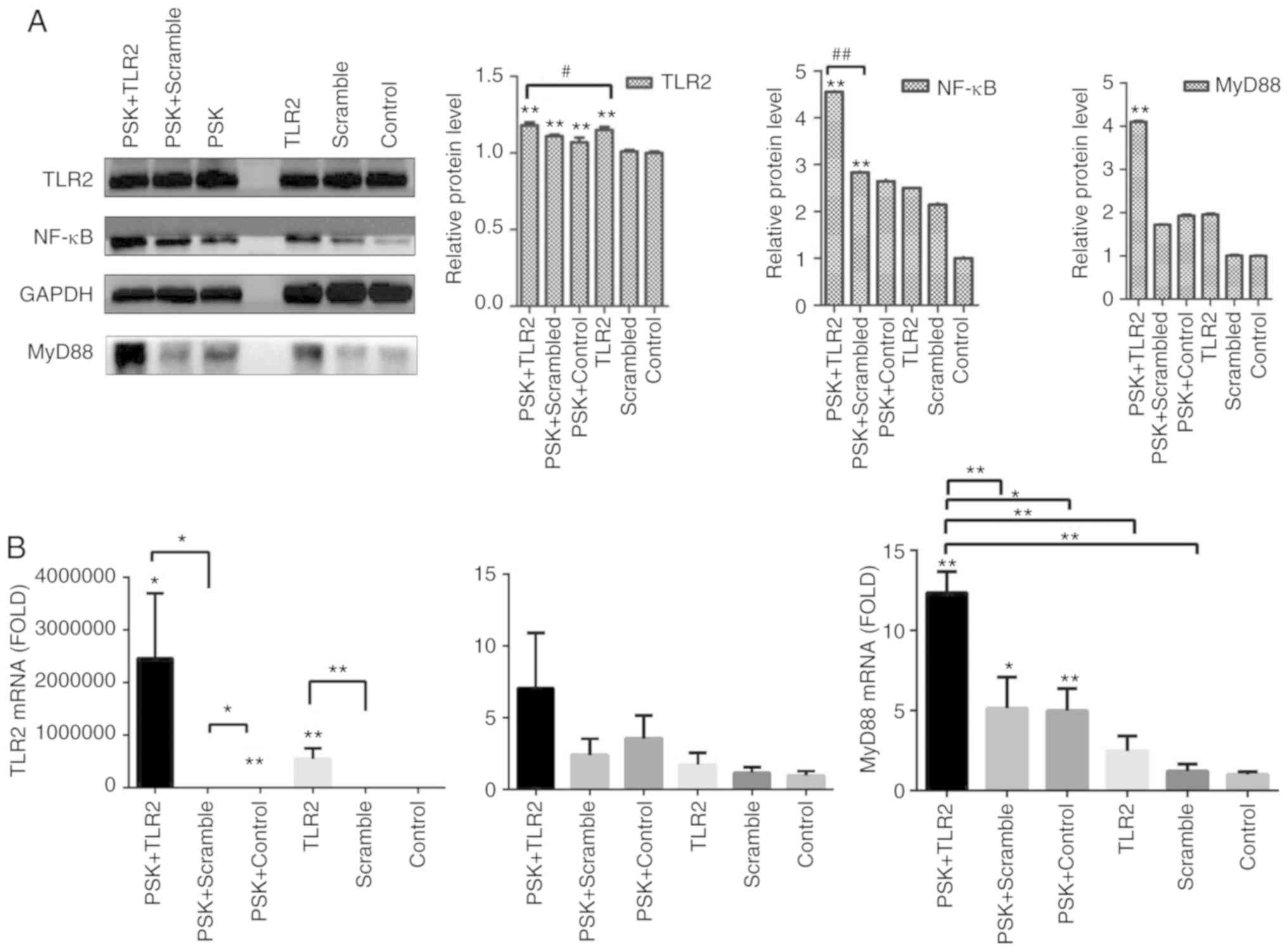
To detect the efficiency of TLR2-overexpression
plasmid and whether the pathway was activated by TLR2 agonist,
B76/Huh7 cells were divided into six groups: TLR2-overexpression
plasmid group; scramble group and the blank group with or without
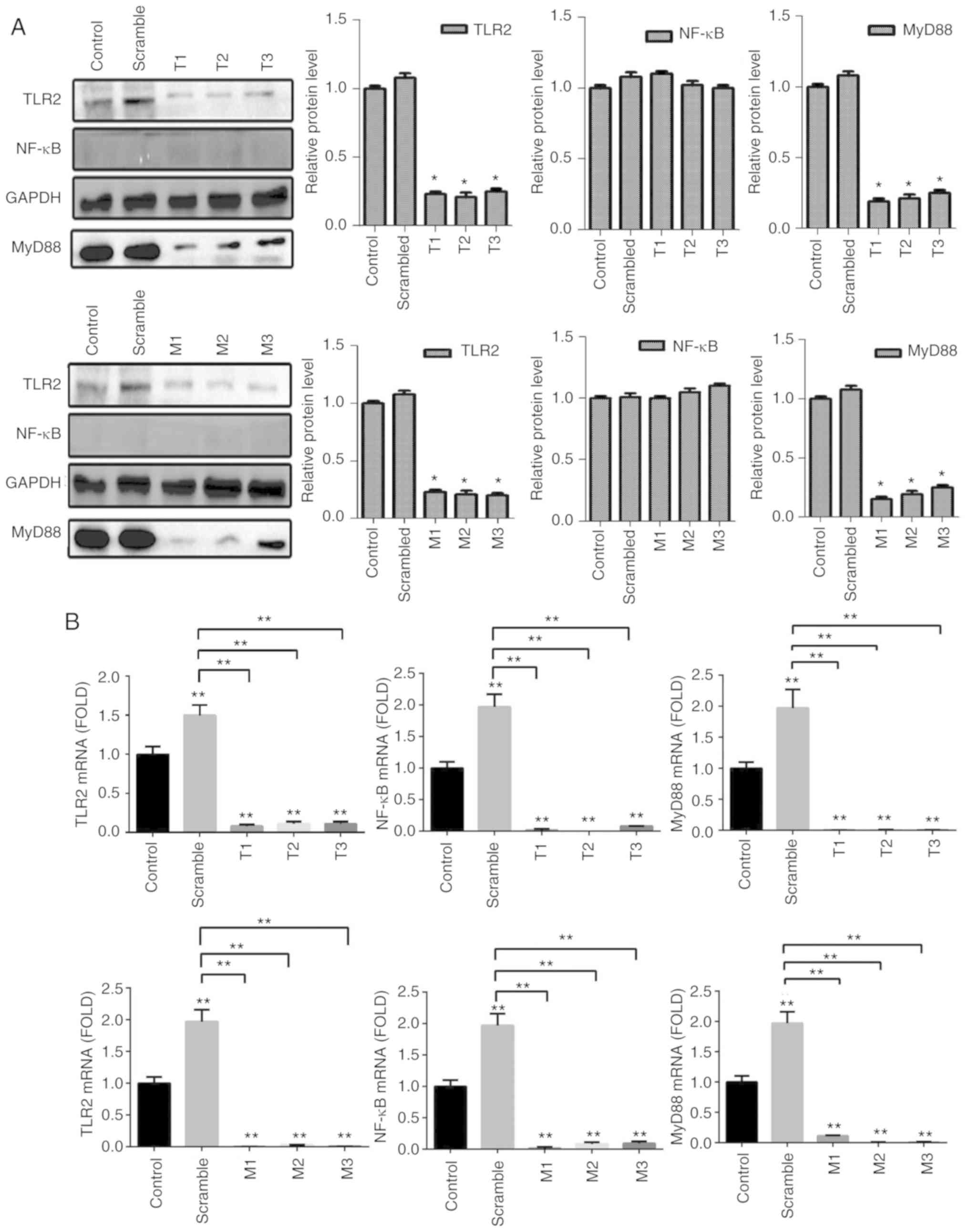
stimulation with Pam3CSK4. The results demonstrated that the
expression of molecules in the TLR2 signaling pathway, such as TLR2
and MyD88 in the TLR2-overexpression groups were significantly
higher than in the scramble groups compared with the control
groups, particularly when stimulated with Pam3CSK4, both at the
protein level (Fig. 3A) and gene
level 48 h after transfection (Fig.
3B). Images of B76/Huh7 cells transfected with the plasmids
were obtained using a fluorescence microscope at the same time
(Fig. S3).
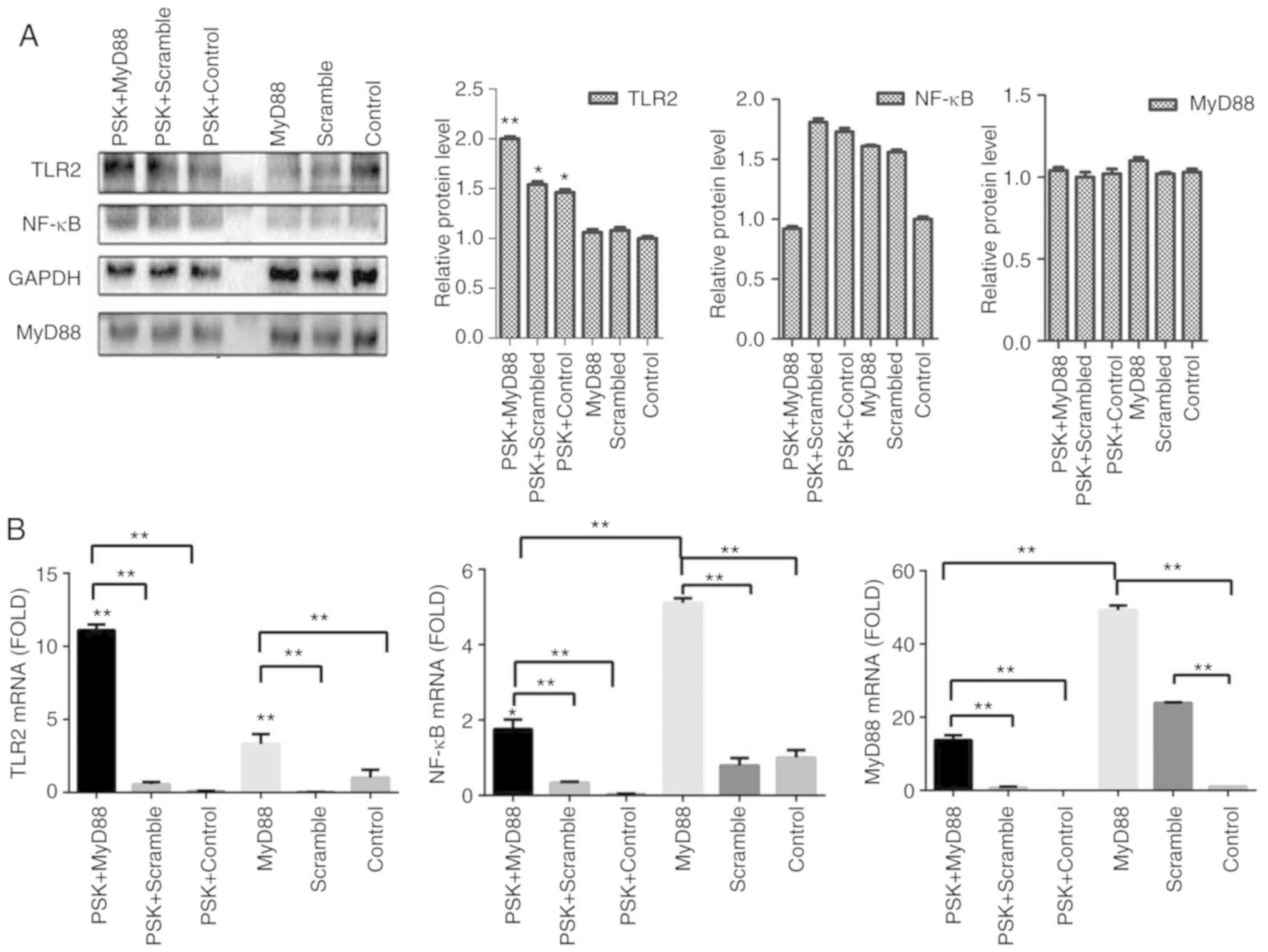
To detect the efficiency of MyD88-overexpression
plasmid, B76/Huh7 cells were also divided into six groups;
MyD88-overexpression plasmid group; scramble group; and the blank
group with or without Pam3CSK4 stimulation. In comparison with the
scramble groups and the control groups, MyD88-overexpression
plasmid group also demonstrated higher expression of TLR2 and MyD88
both at the protein level (Fig. 4A)
and gene level (Fig. 4B) 48 h after
transfection, particularly when stimulated with Pam3CSK4. There was
no consistent corresponding alterations in the expression of NF-κB
following transfection with either of the two plasmids (Figs. 3 and 4).
Images of transfected cells with the plasmids in B76/Huh7 cells
were obtained using a fluorescence microscope (Fig. S4).
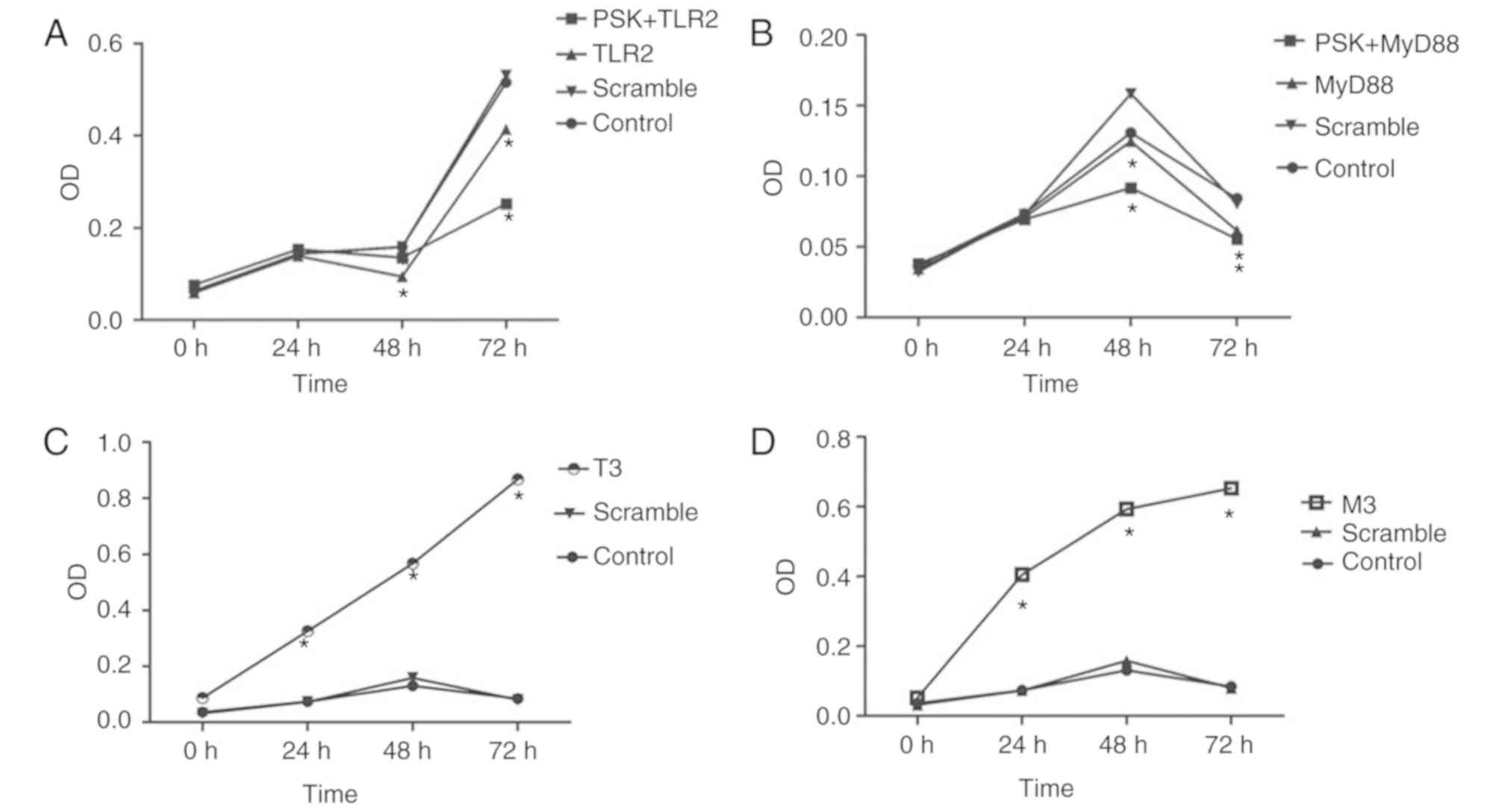
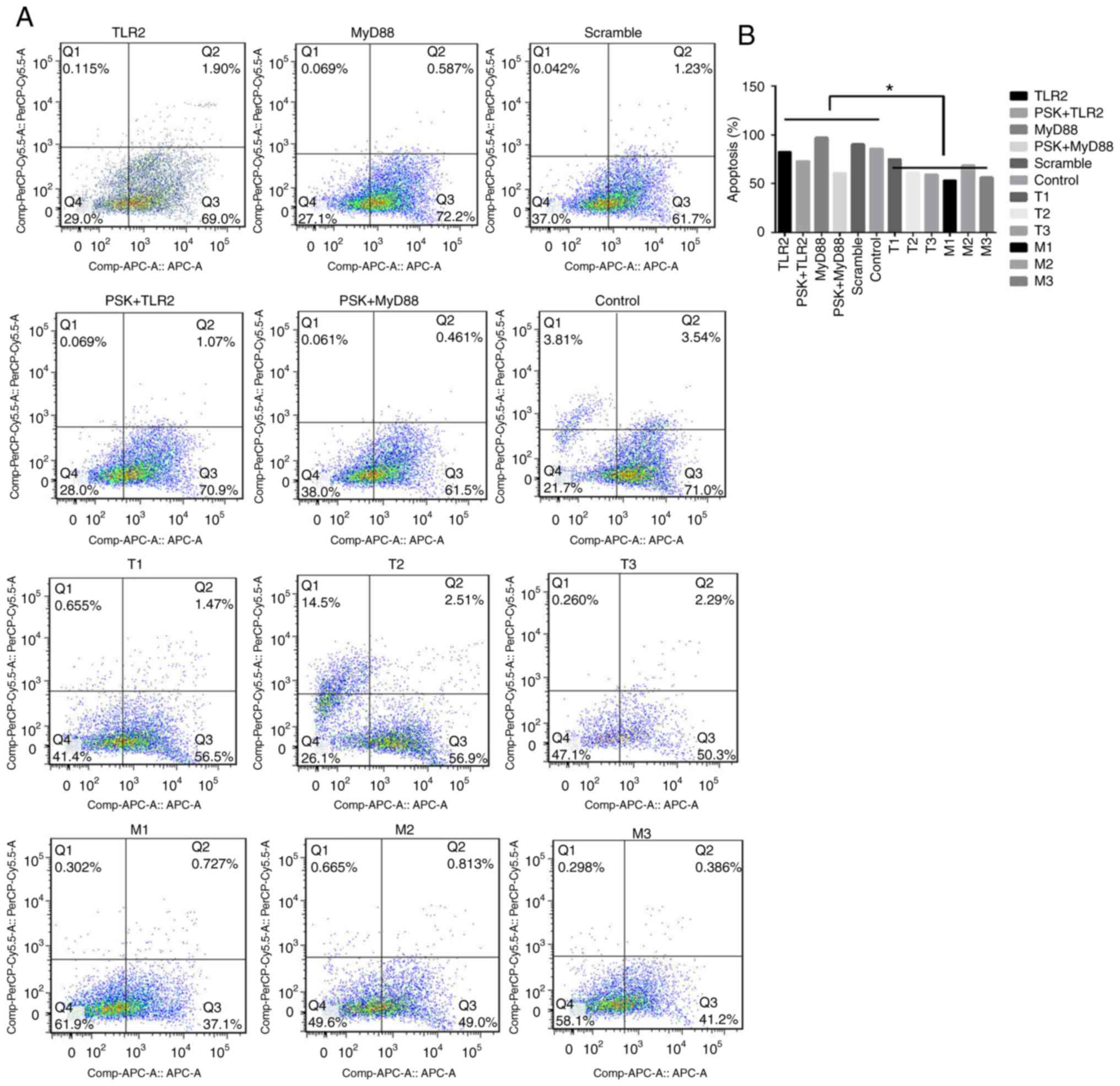
The functional experiments showed that
overexpression of TLR2 and MyD88 significantly inhibited the
proliferation of B76/Huh7 cells (Fig. 6A
and B) and significantly increased apoptosis of B76/Huh7 cells
(Fig. 7A and B).
RNA interference increases
proliferation and decreases apoptosis
To detect the efficiency of TLR2-siRNA or
MyD88-siRNA, cells were divided into five groups per a target RNA:
siRNA1 group, siRNA2 group, siRNA3 group, scramble group and the
blank group. The results of the qPCR and western blotting
demonstrated that the expression of protein and messenger RNA
(mRNA) of TLR2 or MyD88 in the siRNA groups was significantly
decreased compared with the blank groups (P<0.05). It was
observed that all six different siRNAs significantly reduced
expression of TLR2 and MyD88 at the protein (Fig. 5A) and gene expression level (Fig. 5B). The results also showed that the
expression of NF-κB did not significantly differ at the gene and
protein level. The siRNAs with the largest decrease in the
expression levels for both TLR2 and MyD88 were used for the
subsequent experiments. Knockdown of TLR2 or MyD88 significantly
increased proliferation (Fig. 6C and
D) and decreased apoptosis significantly (Fig. 7A and B) in the B76/Huh7 cells.
Inflammatory cytokines serve crucial
roles in antitumor activity
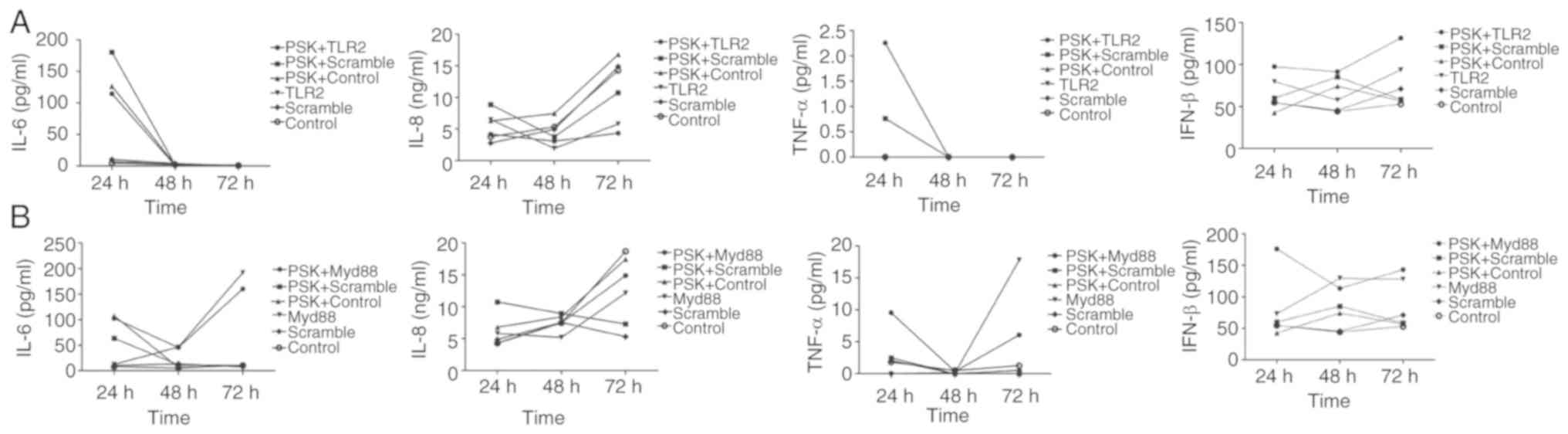
To investigate the role of cytokines through the
activation of the TLR2 signaling pathway, IL-6, IL-8, TNF-α and
IFN-β concentrations were assayed in the supernatant of cells using
ELISA, 24, 48 and 72 h after transfection of the
TLR2-overexpression-plasmid or MyD88-overexpression-plasmid in
B76/Huh7 cell line. The results showed that IL-6 and TNF-α
expression was decreased and IL-8 and IFN-β secretion was increased
slowly between 24 and 48 h in the TLR2-overexpressing cells and
following stimulation with Pam3CSK4 in the supernatants of the
B76/Huh7 cells. IL-6 and TNF-α secretion was decreased in the
MyD88-overexpressing cells and following stimulation with Pam3CSK4
between the 24 and 48 h timepoints in the supernatants of B76/Huh7
cells. IL-6 and TNF-α secretion was increased in the
MyD88-overexpressing cells and Pam3CSK4 stimulated cells between 48
and 72 h. But IL-8 secretion was increased slowly between 24 and 72
h in the supernatants of the B76/Huh7 cells and IFN-β was decreased
slightly in quantitative terms between 24 h and 72 h in the
supernatants of B76/Huh7 cells (Fig.
8). Next it was determined whether the function of the TLR2
pathway was associated with IL-6, IL-8, TNF-α, and IFN-β in
B76/Huh7 cells lines. The results showed that the proinflammatory
cytokines, such as IL-6 and TNF-α were decreased and
anti-inflammatory cytokines, including IL-8 and IFN-β increased
when the cells were transfected with the TLR2 plasmid in B76/Huh7
cells. IL-6 and TNF-α secretion was increased between 48 and 72 h
suggesting that additional MyD88 pathways were also stimulated in
B76/Huh7 cells.
Discussion
TLR2 appears to be an essential stress-sensor the
absence of which reveals an augmented tendency to accumulate damage
to the DNA and reduce cell survival in HCC (5). In the present study, TLR2 expression in
HCC, its effect on cellular proliferation, apoptosis as well as
secretion of cytokines in B76/Huh7 cell line was evaluated. In HCC
tissues, mRNA and protein expression levels of TLR2 were decreased
in tumoral tissues compared with that in the peritumoral tissues.
Additionally, the expression levels of TLR2 were significantly
increased when TLR2 and/or MyD88 were overexpressed and
downregulated when expression of either of these was silenced.
Activation of TLR2 through overexpression of proteins involved in
the signaling TLR2 pathway decreased proliferation of B76/Huh7
cells. The mRNA and protein expression levels of genes encoded by
the plasmids were significantly upregulated in B76/Huh7 cells when
stimulated with Pam3CSK4, suggesting that the TLR2 agonist enhanced
the expression of the plasmids. However, inhibition of the
signaling pathway caused a significant increase in the downstream
signaling cascade, thus increasing proliferation in the B76/Huh7
cell lines. Finally, secretion of IL-6 and TNF-α was decreased,
whereas secretion of IL-8 and IFN-β was increased following
transfection with either of the overexpression plasmids in B76/Huh7
cells.
Several animals (6,7) and in
vitro (10,11,14)
studies have implicated TLR2 in human liver tumors. In wild-type
mice with diethylnitrosamine (DEN)-induced HCC, deficiency of TLR2
resulted in the suppression of immune responses to DEN-induced
injury, thus serving an important role in liver carcinogenesis and
tumor progression (6). TLR1/2
stimulation in mouse LSECs increased CD8+ T cell
immunity in vitro (7). A
previous demonstrated that the activation of the ASK1/p38
MAPK/NF-κB pathway is critical for both neutralizing reactive
oxygen species/endoplasmic reticulum stress and repairing damaged
DNA (15). However, studies on the
underlying mechanism demonstrating a causal link between TLR2 and
tumor proliferation in the liver tumor microenvironment are rare.
In the present study, it was demonstrated that TLR2 expression in
the HCC tissues was associated with increased expression of
molecules involved in the TLR2 signaling pathway. Both TLR2 and
MyD88 are important pathway molecules affecting the proliferation
and apoptosis of HCC cells (16,17). In
theory, we conclude that MyD88 is a more popular signaling pathway
molecule because MyD88 is also included in many Toll like receptor
signaling systems (4,7,8,9,11) and
plays roles in affecting the proliferation and apoptosis of HCC
cells (18). Modulation of TLR2 was
more pronounced when TLR2 was overexpressed compared with MyD88 in
the cell proliferation or apoptosis assays and this may be because
many TLRs potently activate the NF-κB pathway which subsequently
alters MyD88 molecules. The TLR2 agonist, Pam3CSK4, was used to
activate the TLR2 signaling pathway. Pam3CSK4 has been demonstrated
to alter the expression of pathway molecules and inhibit tumor
growth in wild-type mice (11). TLR2
is primarily expressed on traditional immune cells such as
monocytes (19), macrophages
(20), dendritic cells (21), B cells (22) and T cells (23). Recently, studies have demonstrated the
effect of the knockdown of the TLR2 gene in various hepatoma cell
lines (10,24). Huang et al indicated that the
knock down of TLR2 in BEL-7402 cells could resist immune cell
attack and evade immune surveillance (24). Furthermore, downstream expression of
IL-6 was decreased. These results agree with the results of the
present study in B76/Huh7 cells were silencing of TLR2 could
promote tumor proliferation and inhibit apoptosis. Shi et al
(10) found that proliferation,
invasion and migration in the siRNA group was decreased compared
with the control, and the apoptotic ratio was increased in
TLR2-siRNA transfected HepG2 cells treated with recombinant high
mobility group box 1 protein. These results contradict the results
of the present study where silencing of TLR2 and MyD88 in B76/Huh7
cells increased proliferation and inhibited apoptosis. The results
suggest that modulation of the TLR2 pathway may differ in different
cell lines. There are many other TLR2 agonists except for Pam3CSK4
(25). Regulation of TLR2 expression
may be achieved through the NF-κB signaling pathway, which is one
of the major sensors of cytokine secretion (26). Cytokines are important mediators of
cancer-promoting inflammation (27).
IL-6 and TNF-α are proinflammatory mediators in the chronically
injured liver and contribute to the development of HCC (26,28). In
the present study, secretion of IL-6 and TNF-α was decreased
following transfection with the overexpression plasmids. These
cytokines target hepatoma cells, such as primary immune cells, and
reduce their proliferation (29).
IL-8 induced apoptosis of HCC and IFN-β attenuated hepatocellular
carcinoma progression through inhibition of the AKT/FOXO3a pathway
(30,31). IL-8 and IFN-β anti-inflammatory
cytokines were increased and had the opposite effect on hepatoma
cells following transfection with the overexpression plasmids. Many
cytokines in the serum of HCC patients were investigated, and the
volume of tumor was shown to be associated with cytokines (32–35). These
cytokines may serve an important pro- or anti-tumorigenic role
which may be altered by the TLR2 pathway, thus modulating
proliferation (35). IL-6 and TNF-α
secretion was increased between 48 and 72 h after transfection with
pcDNA-MYD88-GFP, suggesting that other MyD88 pathways may be
stimulated in B76/Huh7 cells (36–38). Yet,
we concluded that the content of cytokines was different in serum
of HCC patients and normal individuals. Differences are also
defined in regards to other factors, such as the ethnicity of the
study population and the disease stage (39,27).
Additional experimentation is required to support this
hypothesis.
In summary, the downregulation of TLR2 in the HCC
tissues, otherwise the increasing expression of TLR2 was associated
with a recovery in HCC progression. The liver cells were also
considered as immune cells and associated with signaling network.
The TLR2 signaling pathway is considered as a potential antiviral
mechanism in the hepatitis B infection of hepatic cell lines
(40). The TLR2 signaling pathway in
immune cells is also a subject of increasing study, and it is
hypothesized to trigger the escape of immune-mediated tumor cells
and thus tumor progression (15). The
results of the present study contributed to understanding the role
of the TLR2 pathway in HCC and attempted to partly explain the
carcinogenicity and immunogenicity of TLR2 in the liver. But
experiments in vivo were not conducted in the present study
and further studies of TLR2 in vivo in regards to HCC are
warranted.
TLR2 is an important Toll like receptor family
members which inhibits the development of HCC. The use of
systemically delivered TLR2 to activate the TLR2 signaling pathway
may provide a novel treatment for the prevention of cancer
progression, potentially leading to improved prospects of
survival.
Supplementary Material
Supporting Data
Acknowledgements
The author thank Dr MenJiLu and Dr Yingchao Wang for
providing technical support.
Funding
This study was supported by Fuzhou Science and
Technology Plan Projects (grant no. 2015-S-143-12); the Health and
Family Planning Commission Project of Fujian Province (grant no.
2015-CXB-30); the National Nature Science Foundation of China
(grant no. 81670532); the Provincial Natural Science Foundation of
Fujian (grant no. 2015J01361); and the Fuzhou Infectious Diseases
Medical Center (grant no. 003040008).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YC and ZH conceived and designed the experiments. YC
performed the experiments. YC, ZH and XC collected and analyzed the
data. YC and HY interpreted the findings and wrote the manuscript.
All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the Mengchao Hepatobiliary Hospital of Fujian Medical University
(study license no. 2016-007-01). Informed consent was obtained from
all patients and healthy recruits.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no potential competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
PRR
|
pattern recognition receptor
|
|
TLR2
|
Toll-like receptor 2
|
|
MyD88
|
myeloid differentiation factor 88
|
|
IRAK1
|
interleukin-1 receptor-associated
kinase 1
|
|
TRAF6
|
tumor necrosis factor
receptor-associated factor-6
|
|
ASK1
|
apoptosis signal regulating kinase
1
|
|
p38 MAPK
|
p38 mitogen-activated protein
kinase
|
|
NF-κB
|
nuclear factor-κB
|
|
CCK-8
|
Cell Counting Kit-8
|
|
LSECs
|
liver sinusoidal endothelial cells
|
|
IL-6
|
interleukin-6
|
|
IL-8
|
interleukin-8
|
|
TNF-α
|
tumor necrosis factor-α
|
|
IFN-β
|
interferon-β
|
|
FCM
|
flow cytometry
|
|
Pam3CSK4
|
TLR1/2 agonist
|
|
DEN
|
diethylnitrosamine
|
References
|
1
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen WQ, Zheng RS, Zhang SW, Li N, Zhao P,
Li GL, Wu LY and He J: Report of incidence and mortality in china
cancer registries, 2008. Chin J Cancer Res. 24:171–180. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bieghs V and Trautwein C: The innate
immune response during liver inflammation and metabolic disease.
Trends Immunol. 34:446–452. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nolan JP: Endotoxin, reticuloendothelial
function, and liver injury. Hepatology. 1:458–465. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jorge André Gomes Lopes, Marta
Borges-Canha and PedroPimentel-Nunes: Innate immunity and
hepatocarcinoma: Can toll-like receptors open the door to
oncogenesis? World J Hepatol. 8:162–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oliveira-Nascimento L, Massari P and
Wetzler LM: The role of TLR2 in infection and immunity. Front
Immunol. 3:792012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin H, Yan J, Wang Z, Hua F, Yu J, Sun W,
Li K, Liu H, Yang H, Lv Q, et al: Loss of immunity-supported
senescence enhances susceptibility to hepatocellular carcinogenesis
and progression in toll-like receptor 2-deficient mice. Hepatology.
57:171–182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Jiang M, Ma Z, Dietze KK, Zelinskyy
G, Yang D, Dittmer U, Schlaak JF, Roggendorf M and Lu M: TLR1/2
ligand-stimulated mouse liver endothelial cells secrete IL-12 and
trigger CD8+ T cell immunity in vitro. J Immunol.
191:6178–6190. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Soares JB, Pimentel-Nunes P, Afonso L,
Rolanda C, Lopes P, Roncon-Albuquerque R Jr, Gonçalves N,
Boal-Carvalho I, Pardal F, Lopes S, et al: Increased hepatic
expression of TLR2 and TLR4 in the hepatic
inflammation-fibrosis-carcinoma sequence. Innate Immun. 18:700–708.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shi W, Su L, Li Q, Sun L, Lv J, Li J and
Cheng B: Suppression of toll-like receptor 2 expression inhibits
the bioactivity of human hepatocellular carcinoma. Spring.
35:9627–9637. 2014.
|
|
11
|
Li S, Sun R, Chen Y, Wei H and Tian Z:
TLR2 limits development of hepatocellular carcinoma by reducing
IL18-mediated immunosuppression. Cancer Res. 75:986–995. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Llovet JM, Brú C and Bruix J: Prognosis of
hepatocellular carcinoma: The BCLC staging classification. Semin
Liver Dis. 19:329–338. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu X, Gong J and Xu B: miR-143
down-regulates TLR2 expression in hepatoma cells and inhibits
hepatoma cell proliferation and invasion. Int J Clin Exp Pathol.
8:12738–12747. 2015.PubMed/NCBI
|
|
15
|
Kennedy NJ, Cellurale C and Davis RJ: A
radical role for p38 MAPK in tumor initiation. Cancer Cell.
11:101–103. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Testro AG and Visvanathan K: Toll-like
receptors and their role in gastrointestinal disease. J
Gastroenterol Hepatol. 24:943–954. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Karin M and Greten FR: NF-kappaB: Linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
French SW, Oliva J, French BA, Li J and
Bardag-Gorce F: Alcohol, nutrition and liver cancer: Role of
toll-like receptor signaling. World J Gastroenterol. 16:1344–1348.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang Z, Ge J, Pang J, Liu H, Chen J, Liao
B, Huang X, Zuo D, Sun J, Lu M, et al: Aberrant expression and
dysfunction of TLR2 and its soluble form in chronic HBV infection
and its regulation by antiviral therapy. Antiviral Res. 118:10–19.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Leiguez E, Giannotti KC, Moreira V,
Matsubara MH, Gutiérrez JM, Lomonte B, Rodríguez JP, Balsinde J and
Teixeira C: Critical role of TLR2 and MyD88 for functional response
of macrophages to a group IIA-secreted phospholipase A2 from snake
venom. PLoS One. 9:e937412014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen KY, Song YC, Chen IH, Leng CH, Chen
HW, Li HJ, Chong P and Liu SJ: Molecular mechanisms of
TLR2-mediated antigen cross-presentation in dendritic cells. J
Immunol. 192:4233–4241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bhowmick R, Pore D and Chakrabarti MK:
Outer membrane protein A (OmpA) of Shigella flexneri 2a induces
TLR2-mediated activation of B cells: Involvement of protein
tyrosine kinase, ERK and NF-κB. PLoS One. 9:e1091072014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Derkow K, Krüger C, Dembny P and Lehnardt
S: Microglia induce neurotoxic IL-17+ γδ T cells dependent on TLR2,
TLR4, and TLR9 activation. PLoS One. 10:e01358982015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang Y, Cai B, Xu M, Qiu Z, Tao Y, Zhang
Y, Wang J, Xu Y, Zhou Y, Yang J, et al: Gene silencing of toll-like
receptor 2 inhibits proliferation of human liver cancer cells and
secretion of inflammatory cytokines. PLoS One. 7:e388902012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Landais I, Pelton C, Streblow D,
DeFilippis V, McWeeney S and Nelson JA: Human cytomegalovirus
miR-UL112-3p targets TLR2 and modulates the TLR2/IRAK1/NFκB
signaling pathway. PLoS Pathog. 11:e10048812015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seki E and Schwabe RF: Hepatic
inflammation and fibrosis: Functional links and key pathways.
Hepatology. 61:1066–1079. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Koca YS, Bulbul M and Barut I: The
diagnostic roles of cytokines in hepatobiliary cancers. Biomed Res
Int. 2017:29793072017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heikkilä K, Ebrahim S and Lawlor DA:
Systematic review of the association between circulating
interleukin-6 (IL-6) and cancer. Eur J Cancer. 44:937–945. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kuraishy A, Karin M and Grivennikov SI:
Tumor promotion via injury- and death-induced inflammation.
Immunity. 35:467–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Choi SH, Park JY, Kang W, Kim SU, Kim Y,
Ahn SH, Ro SW and Han KH: Knockdown of HIF-1α and IL-8 induced
apoptosis of hepatocellular carcinoma triggers apoptosis of
vascular endothelial cells. Apoptosis. 21:85–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xie C, Xie DY, Lin BL, Zhang GL, Wang PP,
Peng L and Gao ZL: Interferon-β gene-modified human bone marrow
mesenchymal stem cells attenuate hepatocellular carcinoma through
inhibiting AKT/FOXO3a pathway. Br J Cancer. 109:1198–1205. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Taub R: Hepatoprotection via the
IL-6/Stat3 pathway. J Clin Invest. 112:978–980. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E and
Ben-Neriah Y: NF-kappaB functions as a tumour promoter in
inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Haybaeck J, Zeller N, Wolf MJ, Weber A,
Wagner U, Kurrer MO, Bremer J, Iezzi G, Graf R, Clavien PA, et al:
A lymphotoxin-driven pathway to hepatocellular carcinoma. Cancer
Cell. 16:295–308. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiao P, Long X, Zhang L, Ye Y, Guo J, Liu
P, Zhang R, Ning J, Yu W, Wei F and Yu J: Neurotensin/IL-8 pathway
orchestrates local inflammatory response and tumor invasion by
inducing M2 polarization of tumor-associated macrophages and
epithelial-mesenchymal transition of hepatocellular carcinoma
cells. OncoImmunology. 7:e14401662018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dapito DH, Mencin A, Gwak GY, Pradere JP,
Jang MK, Mederacke I, Caviglia JM, Khiabanian H, Adeyemi A,
Bataller R, et al: Promotion of hepatocellular carcinoma by the
intestinal microbiota and TLR4. Cancer Cell. 21:504–516. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Takeda K and Akira S: Microbial
recognition by toll-like receptors. J Dermatol Sci. 34:73–82. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Seki E and Brenner DA: Toll-like receptors
and adaptor molecules in liver disease: Update. Hepatology.
48:322–335. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dondeti MF, El-Maadawy EA and Talaat RM:
Hepatitis-related hepatocellular carcinoma: Insights into cytokine
gene polymorphisms. World J Gastroenterol. 22:6800–6816. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhan R, Han Q, Zhang C, Tian Z and Zhang
J: Toll-like receptor 2 (TLR2) and TLR9 play opposing roles in host
innate immunity against Salmonella enterica serovar Typhimurium
infection. Infect Immun. 83:1641–1649. 2015. View Article : Google Scholar : PubMed/NCBI
|