Introduction
Cervical cancer (CC) is among the most common
cancers in women worldwide. It mostly affects women in poor
countries that lack public health infrastructures, CC screening,
and vaccines against the human papillomavirus (HPV). In 2016,
511,000 women [95% confidence interval (CI), 414,000-542,000]
developed CC, and 247,000 women (95% CI, 204,000-263,000) succumbed
to CC worldwide (1). Radical surgery
and radiotherapy are well-established and effective; however,
tumors recur or progress in >30% of patients (2,3). The risk
factors for CC are well known, but few targeted therapies are
available. Thus, it is important to identify the genetic and
molecular abnormalities underlying CC for the development of new
and effective therapies.
Rhomboid genes were first described in Drosophila
melanogaster, where they function as serine proteases and key
regulators of epidermal growth factor receptor (EGFR) signaling
(4). The inactive rhomboid protein 1
(iRhom1 or RHBDF1) is a member of the rhomboid family that lacks
protease activity. This protein traverses the membrane 7 times, and
has a long N-terminal cytoplasmic extension that comprises
approximately half of the polypeptide sequence. This protein occurs
in the endoplasmic reticulum and Golgi apparatus, rather than the
plasma membrane. Several recent studies demonstrated that iRhom1
functions as an oncogene in head and neck neoplasms (5), breast (6)
and colorectal cancer (7). Molecular
studies reported that iRhom1 has a critical function in G-protein
coupled receptor (GPCR) ligand-stimulated processes that activate
EGFR ligands in head and neck squamous cancer cells (5) and breast cancer (6). Overexpression of iRhom1 was revealed to
have a significantly positive correlation with poor overall
survival (OS) in patients with colorectal cancer (7).
Inactive rhomboid protein 2 (iRhom2 or RHBDF2) is
also a catalytically inactive member of the rhomboid family. This
protein mediates the maturation of TNFα converting enzyme (TACE,
also called ADAM17) and trafficking to the cell surface (8). iRhom2 functions as an essential
endoplasmic reticulum-to-Golgi trafficking factor for
metalloproteinase ADAM17 when macrophages secrete TNFα (9,10). We
previously demonstrated that ADAM17 expression was an independent
prognostic indicator for CC (11). In
esophageal cancer, the hyperproliferative phenotype was revealed to
be associated with altered iRhom2 levels and increased EGFR
signaling (12,13). Identification of a novel
p63-associated pathway indicated that therapeutic modulation of
iRhom2 has potential for treatment of hyperproliferative skin
disease and neoplasia (14). Blaydon
et al recently reported that missense mutations in iRhom2
were responsible for the autosomal dominant condition tylosis with
oesophageal cancer (TOC) in three families from the United Kingdom,
United States and Germany (10).
Cancer-associated fibroblasts (CAFs) promote tumorigenesis in
several types of cancers. iRhom2 overexpression was revealed to
occur in CAFs isolated from human diffuse-type gastric cancers
(15). However, the levels and roles
of iRhom1 and iRhom2 in the onset and progression of CC remain
unknown.
In the present study, the expression of iRhom1 and
iRhom2 in CC clinical samples was first assessed. Then, their
associations with the clinicopathological features of the CC
patients were determined and their prognostic value was assessed.
Subsequently, HeLa cells were used to evaluate the effects of
knockdown of iRhom1 and iRhom2 on cell proliferation,
cell cycle distribution, and apoptosis. Finally, microarray
analysis was used to identify the molecular mechanisms responsible
for iRhom-mediated promotion of CC by examination of pathways that
have critical roles in the development and progression of CC.
Materials and methods
Patients and tissue collection
The Ethics Committee of Fujian Provincial Cancer
Hospital, which is affiliated with Fujian Medical University,
provided approval of this study. Samples from 83 consecutive
patients (age range, 32–80 years) with CC were collected for
immunohistochemical (IHC) analysis from January 2010 to December
2012. Prior to surgery, none of the enrolled patients received
chemotherapy or radiotherapy. Cancer classification followed the
2009 Federation International of Gynecology and Obstetrics (FIGO)
protocol, and enrolled patients were followed-up until December
2017 or death. Eligibility was determined following hysterectomy
(total, modified-radical, or radical), bilateral
salpingo-oophorectomy, pelvic lymphadenectomy, or para-aortic
lymphadenectomy. Surgical staging was performed no more than 8
weeks before initiation of radiotherapy. The included patients had
hematological, liver, renal function, and other laboratory
variables within normal ranges (creatinine clearance ≥40 ml/min,
leucocytes ≥4.0×109/l, platelets ≥100×109/l,
and hemoglobin ≥10 g/dl). Any patient with a secondary malignancy,
a serious concomitant systemic disorder, or a psychiatric disease
was deemed ineligible. For validation of each diagnosis, two
independent pathologists evaluated the IHC results. After patients
provided written informed consent, samples were used for analysis.
Twenty fresh CC tissues specimens (11 from squamous cell
carcinomas, 5 from adenocarcinomas, 1 from small-cell carcinoma,
and 3 from adenosquamous carcinomas) and matching non-cancerous
adjacent cervical tissue samples were used for immunoblotting.
Study endpoints
Progression-free survival (PFS, the duration from
enrollment to disease progression or death) was the primary
endpoint, and local-regional failure, distant failure, and OS were
the secondary endpoints.
Reagents
The following antibodies were obtained from Abcam:
iRhom1 (ID product code ab81342), iRhom2 (ID product code
ab116139), Ki67 (ID product code ab92742), β-catenin (ID product
code ab32572), Fas (ID product code ab82419), GSK3B (ID product
code ab32391), Myc (ID product code ab32), TGFBR2 (ID product code
ab78419), EGFR (ID product code ab52894), p-EGFR (ID product code
ab40815), GADPH (ID product code ab8245), and actin (ID product
code ab179467). Horseradish peroxidase (HRP)-labeled secondary
antibodies (ID product code sc-2354 and sc-2357) were purchased
from Santa Cruz Biotechnology, Inc.
Immunohistochemistry
IHC analyses employed formalin-fixed and
paraffin-embedded 5 µm-thick sections. These sections were
deparaffinized in xylene and rehydrated in a graded series of
alcohol. For labeling, the avidin-biotin complex (ABC) method was
used. For antigen retrieval, a steamer was used for 20 min in an
EDTA buffer. Then, samples were treated with 3% hydrogen peroxide
in methanol for 20 min to block endogenous peroxidase activity and
the reaction was blocked for 1 h by incubation in a 5% solution of
non-fat milk. Slides were then incubated with polyclonal rabbit
antibodies against the iRhom1 (1:50 dilution), iRhom2 (1:50
dilution), or Ki67 (1:100 dilution) antibody at room temperature
for 1 h. The secondary antibody (MaxVision HRP-polymer anti-rabbit;
Fuzhou Maixin-Biotech Co., Ltd.) was added, and counterstaining was
performed using Mayer's hematoxylin and diaminobenzidine. Tumor
cells that had staining in the cytoplasm were considered iRhom1- or
iRhom2-positive, and tumor cells that had staining in the nucleus
were considered Ki67-positive.
For IHC analysis of iRhom1, iRhom2 and Ki-67, images
of five typical visual fields of each section were analyzed using
software (Image-Pro Plus 6.0; Media Cybernetics, Inc.), and the
mean absorption was used to score expression. All sections were
scored separately by two pathologists who were unaware of the
clinical parameters of the corresponding patients. Stain intensity
was scored as 0 (negative), 1 (weak), 2 (strong). The percentage of
positively-stained area was 0 (fewer than 10% positive cells), 1
(10 to 25% positive cells), 2 (26 to 50% positive cells), and 3
(50% or more positive cells). The final score was determined from
the average of products in stain intensity and percentage with
staining that were immunoreactive on the cell membranes and/or
cytoplasm by examination of 10 typical microscopic fields. The
final IHC score was defined as 0 (−, negative expression), 1–2 (+,
weak expression), 3–4 (++, medium expression), 5–6 (+++, strong
expression).
Cell culture
HeLa cells (Chinese Academy of Sciences Cell Bank)
were plated in 6-well plates (~2×106 cells/well), in
Dulbecco's modified Eagle's medium with 10% fetal bovine serum
(FBS; 1 ml/well). These cells were grown at 37°C with 5%
CO2 and 95% air. After 24 h, cell attachment had
occurred and cells were then incubated in SBF-free medium for 16 to
18 h.
Recombinant lentiviral vector
construction and cell infection
Short hairpin RNAs (shRNAs) that targeted the
iRhom1 gene (NM_022450.3) and iRhom2 gene
(NM_024599.5) were designed, and lentiviruses were constructed to
knockdown these genes. The shRNA targeting iRhom1 was
GAGGCTGGCGGAAGCAGAA, the shRNA targeting iRhom2 was
CGTGTCTGTGGTCTTTCAA, and the negative control (NC) sequence was
TTCTCCGAACGTGTCACGT. Related stem-loop DNA oligonucleotides were
prepared and inserted into a lentiviral vector (pGCSIL-GFP;
Shanghai GeneChem Co., Ltd.). The Lentivector Expression System
(Shanghai GeneChem Co., Ltd.) was used to produce lentiviruses
expressing iRhom1 shRNA, iRhom2 shRNA, or NC
shRNA.
Infection of HeLa cells with
lentivirus
HeLa cells were grown in 12-well plates, then
infected with a shRNA-expressing lentivirus, according to the
desired multiplicity of infection (MOI). Cells were then observed
using fluorescence microscopy (MicroPublisher 3.3RTV; Olympus
Corp.) at 72 h. Cells were harvested at 120 h to determine
knockdown efficiency using immunoblotting.
Western blotting
Samples were initially added to a
radio-immunoprecipitation buffer (Beyotime Institute of
Biotechnology) with 100 nM phenylmethylsulfonyl fluoride (PMSF).
After assessment of protein concentrations with a bicinchoninic
acid kit (Pierce; Thermo Fisher Scientific, Inc.), proteins (50 µg
per lane) were separated by electrophoresis on 12% SDS-PAGE gels,
and then transferred to PVDF membranes (EMD Millipore). These
membranes were incubated with the primary antibody such as iRhom1
(1:50 dilution), iRhom2 (1:50 dilution), β-catenin (1:1,000
dilution), Fas (1:150 dilution), GSK3B (1:100 dilution), Myc
(1:1,000 dilution), TGFBR2 (1:100 dilution), EGFR (1:1000
dilution), p-EGFR (1:1,000 dilution), GADPH (1:1,500 dilution) and
actin (1:1,500 dilution) in Tris-buffered saline with 0.05%
Tween-20 and 5% non-fat dry milk. After incubation at room
temperature for 2 h, the membranes were washed, and then incubated
with the HRP-labeled secondary antibody (1:2,000 dilution) at room
temperature for 1 h. Electrochemiluminescence (Elecsys 2010; Roche
Diagnostics, Basel, Switzerland) was used for visualization of
proteins. Image Lab version 3.0 software was used to perform the
densitometric analysis of blots.
MTT assay
Transfected HeLa cells were added to a 96-well plate
(1×104 cells/well) and incubated for 24, 48, 72 and 96
h. Then MTT (10 µl; 5 mg/ml) in PBS (pH 7.4, Sigma-Aldrich; Merck
KGaA) was added to each well for 4 h. After removal of the
supernatant, dimethyl sulfoxide (DMSO, 100 µl/well; Thermo Fisher
Scientific, Inc.) was added and the absorbance was measured at 490
nm using a microplate reader. Each experiment was performed in
triplicate, and each measurement was also performed in
triplicate.
Cell cycle analysis
Approximately 1×106 HeLa cells were
treated with trypsin, washed twice in PBS, and then incubated
overnight in 10 ml of cold ethanol. Cells were then centrifuged
with 1,000 × g and the ethanol was removed. They were then washed
twice in PBS, added to 100 µl RNase (30 min at 37°C), and then
centrifuged at 1,000 × g. The pellet was added to 400 µl propidium
iodide, and allowed to stand in darkness for 30 min at 4°C. The BD
FACSCalibur system (BD Biosciences) was then used for flow
cytometry and analysis of cell cycle distribution.
Clone formation assay
For each group, ~1×104 HeLa cells were
added to each well of a 6-well culture plate (3 wells/group). The
cells were incubated for 14 days at 37°C, and the growth medium was
replaced every 3 days. After 14 days, the cells were washed two
times with PBS and 0.5% crystal violet was added for staining in 5
min under ambient temperature. Microscopy (light, 10×10) was used
to determine the number of colonies that had at least 50 cells. The
efficiency of plate clone formation was defined as 100% × (number
of colonies/number of cells inoculated), and there were 3
replicates per experiment.
Cell apoptosis assay with Annexin
V-APC single-color staining
After collection of HeLa cells (~2×105),
allophycocyanin (APC)-labeled Annexin V (cat. no. 88-8007;
eBioscience) were added for single-color staining to detect
apoptotic (Annexin V positive) cells. A total of 1.0×106
cells were washed twice with pre-cooled PBS (pH 7.4), and incubated
for 15 min in 100 µl staining buffer including 5 µl APC-labeled
Annexin V. FACS analysis for Annexin V staining was subsequently
performed by flow cytometry (FACSCalibur; BD Biosciences).
Microarray processing and
analysis
Total RNA was extracted from HeLa cells that were
infected with lentivirus expressing NC/shRNA (n=3) or
iRhom2/shRNA (n=3) by use of the TRIzol reagent. RNA was
assessed using the Thermo NanoDrop 2000 and Agilent 2100
Bioanalyzer. Microarray processing and measurement of gene
expression profiles were performed using the Affymetrix human
GeneChip primeview (Affymetrix Inc.; Thermo Fisher Scientific,
Inc.), using the manufacturer's protocols. The GeneChip 3′ IVT
Expression Kit was used to perform reverse transcription, dsDNA
template transformation, in vitro transcription, and biotin
labeling. The GeneChip Hybridization Wash and Stain Kit was then
used to perform microarray hybridization, washing, and staining.
Finally, the GeneChip Scanner 3000 was employed to perform scanning
of the arrays and to produce the raw data.
Integrative analyses
The integrative analyses were generated using
Ingenuity Pathway Analysis (IPA; Ingenuity Systems®;
www.ingenuity.com). A file that had 596 gene IDs
and mean log2(fold-change) data was uploaded to identify
canonical pathways, upstream regulators, relationships with
cellular functions and diseases, effects on regulators, and network
analysis. A P-value and a Z-score were determined for each
pathway/network so that random data had a low probability of
producing significant predictions. The P-value (from a right-sided
Fisher's exact test) considers the number of input genes and the
number of molecules in the database that could be in networks. The
Z-score indicates activation or inhibition of a pathway/network.
The IPA database is based on experiments and results published in
numerous professional journals. The final graphical representation
of each pathway/network has nodes (genes) and edges (biological
relationships between nodes), with unique symbols used for
different classes of molecules.
Statistical analyses
The statistical analyses were performed using SPSS
(version 19.0; IBM Corp.). The correlations in the expression of
different markers were evaluated using Pearson's coefficient, and
the Chi-square test was used to calculate the statistical
significance of differences between groups. Kaplan-Meier survival
curves were plotted, and the log-rank test was used to determine
the significance of differences between these curves. A
multivariate analysis (Cox's non-parametric model) was performed to
assess the relationship of iRhom1, iRhom2 and Ki-67 expression on
progression-free survival (PFS) and overall survival (OS). The
results from western blotting experiments (means ± standard
deviations) were obtained from at least three independent
experiments. The statistical significance of differences was
determined using Student's t-test and a one-way analysis of
variance (ANOVA) with post hoc Student-Newman-Keuls test. For all
data, a P-value <0.05 was considered to indicate a statistically
significant difference.
Results
Expression of iRhom1, iRhom2 and
Ki-67
Tissue specimens from 83 CC patients were examined
using IHC analysis for iRhom1, iRhom2 and Ki-67. iRhoms (−/+) and
iRhoms (++/+++) were based on the average of products in stain
intensity and percentage of positively-stained area. Pathological
analysis indicated squamous cell carcinomas (n=44), adenocarcinomas
(n=29), small-cell carcinomas (n=4), and adenosquamous carcinomas
(n=6). iRhom1 and iRhom2 were present in the cytoplasm of tumor
cells, and Ki67 was mainly in the nucleus, as indicated by dark
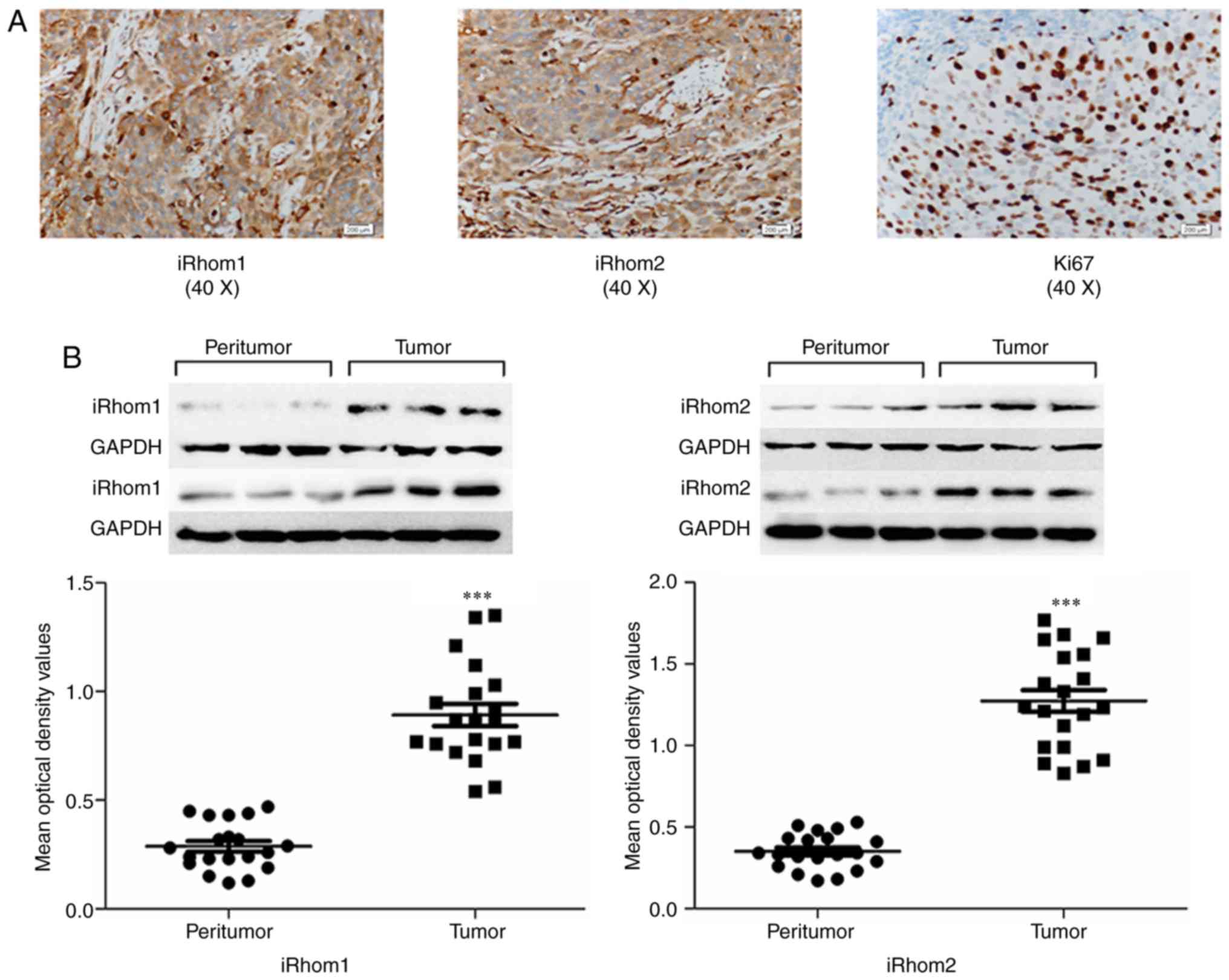
yellow granules (Fig. 1A). There was
high expression of iRhom1 in 55 out of the 83 cases (66.3%), of
iRhom2 in 49 out of the 83 cases (59.1%), and of Ki-67 in 48 out of
the 83 cases (57.8%) (Table I).
 | Table I.Relationship of iRhom1, iRhom2, Ki-67
expression with clinicopathological features of patients with
cervical cancer. |
Table I.
Relationship of iRhom1, iRhom2, Ki-67
expression with clinicopathological features of patients with
cervical cancer.
| Clinical
features | n | iRhom1 (−/+) | iRhom1 (++-
+++) | P-value | iRhom2 (−/+) | iRhom2 (++-
+++) | P-value | Ki-67 (−/+) | Ki-67 (++-
+++) | P-value |
|---|
|
| 83 | 28 | 55 |
| 34 | 49 |
| 35 | 48 |
|
| Age, years |
|
|
| 0.176 |
|
| 0.479 |
|
| 0.146 |
| ≤45 | 40 | 16 | 24 |
| 17 | 23 |
| 14 | 26 |
|
|
>45 | 43 | 12 | 31 |
| 17 | 26 |
| 21 | 22 |
|
| Clinical stage
(FIGO) |
|
0.042a |
|
|
0.010a |
|
|
<0.001a |
|
|
| I | 35 | 16 | 19 |
| 20 | 15 |
| 23 | 12 |
|
| II | 48 | 12 | 36 |
| 14 | 34 |
| 12 | 36 |
|
| Histology |
|
|
| 0.786 |
|
| 0.728 |
|
| 0.116 |
|
Adenocarcinoma | 29 | 10 | 19 |
| 14 | 15 |
| 11 | 18 |
|
|
Squamous carcinoma | 44 | 16 | 28 |
| 16 | 28 |
| 18 | 26 |
|
| Small
cell carcinoma | 4 | 1 | 3 |
| 2 | 2 |
| 4 | 0 |
|
|
Adenosquamous carcinoma | 6 | 1 | 5 |
| 2 | 4 |
| 2 | 4 |
|
| Lymph node
metastasis |
|
|
| 0.081 |
|
| 0.288 |
|
|
0.002a |
|
Positive | 21 | 4 | 17 |
| 7 | 14 |
| 3 | 18 |
|
|
Negative | 62 | 24 | 38 |
| 27 | 35 |
| 32 | 30 |
|
|
Differentiation |
|
|
| 0.287 |
|
| 0.507 |
|
| 0.472 |
|
Well-differentiated | 10 | 4 | 6 |
| 5 | 5 |
| 6 | 4 |
|
|
Moderately-differentiated | 56 | 21 | 35 |
| 24 | 32 |
| 22 | 34 |
|
|
Poorly-differentiated | 17 | 3 | 14 |
| 5 | 12 |
| 7 | 10 |
|
| Tumor size
(cm) |
|
|
|
0.004a |
|
| 0.403 |
|
| 0.069 |
|
>4 | 17 | 1 | 16 |
| 6 | 11 |
| 4 | 13 |
|
| ≤4 | 66 | 27 | 39 |
| 28 | 38 |
| 31 | 35 |
|
| Parametrium
invasion |
|
|
|
0.039a |
|
| 0.136 |
|
| 0.448 |
|
Positive | 16 | 2 | 14 |
| 9 | 7 |
| 6 | 10 |
|
|
Negative | 67 | 26 | 41 |
| 25 | 42 |
| 29 | 38 |
|
The possible function of iRhom1 and iRhom2 was
investigated in CC tumorigenesis by performing western blotting of
normal cervical tissues and adjacent fresh CC tissues. The results
indicated that iRhom1 and iRhom2 had higher expression in CC
tissues than adjacent noncancerous tissues (Fig. 1B, P<0.001).
Correlation of iRhom1, iRhom2 and
Ki-67 with demographic and clinical factors
The correlations of expression of iRhom1, iRhom2 and
Ki-67 with various clinical factors were examined (Table I). All three proteins had a
significant association with FIGO stage (PiRhom1=0.042;
PiRhom2=0.010; PKi-67<0.001). There was
also a significant association between Ki-67 expression and lymph
node metastasis (P=0.002), and significant associations of iRhom1
expression with parametrium invasion (P=0.039) and tumor size
(P=0.04).
Correlation of iRhom1, iRhom2 and
Ki-67 with clinical outcomes
Kaplan-Meier analysis was also used to determine the
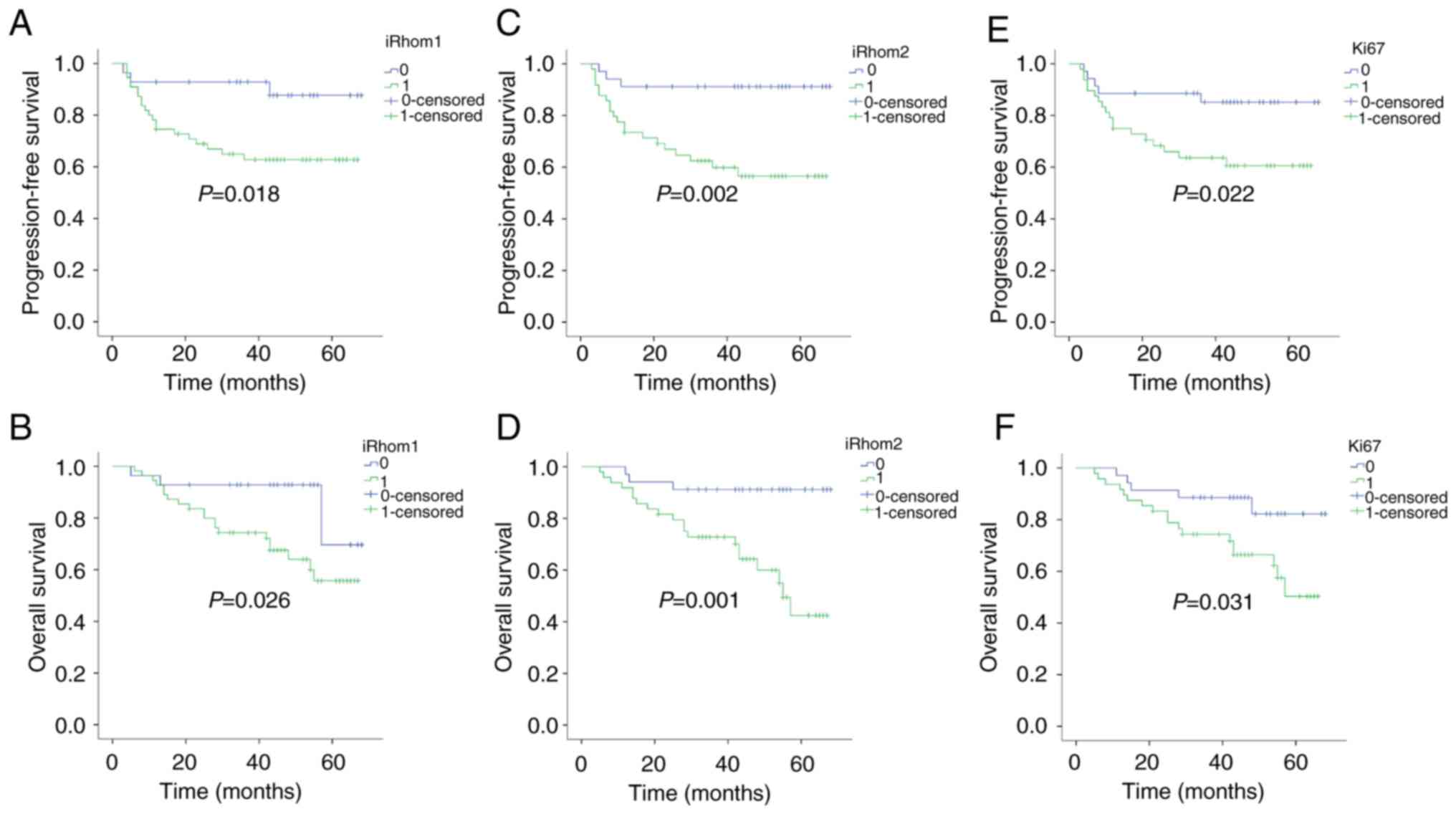
independent prognostic value of iRhom1, iRhom2, and Ki-67 on PFS
and OS (Fig. 2). iRhoms (−/+)
represented weak expression and iRhoms (++/+++) represented strong
expression. The results revealed that patients with strong
expression of each protein had a significantly worse PFS than those
with weak expression (PiRhom1=0.018;
PiRhom2=0.002; PKi-67=0.022). In addition,
patients with strong expression of each protein had significantly
worse OS than those with weak expression (PiRhom1=0.026;
PiRhom2=0.001; PKi-67=0.031).
Multivariate analysis of prognostic
variables
multivariate Cox regression analysis was used to
examine the relationship of OS with iRhom1, iRhom2, Ki-67, age,
histology, stage, metastases to lymph nodes, and invasion of the
parametrium (Table II). The results
indicated that iRhom2 expression [hazard ratio (HR)=4.374; P=0.033;
95% confidence interval (CI)=1.130–16.937] and FIGO stage
(HR=1.778; P=0.027; 95% CI=1.069–2.598) were statistically
significant and independent prognostic factors (Table II). Multivariate analysis of PFS
indicated that iRhom2 expression (HR=4.074; P=0.036; 95%
CI=1.097–15.129) and FIGO stage (HR=1.892; P=0.014; 95%
CI=1.137–3.147) were also statistically significant and independent
prognostic factors (Table II).
| Table II.Multivariate analysis of prognostic
factors for patients with cervical cancer (n=83). |
Table II.
Multivariate analysis of prognostic
factors for patients with cervical cancer (n=83).
| OS |
|---|
|
|---|
| Parameters | Hazard ratio | P-value | 95%CI |
|---|
| Age, years | 1.047 | 0.916 | 0.443–2.476 |
| ≤45 |
|
|
|
| >45 |
|
|
|
| Histology | 0.97 | 0.903 | 0.597–1.576 |
|
Adenocarcinoma |
|
|
|
|
Squamous carcinoma |
|
|
|
| Small
cell carcinoma |
|
|
|
|
Adenosquamous carcinoma |
|
|
|
| Clinical stage
(FIGO) | 1.778 |
0.027a | 1.069–2.598 |
| I |
|
|
|
| II |
|
|
|
| Lymph node
metastasis | 0.475 | 0.152 | 0.156–1.335 |
|
Positive |
|
|
|
|
Negative |
|
|
|
| Parametrium
invasion | 1.767 | 0.342 | 0.546–5.714 |
|
Positive |
|
|
|
|
Negative |
|
|
|
| iRhom1 | 2.737 | 0.119 | 0.771–9.712 |
| iRhom2 | 4.374 |
0.033a | 1.130–16.937 |
| Ki-67 | 1.904 | 0.251 | 0.634–5.719 |
|
| PFS |
|
| Age, years | 1.2 | 0.68 | 0.506–2.846 |
|
≤45 |
|
|
|
|
>45 |
|
|
|
| Histology | 0.986 | 0.956 | 0.597–1.630 |
|
Adenocarcinoma |
|
|
|
|
Squamous carcinoma |
|
|
|
| Small
cell carcinoma |
|
|
|
|
Adenosquamous carcinoma |
|
|
|
| Clinical stage
(FIGO) | 1.892 |
0.014a | 1.137–3.147 |
| I |
|
|
|
| II |
|
|
|
| Lymph node
metastasis | 0.403 | 0.106 | 0.134–1.213 |
|
Positive |
|
|
|
|
Negative |
|
|
|
| Parametrium
invasion | 1.429 | 0.545 | 0.450–4.539 |
|
Positive |
|
|
|
|
Negative |
|
|
|
| iRhom1 | 3.037 | 0.085 | 0.858–10.753 |
| iRhom2 | 4.074 |
0.036a | 1.097–15.129 |
| Ki-67 | 1.876 | 0.272 | 0.611–5.759 |
Oncogenic roles of iRhom1 and iRhom2
knockdown in HeLa cells
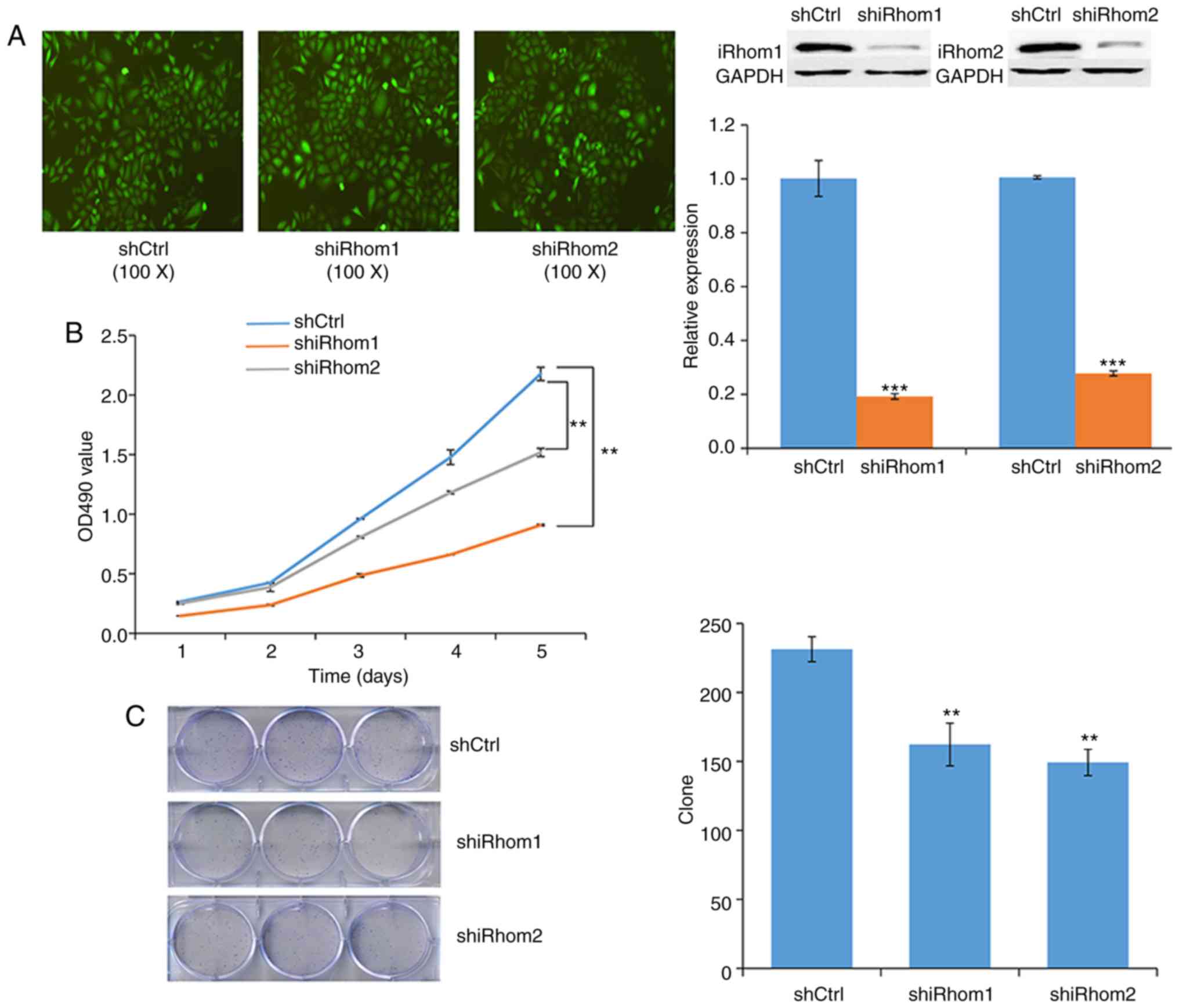
Next, transfected HeLa cells were used to examine
the effect of knockdown of iRhom1 and iRhom2 on CC
cell proliferation in vitro. The reduced expression of each
protein in these cells confirmed the efficiency of silencing
(Fig. 3A). In particular, each shRNA
knocked down expression by >50%. The MTT assay indicated that
cell viability in the controls (OD490 nm) was greater
than in cells with knockdown of iRhom1 or iRhom2 from
day 1 to day 5, and that iRhom2 knockdown had a stronger
effect than iRhom1 knockdown (Fig.
3B). Moreover, knockdown of each gene significantly decreased
the formation of cell colonies (Fig.
3C).
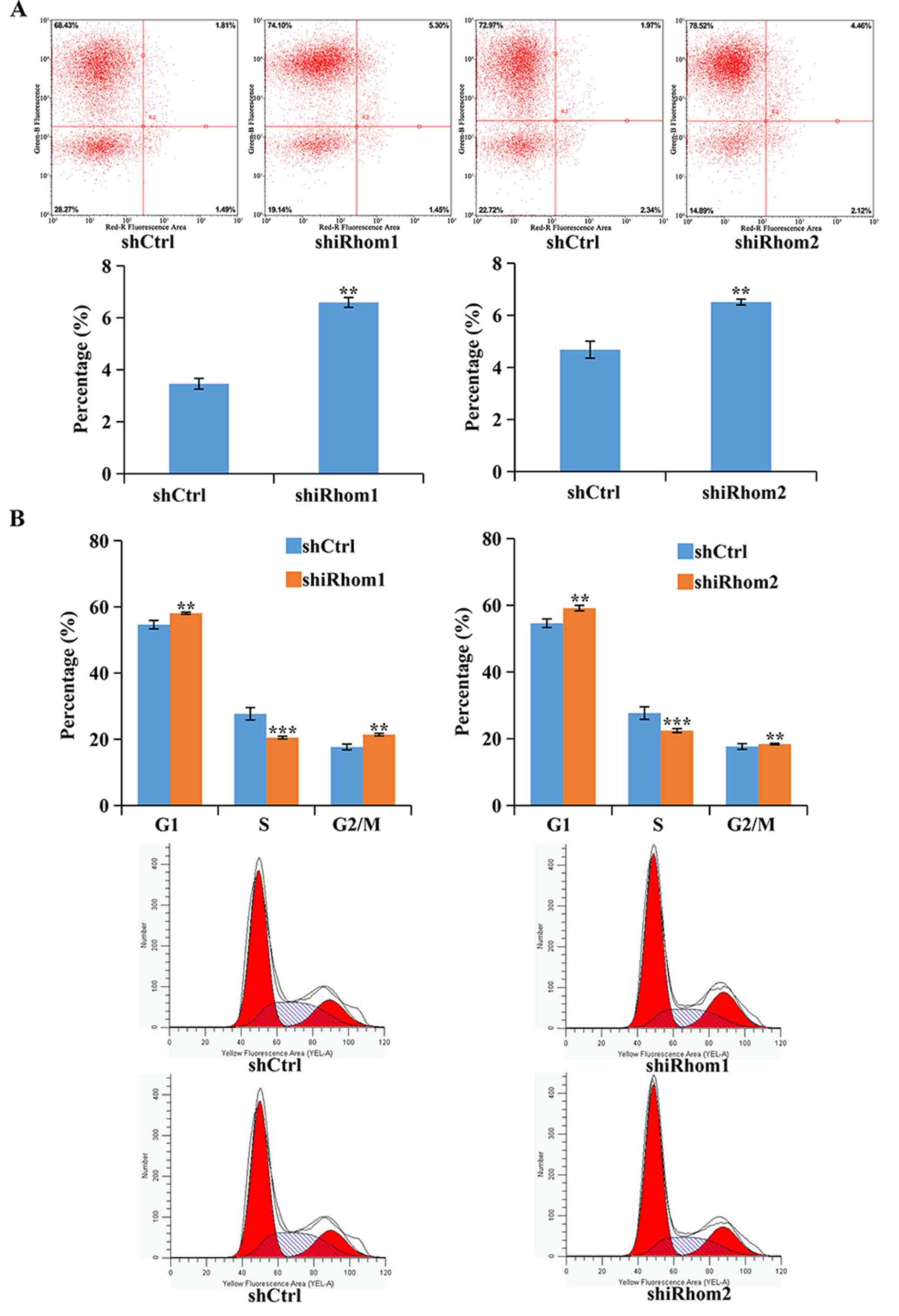
The effect of knockdown of iRhom1 and
iRhom2 was also determined on apoptosis using Annexin V-APC
staining and flow cytometry (Fig.
4A). In these assays, cells in the upper right quadrant
(necrotic) and the lower right quadrant (apoptotic) were apoptotic
(Annexin V positive). The results revealed that knockdown of
iRhom1 or iRhom2 significantly increased the
percentages of cells in apoptosis (Fig.
4A). Dysregulation of the cell cycle in tumors helps to sustain
cell proliferation. Thus, flow cytometry was used to identify the
effects of iRhom1 and iRhom2 knockdown on HeLa cell
cycle progression (Fig. 4B). The
results revealed that downregulation of iRhom1 or
iRhom2 induced cell cycle arrest at the G1 phase, thus,
there was a considerable decrease in the percentage of cells in the
S phase and an increase in the percentage of cells in G1-phase
relative to control cells (Fig. 4B).
However, overexpression of iRhom1 and iRhom2 facilitated cell
proliferation of cervical cancer.
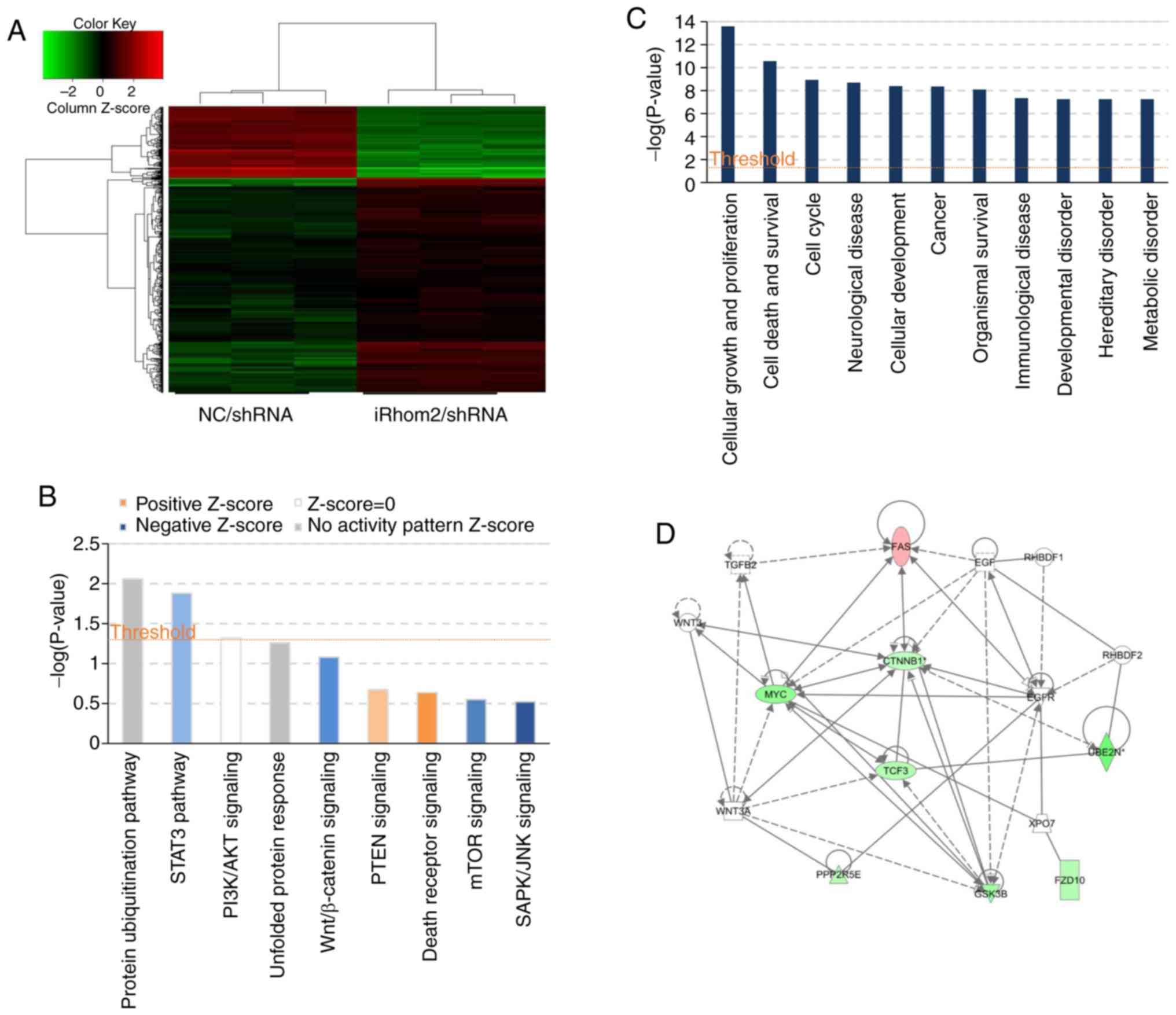
Pathways and genes affected by iRhom2
knockdown
HeLa cells were used infected with
iRhom2/shRNA or NC/shRNA and a microarray platform
for genome-wide gene expression profiling (Fig. 5). The results revealed that iRhom2
knockdown led to significant differences in the expression of 596
genes [P<0.05 for absolute fold change (FC absolute)>1.5],
with 147 upregulated genes and 449 downregulated genes (Fig. 5A). Use of the IPA system for analysis
of the microarray data revealed that iRhom2 knockdown led to
dysregulation of several cellular and molecular functions,
including impairment of cell death and survival, cell growth and
proliferation, cancer, neurological disease, immunological disease,
metabolic disease and cell cycle progression (Fig. 5C). The ‘canonical pathway’ module was
also used for further analysis of the microarray data and
downstream pathways that function in carcinogenesis (STAT3,
PI3K/AKT, Wnt/β-catenin, PTEN, mTOR, unfolded protein response
signaling, Fig. 5B) were identified.
This analysis considers a pathway to be inhibited when the IPA
Z-score is negative. The results indicated significant inhibition
of Wnt/β-catenin signaling, which had an IPA Z-score of −1.134
(Fig. 5D).
Effect of iRhom1 and iRhom2 knockdown
on the Wnt/β-catenin pathway
The present results revealed that expression of
iRhom1 and iRhom2 are linked to carcinogenesis and aggressiveness
of CC cells in vitro, however, the mechanisms of this effect
have not been systematically investigated. The results from the IPA
system indicated that iRhom2 knockdown affected several
proteins that function in Wnt/β-catenin signaling (Table III). This pathway has well
established roles in the migration and invasion of tumor cells.
| Table III.Genes in the Wnt/β-catenin signaling
pathway that are downregulated or upregulated following iRhom2
knockdown in HeLa cells (IPA system). |
Table III.
Genes in the Wnt/β-catenin signaling
pathway that are downregulated or upregulated following iRhom2
knockdown in HeLa cells (IPA system).
| Gene | Regulation | Absolute fold
change |
|---|
| β-catenin | Down | 1.5029672 |
| GSK3B | Down | 2.0011744 |
| Myc | Down | 2.4327161 |
| TGFBR2 | Up | 1.55694 |
| FAS | Up | 1.7245258 |
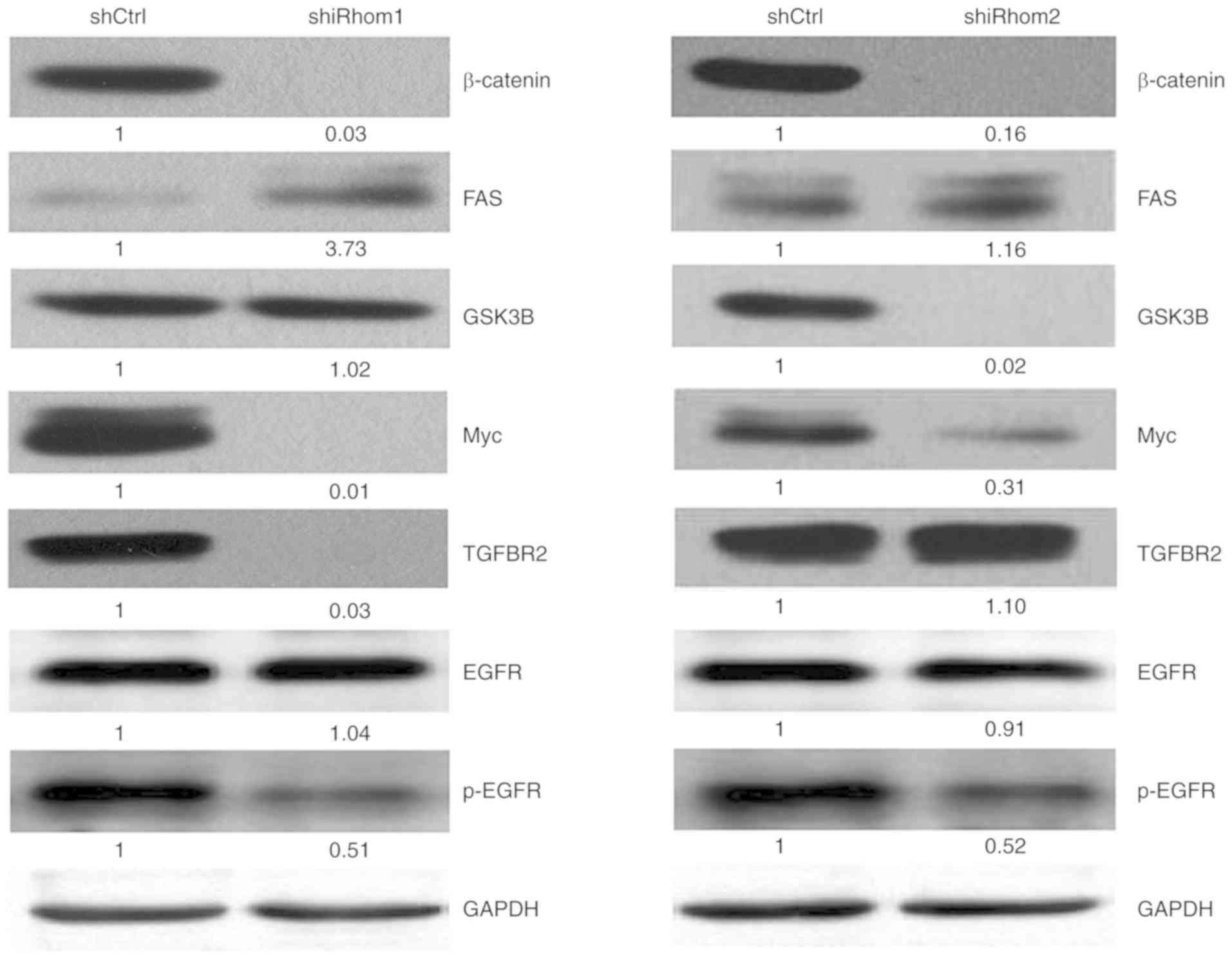
Thus, the effect of iRhom1 and iRhom2
knockdown on the Wnt/β-catenin signaling in HeLa cells was also
determined by assessing the expression of several key proteins in
this pathway (Fig. 6). The western
blotting results indicated that iRhom1 silencing decreased
the expression of β-catenin, Myc, p-EGFR and TGFBR2, and increased
the expression of FAS (P<0.05 for each comparison);
iRhom2 silencing significantly decreased the expression of
β-catenin, GSK3β, p-EGFR and Myc (P<0.05 for each
comparison).
Discussion
CC is a common gynecological cancer. An improved
molecular understanding of its onset and pathogenesis is urgently
required to develop better treatments. The impact of iRhom proteins
on survival from malignant CC has not yet been fully clarified. To
the best of our knowledge, no previous studies have examined the
expression of iRhom proteins in CC, nor examined the prognostic
value of iRhom1 and iRhom2 expression. The present western blot
analyses of CC and adjacent non-malignant normal tissues revealed
that iRhom1 and iRhom2 were upregulated in tumors. Additionally,
the IHC results indicated positive associations of iRhom1, iRhom2,
and Ki-67 expression with cancer stage, tumor size, parametrium
invasion, and patient survival. Multivariate analysis indicated
that cancer stage and iRhom2 expression were independent prognostic
indicators for CC. Previous research reported that expression of
iRhom1 was significantly elevated in clinical specimens of
early-stage breast cancer (13). In
addition, Yuan et al reported that iRhom1 expression was
significantly correlated with tumor size, lymph-node invasion, TNM
stage, and poor prognosis in colorectal cancer (CRC) (7), similar to our findings for CC. Another
study reported that iRhom2 had low or undetectable expression in
most ovarian tumors that were benign or had low malignant potential
(LMP), and in a subset of malignant ovarian tumors (TOV800EPT,
TOV881MT, TOV908DT, TOV974EPT, and TOV1007EPT) (16). These results indicated that expression
of iRhom proteins in tumor cells may be cell-specific.
The present finding of positive correlations of
iRhom1 and iRhom2 expression with several clinicopathological
characteristics suggests that these proteins may have roles in the
onset or progression of CC. Thus, to confirm the roles of iRhom1
and iRhom2 in CC, HeLa cells were used to evaluate the effects of
iRhom1 and iRhom2 knockdown on cell proliferation,
cell cycle progression, and apoptosis. The results indicated that
knockdown of each mRNA inhibited cell proliferation and induced
apoptosis, similar to their effects on CRC (16). To the best of our knowledge, the
present study is the first to quantify the function of
iRhom1 and iRhom2 knockdown on the cell cycle of
cancer cells. Downregulation of each mRNA induced cell cycle arrest
at the G1 phase, and there were therefore fewer cells in the S
phase, and more cells in the G1 phase. These results demonstrated
that iRhom1 and iRhom2 function as oncogenes in CC,
and therefore may have potential as diagnostic and prognostic
biomarkers and as therapeutic targets.
iRhoms1/2 regulated ADAM17-dependent EGFR signaling
during mouse development, suggesting that iRhoms1/2 could emerge as
novel targets for treatment of ADAM17/EGFR-dependent pathologies
(17). Etheridge et al
indicated that mutant iRhom2 proteins present in TOC patients
provide a ‘gain-of-function’, leading to increased ADAM17-mediated
shedding of EGF signaling molecules (including amphiregulin) from
the cell surface (18).
Overexpression of iRhom1 was revealed to coincide with increased
secretion of TGF-α in HeLa cells (5).
The Uev1A-Ubc13 complex with CHIP promoted K63-linked
ubiquitination of iRhom2, increasing its effect on ADAM17
activation, and subsequently blocking TNFα-induced NF-κB signaling
in HeLa cells (19). Our previous
study reported that blockade of ADAM17 decreased the expression of
EMMPRIN, p-EGFR, p-ERK, MMP-2, and MMP-9 proteins in SiHa and HeLa
cells (11). In present study, it was
demonstrated that p-EGFR was downregulated in HeLa cells with
iRhom1 and iRhom2 knockdown, further indicating the activation of
iRhoms1/2-ADAM17-EGFR signaling in cervical cancer. A previous
study also demonstrated that iRhom2 was implicated in epithelial
regeneration and cancer growth through constitutive activation of
epidermal growth factor receptor (EGFR) signaling (20), indicating the positive link between
iRhoms and EGFR signaling (21).
Conversely, Vembar and Brodsky revealed that iRhom1 and iRhom2 bind
to EGF ligands in the ER, downregulate EGFR signaling, and target
these ligands for degradation (22).
ADAM17 was revealed to bind to iRhom2, and activate Notch signaling
in lung cancer stem cells (LCSCs). Moreover, iNOS was revealed to
have an important role in hepatocellular carcinoma due to its
effect on iRhom2/ADAM17 and Notch signaling (23). MET activation in keratinocytes
enhanced Src activity and iRhom expression (particularly
iRhom2), and this led to ADAM17 activation, possibly due to
translocation and maturation of ADAM17 proteolytic activity in
squamous tumors (24). We identified
a critical role of iRhom proteins in CC development and progression
based on studies of clinical samples and HeLa cells. However, the
associated signaling mechanisms remain uncertain.
Genomic microarray analysis of HeLa cells in which
iRhom2 was knocked down was also used. The results indicated
this treatment significantly altered the expression of hundreds of
genes. The IPA data identified multiple pathways that contribute to
the onset and progression of cancer. For instance, knockdown of
iRhom2 enriched the Wnt/β-catenin signaling pathway. This
pathway has been revealed to play a critical role in cell
proliferation, survival, differentiation, and
epithelial-mesenchymal transition (EMT) in cancer cells (25–27).
Studies of canonical Wnt signaling indicated that cytoplasmic
stabilization and nuclear translocation of catenin β1 (CTNNB1), a
regulator of transcription, has a key role (25). However, in the absence of Wnt ligands,
the intracellular levels of β-catenin are low because of the
constitutive activity the ‘β-catenin destruction complex’. This
multiprotein complex consists of two tumor suppressors (APC and
AXIN1), two serine/threonine kinases (CK1 and GSK3) a protein
phosphatase (PP2A), and an E3-ubiquitin ligase (β-TrCP) (28). Wnt/β-catenin signaling plays an
important role in several cancers, including CC (29), lung cancer (30), hepatocellular carcinoma (31), breast cancer (32) and leukemia (33). Although there have not been extensive
studies of Wnt/β-catenin signaling in CC, there is some evidence of
involvement. In particular, previous studies revealed that
epigenetic silencing via hypermethylation of the promoters of
SFRP1, SFRP2, and SFRP4 activated the Wnt pathway,
and promoted cervical adenocarcinoma progression via activation of
EMT (34,35). Another study reported that DKK1 was
downregulated in CC, and this activated the β-catenin pathway
(36). However, it remains unknown
whether iRhom proteins directly or indirectly interact with the
Wnt/β-catenin pathway in CC.
The IPA results revealed that iRhom2
knockdown in HeLa cells altered the expression of five mRNAs
(β-catenin, FAS, Myc, TGFBR2 and GSK3β) that are
involved in Wnt/β-catenin signaling. These results were verified by
western blotting. Thus, the HeLa experiments demonstrated that
iRhom1 silencing significantly decreased the expression of
β-catenin, Myc and TGFBR2, and increased the expression of FAS.
iRhom2 silencing significantly suppressed the expression of
β-catenin, GSK3β, and Myc. A previous study of transgenic mice
reported that upregulated β-catenin-dependent signaling accelerated
carcinogenesis in HPV-mediated CC (37). There is also evidence that
downregulation of a Wnt antagonist (Wnt inhibitory factor 1)
reduced apoptosis and promoted growth, invasion, and angiogenesis
of CC in vivo (38). DAX1 was
revealed to be overexpressed in CC, and promoted cell growth and
tumorigenicity by activating the Wnt/β-catenin pathway via GSK3β
(39). A study on CRC reported that
iRhom1 regulated the activity of APC and stimulated EMT and cell
proliferation, in part via Wnt/β-catenin signaling (39). A study on breast cancer cells reported
that translocation of accumulated β-catenin into the nucleus and
formation of TCF/Lef/β-catenin complexes induced sequential
expression of c-MYC, CCDN1, SNAIL1, and MMP2, and this
increased cell proliferation, migration, and invasion (40). Our previous study of patients with CC
indicated that ADAM17 appears to target matrix metalloproteinase
MMP-2 and MMP-9 through the AREG/EMMPRIN and the EGFR-MEK-ERK
pathway (11).
The HeLa experiments revealed that iRhom1
knockdown decreased the expression of TGFBR2, and increased the
expression of FAS, and that iRhom2 knockdown suppressed the
expression of GSK3β. The reason for the different effects of
iRhom1 and iRhom2 knockdown on Wnt/β-catenin
signaling is unknown. The present study is the first to report
different functions of iRhom1 and iRhom2 in CC. The
downregulation of iRhom proteins could inhibit proliferation,
promote apoptosis maybe in part by altering Wnt/β-catenin signaling
in CC. Our future studies will examine the nature of the
interactions of iRhom proteins with these molecules and their
effect on remodeling of the ECM.
In conclusion, it was revealed that expression of
iRhom1 and iRhom2 were greater in CC tissues than adjacent normal
tissues, and that upregulation of iRhom1 and iRhom2 was correlated
with certain clinical characteristics and with poor OS. Knockdown
of iRhom1 and iRhom2 in HeLa cells inhibited cell
growth, disrupted the cell cycle, and promoted apotosis due to
alterations in the expression of multiple genes and
cancer-associated pathways. Collectively, the present results
demonstrated that iRhom proteins act as oncogenes in CC, and
therefore have potential use as diagnostic and prognostic
biomarkers, as well as therapeutic targets.
Acknowledgements
The authors would like to thank the Department of
Pathology of Fujian Cancer Hospital for their support of the
present study.
Funding
The present study was supported by Grants-in-Aid for
Scientific Research 2015J01379 from the Fujian Natural Science
Foundation, Fujian Province Health Technology Project (No.
2015-ZQN-ZD-8), and the Science and Technology Program of Fujian
Province, China, No. 2018Y2003).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
QX and CC provided substantial contributions to the
conception and design of the work. BL and YL were responsible for
the literature search. YL and CG conducted the clinical studies. DZ
and YX progressed in the experimental studies. JL and LL provided
the data acquisition and carried out the statistical analysis. QX
drafted the work, edited the manuscript and revised it critically
for important intellectual content. All authors critically revised
and approved the final manuscript and agree to be accountable for
all aspects of the work in ensuring that questions related to the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The Ethics Committee of Fujian Provincial Cancer
Hospital which is affiliated with Fujian Medical University,
provided approval of this study. Patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Global Burden of Disease Cancer
Collaboration, ; Fitzmaurice C, Akinyemiju TF, Al Lami FH, Alam T,
Alizadeh-Navaei R, Allen C, Alsharif U, Alvis-Guzman N, Amini E, et
al: Global, regional, and national cancer incidence, mortality,
years of life lost, years lived with disability, and
disability-adjusted life-years for 29 cancer groups, 1990 to 2016:
A systematic analysis for the global burden of disease study. JAMA
Oncol. 4:1553–1568. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim SW, Chun M, Ryu HS, Chang SJ, Kong TW,
Lee EJ, Lee YH and Oh YT: Salvage radiotherapy with or without
concurrent chemotherapy for pelvic recurrence after hysterectomy
alone for early-stage uterine cervical cancer. Strahlenther Onkol.
193:534–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Munro A, Codde J, Spilsbury K, Steel N,
Stewart CJ, Salfinger SG, Tan J, Mohan GR, Leung Y, Semmens JB, et
al: Risk of persistent and recurrent cervical neoplasia following
incidentally detected adenocarcinoma in situ. Am J Obstet Gynecol.
216:272.e1–272.e7. 2017. View Article : Google Scholar
|
|
4
|
Sturtevant MA, Roark M and Bier E: The
Drosophila rhomboid gene mediates the localized formation of wing
veins and interacts genetically with components of the EGF-R
signaling pathway. Genes Dev. 7:961–973. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zou H, Thomas SM, Yan ZW, Grandis JR, Vogt
A and Li LY: Human rhomboid family-1 gene RHBDF1 participates in
GPCR-mediated transactivation of EGFR growth signals in head and
neck squamous cancer cells. FASEB J. 23:425–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li J, Bai TR, Gao S, Zhou Z, Peng XM,
Zhang LS, Dou DL, Zhang ZS and Li LY: Human rhomboid family-1
modulates clathrin coated vesicle-dependent pro-transforming growth
factor α membrane trafficking to promote breast cancer progression.
Ebiomedicine. 36:229–240. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan H, Wei R, Xiao Y, Song Y, Wang J, Yu
H, Fang T, Xu W and Mao S: RHBDF1 regulates APC-mediated
stimulation of the epithelial-to-mesenchymal transition and
proliferation of colorectal cancer cells in part via the
Wnt/β-catenin signalling pathway. Exp Cell Res. 368:24–36. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Adrain C, Zettl M, Christova Y, Taylor N
and Freeman M: Tumor necrosis factor signaling requires iRhom2 to
promote trafficking and activation of TACE. Science. 335:225–228.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McIlwain DR, Lang PA, Maretzky T, Hamada
K, Ohishi K, Maney SK, Berger T, Murthy A, Duncan G, Xu HC, et al:
iRhom2 regulation of TACE controls TNF-mediated protection against
Listeria and responses to LPS. Science. 335:229–232. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blaydon DC, Etheridge SL, Risk JM, Hennies
HC, Gay LJ, Carroll R, Plagnol V, McRonald FE, Stevens HP, Spurr
NK, et al: RHBDF2 mutations are associated with tylosis, a familial
esophageal cancer syndrome. Am J Hum Genet. 90:340–346. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu Q, Ying M, Chen G, Lin A, Xie Y, Ohara
N and Zhou D: ADAM17 is associated with EMMPRIN and predicts poor
prognosis in patients with uterine cervical carcinoma. Tumour Biol.
35:7575–7586. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zettl M, Adrain C, Strisovsky K, Lastun V
and Freeman M: Rhomboid family pseudoproteases use the ER quality
control machinery to regulate intercellular signaling. Cell.
145:79–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yan Z, Zou H, Tian F, Grandis JR, Mixson
AJ, Lu PY and Li LY: Human rhomboid family-1 gene silencing causes
apoptosis or autophagy to epithelial cancer cells and inhibits
xenograft tumor growth. Mol Cancer Ther. 7:1355–1364. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arcidiacono P, Webb CM, Brooke MA, Zhou H,
Delaney PJ, Ng KE, Blaydon DC, Tinker A, Kelsell DP and Chikh A:
p63 is a key regulator of iRHOM2 signalling in the keratinocyte
stress response. Nat Commun. 9:10212018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ishimoto T, Miyake K, Nandi T, Yashiro M,
Onishi N, Huang KK, Lin SJ, Kalpana R, Tay ST, Suzuki Y, et al:
Activation of transforming growth factor beta 1 signaling in
gastric cancer-associated fibroblasts increases their motility, via
expression of rhomboid 5 homolog 2, and ability to induce
invasiveness of gastric cancer cells. Gastroenterology.
153:191–204.e16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wojnarowicz PM, Provencher DM, Mes-Masson
AM and Tonin PN: Chromosome 17q25 genes, RHBDF2 and CYGB, in
ovarian cancer. Int J Oncol. 40:1865–1880. 2012.PubMed/NCBI
|
|
17
|
Li X, Maretzky T, Weskamp G, Monette S,
Qing X, Issuree PD, Crawford HC, McIlwain DR, Mak TW, Salmon JE and
Blobel CP: iRhoms 1 and 2 are essential upstream regulators of
ADAM17-dependent EGFR signaling. Proc Natl Acad Sci USA.
112:6080–6085. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Etheridge SL, Brooke MA, Kelsell DP and
Blaydon DC: Rhomboid proteins: A role in keratinocyte proliferation
and cancer. Tissue Cell Res. 351:301–307. 2013. View Article : Google Scholar
|
|
19
|
Zhang Y, Li Y, Yang X, Wang J, Wang R,
Qian X, Zhang W and Xiao W: Uev1A-Ubc13 catalyzes K63-linked
ubiquitination of RHBDF2 to promote TACE maturation. Cell Signal.
42:155–164. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hosur V, Johnson KR, Burzenski LM, Stearns
TM, Maser RS and Shultz LD: Rhbdf2 mutations increase its protein
stability and drive EGFR hyperactivation through enhanced secretion
of amphiregulin. Proc Natl Acad Sci USA. 111:E2200–E2209. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Siggs OM, Grieve A, Xu H, Bambrough P,
Christova Y and Freeman M: Genetic interaction implicates iRhom2 in
the regulation of EGF receptor signalling in mice. Biol Open.
3:1151–1157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vembar SS and Brodsky JL: One step at a
time: Endoplasmic reticulum-associated degradation. Nat Rev Mol
Cell Biol. 9:944–957. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang R, Li Y, Tsung A, Huang H, Du Q, Yang
M, Deng M, Xiong S, Wang X, Zhang L, et al: iNOS promotes
CD24+CD133+ liver cancer stem cell phenotype
through a TACE/ADAM17-dependent Notch signaling pathway. Proc Natl
Acad Sci USA. 115:E10127–E10136. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cataisson C, Michalowski AM, Shibuya K,
Ryscavage A, Klosterman M, Wright L, Dubois W, Liu F, Zhuang A,
Rodrigues KB, et al: MET signaling in keratinocytes activates EGFR
and initiates squamous carcinogenesis. Sci Signal. 9:ra622016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McCrea PD and Gottardi CJ: Beyond
β-catenin: Prospects for a larger catenin network in the nucleus.
Nat Rev Mol Cell Biol. 17:55–64. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stamos JL and Weis WI: The β-catenin
destruction complex. Cold Spring Harb Perspect Biol. 5:a0078982013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ueda T, Tsubamoto H, Inoue K, Sakata K,
Shibahara H and Sonoda T: Itraconazole modulates hedgehog,
WNT/β-catenin, as well as Akt signalling, and inhibits
proliferation of cervical cancer cells. Anticancer Res.
37:3521–3526. 2017.PubMed/NCBI
|
|
30
|
Mazieres J, He B, You L, Xu Z, Lee AY,
Mikami I, Reguart N, Rosell R, McCormick F and Jablons DM: Wnt
inhibitory factor-1 is silenced by promoter hypermethylation in
human lung cancer. Cancer Res. 64:4717–4720. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Takigawa Y and Brown AM: Wnt signaling in
liver cancer. Curr Drug Targets. 9:1013–1024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Khramtsov AI, Khramtsova GF, Tretiakova M,
Huo D, Olopade OI and Goss KH: Wnt/beta-catenin pathway activation
is enriched in basal-like breast cancers and predicts poor outcome.
Am J Pathol. 176:2911–2920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu D, Zhao Y, Tawatao R, Cottam HB, Sen M,
Leoni LM, Kipps TJ, Corr M and Carson DA: Activation of the Wnt
signaling pathway in chronic lymphocytic leukemia. Proc Natl Acad
Sci USA. 101:3118–3123. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chung MT, Lai HC, Sytwu HK, Yan MD, Shih
YL, Chang CC, Yu MH, Liu HS, Chu DW and Lin YW: SFRP1 and SFRP2
suppress the transformation and invasion abilities of cervical
cancer cells through Wnt signal pathway. Gynecol Oncol.
112:646–653. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin YW, Chung MT, Lai HC, De Yan M, Shih
YL, Chang CC and Yu MH: Methylation analysis of SFRP genes family
in cervical adenocarcinoma. J Cancer Res Clin Oncol. 135:1665–1674.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee J, Yoon YS and Chung JH: Epigenetic
silencing of the WNT antagonist DICKKOPF-1 in cervical cancer cell
lines. Gynecol Oncol. 109:270–274. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bulut G, Fallen S, Beauchamp EM, Drebing
LE, Sun J, Berry DL, Kallakury B, Crum CP, Toretsky JA, Schlegel R
and Üren A: Beta-catenin accelerates human papilloma virus type-16
mediated cervical carcinogenesis in transgenic mice. PLoS One.
6:e272432011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ramachandran I, Thavathiru E, Ramalingam
S, Natarajan G, Mills WK, Benbrook DM, Zuna R, Lightfoot S, Reis A,
Anant S and Queimado L: Wnt inhibitory factor 1 induces apoptosis
and inhibits cervical cancer growth, invasion and angiogenesis in
vivo. Oncogene. 31:2725–2737. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu XF, Li XY, Zheng PS and Yang WT: DAX1
promotes cervical cancer cell growth and tumorigenicity through
activation of Wnt/β-catenin pathway via GSK3β. Cell Death Dis.
9:3392018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Perez-Yepez EA, Ayala-Sumuano JT, Lezama R
and Meza I: A novel β-catenin signaling pathway activated by IL-1β
leads to the onset of epithelial-mesenchymal transition in breast
cancer cells. Cancer Lett. 354:164–171. 2014. View Article : Google Scholar : PubMed/NCBI
|