Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common malignant tumor worldwide and the third most common cause of
cancer-related death (1).
Unfortunately, HCC tumors are aggressive, difficult to detect and
exhibit a complex pathology, which makes them highly refractory in
nature. While less than 30% of patients with HCC are suitable for
resection and orthotopic liver transplantation, which is commonly
accepted as the most effective treatment (2), advanced HCC cases lack adequate
therapeutic treatments. Moreover, high rates of metastasis and
relapse limit the long-term efficacy of surgery (3) and HCC frequently demonstrates
resistance to systemic chemotherapy, with the National
Comprehensive Cancer Network guidelines recommending that
chemotherapeutics are not prescribed for the treatment of HCC in
clinical practice (4). Although
sorafenib has demonstrated potential as a drug for advanced liver
cancer in the SHARP and START (5–7)
studies, its application in advanced HCC is limited by poor
responses and various side effects (8). Similarly, radiotherapy in HCC is also
limited due to the radiation dosage and tumor positioning (9–11).
Currently, the most commonly used non-surgical therapies for liver
cancer are transarterial chemoembolization and transarterial
embolization. Both treatments significantly improve the prognosis
of patients with multiple lesions and a large tumor mass whose
livers are not tolerant to surgery (12,13);
however, they are often associated with incomplete embolization,
the formation of collateral circulation and liver function damage
following repeated procedures (14). Additionally, they do not effectively
control distant metastasis rates. Thus, there is unanimous
agreement for the urgent requirement to develop novel, more
effective drugs and treatment methods for the clinical treatment of
HCC (5,15). One suggested approach is to identify
drugs that are derived from natural compounds with anticancer
activity.
Musk and its preparations have been used as
traditional analgesics for treating pain resulting from various
types of cancer, including liver cancer (16); it is particularly popular among the
Tibetans in Sichuan, China, which due to their health philosophy,
receive no surgical intervention or non-surgical treatments. Over
the past decade, follow-up studies in dozens of patients with
advanced liver cancer from the Tibetan region observed that the use
of musk as an analgesic increased their quality of life and
lifespan, suggesting that the ingredients within musk may inhibit
liver cancer progression in addition to their analgesic
properties.
Since these observations, modern basic and clinical
pharmacological studies have confirmed that musk and its
preparations are effective in the treatment of central nervous
system and cardiovascular/cerebrovascular diseases, as demonstrated
in a previous study of the Shexiang Baoxin pill (17). Previous studies have investigated
the antitumor properties of musk, including Shengjizonglu and
TaipingHuiminHejijuFang (18), and
musk and muscone were found to inhibit cell growth in Ehrlich
ascites carcinoma, sarcoma −180 and HCC (19). Clinical studies have also reported
that musk-containing compound drugs, such as Xihuangwan,
Xiaojindan, and Liushenwan demonstrated certain anticancer effects
(20). The active anticancer
ingredients of musk are thought to include the macrocyclic ketones,
of which 3-methylcyclohexanone (muscone) is the major one (21), and synthetic dextral muscone has
demonstrated activities similar to natural sinistral muscone
(22). However, the antitumor
mechanisms of musk/muscone in liver cancer are unclear.
Based on the clinical observations that have
suggested that muscone exhibits anticancer properties, the present
study aimed to investigate whether it could be used as an
anti-liver cancer drug for the non-surgical treatment of advanced
liver cancer. The inhibitory effects of synthetic muscone were
tested on various liver cancer cell lines, and its target sites and
mechanisms of action were also explored. In addition, the in
vitro findings were validated in vivo, in nude mice with
transplanted tumors, as well as in clinical HCC specimens.
Materials and methods
Reagents and antibodies
Muscone was obtained from Chengdu Kangfei Co., Ltd.
and was dissolved in DMSO (Amresco, LLC); and 4-phenylbutyrate
(4-PBA) was purchased from Sigma-Aldrich; Merck KGaA. Primary
antibodies against phosphorylated (p)-protein kinase R-like
endoplasmic reticulum kinase (PERK) (dilution 1:1,000; cat. no.
SAB4301310), p-eukaryotic initiation factor (eIF)2α (dilution
1:1,000; cat. no. SAB4504388), p-AMP kinase (p-AMPK) (dilution
1:1,000; cat. no. SAB4503754) and p-mTOR (dilution 1:1,000; cat.
no. SAB4504282) were purchased from Sigma-Aldrich; Merck KGaA; the
primary antibody against light chain 3B (LC3B; dilution 1:1,000;
cat. no. 2775) was purchased from Cell Signaling Technology, Inc.;
antibodies against Bcl-2 (dilution 1:500; cat. no. sc-7382), pro
caspase-3 (dilution 1:500; cat. no. sc-7272), AMPK (dilution 1:500;
cat. no. sc-74461), Bax (dilution 1:500; cat. no. sc-7480) and mTOR
(dilution 1:500; cat. no. sc-517464) were purchased from Santa Cruz
Biotechnology, Inc.; anti-3-methyladenine (3-MA) primary antibodies
were purchased from Selleck Chemicals; the anti-activating
transcription factor (ATF)4 (dilution 1:500; cat. no. AF2560) and
anti-DNA damage inducible transcript (DDIT) 3 (dilution 1:500; cat.
no. AF6684) antibodies were purchased from Beyotime Institute of
Biotechnology; and the cleaved caspase-3 (dilution 1:1,000; cat.
no. 19677-1-AP), anti-sestrin (SESN)2 (dilution 1:1,000; cat. no.
10795-1-AP), anti-β-actin primary antibodies (dilution 1:5,000;
cat. no. 66009-1-Ig), horseradish peroxidase (HRP)-conjugated
secondary antibodies including HRP-conjugated Affinipure goat
anti-mouse IgG (H+L) (dilution 1:10,000; cat. no. SA00001-1),
HRP-conjugated Affinipure goat anti-rabbit IgG (H+L) (dilution
1:10,000; cat. no. SA00001-2) and HRP-conjugated Affinipure rabbit
anti-goat IgG (H+L) (dilution 1:10,000; cat. no. SA00001-4) were
purchased from ProteinTech Group, Inc.
Cell culture and small interfering RNA
(siRNA) transfection
Human hepatoma/liver cancer HepG2 cells, Hep3B
cells, human lung carcinoma A549 cells and human colorectal cancer
HCT116 cells were purchased from The Cell Bank of the Type Culture
Collection of the Chinese Academy of Sciences and were validated
using short tandem repeat DNA profiling. HepG2, Hep3B and A549
cells were cultured in RPMI-1640 medium (Logan), and HCT116 cells
were cultured in DMEM (Logan). Both media were supplemented with
100 U/ml penicillin, 100 µg/ml streptomycin and 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and cells were cultured in a
humidified atmosphere at 37°C in 5% CO2.
siRNA against SENS2 (5′-GACCAUGGCUACUCGCUGATT-3′)
and the negative control (NC; 5′-UUCUCCGAACGUGUCACGUTT-3′) were
purchased from Genepharm (Shanghai, China). All siRNAs were
subjected to BLAST search to ensure the absence of hits with >17
nucleotide matches found in the corresponding genomes. Transfection
was carried out using 100 pmol of siRNA and
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
medium was changed after 6 h.
Flow cytometric analysis of
apoptosis
Following treatment with DMSO or muscone for 24 h,
the apoptotic nuclear morphology of HepG2 cells was observed under
a fluorescence microscope at an ×40 magnification (Olympus IX71)
with 2 µg/ml DAPI staining (Sigma-Aldrich; Merck KGaA) for 20 min
at room temperature. Cells were subsequently double stained with an
Annexin V-FITC/propidium iodide (PI) kit (Nanjing KeyGen Biotech
Co., Ltd.), according to the manufacturer's protocol. Apoptotic
cells were subsequently analyzed using a flow cytometer (BD
Pharmingen; BD Biosciences). The data were analyzed using FlowJo
software (version 7.6.1; Tree Star, Inc., Ashland, OR, USA).
Hoechst 33342 staining
Cells were plated into 6-well plates at a density of
1×105 cells per well and pretreated with 1 mM 3-MA
(Selleck Chemicals) for 2 h, prior to treatment with muscone. Cells
were rinsed three times with PBS and then stained with 10 µg/ml
Hoechst 33342 (Sigma-Aldrich; Merck KGaA) for 15 min at 37°C before
being washed three times with PBS for a second time. Stained cells
were observed under a fluorescence microscope at an ×200
magnification (Olympus IX71). Apoptotic cells were identified by
the fragmentation of nuclei and condensation of chromatin, and the
apoptotic index was calculated using the following equation: Number
of apoptotic cells/total cell count. Experiments were performed in
triplicate.
Autophagy assay
Following incubation with DMSO or muscone at 37°C
for 24 h, 3×104 cells/well were stained with
monodansylcadaverine (MDC, 0.1 mmol/l; Nanjing KeyGen Biotech Co.,
Ltd.) for 25 min at 37°C and observed using a fluorescence
microscope (Olympus IX71) at an ×100 magnification. The
fluorescence intensity was analyzed using a Varioskan Flash
fluorescence microplate reader (Thermo Fisher Scientific,
Inc.).
To examine the formation of autophagosomes, a total
of 3×104 cells/well were plated into 48-well plates and
~24 h later, the cells were rinsed three times PBS. Subsequently,
cells were treated with 1 mg/ml acridine orange (AO; Nanjing KeyGen
Biotech Co., Ltd.) for 30 min at 37°C, and then rinsed again three
times with PBS. Stained cells were visualized using a fluorescence
microscope at an ×100 magnification (Olympus IX71) and the
percentage of acidic vesicular organelle (AVO)-positive cells with
≥10 AVO aggregations was calculated and analyzed statistically
(≥600 cells were observed). Experiments were performed in
triplicate.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and
subsequently treated with DNase I (Thermo Fisher Scientific, Inc.).
Total RNA was reverse transcribed into cDNA using random hexamer
primers and RevertAid Reverse Transcriptase (Thermo Fisher
Scientific, Inc.). qPCR was subsequently performed using a
SYBR® Premix Ex Taq II kit (Takara Biotechnology Co.,
Ltd.) and a StepOnePlus Real-time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The following primer pairs were
used for the qPCR: AFT4 forward, 5′-GTCAGTCCCTCCAACAACAGCAA-3′ and
reverse, 5′-GAAGGTCATCTGGCATGGTTTCC-3′; DDIT3 forward,
5′-TTGCCTTTCTCCTTCGGGACACT-3′ and reverse,
5′-CTTGTGACCTCTGCTGGTTCTGG; FGF21 forward,
5′-CTGGAGATCAGGGAGGATGGGA-3′ and reverse,
5′-FCGTGGGCTTCGGACTGGTAAA-3′; TRIB3 forward,
5′-AAGCGGTTGGAGTTGGATGAC-3′ and reverse,
5′-CACGATCTGGAGCAGTAGGTG-3′; SESN2 forward,
5′-AAGGACTACCTGCGGTTCG-3′ and reverse, 5′-CGCCCAGAGGACATCAGTG-3′;
and GAPDH forward, 5′-CCATGTTCGTCATGGGTGTGAACCA-3′ and reverse,
5′-GCCAGTAGAGGCAGGGATGATGTTC-3′. Expression levels were quantified
using the 2−ΔΔCq method (23) and GAPDH was used as the internal
reference control. The thermocycling conditions were as follows:
95°C for 3 min, followed by 40 cycles of 95°C for 15 sec, 60°C for
15 sec, 72°C for 20 sec; 95°C, 15 sec; 60°C, 1 min; 95°C, 15 sec;
60°C, 15 sec. Each sample was determined in duplicate. All PCR
products were confirmed by 2.0% agarose gel electrophoresis.
Whole transcriptome library
preparation and sequencing
Novogene Bioinformatics Technology Cooperation
prepared the whole transcriptome libraries and performed deep
sequencing. Following incubation with DMSO or muscone for 24 h,
total RNA from HepG2 cells was extracted. Total RNA was quantified
using a K5500 micro-spectrophotometer (Drawell International
Technology Co., Ltd.) and the quality was determined using an
Agilent 2200 TapeStation system (Agilent Technologies, Inc.).
Ribosomal RNA was removed following the generation of
strand-specific sequencing libraries, according to the
manufacturer's protocol (Novogene Co., Ltd.). RNA sequencing
(RNA-Seq) was performed using an Illumina HiSeq 2000 platform and
100 bp paired-end reads were generated according to the
manufacturer's protocol (Illumina, Inc.).
RNA-Seq data analysis
Raw image data obtained from high-throughput
sequencing (Illumina HiSeqTMPE125/PE150) were processed by CASAVA
(version 1.8.2; Illumina, Inc.) for base calling and sequenced
reads were obtained. Raw data were stored as FASTQ files, and raw
reads with adapters and low-quality portions were filtered to
obtain clean reads for subsequent analysis. TopHat2 (version 2.1.1)
was used to map the reads to the reference genome. During RNA-Seq
analysis, expression levels were determined by counting the reads
that were mapped to genomic regions or exons. Subsequently,
fragments/kilobases/million reads were used for differential
expression analysis. Pathway enrichment associated with
differentially expressed genes (DEGs) was performed using
hypergeometric analysis of corresponding Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathways (http://www.genome.jp/kegg/ or http://www.kegg.jp/).
Western blotting
Cells were lysed using NP-40 lysis buffer (50 mM
Tris-HCl, pH 7.4, 150 mM NaCl, 1% NP-40, 0.5% NaDC, 0.1% SDS)
containing a protease inhibitor cocktail proteinase inhibitor
cocktail solution (Roche, Basel, Switzerland). Total protein in
cell lysates was quantified using a bicinchoninic acid protein
assay kit (Thermo Fisher Scientific, Inc.). Equal amounts (30 µg)
of protein were separated by 10% SDS-PAGE. Separated proteins were
subsequently transferred onto nitrocellulose membranes and blocked
with PBS-0.2% Tween-20 (PBST) containing 5% skimmed milk powder
(SMP) at room temperature for 1 h. The membranes were incubated
overnight at 4°C with primary antibodies in PBST containing 5% SMP.
The membranes were subsequently washed five times with PBST and
incubated with secondary antibodies (1:10,000) in PBST containing
5% SMP at room temperature for 1 h. Membranes were washed five
times with PBST and protein bands were visualized on X-ray film
(Carestream Health, Inc.) using ECL chemiluminescence reagents
(Thermo Fisher Scientific, Inc.), with β-actin as the loading
control. Protein expression was quantified using ImageJ 1.51 k
software (National Institutes of Health).
Cell viability assay
Cells (1×104) were incubated with muscone
(concentrations of 0.5, 1, 1.5, 2 and 2.5 µM) or a corresponding
volume of DMSO for 24 h, and cell viability was measured using a
Cell Counting Kit-8 assay (CCK-8; Nanjing KeyGen Biotech Co.,
Ltd.), according to the manufacturer's protocol. Brielfly, 10 µl of
CCK-8 solution was added to each well followed by 1 h of incubation
at 37°C. Subsequently, the absorbance value (OD) of the cells at
450 nm was measured by using a universal microplate
spectrophotometer (BioTek Instruments, Inc.).
Animal studies
All animal studies were approved by the Committee on
the Use of Live Animals in Teaching and Research of Sichuan
University, and were conducted in accordance with approved
guidelines. BALB/c nude mice (number, 20; age, 6 weeks; sex, male;
weighing approximately 18–22 g) were obtained from Chengdu Dossy
Biological Technology Company (http://www.cd-dossy.cn/intro/50.html) (Chengdu, China;
certificate no: SCXK Sichuan 2015-030). All mice were maintained
under special pathogen-free conditions, with temperature maintained
at 23±2°C, humidity of 60±10%, 12 h light-dark cycle with free
access to standard rodent chow and water. A total of
3×106 HepG2 cells were resuspended in 0.2 ml serum-free
DMEM with 50% Matrigel (Corning, Inc.) subcutaneously injected into
the lateral flank of mice and 14 days later, mice with an average
tumor diameter of 6 mm were selected for the intraperitoneal
injection of 0.1 mmol/kg, 0.2 ml/20 g bw DMSO or muscone. Maximum
tumor diameter exceeding 15 mm was deemed the humane endpoint of
the study. The mice were sacrificed by CO2 exposure
followed by cervical dislocation and efforts were made to minimize
suffering. Then xenograft tumors were collected and measured.
Patient studies
The present study was approved by the Institutional
Ethics Committee of the Changchun University of Traditional Chinese
Medicine (approval no. CCZYFYCC2017-071) and adhered to the
principles in the Declaration of Helsinki. Informed consent was
obtained from each patient prior to tissue collection for
experimentation. A total of 14 human HCC tissue samples were
obtained from the Affiliated Hospital of Changchun University of
Chinese Medicine (age range, 40–55 years; 10 males and 4 females;
collected from March 2016 to March 2017). All tissues were
collected from patients who underwent surgery at the hospital. The
patients were diagnosed with HCC based on pathology and all
patients had no history of anticancer therapy. All of the specimens
were examined and evaluated by two independent pathologists. The
tissues were snap-frozen and stored in liquid nitrogen immediately
after dissection until required for further analysis using western
blotting and RT-qPCR.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 6 software (GraphPad Software, Inc.) and based on
three independent experiments; data are expressed as the mean ± SD.
Student's t-tests was used to determine statistical differences
between groups. Differences among three or more groups were
compared using one-way ANOVA followed by a Tukey's test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Muscone inhibits the growth of liver
cancer cells
To investigate the effects of muscone on the cell
viability of liver cancer cells, HepG2 and Hep3B cells were treated
with muscone (0.5, 1, 1.5, 2 and 2.5 µM) for 24 h. Results from the
CCK-8 assay demonstrated that the viability of cells treated with
muscone was significantly decreased compared with cells treated
with DMSO, with muscone exhibiting a dose-dependent effect on cell
viability (Fig. 1A and C). HepG2
cells were observed to be more sensitive to muscone treatment
(Fig. 1A and C), with a half
maximal inhibitory concentration (IC50) value of 0.663
µM, whereas the Hep3B IC50 value was 1.416 µM. Whilst
the corresponding volume of DMSO had no effect on cell growth,
0.663 µM muscone significantly inhibited cell growth following 24
and 48 h of treatment (Fig. 1B and
D). Thus, 0.663 µM muscone and the corresponding volume of DMSO
were used in the following HepG2 cell experiments. To further
identify whether muscone treatment affected other malignant tumors,
1 µM muscone was used to treat malignant tumor cell lines A549 and
HCT116, and was found to significantly inhibit the cell viability
compared with the control group (Fig.
1E and F).
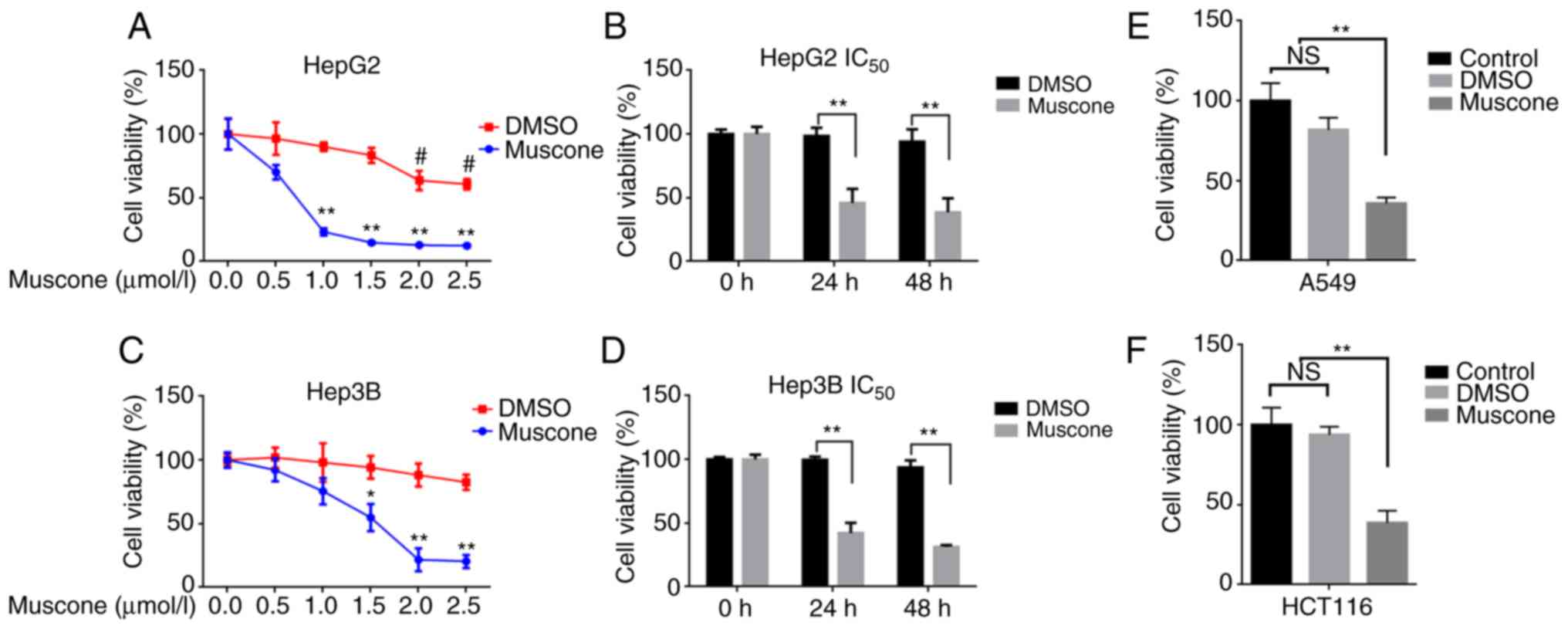 | Figure 1.Muscone inhibits tumor cell
viability. (A) Cell viability and IC50 of HepG2 cells
treated with 0, 0.5, 1, 1.5, 2 or 2.5 µMol/l muscone or DMSO for 24
h were determined using CCK-8 assays. (B) Cell viability of HepG2
cells was determined using CCK-8 assays following treatment with
0.663 µM muscone or the corresponding volume of DMSO for 24 or 48
h. (C) Cell viability and IC50 of Hep3B cells treated
with 0, 0.5, 1, 1.5, 2 or 2.5 µMol/l muscone or DMSO for 24 h was
determined using CCK-8 assays. (D) Cell viability of Hep3B cells
was determined using CCK-8 assays following treatment with 0.663 µM
muscone or the corresponding volume of DMSO for 24 or 48 h. (E and
F) Cell viability of (E) A549 and (F) HCT116 cells treated with 1
µM muscone or DMSO for 24 h was assessed using a CCK-8 assay.
Control represents cells that did not receive treatment.
*P<0.05, **P<0.01 vs. Control group; #P<0.05
vs. muscone group; n=3; NS, not significant; IC50, half
maximal inhibitory concentration; CCK-8, Cell Counting Kit-8. |
Muscone induces HepG2 cell apoptosis
and autophagy
Next, it was investigated whether the effect of
muscone treatment on the survival of HepG2 cells was associated
with apoptosis. HepG2 cells treated with muscone and stained with
DAPI demonstrated an apoptotic phenotype (Fig. 2A). Cells were subsequently subjected
to double staining with Annexin V-FITC/PI and examined by flow
cytometry. The apoptotic rate of HepG2 cells was significantly
increased following treatment with muscone compared with the
control (Fig. 2B). Apoptosis is
induced following the cleavage and subsequent activation of
pro-caspase-3 (24–26), as caspase-3 is an important
regulator of apoptosis in the downstream caspase cascade reaction
(27,28). Bcl-2 and Bax proteins belonging to
the Bcl-2 family are also important regulators of apoptosis
(29), thus the expression levels
of Bax, Bcl-2 and caspase-3 proteins were analyzed using western
blotting (Fig. 2C). Following
treatment of HepG2 cells with muscone, the expression levels of
cleaved caspase-3 and Bax proteins were markedly increased, whereas
those of the Bcl-2 protein were decreased compared with the
control.
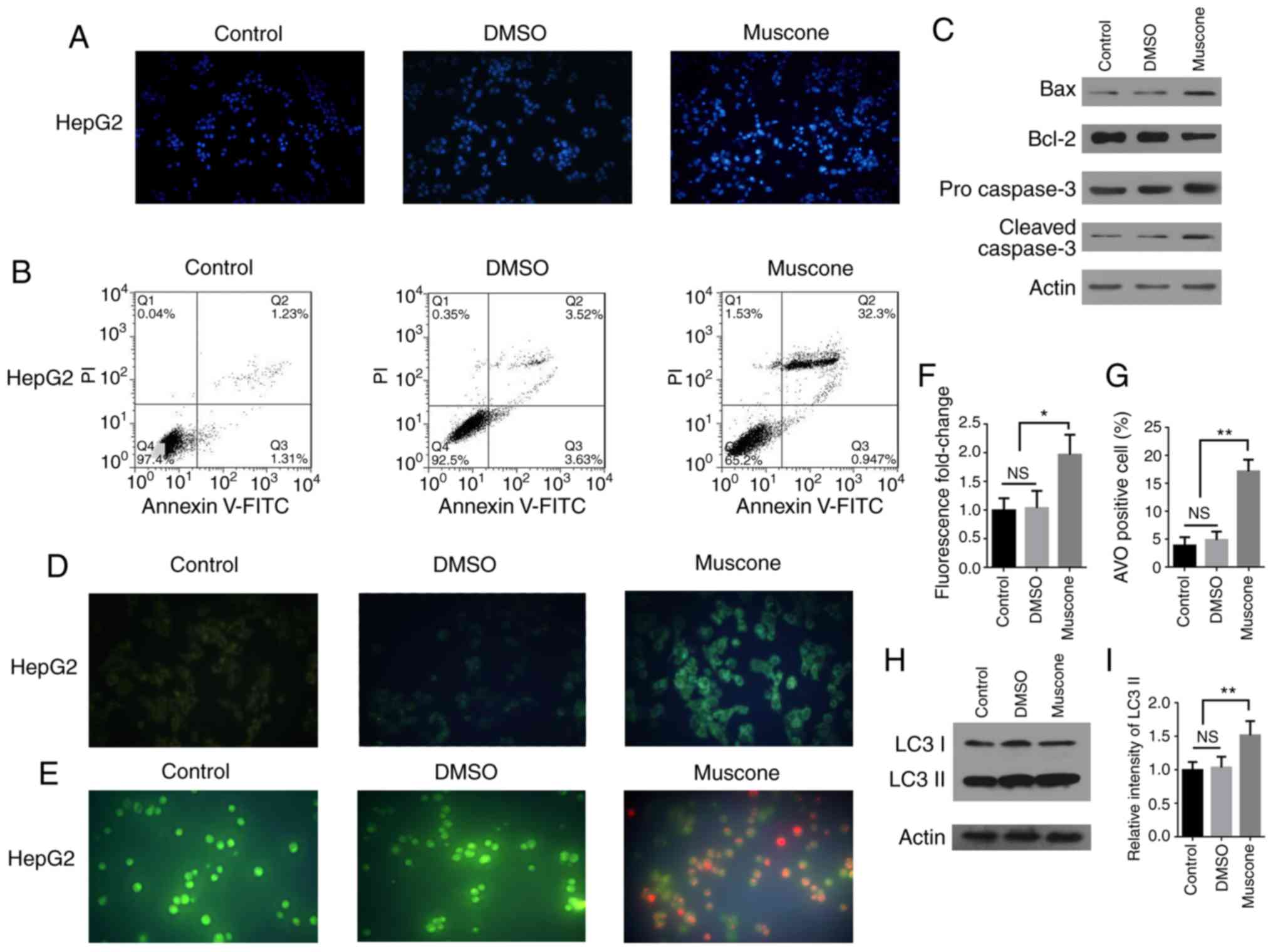 | Figure 2.Muscone induces apoptosis and
autophagy in HepG2 cells. (A) HepG2 cells were treated with DMSO or
muscone for 24 h, and then stained with DAPI and examined by
fluorescence microscopy at an ×40 magnification (Olympus IX71).
Blue staining represents the cell nucleus and the control
represents cells that did not receive treatment. (B) HepG2 cells
were treated with DMSO or muscone for 24 h and subsequently double
stained with Annexin V-FITC/PI. Apoptosis was analyzed using flow
cytometry. (C) HepG2 cells were treated with DMSO or muscone for 24
h and the protein expression levels of Bax, Bcl-2, pro-caspase-3
and cleaved caspase-3 were analyzed using western blotting. (D and
E) Representative micrographs (×100 magnification, Olympus IX71) of
(D) MDC and (E) AO staining following treatment of HepG2 cells with
DMSO or muscone for 24 h. (F) MDC fluorescence intensity
fold-change was semi-quantified using a fluorescence microplate
reader. (G) Positive AO staining and cell counting. (H) HepG2 cells
were treated with DMSO or muscone for 24 h and LC3 I and LC3 II
expression levels were analyzed using western blotting. (I) LC3 II
levels were quantified by densitometric analysis relative to LC3 I.
Error bars represent the standard deviations for n=3. *P<0.05,
**P<0.01; n=3; NS, not significant; PI, propidium iodide; MDC,
monodansylcadaverine; AO, acridine orange; LC3, light chain 3. |
To investigate the effect of muscone on autophagy
within HepG2 cells, AVOs were visualized using MDC staining and AO
staining (Fig. 2D and E) (30). Muscone treatment significantly
increased the percentage of AVO-positive HepG2 cells, which was
demonstrated using fluorescence analysis (Fig. 2F) and cell counts (Fig. 2G). Moreover, western blotting was
used to analyze the expression levels of microtubule-associated
proteins 1A/1B light chain 3 (LC3) II protein, the lipidated form
of LC3 associated with the autophagosome membrane (31), in the HepG2 cells and it was
demonstrated that LC3 II expression levels were markedly increased
following muscone treatment (Fig. 2H
and I). These results suggest that muscone treatment may induce
apoptosis and autophagy in HepG2 cells.
Muscone-induced HepG2 cell apoptosis
is associated with endoplasmic reticulum (ER) stress through the
PERK/ATF4/DDIT3 signaling pathway
To determine the molecular mechanism used by muscone
to induce apoptosis, transcriptome sequencing analysis was
performed in HepG2 cells treated with muscone. The differentially
expressed genes (DEGs) [|log2 (fold-change|)>1 and q-value
<0.005] were screened according to the two threshold values:
Fold-change and significance level. A total of 170 genes
demonstrated differential expression following muscone treatment,
of which 131 genes had increased expression levels and 39 genes
were found to have reduced expression levels (Fig. 3A). Statistical analysis of these
data indicated that genes associated with ER stress, such as ATF4,
DDIT3, TIRB3 and FGF21 (32–35),
had markedly increased expression levels (Fig. 3B). These results were further
confirmed by RT-qPCR analysis (Fig.
3C-F). This indicated that muscone treatment may induce HepG2
cell apoptosis through ER stress. The DDIT3 transcription factor is
an important pro-apoptotic factor in ER stress that is highly
expressed under such conditions (36). It is activated by three
transmembrane signaling proteins on the ER membrane, including PERK
(37), and during periods of
excessive or long-lived ER stress, activated PERK forms a complex
with eIF2α and ATF4 to induce DDIT3 expression (38,39).
DDIT3 subsequently accumulates in the nucleus, promoting apoptosis
and inhibiting the expression of the anti-apoptotic protein, Bcl-2
(40). Since ATF4 and DDIT3
expression levels were significantly increased following muscone
treatment compared with the control (Fig. 3C and D), it was hypothesized that
muscone may induce HepG2 cell apoptosis through the PERK/ATF4/DDIT3
signaling pathway. To verify this, the changes in the expression
levels of relevant proteins in this pathway were investigated.
Muscone treatment markedly increased the phosphorylation levels of
PERK and eIF2α proteins, and the expression levels of ATF4 and
DDIT3 in the downstream signaling pathway (Fig. 3G). To confirm the effects of ER
stress on muscone-induced growth inhibition, ER stress was blocked
by treatment with 3 mM 4-PBA for 1 h, followed by treatment with
muscone for 24 h. DAPI staining and flow cytometric analysis
revealed that 4-PBA treatment inhibited muscone-induced apoptosis
(Fig. 3H and I). Compared with the
muscone treatment group, 4-PBA treatment markedly decreased the
phosphorylation of PERK and eIF2α proteins, as well as the protein
expression levels of ATF4 and DDIT3 (Fig. 3J), 4-PBA treatment also partially
reversed muscone-induced cell viability inhibition (Fig. 3K). These results further confirmed
that muscone may induce HepG2 cell apoptosis through the
PERK/ATF4/DDIT3 signaling pathway.
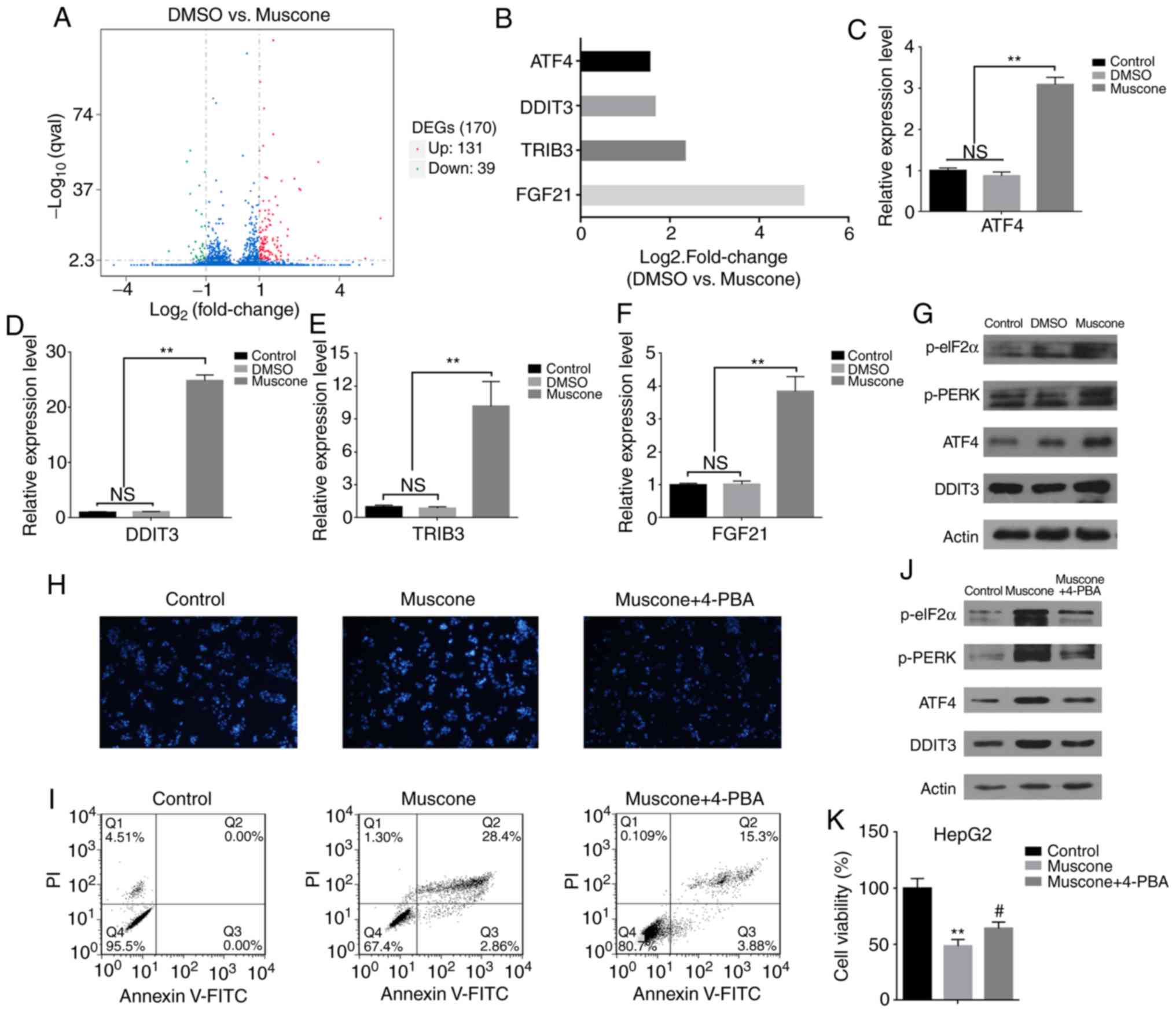 | Figure 3.Transcriptome sequencing and gene
expression analysis of HepG2 cells revealed that muscone influences
the expression levels of genes associated with ER stress. (A)
Volcano plot demonstrating the DEGs identified in HepG2 cells
following muscone treatment. Overall, 131 upregulated genes and 39
downregulated genes were identified in the muscone group compared
with the DMSO group. DEGs were screened according to fold-change
and significance level [|log2 (Fold Change) | >1 and q-value
<0.005]. Red, upregulated; green, downregulated; blue, no
change. (B) Fold-change of genes associated with ER stress (ATF4,
DDIT3, TRIB3 and FGF21) in HepG2 cells (DMSO vs. muscone) following
transcriptome sequencing and gene expression analysis. (C-F)
Reverse transcription-quantitative PCR analysis of (C) ATF4, (D)
DDIT3, (E) TRIB3 and (F) FGF21 expression levels in HepG2 cells
treated with DMSO or muscone for 24 h. Expression levels were
normalized to the control group (no treatment). (G) Expression
levels of proteins involved in the PERK/ATF4/DDIT3 signaling
pathway in HepG2 cells treated with DMSO or muscone for 24 h were
detected using western blotting. (H) Effects of 4-PBA treatment on
muscone-induced apoptosis. Apoptotic cells were stained with DAPI
and visualized using a microscope at an ×40 magnification (Olympus
IX71). (I) Apoptotic rate was determined in muscone and muscone +
4-PBA treated cells using flow cytometry based on the percentage of
Annexin V/FITC-positive cells. (J) Expression levels of
PERK/ATF4/DDIT3 signaling pathway-related proteins were detected
using western blotting. (K) Effects of 4-PBA inhibition on muscone
cell viability in HepG2 cells. **P<0.01 vs. control group;
#P<0.05 vs. muscone group; n=3; NS, not significant.
PI, propidium iodide; DEGs, differentially expressed genes; ER,
endoplasmic reticulum; ATF4, anti-activating transcription factor
4; DDIT3, anti-DNA damage inducible transcript 3; TRIB3, Tribbles
pseudokinase 3; FGF21, fibroblast growth dactor 21; 4-PBA, 4-phenyl
butyric acid. |
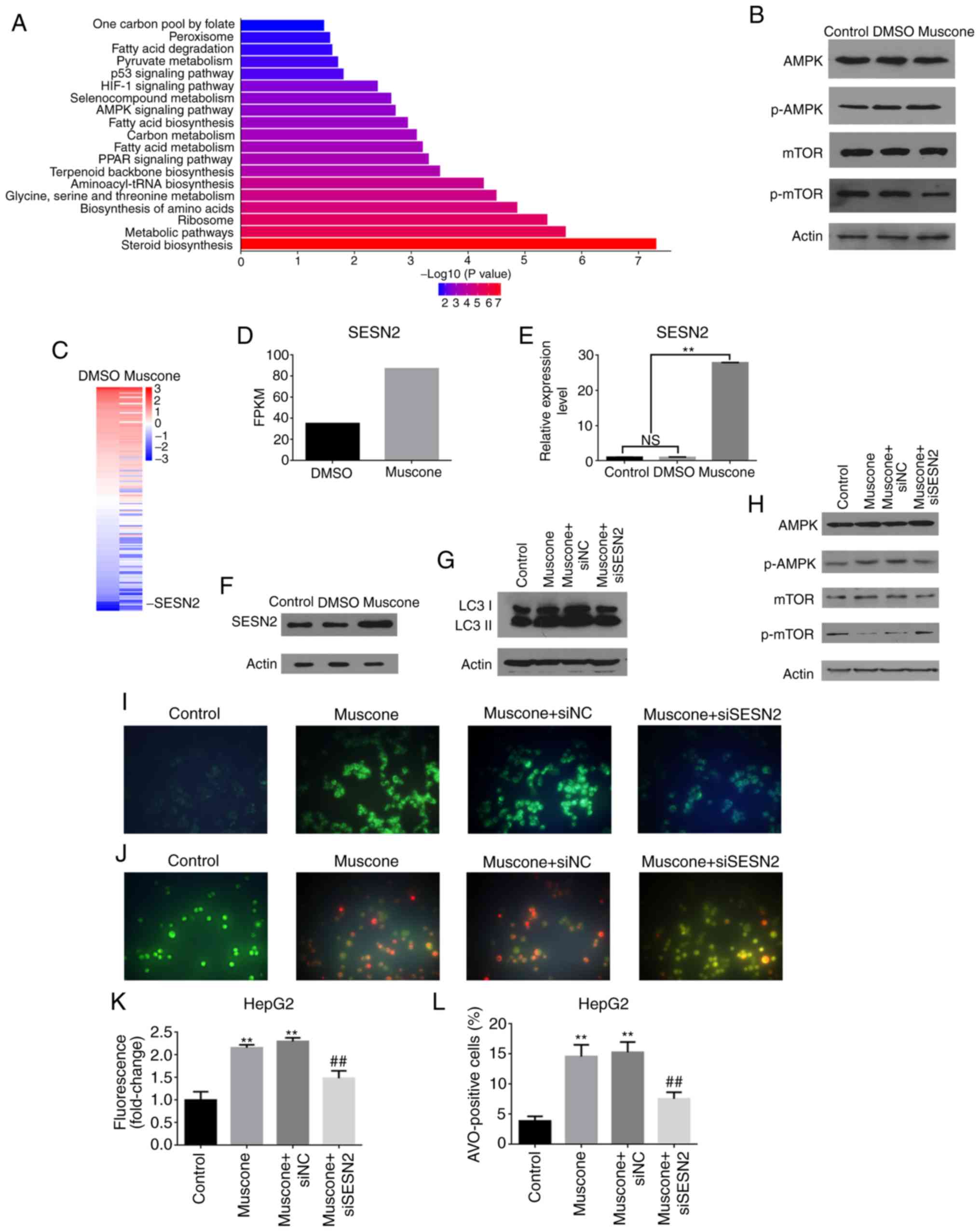
Muscone induces HepG2 cell autophagy
through the SESN2/AMPK/mTOR1 signaling pathway
To further investigate which autophagy-related
signaling pathways are regulated by muscone, transcriptome sequence
analysis of HepG2 cells treated with DMSO or muscone was performed.
GO analysis demonstrated that the function of DEGs was associated
with ‘aminoacyl tRNA ligase activity’, ‘synthesis pathways’ and
‘organic nitrogen complex metabolic pathways’ (data not shown).
KEGG pathway analysis reported that the DEGs were mainly enriched
in the following pathways: ‘One-carbon pool by folate’,
‘peroxisome’, ‘fatty acid metabolism (degradation, synthesis and
metabolism)’, ‘pyruvate metabolism’, ‘p53 signaling’, ‘HIF-1
signaling’, ‘organic selenium compound metabolism’, ‘AMPK
signaling’ and ‘PPAR signaling’ (Fig.
4A). Western blotting further revealed that muscone increased
AMPK expression levels and suppressed the mTOR pathway (Fig. 4B). Among the DEGs, the
autophagy-related gene SESN2 (41)
was reported to be highly activated by muscone treatment (Fig. 4C and D).
The involvement of SESN2 in muscone-associated
autophagy was investigated further, and it was observed that the
expression levels of SESN2 were significantly increased at both the
mRNA and protein level by muscone treatment (Fig. 4E and F). Using siRNA targeting human
SESN2 to knock down its endogenous expression in HepG2 cells, it
was demonstrated that LC3 II expression levels were decreased in
SESN2-knockdown cells treated with muscone compared with the
control (Fig. 4G). In addition,
SESN2 depletion significantly attenuated the muscone-mediated
induction of AMPK and mTOR phosphorylation (Fig. 4H). AVOs were visualized using MDC
staining and AO staining (Fig. 4I and
J). Results showed a significantly reduced percentage of
AVO-positive cells which was demonstrated using fluorescence
analysis (Fig. 4K) and cell counts
(Fig. 4L). These findings suggested
that muscone may induce autophagy through the SESN2-mediated
activation of the AMPK pathway.
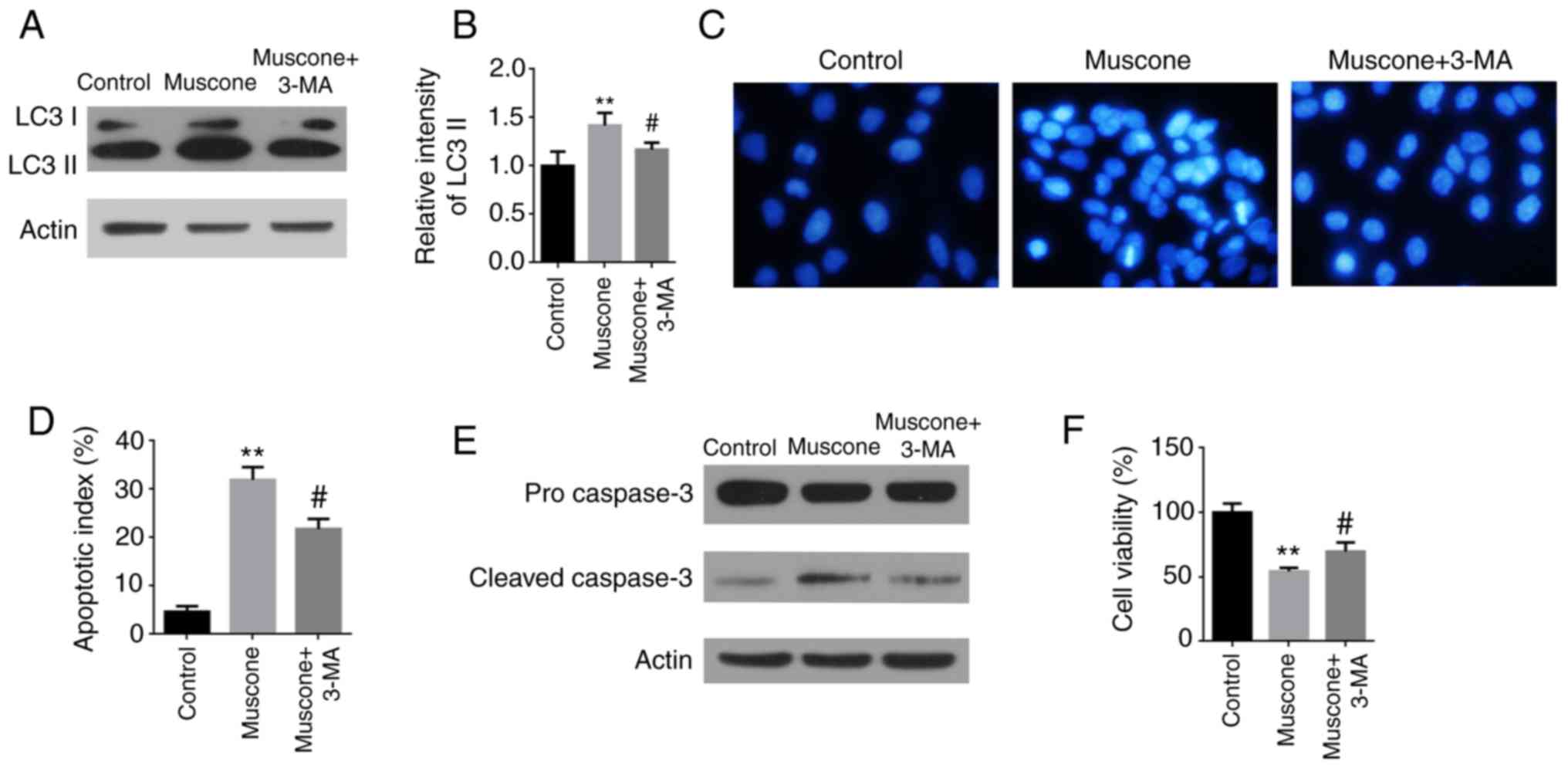
Muscone induces autophagy-dependent
apoptosis in HepG2 cells
Taken together, the results of the present study
demonstrated that muscone triggered apoptotic cell death and
simultaneously activated autophagy in liver cancer cells. To
investigate whether autophagy affected muscone-induced apoptotic
cell death, the autophagy inhibitor, 3-MA was used. Compared with
the muscone group, the pretreatment of cells with 3-MA reduced
muscone-induced autophagy, which was demonstrated through decreased
expression levels of LC3 II (Fig. 5A
and B). The 3-MA inhibitor relieved the muscone-induced
chromatin condensation in HepG2 cells and attenuated the
muscone-induced increased expression levels of caspase-3 in HepG2
cells (Fig. 5C-E). Additionally,
the pretreatment of cells with 3-MA relieved the significantly
decreased levels of cell viability induced by muscone (Fig. 5F). These data demonstrated that the
inhibition of autophagosome formation by 3-MA attenuated
muscone-induced apoptosis, suggesting that muscone-induced
apoptosis may be partially autophagy-dependent.
Muscone suppresses subcutaneous tumor
growth in mice
The effects of muscone on tumor growth in
vivo were subsequently investigated. HepG2 cells were injected
subcutaneously into athymic nude mice as previously described
(42). The transplantation of HepG2
cells into nude mice successfully induced the in-situ
formation of liver cancer (Fig.
6A). Treatment with muscone for 1 week significantly reduced
the tumor volume and weight compared with DMSO treatment (Fig. 6B-D). The role of apoptosis in
muscone-inhibited subcutaneous tumor growth was analyzed. The
apoptosis-related proteins, including Bax, Bcl-2 and caspase-3 were
assayed using western blotting of two different groups (group 1:
No. 1 and No. 5 subcutaneous tumor in Fig. 6B; group 2: No. 2 and No. 6
subcutaneous tumor in Fig. 6B). The
protein expression levels of cleaved caspase-3 and Bax were
markedly increased, whereas Bcl-2 expression levels were decreased
in the muscone-treated group compared with the control (Fig. 6E). To further confirm that muscone
induced subcutaneous tumor apoptosis via the PERK/ATF4/DDIT3
signaling pathway, the phosphorylation levels of eIF2α and PERK, as
well as the protein expression levels of ATF4 and DDIT3 were
examined. Muscone treatment increased the expression levels of the
PERK/ATF4/DDIT3 signaling pathway-related proteins compared with
the DMSO group (Fig. 6E). Thus, the
role of autophagy in muscone-induced inhibition of tumor growth was
further investigated by analyzing the protein expression levels of
p-AMPK, p-mTOR, SESN2 and LC3-II. Muscone treatment markedly
increased the expression levels of p-AMPK, LC3-II and SENS2, and
reduced the expression levels of p-mTOR compared with the DMSO
group (Fig. 6F). These results
suggest that muscone may inhibit HCC-transplanted subcutaneous
tumor growth in vivo by inducing apoptosis and
autophagy.
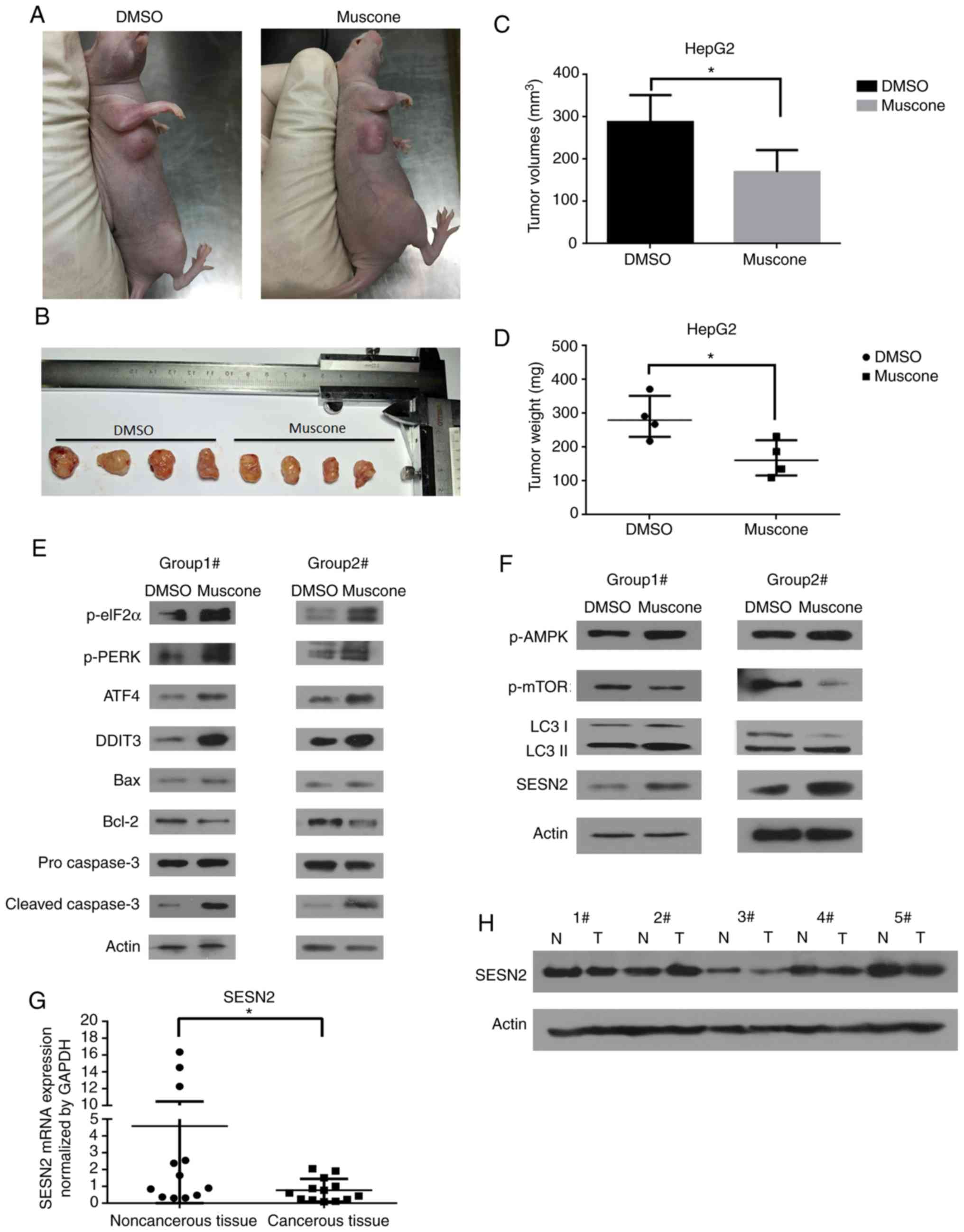 | Figure 6.Effects of muscone on subcutaneous
tumor growth in HepG2 cells. (A and B) Morphology of HepG2 cell
subcutaneous tumors in BALB/c nude mice injected with HepG2 cells
and treated with muscone (no. 5-8) or DMSO (no. 1-4). (C) Tumor
volumes of BALB/c nude mice injected with HepG2 cells and treated
with DMSO or muscone. (D) Tumor weight (in mg) of subcutaneous
tumors in BALB/c nude mice injected with HepG2 cells and treated
with muscone or DMSO. (E) Expression levels of PERK/ATF4/DDIT3
signaling pathway-related proteins (p-eIF2α, p-PERK, ATF4 and
DDIT3) and apoptosis-related markers (Bax, Bcl-2 and caspase-3)
were detected by western blotting from two different groups of
tumor tissues (group 1: no. 1 and no. 5 subcutaneous tumors; group
2: no. 2 and no. 6 subcutaneous tumors). (F) Western blotting was
used to analyze the expression levels of autophagy-related markers
in two different groups of tumor tissues (group 1: no. 1 and no. 5;
group 2: no. 2 and no. 6). Actin was used as a loading control. (G)
Reverse transcription-quantitative PCR analysis was used to analyze
SESN2 expression levels in HCC tissues (T) compared with
corresponding non-cancerous tissues (N). *P<0.05. (H) Western
blotting was used to investigate SESN2 expression levels in 5
samples randomly selected from HCC tumor samples compared with
corresponding non-cancerous tissue samples. p-eiF2α, phosphorylated
eukaryotic initiation factor 2α; p-PERK, phosphorylated protein
kinase R-like endoplasmic reticulum kinase; ATF4, anti-activating
transcription factor 4; DDIT3, anti-DNA damage inducible transcript
3; p-AMPK, phosphorylated AMP-activated protein kinase; p-mTOR1,
phosphorylated mechanistic target of rapamycin kinase 1; SESN2,
anti-sestrin 2. |
Finally, the prognostic role of SESN2 expression
levels in HCC was investigated. RT-qPCR and western blotting
demonstrated that both SESN2 mRNA and protein expression levels
were significantly decreased in the 14 human HCC tissues compared
with the non-cancerous tissue samples (Fig. 6G and H). Thus, these findings
indicated that SESN2 may be a potential candidate for understanding
the molecular mechanisms of hepatocarcinogenesis, and especially
for HCC diagnosis and therapy.
Discussion
In the present study, the effects of muscone on the
induction of liver cancer cell apoptosis and autophagy were
investigated. Muscone exhibited a broad range of antitumor
activities that were mediated by apoptotic and autophagic pathways,
which is consistent with that observed in previous clinical
studies. Using RNA-Seq to detect differentially expressed genes
(DEGs) and signaling pathways in HepG2 cells treated with muscone,
it was demonstrated that ER stress was mediated by the
muscone-induced PERK/ATF4/DDIT3 apoptotic pathway and the
SENS2/AMPK autophagy signaling pathway. In vivo experiments
also confirmed that the intraperitoneal injection of muscone
inhibited the proliferation of transplanted tumors in nude mice;
this inhibition was related to the PERK/ATF4/DDIT3 apoptotic
signaling pathway and the SESN2/AMPK/mTOR autophagy signaling
pathway.
In previous studies, both autophagy and apoptosis
have been reported to occur in the same cells (43–46),
and are suggested to intersect with each other through the mTOR and
AMPK signaling pathways (47,48).
Several coinciding apoptotic signals, such as the AMPK signaling
pathway and Bax have also been demonstrated to activate autophagy
(49). In fact, autophagy-induced
apoptosis has been reported by numerous studies (50,51)
and it is currently hypothesized that these processes most commonly
occur in sequence, with autophagy preceding apoptosis (52). The results of the present study
revealed that the inhibition of autophagosome formation attenuated
muscone-induced apoptosis, suggesting that muscone-induced
apoptosis may be partially autophagy-dependent. In support of
autophagy-dependent apoptosis, it has been suggested that autophagy
may function upstream of apoptosis and participate in the process
of membrane blebbing by maintaining cellular ATP levels (53,54);
however, this possibility remains to be elucidated in future
studies.
The present study indicated that the SESN2/AMPK/mTOR
pathway may be an important pathway for mediating muscone-induced
autophagy in liver cancer cells. The SESN family of proteins
consists of a highly conserved group of stress-inducible proteins
with molecular weights ranging between 54–57 kDa. They are
activated by different environmental stresses, such as DNA damage,
oxidative stress and hypoxia, and following activation, they
promote the recycling of peroxiredoxin through their sulfonic
reductase activity to reduce the levels of reactive oxygen species
(55). SESNs have also been
reported to regulate the AMPK/mTOR complex 1 signaling pathway,
which is involved in cellular autophagy (56,57).
Our study detected decreased expression levels of SESN2 in liver
cancer tissues compared with paracancerous tissues (58), which provided clinical evidence that
muscone may inhibit tumor development.
In conclusion, to the best of our knowledge, the
findings from the present study were among the first to suggest
that in addition to its analgesic effects, muscone may also induce
liver cancer cell autophagy and apoptosis. Autophagy and apoptosis
prevent necrosis-induced inflammatory responses in tumor tissues
and ameliorate tissue pain caused by the compression induced by
tumor growth. Another advantage of muscone is that its low toxicity
prevents toxic inflammatory responses from occurring in healthy
liver cells, which is of great importance for clinical practice.
Thus, these findings revealed that muscone demonstrated potential
for its use clinically as an anticancer drug, which may provide the
basis for the development of more effective anticancer drugs
derived from natural substances. Future studies are required to
develop highly efficient anticancer drugs that target tumor
cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
31300674).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SX and LX designed the study; WQ, ZL, CY, JD, QZ, DW
and CW performed the experiments and analyzed the data; SX and WQ
wrote the manuscript; and LX and ZL revised the manuscript. All
authors read and approved the final manuscript and agreed to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Institutional Ethics
Committee of the Changchun University of Traditional Chinese
Medicine (approval no. CCZYFYCC2017-071) and adhered to the
principles in the Declaration of Helsinki. Informed consent was
obtained from each patient before tissue collection for
experimentation. All animal studies were approved by the Committee
on the Use of Live Animals in Teaching and Research of Sichuan
University, and were conducted in accordance with approved
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
World Health Statistics 2017, . Monitoring
health for the SDGs. https://www.who.int/gho/publications/world_health_statistics/2017/en/WHO;
May 17–2017
|
|
2
|
Schwarz RE and Smith DD: Trends in local
therapy for hepatocellular carcinoma and survival outcomes in the
US population. Am J Surg. 195:829–836. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Llovet JM, Di Bisceglie AM, Bruix J,
Kramer BS, Lencioni R, Zhu AX, Sherman M, Schwartz M, Lotze M,
Talwalkar J, et al: Panel of experts in HCC-design clinical trials
in hepatocellular carcinoma. J Natl Cancer Inst. 100:698–711. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan L and Cheng P: Advances in treatment
of advanced hepatocellular cancer. J Clin Hepatol. 10:47–60.
2014.
|
|
5
|
Moriguchi M, Umemura A and Itoh Y: Current
status and future prospects of chemotherapy for advanced
hepatocellular carcinoma. Clin J Gastroenterol. 9:184–190. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
et al: Sorafenib in advanced hepatocellular carcinoma. N Engl J
Med. 359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S,
Kim JS, Luo R, Feng J, Ye S, Yang TS, et al: Efficacy and safety of
sorafenib in patients in the Asia-Pacific region with advanced
hepatocellular carcinoma: A phase III randomised, double-blind,
placebo-controlled trial. Lancet Oncol. 10:25–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo X: Clinical observation of compound
musk injection for treating primary liver cancer pain. Guid J
Tradit Chin Med Pharm. 12:41–43. 2014.
|
|
9
|
Lewandowski RJ, Kulik LM, Riaz A,
Senthilnathan S, Mulcahy MF, Ryu RK, Ibrahim SM, Sato KT, Baker T,
Miller FH, et al: A comparative analysis of transarterial
downstaging for hepatocellular carcinoma: Chemoembolization versus
radioembolization. Am J Transplant. 9:1920–1928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Krishnan S, Dawson LA, Seong J, Akine Y,
Beddar S, Briere TM, Crane CH and Mornex F: Radiotherapy for
hepatocellular carcinoma: An overview. Ann Surg Oncol.
15:1015–1024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Salem R, Lewandowski RJ, Mulcahy MF, Riaz
A, Ryu RK, Ibrahim S, Atassi B, Baker T, Gates V, Miller FH, et al:
Radioembolization for hepatocellular carcinoma using Yttrium-90
microspheres: A comprehensive report of long-term outcomes.
Gastroenterology. 138:52–64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi JW, Park JY, Ahn SH, Yoon KT, Ko HK,
Lee DY, Lee JT, Kim KS, Choi JS, Han KH, et al: Efficacy and safety
of transarterial chemoembolization in recurrent hepatocellular
carcinoma after curative surgical resection. Am J Clin Oncol.
32:564–569. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takayasu K, Arii S, Kudo M, Ichida T,
Matsui O, Izumi N, Matsuyama Y, Sakamoto M, Nakashima O, Ku Y, et
al: Superselective transarterial chemoembolization for
hepatocellular carcinoma. Validation of treatment algorithm
proposed by Japanese guidelines. J Hepatol. 56:886–892. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lo CM, Ngan H, Tso WK, Liu CL, Lam CM,
Poon RT, Fan ST and Wong J: Randomized controlled trial of
transarterial lipiodol chemoembolization for unresectable
hepatocellular carcinoma. Hepatology. 35:1164–1171. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schwarz RE, Abou-Alfa GK, Geschwind JF,
Krishnan S, Salem R and Venook AP; American
Hepato-Pancreato-Biliary Association, Society of Surgical Oncology
and Society for Surgery of the Alimentary Tract, : Nonoperative
therapies for combined modality treatment of hepatocellular cancer:
Expert consensus statement. HPB (Oxford). 12:313–320. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong N, Li Y and Hua Y: Research progress
of chinese medicine in the treatment of cancer pain. World Chin
Med. 8:1501–1504. 2013.
|
|
17
|
Liu R, Runyon RS, Wang Y, Oliver SG, Fan T
and Zhang W: Deciphering ancient combinatorial formulas: The
Shexiang Baoxin pill. Science. 347:S40–S42. 2015.
|
|
18
|
Choi HB: Knowledge transmission of medical
prescription and the role of the literati officials from the Song
dynasty: Focusing on huadupainongneibusan. Uisahak. 27:89–130.
2018.PubMed/NCBI
|
|
19
|
Xu L and Cao Y: Native musk and synthetic
musk ketone strongly induced the growth repression and the
apoptosis of cancer cells. BMC Complement Altern Med. 16:5112016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Zhang J and Li L: Comparison of the
therapeutic effects of different compositions of muskone in the
treatment of experimental myocardial infarct in rats and analgesia
in mice. Phytother Res. 22:1219–1223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma LF, Hao LP, Li LJ, Chen H and Ma JH:
Process of the Study on Muscone. Hebei Chem Industry. 33:11–14.
2010.(In Chinese).
|
|
22
|
Guo Y, Gu H and Shi Z: Synthesis of
2,15-hexadecanedione as a precursor of muscone. Chin J Chem.
23:334–336. 2005. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuribayashi K, Mayes PA and El-Deiry WS:
What are caspases 3 and 7 doing upstream of the mitochondria?
Cancer Biol Ther. 5:763–765. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lockshin RA: Programmed cell death:
History and future of a concept. J Soc Biol. 199:169–173. 2005.(In
French). View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Visconti R and D'Adamio L: Functional
cloning of genes regulating apoptosis in neuronal cells. Methods
Mol Biol. 399:125–131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu Y, Wang D, Wang X, Wang Y, Ren F, Chang
D, Chang Z and Jia B: Caspase 3 is activated through caspase 8
instead of caspase 9 during H2O2-induced apoptosis in HeLa cells.
Cell Physiol Biochem. 27:539–546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thornberry NA and Lazebnik Y: Caspases:
Enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vermeulen K, Berneman ZN and Van
Bockstaele DR: Cell cycle and apoptosis. Cell Prolif. 36:165–175.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim EH, Sohn S, Kwon HJ, Kim SU, Kim MJ,
Lee SJ and Choi KS: Sodium selenite induces superoxide-mediated
mitochondrial damage and subsequent autophagic cell death in
malignant glioma cells. Cancer Res. 67:6314–6324. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ueno T, Sato W, Horie Y, Komatsu M, Tanida
I, Yoshida M, Ohshima S, Mak TW, Watanabe S and Kominami E: Loss of
Pten, a tumor suppressor, causes the strong inhibition of autophagy
without affecting LC3 lipidation. Autophagy. 4:692–700. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ohoka N, Yoshii S, Hattori T, Onozaki K
and Hayashi H: TRB3, a novel ER stress-inducible gene, is induced
via ATF4-CHOP pathway and is involved in cell death. EMBO J.
24:1243–1255. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bromati CR, Lellis-Santos C, Yamanaka TS,
Nogueira TCA, Leonelli M, Caperuto LC, Gorjão R, Leite AR, Anhê GF
and Bordin S: UPR induces transient burst of apoptosis in islets of
early lactating rats through reduced AKT phosphorylation via
ATF4/CHOP stimulation of TRB3 expression. Am J Physiol Regul Integr
Comp Physiol. 300:R92–R100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ye J and Koumenis C: ATF4, an ER stress
and hypoxia-inducible transcription factor and its potential role
in hypoxia tolerance and tumorigenesis. Curr Mol Med. 9:411–416.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang S, Yan C, Fang QC, Shao ML, Zhang
YL, Liu Y, Deng YP, Shan B, Liu JQ, Li HT, et al: Fibroblast growth
factor 21 is regulated by the IRE1α-XBP1 branch of the unfolded
protein response and counteracts endoplasmic reticulum
stress-induced hepatic steatosis. J Biol Chem. 289:29751–29765.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamaguchi Y, Larkin D, Lara-Lemus R,
Ramos-Castañeda J, Liu M and Arvan P: Endoplasmic reticulum (ER)
chaperone regulation and survival of cells compensating for
deficiency in the ER stress response kinase, PERK. J Biol Chem.
283:17020–17029. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Roidl D, Hellbach N, Bovio PP, Villarreal
A, Heidrich S, Nestel S, Grüning BA, Boenisch U and Vogel T: DOT1L
activity promotes proliferation and protects cortical neural stem
cells from activation of ATF4-DDIT3-mediated ER stress in vitro.
Stem Cells. 34:233–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ma J, Yang YR, Chen W, Chen MH, Wang H,
Wang XD, Sun LL, Wang FZ and Wang DC: Fluoxetine synergizes with
temozolomide to induce the CHOP-dependent endoplasmic reticulum
stress-related apoptosis pathway in glioma cells. Oncol Rep.
36:676–684. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
McCullough KD, Martindale JL, Klotz LO, Aw
TY and Holbrook NJ: Gadd153 sensitizes cells to endoplasmic
reticulum stress by down-regulating Bcl2 and perturbing the
cellular redox state. Mol Cell Biol. 21:1249–1259. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sanli T, Steinberg GR, Singh G and
Tsakiridis T: AMP-activated protein kinase (AMPK) beyond
metabolism: A novel genomic stress sensor participating in the DNA
damage response pathway. Cancer Biol Ther. 15:156–169. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yu DC, Lee JS, Yoo JY, Shin H, Deng H, Wei
Y and Yun CO: Soluble vascular endothelial growth factor decoy
receptor FP3 exerts potent antiangiogenic effects. Mol Ther.
20:938–947. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shao Y, Gao Z, Marks PA and Jiang X:
Apoptotic and autophagic cell death induced by histone deacetylase
inhibitors. Proc Natl Acad Sci USA. 101:18030–18035. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kumar D, Shankar S and Srivastava RK:
Rottlerin induces autophagy and apoptosis in prostate cancer stem
cells via PI3K/Akt/mTOR signaling pathway. Cancer Lett.
343:179–189. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jing K, Song KS, Shin S, Kim N, Jeong S,
Oh HR, Park JH, Seo KS, Heo JY, Han J, et al: Docosahexaenoic acid
induces autophagy through p53/AMPK/mTOR signaling and promotes
apoptosis in human cancer cells harboring wild-type p53. Autophagy.
7:1348–1358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nguyen TM, Subramanian IV, Kelekar A and
Ramakrishnan S: Kringle 5 of human plasminogen, an angiogenesis
inhibitor, induces both autophagy and apoptotic death in
endothelial cells. Blood. 109:4793–4802. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Baehrecke EH: Autophagy: Dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Levine B and Yuan J: Autophagy in cell
death: An innocent convict? J Clin Invest. 115:2679–2688. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kandala PK and Srivastava SK: Regulation
of macroautophagy in ovarian cancer cells in vitro and in vivo by
controlling glucose regulatory protein 78 and AMPK. Oncotarget.
3:435–449. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shimizu S, Yoshida T, Tsujioka M and
Arakawa S: Autophagic cell death and cancer. Int J Mol Sci.
15:3145–3153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yao X, Ma Y, Kang J, Wang Y, Jiang L, Chen
M, Sha S, Sun X and Cao J: Perfluorooctane sulfonate induces
autophagy-dependent apoptosis through spinster 1-mediated
lysosomal-mitochondrial axis and impaired mitophagy. Toxicol Sci.
153:198–211. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Y, Liu Y, Liu X, Jiang L, Yang G, Sun
X, Geng C, Li Q, Yao X and Chen M: Citreoviridin induces
autophagy-dependent apoptosis through lysosomal-mitochondrial axis
in human liver HepG2 cells. Toxins (Basel). 7:3030–3044. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sy LK, Yan SC, Lok CN, Man RY and Che CM:
Timosaponin A-III induces autophagy preceding mitochondria-mediated
apoptosis in HeLa cancer cells. Cancer Res. 68:10229–10237. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Qu X, Zou Z, Sun Q, Luby-Phelps K, Cheng
P, Hogan RN, Gilpin C and Levine B: Autophagy gene-dependent
clearance of apoptotic cells during embryonic development. Cell.
128:931–946. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu HJ, Shi ZY, Lin XL, Chen SM, Wang QY
and Tang SY: Upregulation of Sestrin2 expression protects against
macrophage apoptosis induced by oxidized low-density lipoprotein.
DNA Cell Biol. 34:296–302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Budanov AV, Lee JH and Karin M: Stressin'
Sestrins take an aging fight. EMBO Mol Med. 2:388–400. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lee JH, Budanov AV and Karin M: Sestrins
orchestrate cellular metabolism to attenuate aging. Cell Metab.
18:792–801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen S, Yan W, Lang W, Yu J, Xu L, Xu X,
Liu Y and Bao H: SESN2 correlates with advantageous prognosis in
hepatocellular carcinoma. Diagn Pathol. 12:13–19. 2017. View Article : Google Scholar : PubMed/NCBI
|