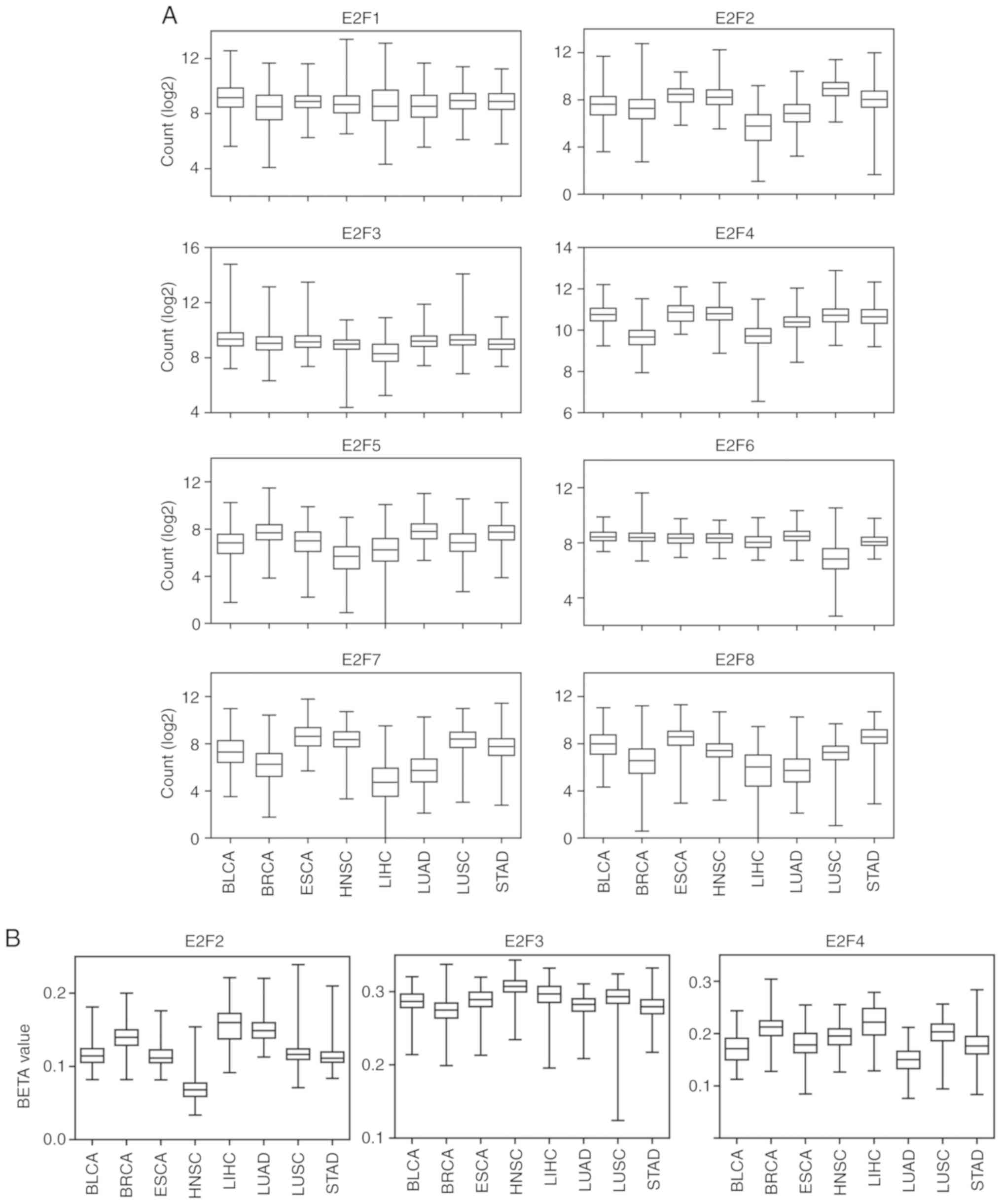
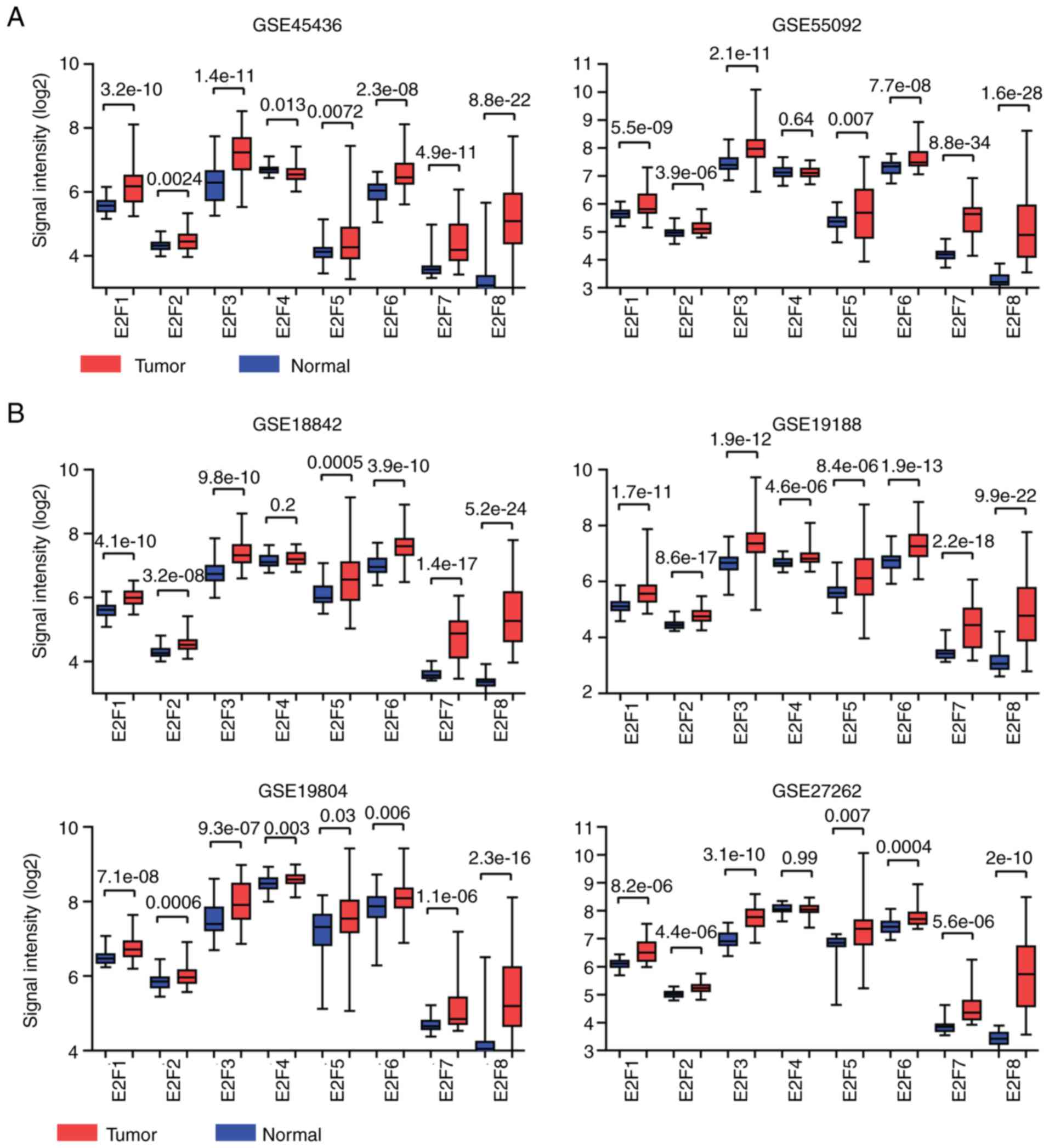
Introduction
E2F transcription factors include eight genes,
E2F1-8 (1), which play critical
roles in the regulation of cell proliferation and cell cycle
progression through the mediated transcriptional activation of
specific downstream target genes (2–4). E2F
family genes also regulate autophagy (5), mitochondrial functions (6) and the DNA damage response (7). Increased expression levels of the E2F
family genes are observed in a number of different types of cancer.
For example, E2F1 and E2F3 are increased in liver cancer, and the
overexpression of E2F1 or E2F3 could induce spontaneous liver
cancer development in mice (8,9).
Furthermore, knockout of E2F8 could protect mice against the
development of liver cancer (10).
E2F8 is also a therapeutic target for lung cancer. Compared with
normal lung cells, lung cancer cells exhibit upregulated E2F8
levels (11). Loss of E2F2
expression could decrease the level of susceptible breast tumor
progression by alleviating the Myc-mediated proliferative effects
(12). The global E2F signature is
associated with clinical outcomes of patients with pancreatic
adenocarcinoma (13). Restoring the
balance between E2F1 and E2F7 is a therapeutic strategy in head and
neck squamous cell carcinomas (14). These findings indicate the universal
functions of the E2F family genes in the development of cancer.
However, previous studies on the E2F family genes
have focused on a single E2F gene in a limited number of cancer
types (8–14). Therefore, there is an urgent need to
comprehensively analyze E2F family genes across tumor types. The
Cancer Genome Atlas (TCGA) project contains gene expression data
and DNA methylation profiles of normal and malignant tissues across
several types of cancer (15–18).
In addition, genetic aberrations, including gene mutations and DNA
amplification, are documented in TCGA database (19–23).
The majority of data in the present study was derived from TCGA,
allowing the functions of E2F family genes to be investigated in a
comprehensive manner.
The present study compared the expression levels of
the E2F family genes between normal and cancer tissues in each
tumor type. The present study also analyzed the prognostic effects
of the E2F family genes. The expression and methylation level of
the E2F family genes in different pathological stages and molecular
subtypes of different types of tumor were studied. Correlation
efficiency and multivariate cox regression analysis were used to
determine the associations between E2F family genes. Furthermore,
the present study identified key genomic mutations, DNA
amplification or epigenetic DNA methylation that contributed to the
increased expression levels of the E2F family genes. Overall, the
analysis of TCGA database in the present study allowed an improved
understanding of the functions of the E2F family genes. The results
also indicated the E2F family genes that may serve as potential
biomarkers for further therapeutic studies.
Materials and methods
Data acquisition
TCGA data were downloaded from TCGA hub (tcga.xenahubs.net). Pathological and molecular subtype
information of patients with liver hepatocellular carcinoma (LIHC)
(24) and lung adenocarcinoma
(LUAD) (25) was obtained from
previously published studies. Oncoprints of the E2F family genes in
different types of tumor were downloaded from cbioportal (version
3.2.0; www.cbioportal.org/index.do).
The gene expression series matrix of normal and
cancerous liver tissues was downloaded from the Gene Expression
Omnibus (GEO) website (www.ncbi.nlm.nih.gov/geo), and included the GEO
datasets GSE45436 (26) and
GSE55092 (27). The gene expression
series matrix of normal and cancerous lung tissues were downloaded
from the GSE18442 (28), GSE19188
(29), GSE19804 (30) and GSE27262 (31) datasets. The gene expression and
survival data of patients with lung cancer were downloaded from the
GSE30219 dataset (32).
TCGA data analysis
Gene expression profiles across cancer types were
analyzed using RNA-sequencing data (TCGA HiSeqV2 data). TP53
mutations were analyzed using DNA-sequencing data (TCGA mutation
broad data). The DNA methylation profiles were analyzed using
HumanMethylation450 microarray data
(Methylation_Preprocess.Level3).
iCluster classification of patients
with LIHC
iCluster is a novel classification system of
patients with LIHC, proposed by TCGA network (24). The DNA copy number, DNA methylation,
mRNA expression, miRNA expression and protein expression of 363
patients with LIHC were integrated to divide the patients into
three iClusters using a joint multivariate regression (24). The iCluster information of each
patient with LIHC was obtained from a previous study (24). The expression levels of the E2F
family genes were further determined in each iCluster subtype of
patients with LIHC.
GEO data analysis
The expression profiles obtained from the GEO
datasets were processed separately. The expression values were
averaged if multiple probes corresponded to the same gene symbol
using the ‘plyr’ package (version 1.8.5; cran.r-project.org/web/packages/plyr/index.html) in R
software (version 3.5.0; www.r-project.org). The paired Student's t-test was
used to determine differences in gene expression between normal and
cancer samples in R.
Kyoto Encyclopedia of Gens and Genomes
signaling pathway enrichment analysis
The signaling pathway enrichment analysis was
performed using the Database for Annotation, Visualization and
Integrated Discovery (version 6.8; david.ncifcrf.gov).
Gene set enrichment analysis
(GSEA)
The GSEA of the E2F transcription factors was
performed using GSEA software (version 2.0; www.broad.mit.edu/gsea/index.html). Genes were ranked
by the signal-to-noise ratio, and statistical significance was
determined by 1,000 gene set permutations.
Survival analysis
The Kaplan-Meier Plotter (kmplot.com/analysis/index.php?p=service) was used
to identify the association between the expression levels of the
E2F family genes and overall survival in patients with LIHC and
LUAD derived from TCGA database and GEO datasets. The associations
between the expression levels of the E2F family genes and overall
survival in patients with lung cancer from the GSE30219 dataset
were determined using the ‘survival’ package (version 3.1;
cran.r-project.org/web/packages/survival/index.html)
in R. Patients were divided into high and low expression groups
using the mean expression level of E2F family genes. P-values were
determined by the log-rank test.
Heatmaps
Heatmaps were created using the ‘pheatmap’ package
(version 1.0.12; cran.r-project.org/web/packages/pheatmap/index.html)
in R. The clustering scale was determined by the ‘average’ method.
The clustering distance was determined by the ‘correlation’
method.
Statistical analysis
Box plots and contingency graphs were generated
using GraphPad Prism software (version 5.0; GraphPad Software,
Inc.). Statistical analysis was performed using the paired
Student's t-test or χ2 test using R software.
Multivariate cox regression analyses of the E2F family genes were
performed using the ‘survival’ package (version 3.1; cran.r-project.org/web/packages/survival/index.html)
in R. Correlation plots of the E2F family genes were created using
the ‘corrplot’ package (version 0.84; cran.r-project.org/web/packages/corrplot/index.html)
in R. The Spearman's correlation test was used to demonstrate the
correlation between E2F family genes. P<0.05 was considered to
indicate a statistically significant difference.
Results
E2F family genes are upregulated in
several types of tumor
The E2F transcription factor family contains eight
members, E2F1-8 (3). The present
study identified the enriched signaling pathways of the eight E2F
family genes, and the results revealed that the cell cycle
signaling pathway was the most highly enriched (Fig. 1A). Notably, the majority of the
enriched pathways were tumor-specific-associated pathways, for
example, bladder cancer, non-small cell lung cancer and pancreatic
cancer pathways (Fig. 1A). E2F
family genes were also associated with the transforming growth
factor β (TGFβ) signaling pathway (Fig.
1A). All the enriched signaling pathways highlighted the
universal functions of the E2F family genes in cancer development
in several types of tumor. However, the E2F family genes have not
yet been studied in a comprehensive pan-cancer manner.
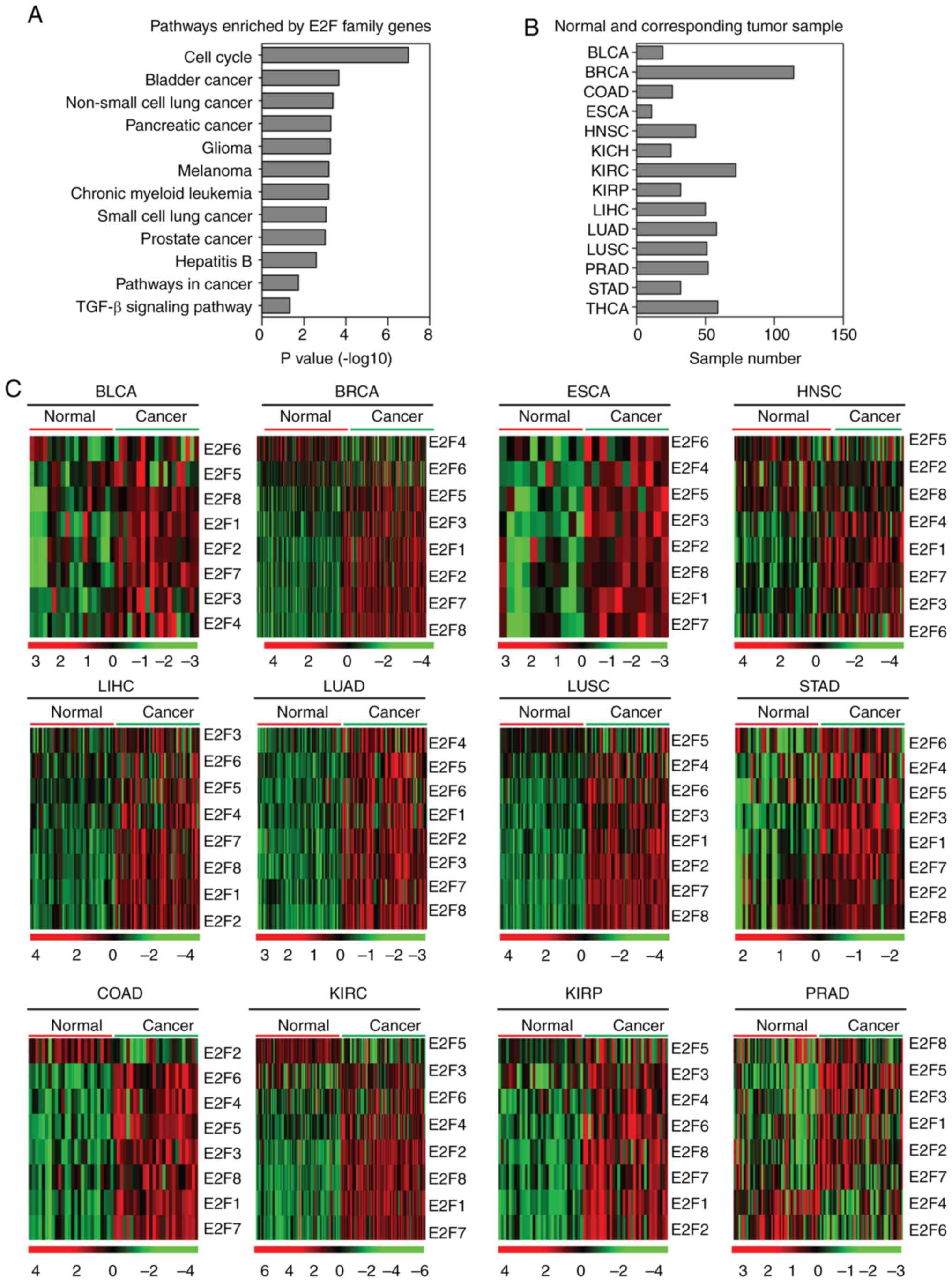 | Figure 1.Expression of E2F family genes is
increased in several tumor types. (A) Functional pathway enrichment
analysis of the E2F family genes using the Database for Annotation,
Visualization and Integrated Discovery. The most significantly
enriched pathways were shown. (B) Box plots demonstrated the number
of normal and corresponding tumor samples used in the present
study. (C) Heatmaps demonstrated the expression levels (log2 count)
of E2F family genes in normal and tumor samples in BLCA, BRCA,
COAD, ESCA, HNSC, KIRC, KIRP, LIHC, LUAD, LUSC, PRAD and STAD.
Upregulated (red), downregulated (green) and unchanged (black)
genes were delineated. BLCA, bladder urothelial carcinoma; BRCA,
breast invasive carcinoma; COAD, colon adenocarcinoma; ESCA,
esophageal carcinoma; HNSC, head and neck squamous cell carcinoma;
KIRC, kidney renal clear cell carcinoma; KIRP, kidney renal
papillary cell carcinoma; LIHC, liver hepatocellular carcinoma;
LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma;
PRAD, prostate adenocarcinoma; STAD, stomach adenocarcinoma. |
The present study used TCGA database to integrate
the analysis of the E2F family genes across different types of
cancer. The expression levels of the E2F family genes in normal
tissues and corresponding tumor tissues were investigated. As
illustrated in the heatmaps (Fig.
1B), the majority of the E2F family genes were upregulated in
bladder urothelial carcinoma (BLCA), breast invasive carcinoma
(BRCA), colon adenocarcinoma (COAD), esophageal carcinoma (ESCA),
head and neck squamous cell carcinoma (HNSC), kidney renal clear
cell carcinoma (KIRC), kidney renal papillary cell carcinoma
(KIRP), LIHC, LUAD, lung squamous cell carcinoma (LUSC) and stomach
adenocarcinoma (STAD) tumor tissues compared with normal tissues
(Fig. 1C). In particular, compared
with the normal tissues, all the E2F family genes were highly
expressed in LIHC, LUAD and STAD tumor tissues (Fig. 1C). However, it was revealed that the
majority of the E2F family genes were not altered in prostate
adenocarcinoma (PRAD; Fig. 1C). For
the other types of tumor, the E2F family genes were all highly
expressed in the corresponding tumor samples, excluding E2F4 in
BRCA, E2F2 in COAD, and E2F5 in HNSC, KIRC, KIRP and LUSC.
E2F transcription factors are highly
enriched in several types of tumors
The present study identified the transcription
factors enriched in tumors using GSEA (33). Among the transcription factors, E2F
was significantly enriched in BLCA, BRCA, ESCA, HNSC, LIHC, LUAD,
LUSC and STAD tumors, representing the most frequently enriched
transcription factor (Fig. 2).
These results indicated the universal importance of E2F
transcription factors in the development of cancer.
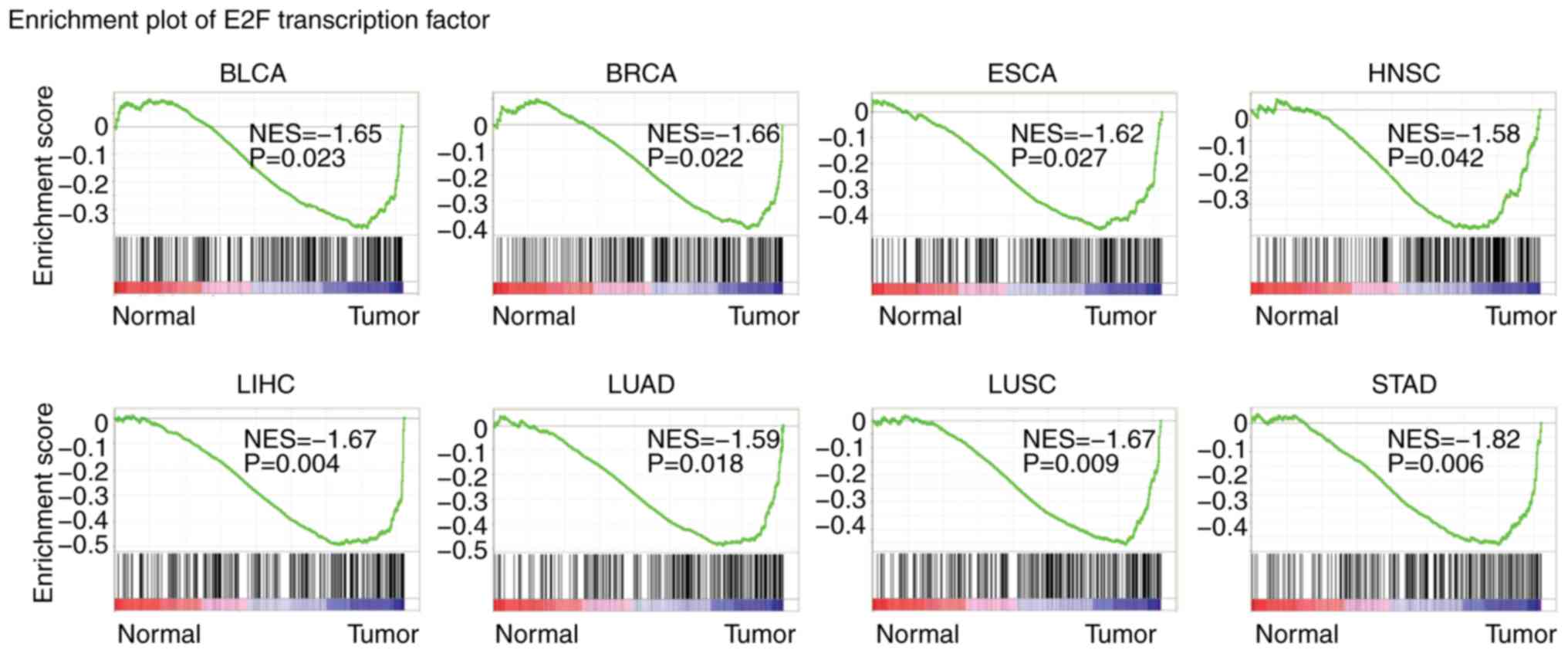 | Figure 2.E2F transcription factors are highly
enriched in several types of tumor. Enrichment plots demonstrated
the E2F transcriptions factors in BLCA, BRCA, ESCA, HNSC, LIHC,
LUAD, LUSC and STAD. Enrichment of NES and P-values are presented.
BLCA, bladder urothelial carcinoma; BRCA, breast invasive
carcinoma; COAD, colon adenocarcinoma; ESCA, esophageal carcinoma;
HNSC, head and neck squamous cell carcinoma; KIRC, kidney renal
clear cell carcinoma; KIRP, kidney renal papillary cell carcinoma;
LIHC, liver hepatocellular carcinoma; LUAD, lung adenocarcinoma;
LUSC, lung squamous cell carcinoma; PRAD, prostate adenocarcinoma;
STAD, stomach adenocarcinoma; NES, normalized enrichment score. |
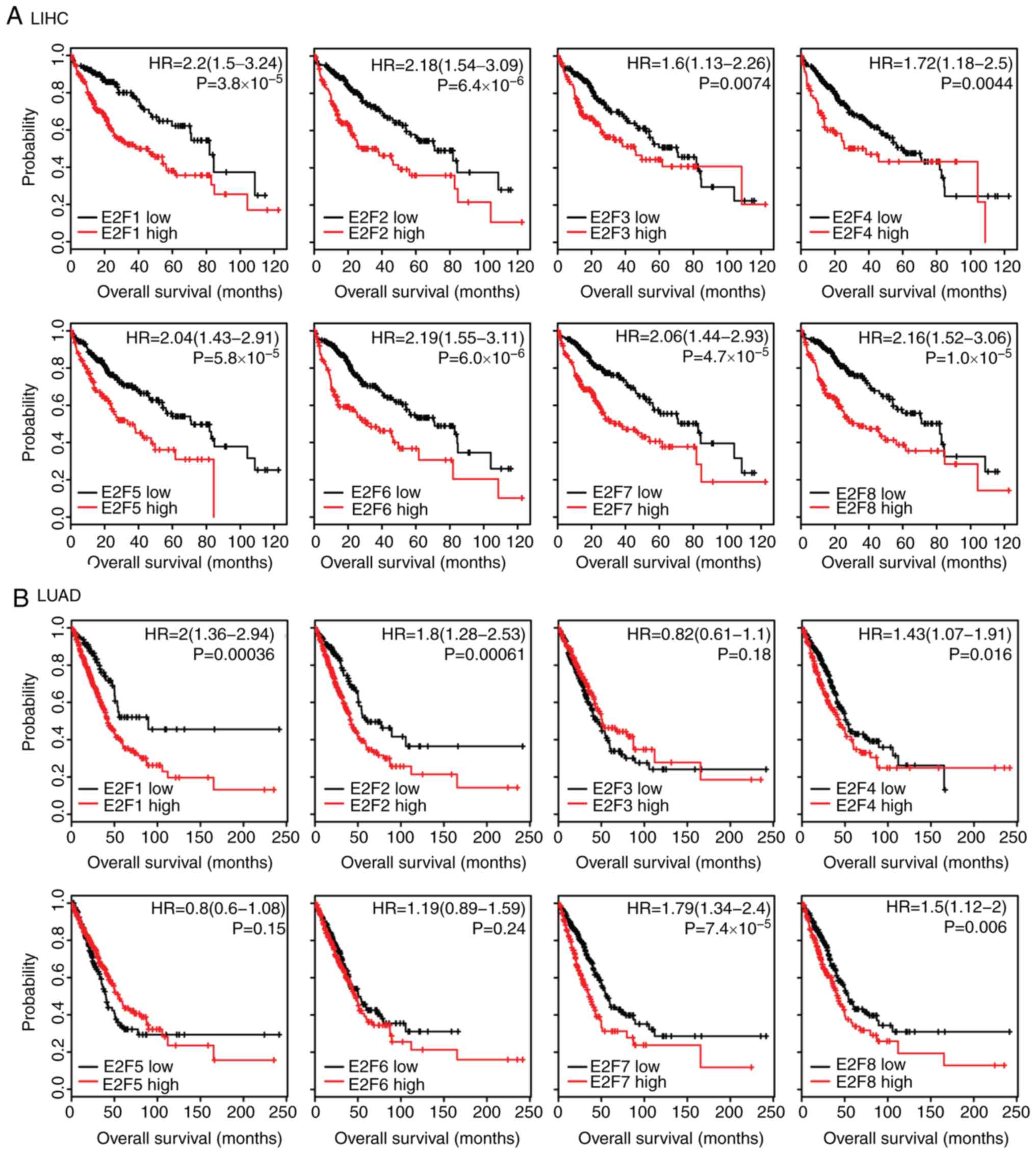
Expression levels of E2F family genes
are associated with the overall survival in patients with LIHC and
LUAD
The present study assessed whether the E2F family
genes had prognostic effects in tumor progression. Using the
Kaplan-Meier Plotter (34), the
present study identified the association between the expression
levels of the E2F family genes and overall survival in several
types of cancer. The results revealed that, high expression levels
of the E2F family genes were associated with an unfavorable
prognosis, particularly in LIHC and LUAD (Fig. 3). Patients with LIHC with higher
expression levels of E2F1-8 had worse prognosis than patients with
low expression (Fig. 3A).
Furthermore, the Kaplan-Meier survival analysis revealed that E2F1,
E2F2, E2F4, E2F7 and E2F8 were all associated with overall survival
in patients with LUAD (Fig. 3B).
However, the E2F family genes had no or little prognostic effect in
other types of tumor, including LUSC (Fig. S1A) or STAD (Fig. S1B). These results suggested that
although the E2F family genes were highly expressed across tumor
types, the E2F family genes were more important in the development
of LIHC and LUAD compared with other tumor types.
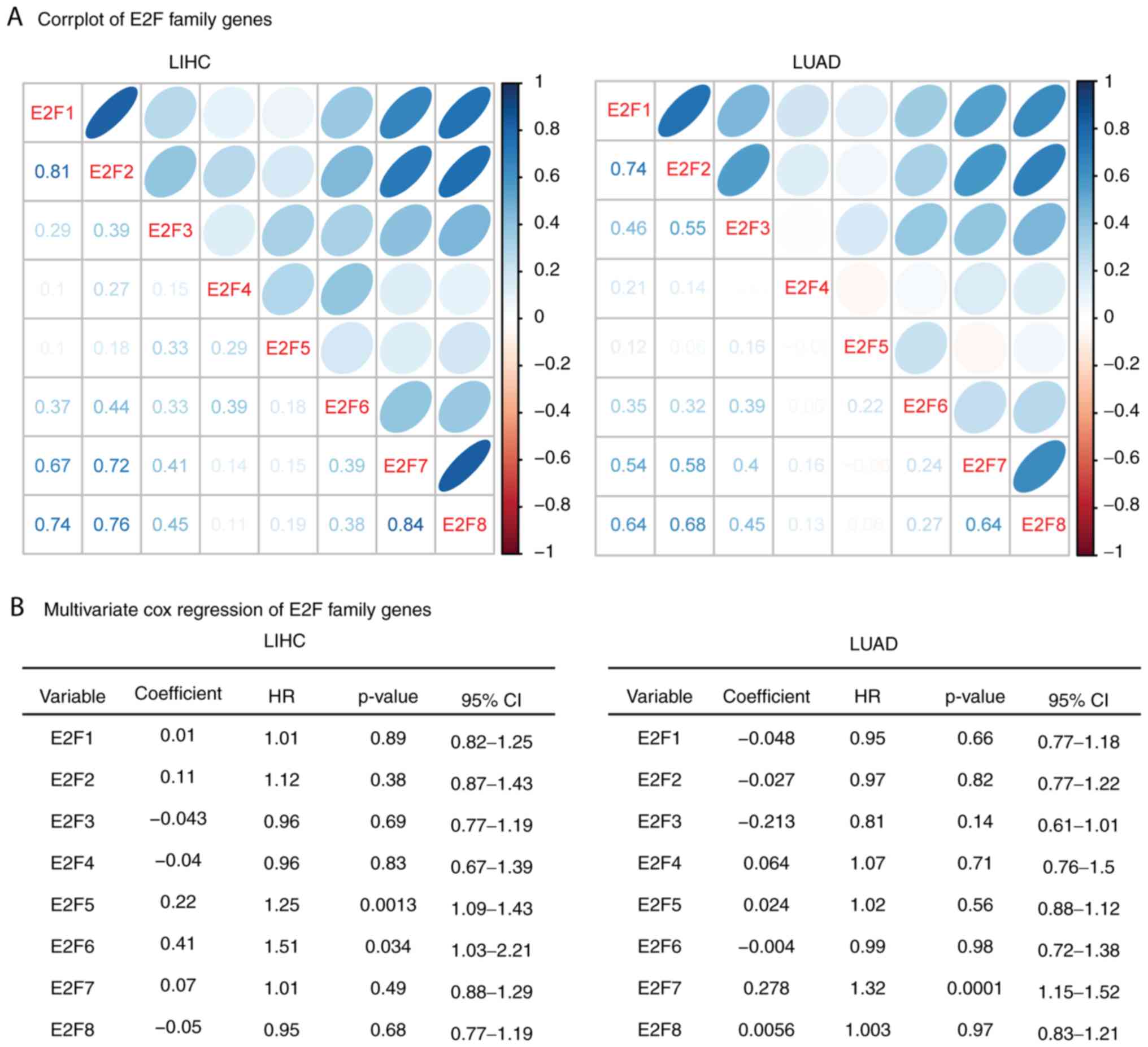
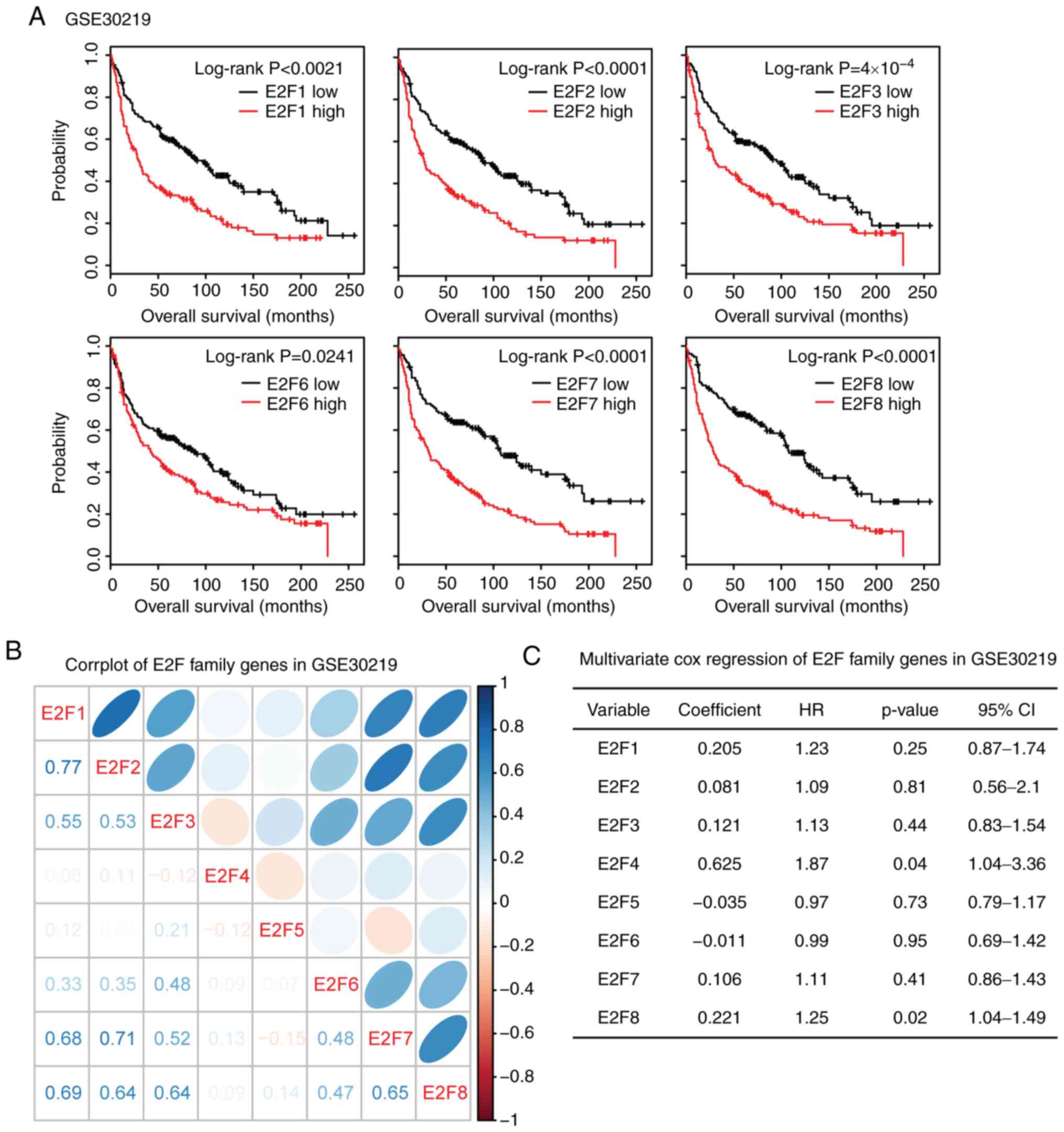
Correlation or independence of the E2F
family genes is identified in patients with LIHC and LUAD
The results of the present study demonstrated the
similar functions of the E2F family genes in overall survival in
patients with LIHC and LUAD. Thus, the present study aimed to
determine the connection between E2F family genes. The Spearman's
correlation test demonstrated a high correlation between E2F1, E2F7
and E2F8 in LIHC and LUAD expression datasets (Fig. 4A). E2F2 was also highly correlated
with E2F7 and E2F8 in LIHC and LUAD. Additionally, E2F7 and E2F8
were correlated with each other (Fig.
4A).
The present study also used multivariate cox
regression analyses to reveal the connection that the E2F family
genes have in determining the overall survival of patients with
LIHC and LUAD. It was revealed that E2F5 and E2F6 were independent
prognostic markers in patients with LIHC, and E2F7 was an
independent prognostic marker in patients with LUAD (Fig. 4B). These results were consistent
with the gene expression data, and E2F5 and E2F6 demonstrated
little connection with other E2F family genes (Fig. 4A). E2F1-4, E2F7 and E2F8 genes were
interconnected with each other, so those genes were not independent
prognostic markers (Fig. 4B).
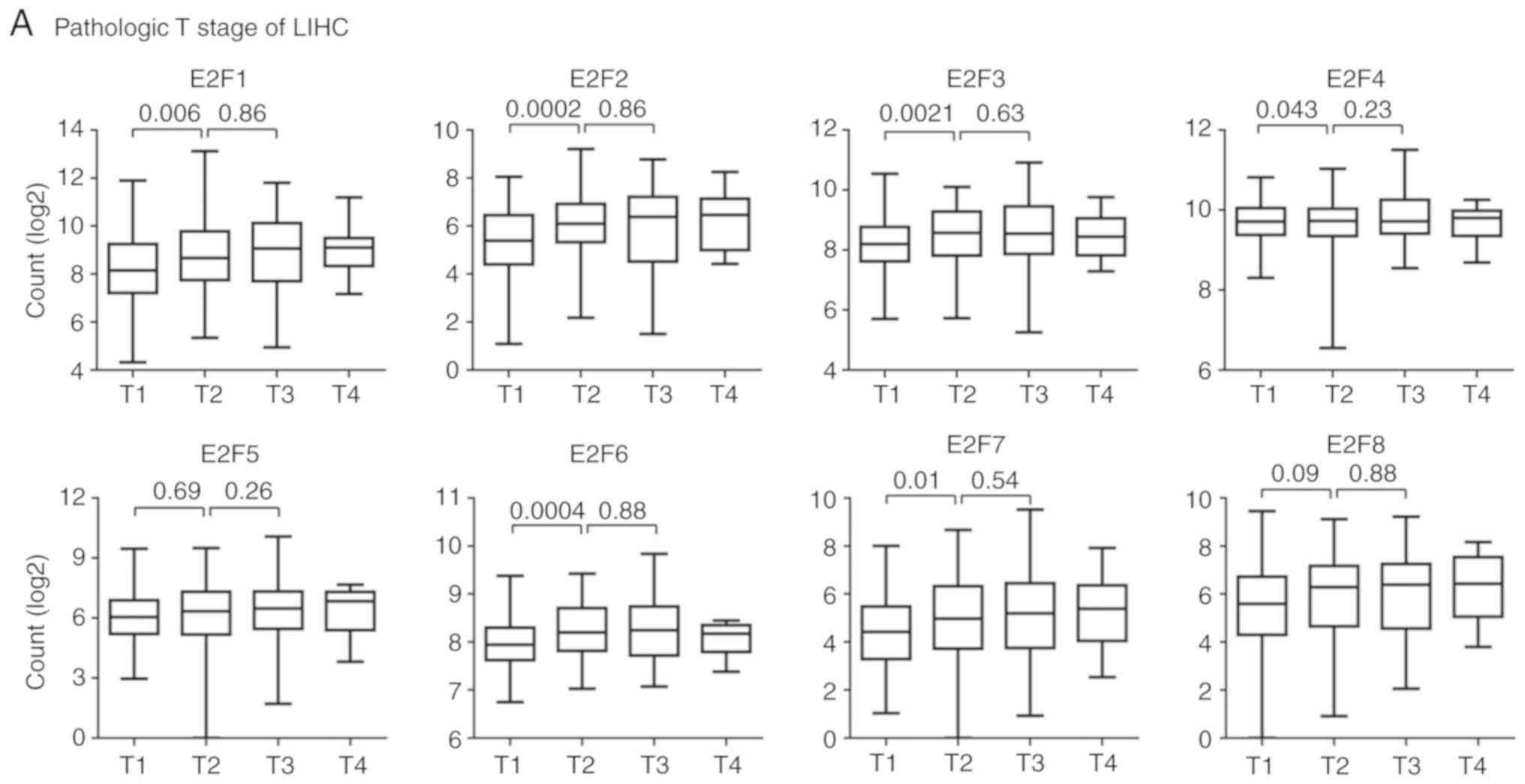
E2F family genes are downregulated in
patients with T1 stage and iCluster2 LIHC subtypes
The Kaplan-Meier Plotter analysis suggested that the
E2F family genes may play more important roles in LIHC than other
tumor types (Fig. 3A). LIHC is a
heterogeneous disease, reflected by differences in genomic or
epigenetic aberrations, pathological stages and responses to
therapies (35–37). Pathological stages are basic
prognostic information to evaluate the malignant characteristics of
the disease (38). The present
study then tested the expression levels of E2F family genes in
patients with different pathological stages of LIHC.
The majority of patients (98%) with LIHC in TCGA
database were in the M0 or N0 stage, without distant or lymphatic
metastasis. However, the expression levels of the E2F family genes
in different pathological T stages were quite different. Compared
with patients with T1 stage LIHC, E2F1-4, E2F6 and E2F7 were all
highly expressed in patients with T2 stage LIHC (Fig. 5A). However, the expression levels of
E2F5 and E2F8 in patients with T1 and T2 stage LIHC were not
different. Furthermore, there were no differences observed between
the expression level of the E2F family genes and patients with T2
stage or T3 stage LIHC (Fig.
5A).
Besides the basic pathological classification of
LIHC, patients with LIHC were divided into three iClusters based on
genomic alterations, gene expression profiles and DNA methylation
aberrations (24). Furthermore,
patients with different iCluster subtype demonstrated different
clinical outcomes. Patients with the iCluster1substype had a worse
prognosis than patients with the iCluster2 and iCluster3 subtypes
(24). The present study then
assessed the expression levels of the E2F family genes in patients
with different iCluster subtypes of LIHC. Compared with the
iCluster1 or iCluster3 subtypes, the expression levels of E2F1-4,
E2F6 and E2F7 were relatively lower in patients with iCluster2
subtype of LIHC (Fig. 5B). This low
expression of the E2F family genes in iCluster2 was not due to the
high proportion of patients with T1 stage LIHC (Fig. 5C).
The present study also assessed the expression
levels of the E2F family genes in different pathological stages or
molecular subtypes of patients with LUAD (25). However, no significantly different
expression levels of E2F family genes were observed in different
pathological stages (Fig. S2).
E2F2 and E2F8 were highly expressed in patients with T2 stage LUAD
compared with patients with T1 stage (Fig. S2).
Increased expression levels of the E2F
family genes are induced by multiple levels of genomic or
epigenetic aberrations
The present study aimed to determine the mechanisms
that induced the activation of the E2F family genes in tumor
development. The high expression levels of oncogenes are usually
mediated by hypo-DNA methylation, DNA amplification and gene
mutation (16). Compared with
normal tissues, it was revealed that the E2F family genes exhibited
hypo-DNA methylation patterns in LIHC and LUAD tumor tissues
(Fig. 6A). Particularly LIHC, E2F2,
E2F4, E2F5, E2F5 and E2F8 genes exhibited hypo-DNA methylation in
tumor samples (Fig. 6A), and E2F1,
E2F3 and E2F6 genes exhibited hypo-DNA methylation in LUAD
(Fig. 6A). These observations
suggested that DNA methylation was partially contributing to the
activation of E2F family genes in the tumor cells.
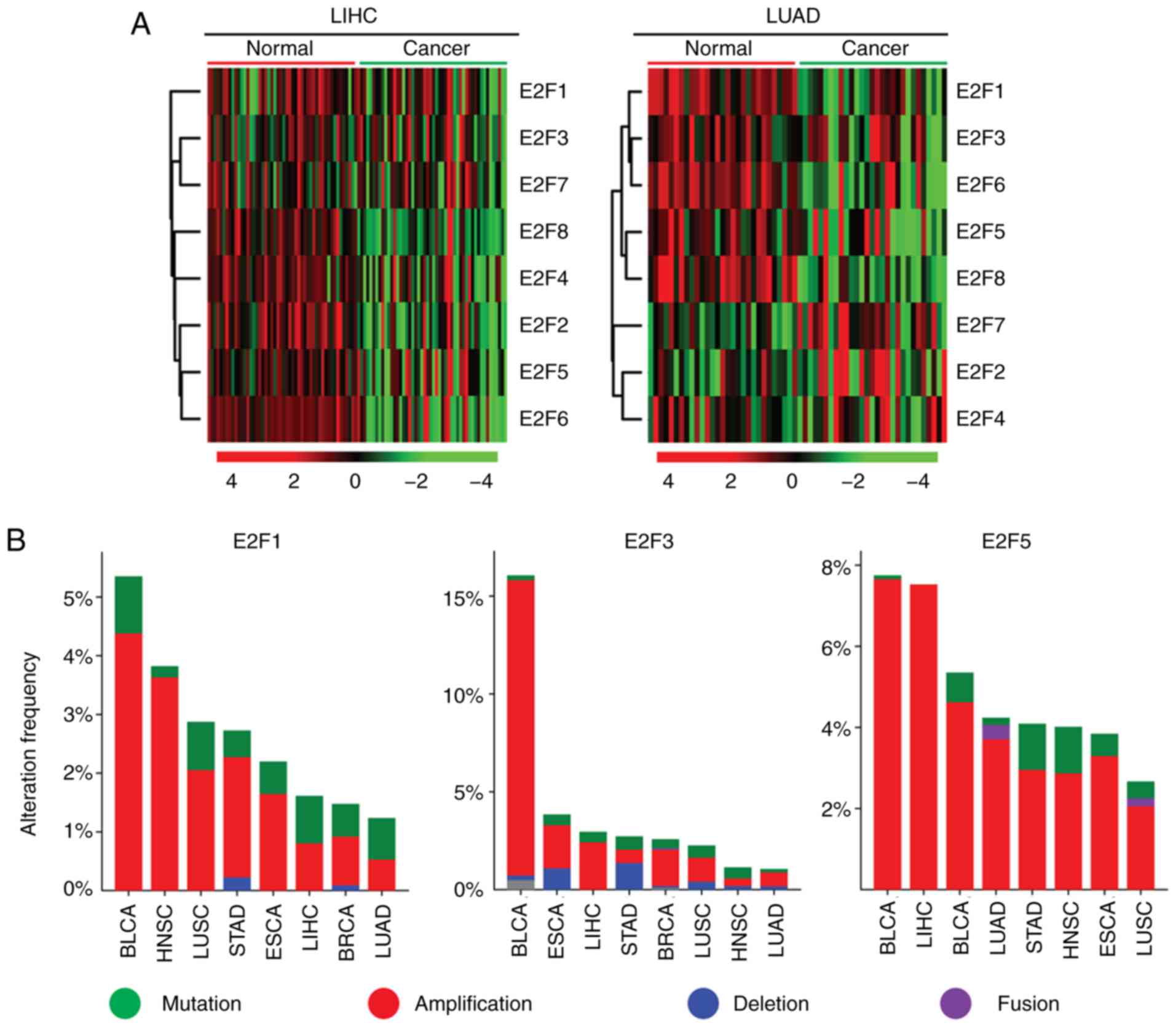 | Figure 6.Increased expression levels of the
E2F family genes are induced by multiple levels of genomic or
epigenetic aberrations. (A) Heatmaps demonstrated the methylation
level (β value) of the E2F family genes in normal and tumor samples
in patients with LIHC and LUAD. Hypermethylated (red),
hypomethylated (green) and unchanged (black) genes were delineated.
(B) Oncoprints demonstrated the alteration frequency of E2F1, E2F3
and E2F5 genes in patients with BLCA, HNSC, LUSC, STAD, ESCA, LIHC,
BRCA and LUAD. (C) Box plots demonstrated the expression levels of
the E2F family genes in patients with LIHC and LUAD. P-values
indicated the differences between patients with TP53 mutant and
TP53 wild-type cancer types. LIHC, liver hepatocellular carcinoma;
LUAD, lung adenocarcinoma; BLCA, bladder urothelial carcinoma;
BRCA, breast invasive carcinoma; ESCA, esophageal carcinoma; HNSC,
head and neck squamous cell carcinoma; LIHC, liver hepatocellular
carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell
carcinoma; STAD, stomach adenocarcinoma. |
Another factor determining the activation of E2F
genes in tumor cells was genomic aberration, particularly DNA
amplification. It was revealed that the E2F1 and E2F3 genes were
present in higher proportions of DNA amplification in BLCA
(Fig. 6B). BRCA, LIHC and BLCA
tumor types also exhibited high levels of E2F5 amplification
(Fig. 6B).
TP53 is a driver of mutations in several types of
tumor (23,40,41).
Loss of TP53 functions induces uncontrolled cell cycle progression,
and resistance to cell apoptosis (42–44).
The present study assessed whether TP53 regulated the expression
levels of the E2F family genes. It was revealed that, except for
E2F5, the other E2F family genes were all highly expressed in
patients with TP53 mutant LIHC and LUAD, compared with those
patients with TP53 wild-type LIHC and LUAD (Fig. 6C). Overall, the present study
speculated that hypo-DNA methylation, DNA amplification and TP53
mutation were contributing to the high expression levels of E2F
family genes in cancer cells.
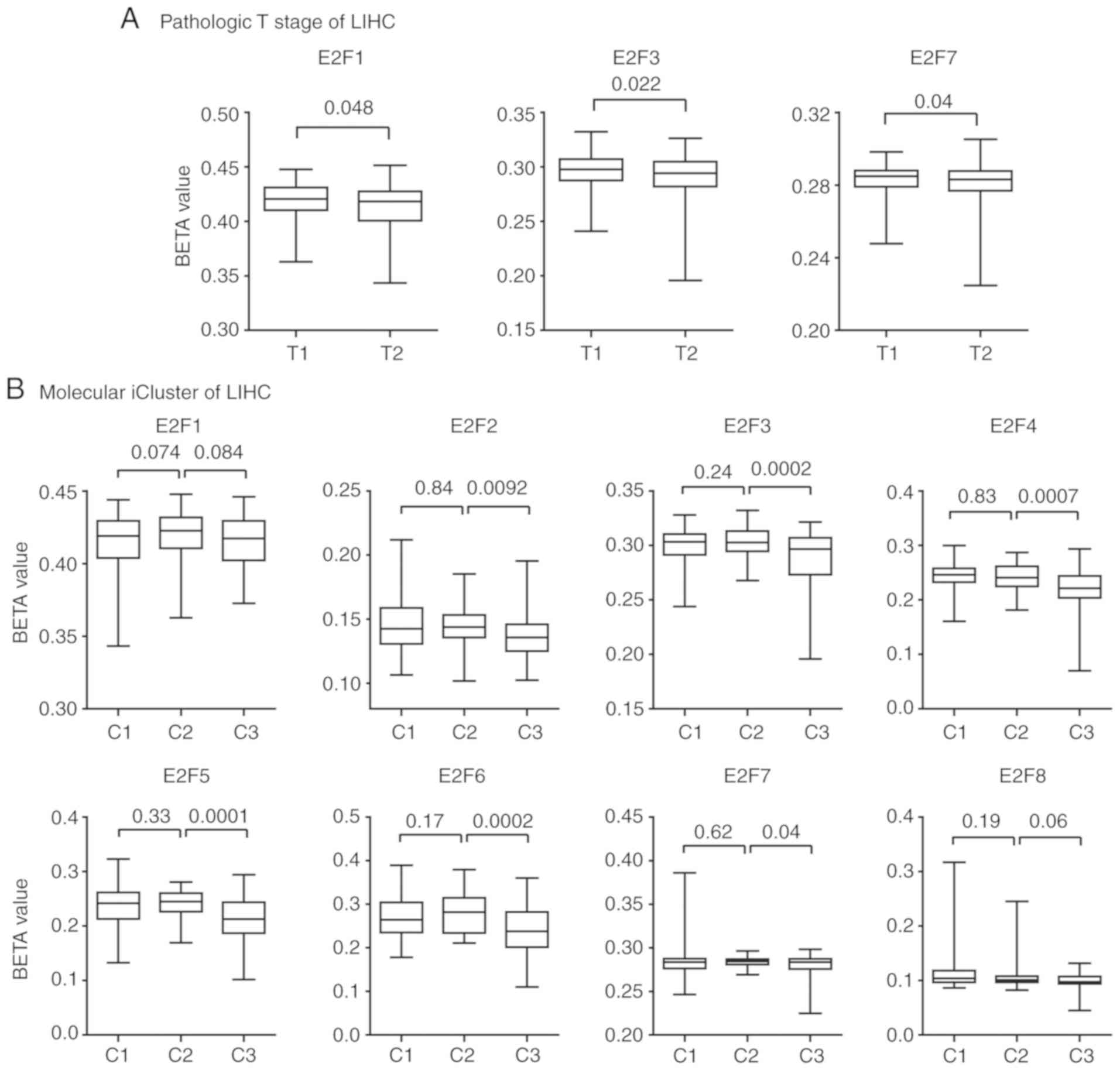
E2F family genes exhibit hyper-DNA
methylation patterns in patients with iCluster2 subtype LIHC
The present study demonstrated that the E2F family
genes were downregulated in patients with pathological T1 stage and
molecular iCluster2 subtypes of LIHC. However, the mechanisms
underlying this phenotype were not clear. The present study
analyzed the methylation patterns of the E2F family genes in
patients with LIHC. It was revealed that the E2F family genes E2F1,
E2F3 and E2F7 exhibited hyper-DNA methylation patterns in patients
with T1 stage LIHC compared with those patients with T2 stage LIHC
(Fig. 7A). No significantly
different methylation intensities of other E2F family genes were
observed between patients with pathological T1 and T2 LIHC.
The methylation patterns of the E2F family genes in
patients with different molecular iClusters of LIHC were also
analyzed. Consistent with the low expression levels of the E2F
family genes in patients with iCluster2 subtype of LIHC, the
present study revealed that the methylation intensity of E2F2-7 in
patients with LIHC was relatively higher, compared with patients
with iCluster3 subtype LIHC. However, there was no statistically
significant difference in methylation intensity in the E2F family
genes between patients with iCluster1 and iCluster2 subtypes of
LIHC (Fig. 7B).
E2F family genes are expressed in
particularly low levels in patients with LIHC
The present study compared the expression levels of
E2F family genes in BLCA, BRCA, ESCA, HNSC, LIHC, LUAD, LUSC and
STAD. It was revealed that, compared with other tumor types,
E2F2-4, E2F7 and E2F8 were all expressed at relatively low levels
in LIHC tumors (Fig. 8A). In
addition, the methylation intensity of E2F2-4 was relatively higher
in LIHC compared with other tumor types (Fig. 8B). However, the methylation level of
E2F3 was higher in HNSC than in LIHC (Fig. 8B).
Expression and prognostic effects of
the E2F family genes are validated from GEO datasets
From TCGA database, it was revealed that the
majority of the E2F family genes were upregulated in several types
of tumor tissue, and the upregulated E2F family genes were
associated with worse prognosis in LIHC and LUAD tumor types. In
order to further confirm these findings, the present study analyzed
the functions of the E2F family genes from GEO datasets.
The GSE45436 and GSE55092 datasets included the
expression profiles derived from normal liver and malignant liver
tissues (27). The expression
levels of the E2F family genes varied significantly between normal
liver or malignant tissues. E2F3 was highly expressed, while the
expression levels of E2F7 and E2F8 were relatively lower in liver
tissues (Fig. 9A). However,
compared with the normal liver tissues, the E2F family genes E2F1-8
were all highly expressed in liver cancer tissues (Fig. 9A). However, the expression of E2F4
between normal liver and cancerous liver tissues was not
significantly different in the GSE55092 dataset (Fig. 9A).
The expression levels of the E2F family genes
between normal lung and malignant lung tissues were also analyzed
using the GSE18842, GSE19188, GSE19804 and GSE27262 GEO datasets
(28–31). Similar to liver tissues, E2F3 was
highly expressed, while the expression levels of E2F7 and E2F8 were
relatively lower in normal lung or malignant tissues (Fig. 9B). In addition, compared with the
normal lung tissues, the E2F family genes were all highly expressed
in lung cancer tissues, except E2F4 (Fig. 9B). These results were relatively
consistent with the findings derived from TCGA database.
The Kaplan-Meier Plotter analysis also provided
prognostic values of genes using combined GEO transcriptional data
in non-small-cell lung cancer (45). Similar to TCGA data, high expression
levels of the E2F family genes E2F1, E2F2, E2F4, E2F7 and E2F8 were
all associated with low overall survival in patients with lung
cancer, while, high expression levels of E2F3, E2F5 and E2F6 were
associated with an improved prognosis in patients with LUAD
(Fig. 9C).
Correlation between E2F family genes
is validated in the GSE30219 lung cancer expression dataset
The present study validated the correlation of E2F
family genes in the GSE30219 lung cancer expression dataset. This
dataset was selected as it included both gene expression and
overall survival time of patients with lung cancer (32). High expression levels of E2F1-3 and
E2F6-8 were associated with poor overall survival (Fig. 10A). Results derived from this
dataset suggested a strong correlation among E2F1, E2F7 and E2F8
(Fig. 10B). E2F2 was also strongly
correlated with E2F7 and E2F8. In addition, E2F7 and E2F8 were
correlated with each other (Fig.
10B). Multivariate cox regression analysis revealed those genes
interconnected with each other in determining the overall survival
of patients with LUAD. Only E2F4 and E2F8 were independent
prognostic markers (Fig. 10C). All
these results were quite similar to those derived from TCGA
database.
Discussion
Using TCGA database, the present study analyzed the
transcriptional profiles, DNA methylation characteristics, DNA
amplification features and prognostic effects of the E2F family
genes in several tumor types. Compared with the normal tissues, E2F
family genes were highly expressed in multiple cancer tissues,
including BLCA, BRCA, COAD, ESCA, HNSC, KIRC, KIRP, LIHC, LUAD,
LUSC and STAD. Furthermore, the E2F transcription factors were
highly enriched in the aforementioned tumors. The results of the
present study suggested that the E2F family genes played important
and universal roles in cancer development. Furthermore, since the
E2F family genes were mostly associated with the cell cycle
signaling pathway, the results supported previous observations that
uncontrolled cell cycle progression was a hallmark of cancer
(46,47).
The potential prognostic roles of E2F family genes
in breast cancer (48), gastric
cancer (49), liver cancer
(50) and non-small-cell lung
cancer (51) have been previously
studied. For example, higher E2F1, E2F3, E2F5, E2F7 and E2F8
expression levels were significantly associated with lower overall
survival in patients with breast cancer (48). E2F1, E2F3 and E2F4 were
significantly associated with unfavorable outcomes in patients with
gastric cancer (49). E2F1 and E2F2
upregulation was significantly correlated with poor prognosis in
patients with non-small-cell lung cancer (51). However, in the present study, the
E2F family genes had no or little prognostic effect in LUSC and
STAD. On the contrary, the high expression levels of E2F family
genes, E2F1-8, were all associated with unfavorable outcomes in
patients with LIHC. In patients with LUAD, E2F1, E2F2, E2F4, E2F7
and E2F8, were significantly associated with unfavorable outcomes.
The data from the present study suggested that the E2F family genes
had more important functions in LIHC and LUAD than in other tumor
types.
The expression levels of E2F family genes in tumor
cells are regulated by several molecular mechanisms. In
retinoblastoma (RB), RB protein was a critical regulator of E2F
family genes (52). The present
study demonstrated that, in LIHC and LUAD patients, the
hypomethylation of the E2F family genes in tumor samples may
explain the high expression levels of these genes in the tumor
tissues. Moreover, in BLCA, the high expression levels of the E2F
family genes may be due to DNA amplification. Furthermore, BRCA and
LIHC tumor types exhibited high levels of E2F5 amplification. TP53
tumor suppressor is also a critical regulator of E2F family genes.
E2F7 is a TP53 direct transcriptional target gene, connecting TP53
with RB protein (53). E2F family
genes and TP53 bind the same promoter region to regulate downstream
target genes (54). The present
study demonstrated that, except for E2F5, the E2F family genes were
all highly expressed in patients with TP53 mutant LIHC and LUAD,
compared with patients with TP53 wild-type LIHC and LUAD.
Overall, the analysis in the present study provided
a more in-depth understanding of the biological functions of the
E2F family genes. However, the results require further clinical
validation. The results of the present study suggested that the E2F
family genes may serve as important biomarkers and therapeutic
targets, particularly for patients with LIHC and LUAD.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Fujian Provincial Maternity and the Children's Hospital (grant nos.
YCXB 18-10 and YCXM 19-04).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in TCGA (tcga.xenahubs.net) and GEO (www.ncbi.nlm.nih.gov/geo) repositories.
Authors' contributions
HW designed the study and wrote the manuscript. HW,
XW and LX performed the data analysis. HC and JZ designed the study
and supervised the work.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kent LN and Leone G: The broken cycle: E2F
dysfunction in cancer. Nat Rev Cancer. 19:326–338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu H, Tang X, Srivastava A, Pécot T,
Daniel P, Hemmelgarn B, Reyes S, Fackler N, Bajwa A, Kladney R, et
al: Redeployment of Myc and E2f1-3 drives Rb-deficient cell cycles.
Nat Cell Biol. 17:1036–1048. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morgunova E, Yin Y, Jolma A, Dave K,
Schmierer B, Popov A, Eremina N, Nilsson L and Taipale J:
Structural insights into the DNA-binding specificity of E2F family
transcription factors. Nat Commun. 6:100502015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen HZ, Tsai SY and Leone G: Emerging
roles of E2Fs in cancer: An exit from cell cycle control. Nat Rev
Cancer. 9:785–797. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang H, Martin V, Gomez-Manzano C,
Johnson DG, Alonso M, White E, Xu J, McDonnell TJ, Shinojima N and
Fueyo J: The RB-E2F1 pathway regulates autophagy. Cancer Res.
70:7882–7893. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Benevolenskaya EV and Frolov MV: Emerging
links between E2F control and mitochondrial function. Cancer Res.
75:619–623. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bertoli C, Herlihy AE, Pennycook BR,
Kriston-Vizi J and de Bruin RAM: Sustained E2F-dependent
transcription is a key mechanism to prevent
replication-stress-induced DNA damage. Cell Rep. 15:1412–1422.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kent LN, Bae S, Tsai SY, Tang X,
Srivastava A, Koivisto C, Martin CK, Ridolfi E, Miller GC, Zorko
SM, et al: Dosage-Dependent copy number gains in E2f1 and E2f3
drive hepatocellular carcinoma. J Clin Invest. 127:830–842. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tarangelo A, Lo N, Teng R, Kim E, Le L,
Watson D, Furth EE, Raman P, Ehmer U and Viatour P: Recruitment of
pontin/reptin by E2f1 amplifies E2f transcriptional response during
cancer progression. Nat Commun. 6:100282015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kent LN, Rakijas JB, Pandit SK, Westendorp
B, Chen HZ, Huntington JT, Tang X, Bae S, Srivastava A, Senapati S,
et al: E2f8 mediates tumor suppression in postnatal liver
development. J Clin Invest. 126:2955–2969. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park SA, Platt J, Lee JW, Lopez-Giraldez
F, Herbst RS and Koo JS: E2F8 as a novel therapeutic target for
lung cancer. J Natl Cancer Inst. 18:1072015.
|
|
12
|
Fujiwara K, Yuwanita I, Hollern DP and
Andrechek ER: Prediction and genetic demonstration of a role for
activator E2Fs in myc-induced tumors. Cancer Res. 71:1924–1932.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lan W, Bian B, Xia Y, Dou S, Gayet O,
Bigonnet M, Santofimia-Castaño P, Cong M, Peng L, Dusetti N and
Iovanna J: E2F signature is predictive for the pancreatic
adenocarcinoma clinical outcome and sensitivity to E2F inhibitors,
but not for the response to cytotoxic-based treatments. Sci Rep.
8:83302018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saenz-Ponce N, Pillay R, de Long LM,
Kashyap T, Argueta C, Landesman Y, Hazar-Rethinam M, Boros S,
Panizza B, Jacquemyn M, et al: Targeting the XPO1-dependent nuclear
export of E2F7 reverses anthracycline resistance in head and neck
squamous cell carcinomas. Sci Transl Med. 27:4472018.
|
|
15
|
Yan X, Hu Z, Feng Y, Hu X, Yuan J, Zhao
SD, Zhang Y, Yang L, Shan W, He Q, et al: Comprehensive genomic
characterization of long non-coding RNAs across human cancers.
Cancer Cell. 28:529–540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun W, Bunn P, Jin C, Little P,
Zhabotynsky V, Perou CM, Hayes DN, Chen M and Lin DY: The
association between copy number aberration, DNA methylation and
gene expression in tumor samples. Nucleic Acids Res. 46:3009–3018.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weisenberger DJ: Characterizing DNA
methylation alterations from the cancer genome atlas. J Clin
Invest. 124:17–23. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saghafinia S, Mina M, Riggi N, Hanahan D
and Ciriello G: Pan-Cancer landscape of aberrant DNA methylation
across human tumors. Cell Rep. 25:1066–1080. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weinstein JN, Collisson EA, Mills GB, Shaw
KR, Ozenberger BA, Ellrott K, Shmulevich I, Sander C and Stuart JM;
Cancer Genome Atlas Research Network, : The cancer genome atlas
pan-cancer analysis project. Nat Genet. 45:1113–1120. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hutter C and Zenklusen JC: The cancer
genome atlas: Creating lasting value beyond its data. Cell.
173:283–285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uhlen M, Zhang C, Lee S, Sjöstedt E,
Fagerberg L, Bidkhori G, Benfeitas R, Arif M, Liu Z, Edfors F, et
al: A pathology atlas of the human cancer transcriptome. Science.
35:eaan2507. 7–2017.
|
|
22
|
Hoadley KA, Yau C, Hinoue T, Wolf DM,
Lazar AJ, Drill E, Shen R, Taylor AM, Cherniack AD, Thorsson V, et
al: Cell-of-origin patterns dominate the molecular classification
of 10,000 tumors from 33 types of cancer. Cell. 173:291–304. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bailey MH, Tokheim C, Porta-Pardo E,
Sengupta S, Bertrand D, Weerasinghe A, Colaprico A, Wendl MC, Kim
J, Reardon B, et al: Comprehensive characterization of cancer
driver genes and mutations. Cell. 173:371–385. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cancer Genome Atlas Research Network.
Electronic address, . wheeler@bcm.edu and CancerGenome Atlas
Research Network: Comprehensive and integrative genomic
characterization of hepatocellular carcinoma. Cell. 169:1327–1341.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular profiling of lung adenocarcinoma. Nature.
511:543–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang HW, Hsieh TH, Huang SY, Chau GY, Tung
CY, Su CW and Wu JC: Forfeited hepatogenesis program and increased
embryonic stem cell traits in young hepatocellular carcinoma (HCC)
comparing to elderly HCC. BMC Genomics. 14:7362013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Melis M, Diaz G, Kleiner DE, Zamboni F,
Kabat J, Lai J, Mogavero G, Tice A, Engle RE, Becker S, et al:
Viral expression and molecular profiling in liver tissue versus
microdissected hepatocytes in hepatitis B virus-associated
hepatocellular carcinoma. J Transl Med. 12:2302014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sanchez-Palencia A, Gomez-Morales M,
Gomez-Capilla JA, Pedraza V, Boyero L, Rosell R and Fárez-Vidal ME:
Gene expression profiling reveals novel biomarkers in nonsmall cell
lung cancer. Int J Cancer. 129:355–364. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hou J, Aerts J, den Hamer B, van Ijcken W,
den Bakker M, Riegman P, van der Leest C, van der Spek P, Foekens
JA, Hoogsteden HC, et al: Gene expression-based classification of
non-small cell lung carcinomas and survival prediction. PLoS One.
5:e103122010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fouret R, Laffaire J, Hofman P,
Beau-Faller M, Mazieres J, Validire P, Girard P, Camilleri-Bröet S,
Vaylet F, Leroy-Ladurie F, et al: A comparative and integrative
approach identifies ATPase family, AAA domain containing 2 as a
likely driver of cell proliferation in lung adenocarcinoma. Clin
Cancer Res. 18:5606–5616. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei TY, Juan CC, Hisa JY, Su LJ, Lee YC,
Chou HY, Chen JM, Wu YC, Chiu SC, Hsu CP, et al: Protein arginine
methyltransferase 5 is a potential oncoprotein that upregulates G1
cyclins/cyclin-dependent kinases and the phosphoinositide
3-kinase/AKT signaling cascade. Cancer Sci. 103:1640–1650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rousseaux S, Debernardi A, Jacquiau B,
Vitte AL, Vesin A, Nagy-Mignotte H, Moro-Sibilot D, Brichon PY,
Lantuejoul S, Hainaut P, et al: Ectopic activation of germline and
placental genes identifies aggressive metastasis-prone lung
cancers. Sci Transl Med. 5:1862013. View Article : Google Scholar
|
|
33
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nagy A, Lanczky A, Menyhart O and Gyorffy
B: Validation of miRNA prognostic power in hepatocellular carcinoma
using expression data of independent datasets. Sci Rep. 8:92272018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ye QH, Qin LX, Forgues M, He P, Kim JW,
Peng AC, Simon R, Li Y, Robles AI, Chen Y, et al: Predicting
hepatitis B virus-positive metastatic hepatocellular carcinomas
using gene expression profiling and supervised machine learning.
Nat Med. 9:416–423. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hoshida Y, Villanueva A, Kobayashi M, Peix
J, Chiang DY, Camargo A, Gupta S, Moore J, Wrobel MJ, Lerner J, et
al: Gene expression in fixed tissues and outcome in hepatocellular
carcinoma. N Engl J Med. 359:1995–2004. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim SM, Leem SH, Chu IS, Park YY, Kim SC,
Kim SB, Park ES, Lim JY, Heo J, Kim YJ, et al: Sixty-five
gene-based risk score classifier predicts overall survival in
hepatocellular carcinoma. Hepatology. 55:1443–1452. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kate RJ and Nadig R: Stage-Specific
predictive models for breast cancer survivability. Int J Med
Inform. 97:304–311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jamal-Hanjani M, Wilson GA, McGranahan N,
Birkbak NJ, Watkins TBK, Veeriah S, Shafi S, Johnson DH, Mitter R,
Rosenthal R, et al: Tracking the evolution of non-small-cell lung
cancer. N Engl J Med. 376:2109–2121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peifer M, Fernandez-Cuesta L, Sos ML,
George J, Seidel D, Kasper LH, Plenker D, Leenders F, Sun R, Zander
T, et al: Integrative genome analyses identify key somatic driver
mutations of small-cell lung cancer. Nat Genet. 44:1104–1110. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bertucci F, Ng CKY, Patsouris A, Droin N,
Piscuoglio S, Carbuccia N, Soria JC, Dien AT, Adnani Y and Kamal M:
Genomic characterization of metastatic breast cancers. Nature.
569:560–564. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vousden KH and Lane DP: P53 in health and
disease. Nat Rev Mol Cell Biol. 8:275–283. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen Z, Trotman LC, Shaffer D, Lin HK,
Dotan ZA, Niki M, Koutcher JA, Scher HI, Ludwig T, Gerald W, et al:
Crucial role of p53-dependent cellular senescence in suppression of
pten-deficient tumorigenesis. Nature. 436:725–730. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Meek DW: Tumour suppression by p53: A role
for the DNA damage response? Nat Rev Cancer. 9:714–723. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gyorffy B, Surowiak P, Budczies J and
Lanczky A: Online survival analysis software to assess the
prognostic value of biomarkers using transcriptomic data in
non-small-cell lung cancer. PLoS One. 8:e822412013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li Y, Huang J, Yang D, Xiang S, Sun J, Li
H and Ren G: Expression patterns of E2F transcription factors and
their potential prognostic roles in breast cancer. Oncol Lett.
15:9216–9230. 2018.PubMed/NCBI
|
|
49
|
Manicum T, Ni F, Ye Y, Fan X and Chen BC:
Prognostic values of E2F mRNA expression in human gastric cancer.
Biosci Rep. 21:382018.
|
|
50
|
Huang YL, Ning G, Chen LB, Lian YF, Gu YR,
Wang JL, Chen DM, Wei H and Huang YH: Promising diagnostic and
prognostic value of E2Fs in human hepatocellular carcinoma. Cancer
Manag Res. 11:1725–1740. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gao Z, Shi R, Yuan K and Wang Y:
Expression and prognostic value of E2F activators in NSCLC and
subtypes: A research based on bioinformatics analysis. Tumour Biol.
37:14979–14987. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Narita M, Nunez S, Heard E, Narita M, Lin
AW, Hearn SA, Spector DL, Hannon GJ and Lowe SW: Rb-Mediated
heterochromatin formation and silencing of E2F target genes during
cellular senescence. Cell. 113:703–716. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Aksoy O, Chicas A, Zeng T, Zhao Z,
McCurrach M, Wang X and Lowe SW: The atypical E2F family member
E2F7 couples the p53 and RB pathways during cellular senescence.
Genes Dev. 26:1546–1557. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ran LK, Chen Y, Zhang ZZ, Tao NN, Ren JH,
Zhou L, Tang H, Chen X, Chen K, Li WY, et al: SIRT6 overexpression
potentiates apoptosis evasion in hepatocellular carcinoma via
BCL2-associated X protein-dependent apoptotic pathway. Clin Cancer
Res. 22:3372–3382. 2016. View Article : Google Scholar : PubMed/NCBI
|