Introduction
Hepatocellular carcinoma (HCC) is the most common
primary malignant tumor of the liver. It is the fifth most common
cancer in the word and the third most common cause of
cancer-related death (1,2). HCC is commonly diagnosed at the
advanced stage due to the asymptomatic features of early HCC, and
patients with advanced HCC have a poor life expectancy with an
average survival time of 7 months (3). Sorafenib, a multi-kinase inhibitor, is
the only treatment option for advanced liver cancer (4). Although the use of sorafenib has
improved the survival rate of patients with liver cancer, treatment
resistance associated with sorafenib has become a major obstacle to
treatment efficacy (2). Therefore,
it is important to determine the molecular mechanisms underlying
sorafenib resistance.
Long non-coding RNAs (lncRNAs) are
non-protein-coding RNAs with more than 200 nucleotides. Many
studies have shown that lncRNAs contribute to the drug resistance
in many types of cancer (5,6). For example, lncRNA H19 contributes to
5-fluorouracil (5-FU) resistance in colorectal cancer by promoting
autophagy (7). Downregulation of
lncRNA HOTAIR was found to increase the sensitivity of cisplatin to
ovarian cancer by inhibiting cisplatin-induced autophagy (8). H19 was demonstrated to mediate the
chemosensitivity of breast cancer cells via the Wnt pathway and
epithelial-mesenchymal transition (EMT) (9). lncRNA UCA1 confers tamoxifen
resistance in endocrine therapy of breast cancer by regulating the
EZH2/p21 axis and the PI3K/AKT signaling pathway (10). lncRNA HULC was found to attenuate
the chemosensitivity of HCC cells by stabilizing Sirt1 to trigger
autophagy (11). Among the lncRNAs,
H19 was first described as an oncofetal transcript more than 30
years ago (12). Since its
discovery, emerging evidence shows that H19 expression is
upregulated in many types of cancer (7). However, research on lncRNA H19 in
sorafenib resistance in HCC is quite limited.
The aim of the present study was to determine the
mechanism of sorafenib resistance and to lay a foundation for
achieving greater treatment efficacy. We focused on the mechanism
by which lncRNA H19 regulates sorafenib resistance in liver cancer.
Our findings provide new clues for further clinical treatment of
patients with sorafenib-resistant liver cancer.
Materials and methods
Cell lines and transfection
Human HCC cell lines (Huh7, Hep3B, SNU-449, SNU-387)
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). Huh7 was cultured in Dulbecco's modified
Eagle's medium (DMEM, Gibco; Thermo Fisher Scientific, Inc.). Hep3B
cells were cultured in Mimimum Essential Medium (MEM) (Gibco;
Thermo Fisher Scientific, Inc.) SNU-387 and SNU-449 cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.). All cells were placed in a humidified incubator containing
5% carbon dioxide (CO2) at 37°C with 10% fetal bovine
serum (FBS, Gibco; Thermo Fisher Scientific, Inc.).
For transfection, H19 siRNA or the negative control
were purchased from GenePharma (Shanghai, China). We used the mix
of three sequences of siRNAs for knockdown. The sequences of the
H19 siRNAs are as follows: H19-Homo-1552: Sense,
5′-CCCACAACAUGAAAGAAAUUU-3′ and antisense,
5′-AUUUCUUUCAUGUUGUGGGUU-3′; H19-Homo-1617: Sense,
5′-CCUCUAGCUUGGAAAUGAAUU-3′ and antisense,
5′-UUCAUUUCCAAGCUAGAGGUU-3′; and H19-Homo-1712: Sense,
5′-UCAUCAGCCCAACAUCAAAUU-3′ and antisense,
5′-UUUGAUGUUGGGCUGAUGAUU-3′; negative control (NC): Sense:
5′-UUCUCCGAACGUGUCACGUUU-3′ and antisense:
5′-ACGUGACACGUUCGGAGAAUU-3′; miR-675 mimic
(5′-UGGUGCGGAGAGGGCCCACAGUG-3′ and 5′-CUGUGGGCCCUCUCCGCACCAUU-3′);
inhibitor (5′-CACUGUGGGCCCUCUCCGCACCA-3′), and negative control
were purchased by RIBO (Guangzhou, China). Transfections were
achieved using Lipofectamine 2000 reagent (Life Technologies;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The transfected HCC cells were collected after 48 h
of transfection for further experiments.
RT-qPCR
Total RNA was extracted from the cultured cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
and RNA concentration was determined by spectrophotometry.
Single-stranded cDNA was synthesized using a cDNA synthesis kit
(Takara) according to the manufacturer's instructions. Reverse
transcription-polymerase chain reaction assays were performed using
Applied Biosystems SYBR Green Mix kits (Applied Biosystems). The
amplification conditions were: 40 cycles of 5 sec at 95°C and 30
sec at 60°C. The 2−ΔΔCq method was used for quantitative
gene expression (13). The
following primers were used: H19-homo-F, 5′-TCCCAGAACCCACAACATGA-3′
and H19-homo-R, 5′-TTCACCTTCCAGAGCCGATT-3′; ACTB-homo-F,
5′-TGGCACCCAGCACAATGAA-3′ and ACTB-homo-R,
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′; Hsa-miR-675:
5′-TGGTGCGGAGAGGGCCCACAGTG-3′.
EdU assay
For the EdU staining assay, cells
(1×105/well) were permeabilized and stained with
Click-iT EdU Imaging Kit (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. Briefly, HCC
cells under different conditions were incubated for 24 h. After
removing the culture medium, the cells were fixed with 1 ml of 3.7%
formaldehyde in phosphate-buffered saline (PBS) for 15 min and
incubated with 0.5% Triton X-100 in PBS for 20 min. Then, the cells
were incubated with 0.5 ml of Click-iT reaction cocktail at room
temperature for 30 min in the dark. After removal of the reaction
cocktail, cells were washed with 3% BSA in PBS followed by PBS.
After addition of DAPI in PBS, cells were incubated for 20 min at
room temperature in the dark. The cells were then washed with PBS,
and images were obtained under an inverted fluorescence microscope
(magnification ×200; Olympus Corp.).
Western blot analysis
Total proteins were extracted using RIPA buffer
(Beyotimes Biotechnology, Guangzhou) and quantified using the
bicinchoninic acid method. Proteins (40 µg/lane) were separated on
a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and then transferred to PVDF membranes (Millipore). The
blots were incubated overnight with the following primary
antibodies: E-cadherin (cat. no. 3195, dilution 1:1,000; Cell
Signaling Technology, Inc.), vimentin (cat. no. 5741, dilution
1:1,000; Cell Signaling Technology, Inc.), and GAPDH (cat. no.
2118, dilution 1:1,000; Cell Signaling Technology, Inc.). After
three washes, the membranes were incubated with a horseradish
peroxidase-conjugated secondary antibody (anti-rabbit-HPR, cat. no.
7074, dilution 1:2,000; Cell Signaling Technology, Inc.) and
visualized by electrochemiluminescence (ECL).
CCK-8 assay
Cell viability was detected using the Cell Counting
Kit-8 (CCK-8) assay (Dojindo). Briefly, cells were seeded at 5,000
cells per well in 96-well plates and cultured for 12 h at 37°C. The
cells were then subjected to different concentrations of sorafenib
for 48 h. Then CCK-8 solution (10 µl/ well) was added to each well
and the cells were incubated for additional 2–4 h at 37°C. The
values of absorbance at 490 nm were measured using a microplate
reader (Bio-Tek Instruments, Inc.).
Statistical analysis
Data are expressed as the mean ± SD. All statistical
analyses were performed using GraphPad 6.0 statistical software
(GraphPad Software, Inc.). Differences between two groups were
analyzed using the Student's t-test. One-way analysis of variance
(ANOVA) with Tukey's post hoc test was used to analyze differences
among multiple groups. P<0.05 was assigned to indicate a
statistical significance.
Results
Expression of H19 correlates with
sorafenib sensitivity in HCC cells
We collected samples from 18 patients with HCC and
examined the expression of H19 by RT-qPCR. We found significant
upregulation of H19 in HCC tissue samples, compared with that noted
in the matched adjacent normal tissues (Fig. S1A). In order to investigate the
role of lncRNA H19 in drug resistance, HCC cells were treated with
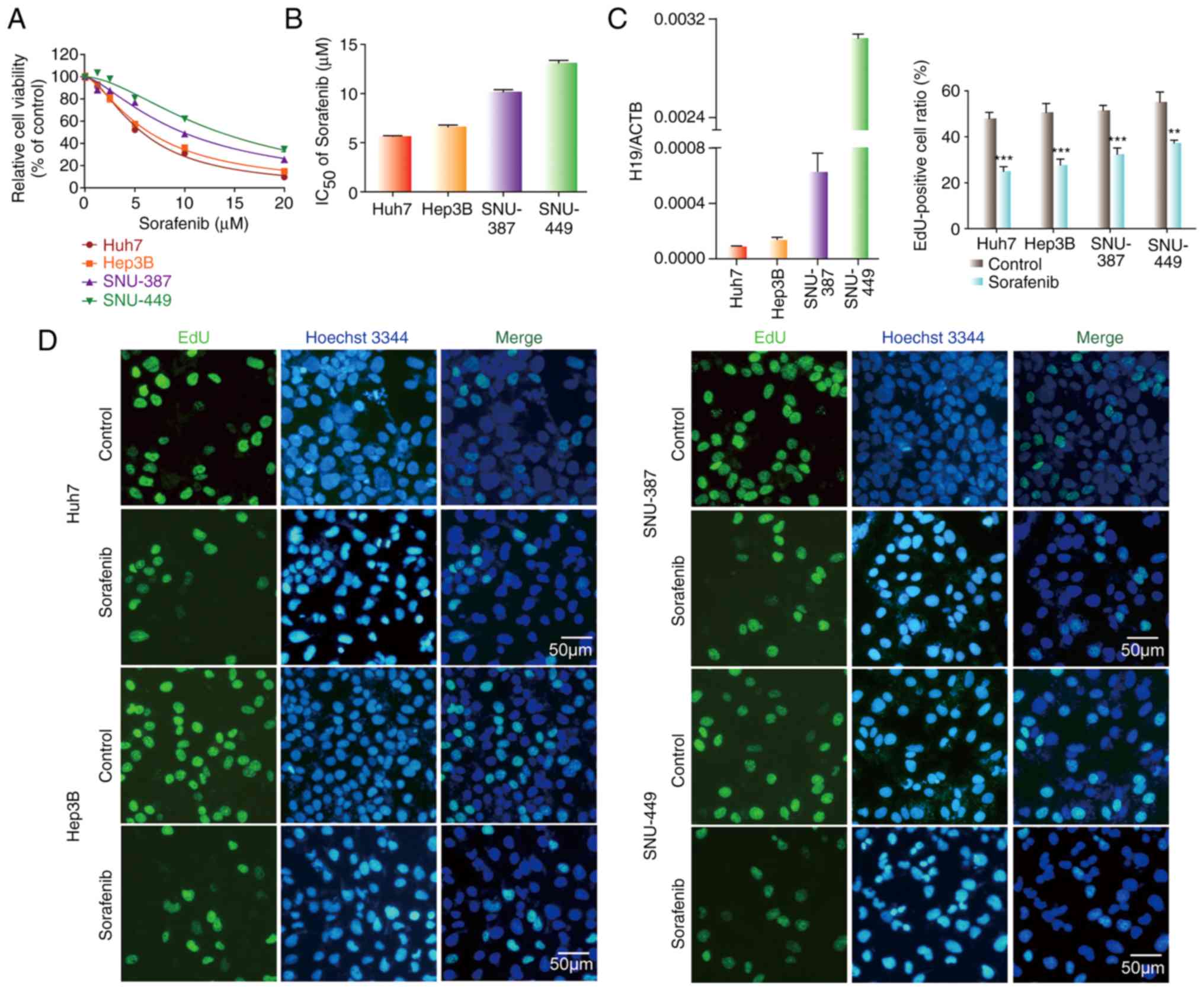
various concentrations of sorafenib for 48 h, and the CCK-8 assay
was then performed. IC50 values of sorafenib in SNU-387
and SNU-449 cells were higher than these values in the Huh7 and
Hep3B cells (Fig. 1A and B).
Meanwhile, we detected the level of H19 in HCC cell lines. The
results revealed that the expression of H19 in SNU-387 and SNU-449
cells was higher than that in the Huh7 and Hep3B cells (Fig. 1C). Compared with the untreated
control, the EdU-positive cell ratio in all four HCC cell lines
exposed to sorafenib was significantly lower (Fig. 1D). The EdU-positive cell ratios in
the SNU-387 and SNU-449 cells with higher H19 expression were less
than that in the Huh7 and Hep3B cells with lower H19 expression.
These results show that H19 expression is negatively related to
sorafenib sensitivity in HCC cells, suggesting that H19 is an
oncogene.
H19 knockdown sensitizes HCC cells to
sorafenib in vitro
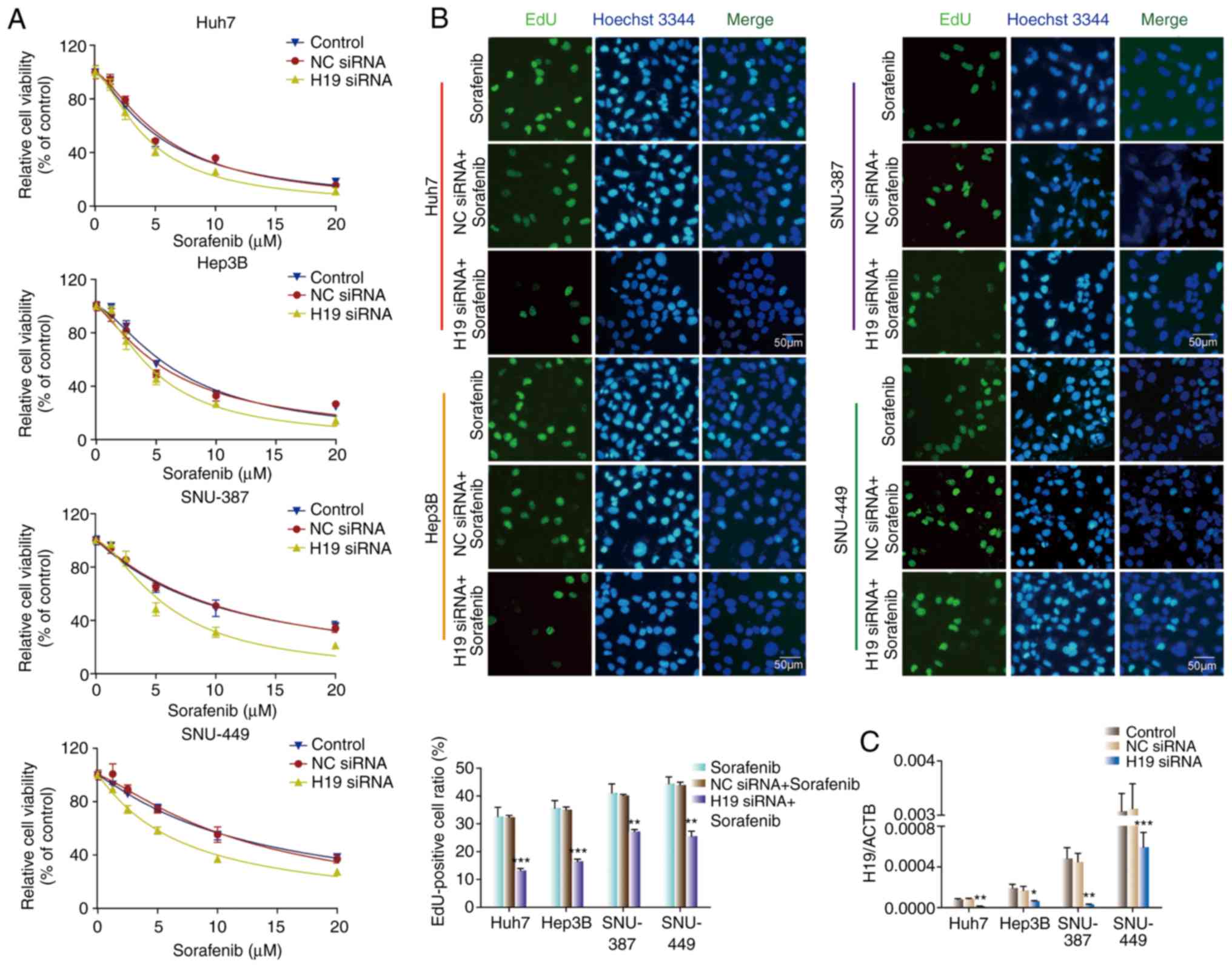
To further investigate the effect of H19 on the
chemoresistance of HCC cells against sorafenib, four HCC cell lines
were transfected with H19 siRNA. CCK-8 and EdU assays were utilized
to evaluate cell proliferative capacity. As shown in Fig. 2A, there was no significant
difference in sensitivity to sorafenib between the control group
and the NC siRNA group for all cell lines. The H19 siRNA groups
showed significantly higher sensitivity to sorafenib than the
control group and the NC siRNA group for all four HCC cell lines.
Consistent with the CCK-8 results, H19 downregulation attenuated
the EdU-positive cell ratio. The EdU-positive cell ratio in the H19
siRNA group was lower than that in the control group and NC siRNA
group in all cell lines (Fig. 2B).
H19 knockdown efficiency was evaluated by RT-qPCR (Fig. 2C). These results suggest that H19
knockdown sensitizes HCC cells to sorafenib in vitro.
H19 knockdown suppresses
epithelial-mesenchymal transition (ETM)
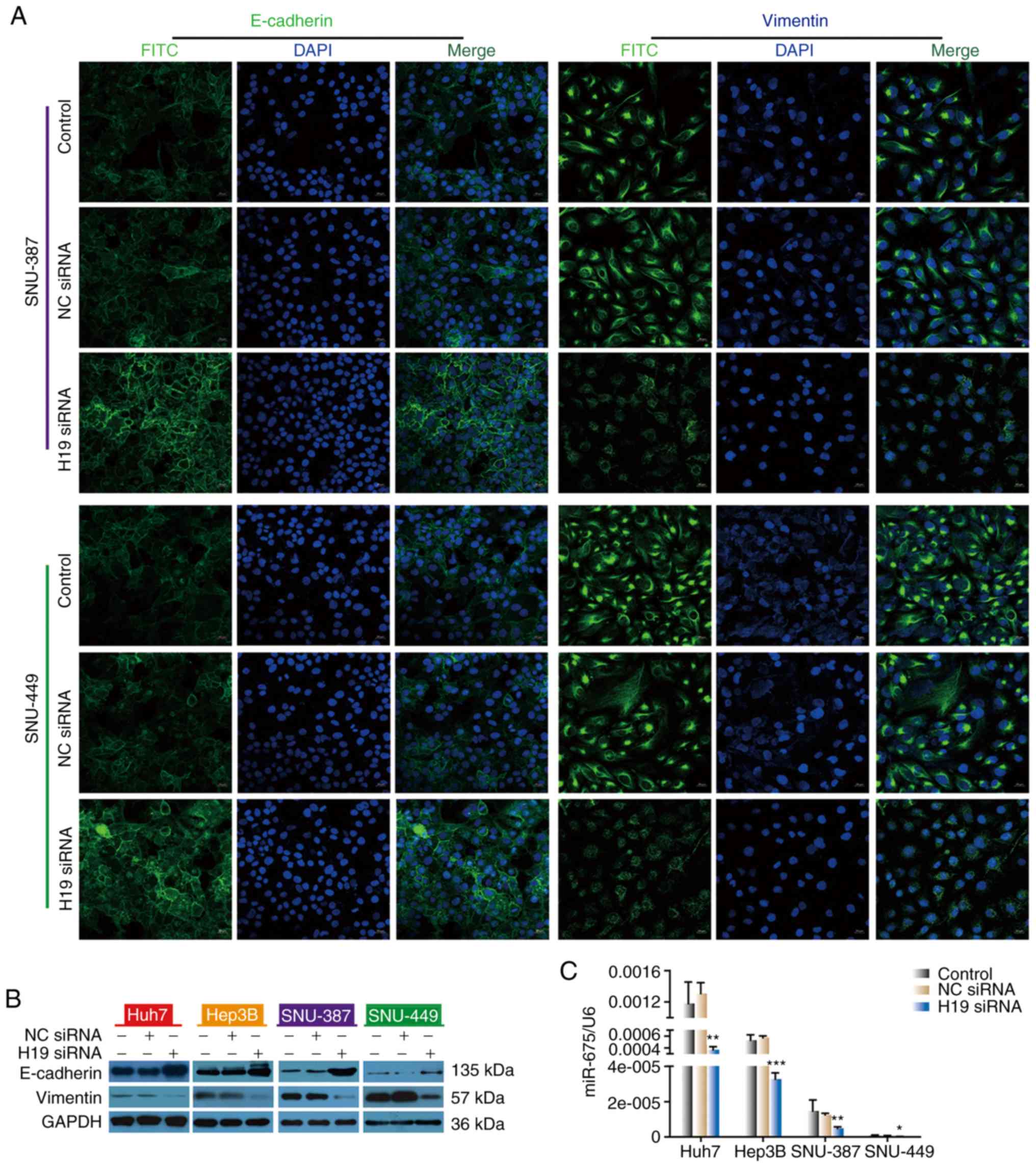
To investigate the mechanism by which H19 knockdown
sensitizes HCC cells to sorafenib, immunofluorescence staining and
western blot analysis were performed. HCC cells were transfected
with NC siRNA or H19 siRNA. We found that siRNA H19-transfected
cells showed stronger E-cadherin immunofluorescence compared with
the untreated control and NC siRNA-transfected cells, while
vimentin immunofluorescence was less than that noted in the
untreated control and NC siRNA-transfected cells (Fig. 3A). Consistent with
immunofluorescence results, western blot analysis showed that
silencing of H19 by siRNA transfection resulted in a significant
increase in E-cadherin and a decrease in vimentin in all four HCC
cell lines examined (Fig. 3B).
Moreover, the results of the migration and invasion assays revealed
that H19 siRNA exerted a distinct inhibitory effect on the
migration distance and cell numbers that had migrated in the four
HCC cell lines (Fig. S2). These
findings indicate that knockdown of lncRNA H19 may enhance
sorafenib sensitivity in HCC cells by inhibiting EMT.
As previously reported, H19 was found to play a role
in breast cancer by upregulating the expression of miR-675
(14). To study the effect of
miR-675 on HCC, we first detected miR-675 expression in the
collected samples. Notably, the expression of miR-675 in HCC
tissues was significantly higher than that in normal tissues
(Fig. S1B). Our results showed
that H19 positively regulates miR-675. We next evaluated the level
of miR-675 in HCC cells after transfection with H19 siRNA. The
level of miR-675 in the siRNA H19-transfected group was
significantly lower than that noted in the control group and NC
group, indicating that miR-675 may be a target gene of lncH19
(Fig. 3C).
miR-675 regulates the sensitivity of
HCC cells to sorafenib
To investigate the function of miR-675 on sorafenib
sensitivity of HCC cells, we transfected the four HCC cell lines
with miR-675 mimic, inhibitor, or negative control. miR-675 mimic
upregulated miR-675 expression while the inhibitor downregulated
its expression (Fig. 4C). CCK-8 and
EdU assays were used to assess the impact of miR-675 on cell
proliferation. As shown in Fig. 4A and
B, miR-675 mimic increased the cell viability and the number of
EdU-positive cells compared with the control and NC groups. In
contrast, miR-675 inhibitor reduced the cell viability and the
number of EdU-positive cells compared with the control and NC
groups. These results showed that miR-675 is involved in the
regulation of cell sensitivity to sorafenib in HCC cells.
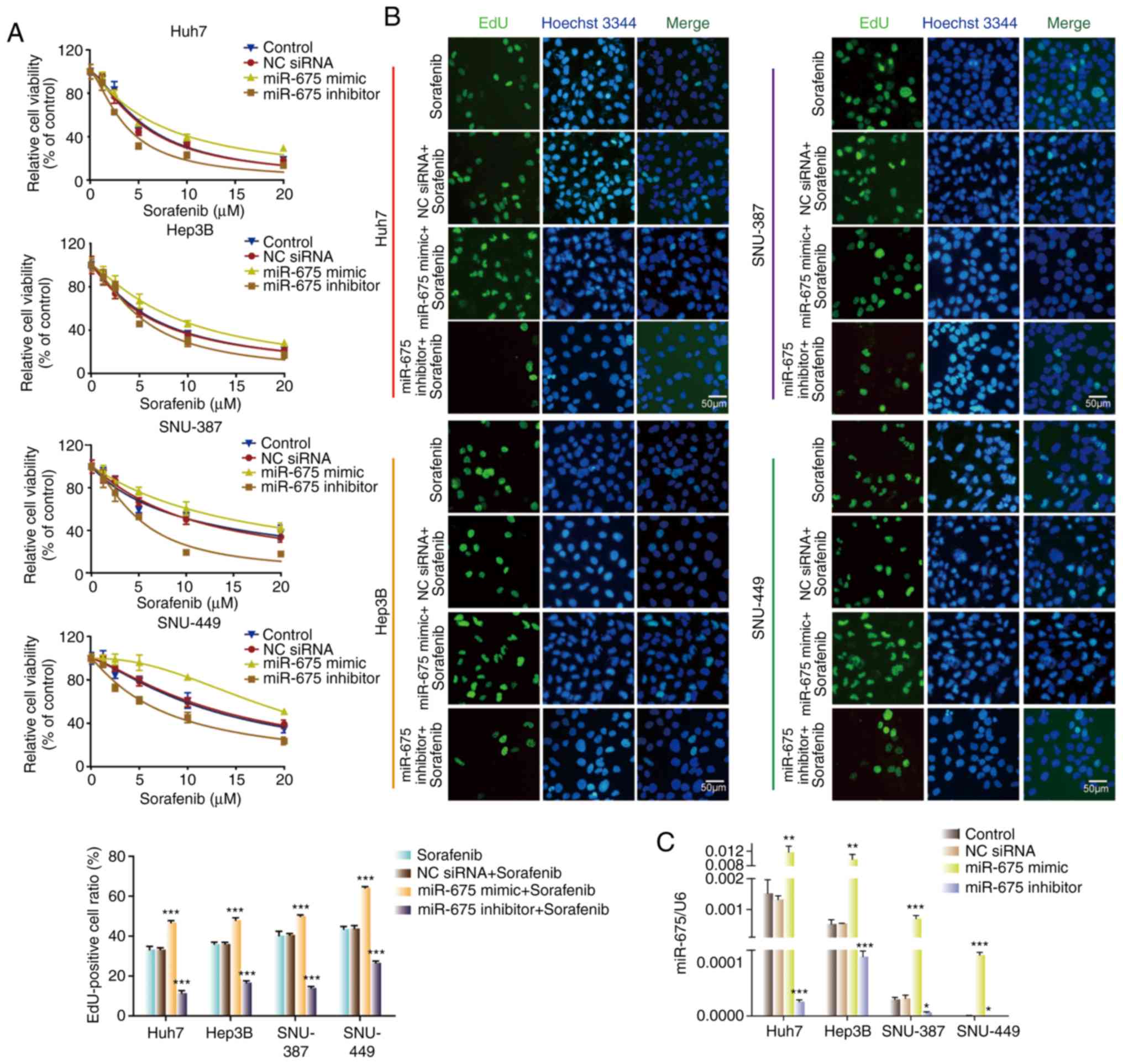 | Figure 4.miR-675 regulates sensitivity of HCC
cells to sorafenib. (A) Sorafenib sensitivity of HCC cells
transfected with NC siRNA, miR-675 mimic, or miR-675 inhibitor. (B)
Proliferation of HCC cells transfected with NC siRNA, miR-675
mimic, or miR-675 inhibitor in the presence of sorafenib.
***P<0.001, compared with the sorafenib group. (C) Expression of
miR-675 in HCC cells transfected with NC siRNA, miR-675 mimic, or
miR-675 inhibitor in the presence of sorafenib. *P<0.05,
**P<0.01, ***P<0.001. HCC, hepatocellular carcinoma; NC,
negative control. |
miR-675 influences
epithelial-mesenchymal transition (ETM)
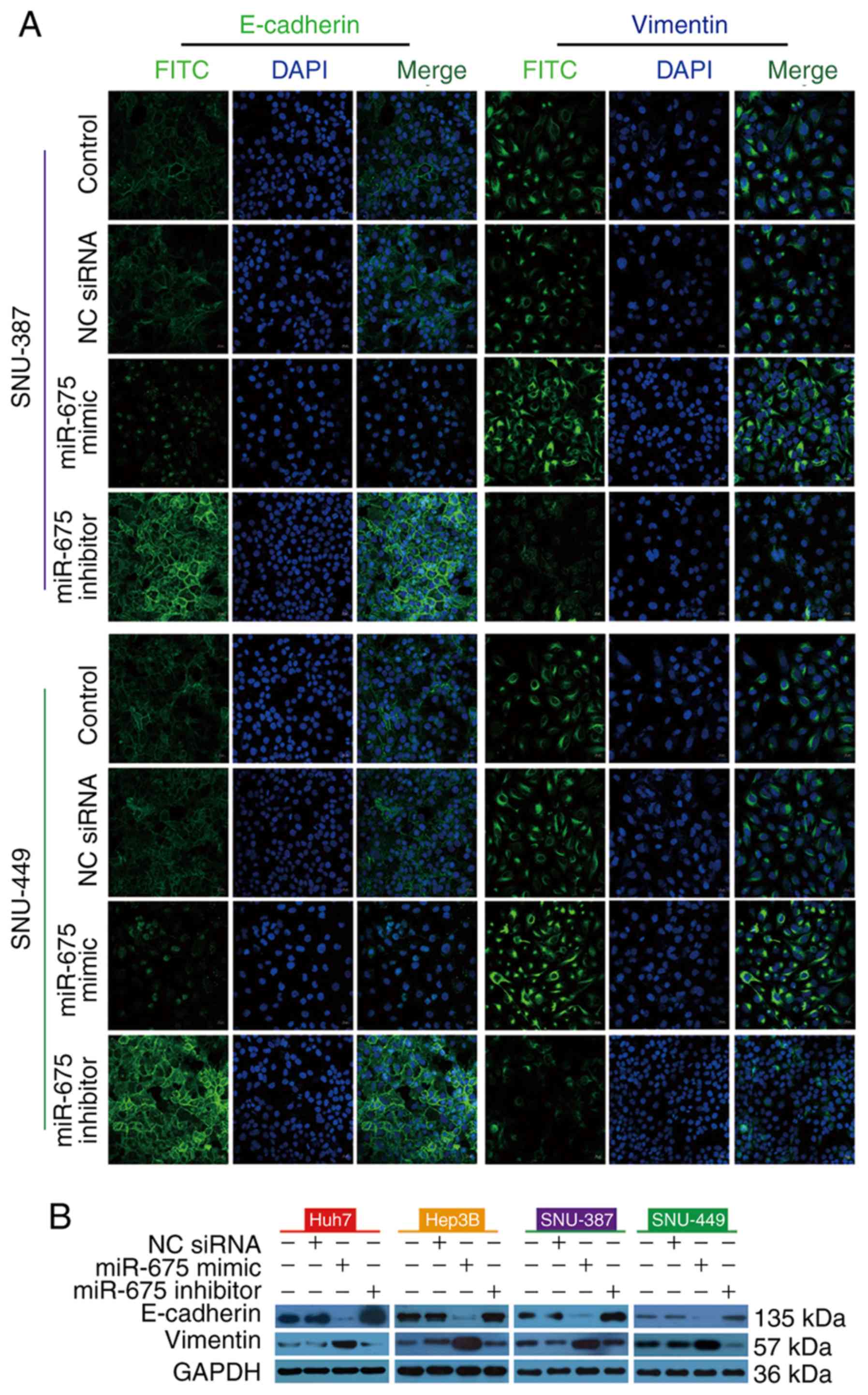
To investigate the effect of miR-675 on EMT, we
transfected HCC cells with miR-675 mimic, inhibitor, or negative
control. Immunofluorescence staining showed that miR-675 mimic
reduced E-cadherin immunofluorescence and enhanced vimentin
immunofluorescence. In contrast, miR-675 inhibitor increased
E-cadherin immunofluorescence and decreased vimentin
immunofluorescence (Fig. 5A).
Consistent with the results of immunofluorescence staining, miR-675
mimic reduced the expression of E-cadherin protein and enhanced the
expression of vimentin protein in the four HCC lines while a
miR-675 inhibitor had the opposite effect (Fig. 5B). Taken together, these findings
suggest that overexpression of miR-675 promotes EMT while
inhibition of miR-675 can suppress EMT.
miR-675 mediates the regulatory effect
of lncRNA H19 on sorafenib sensitivity
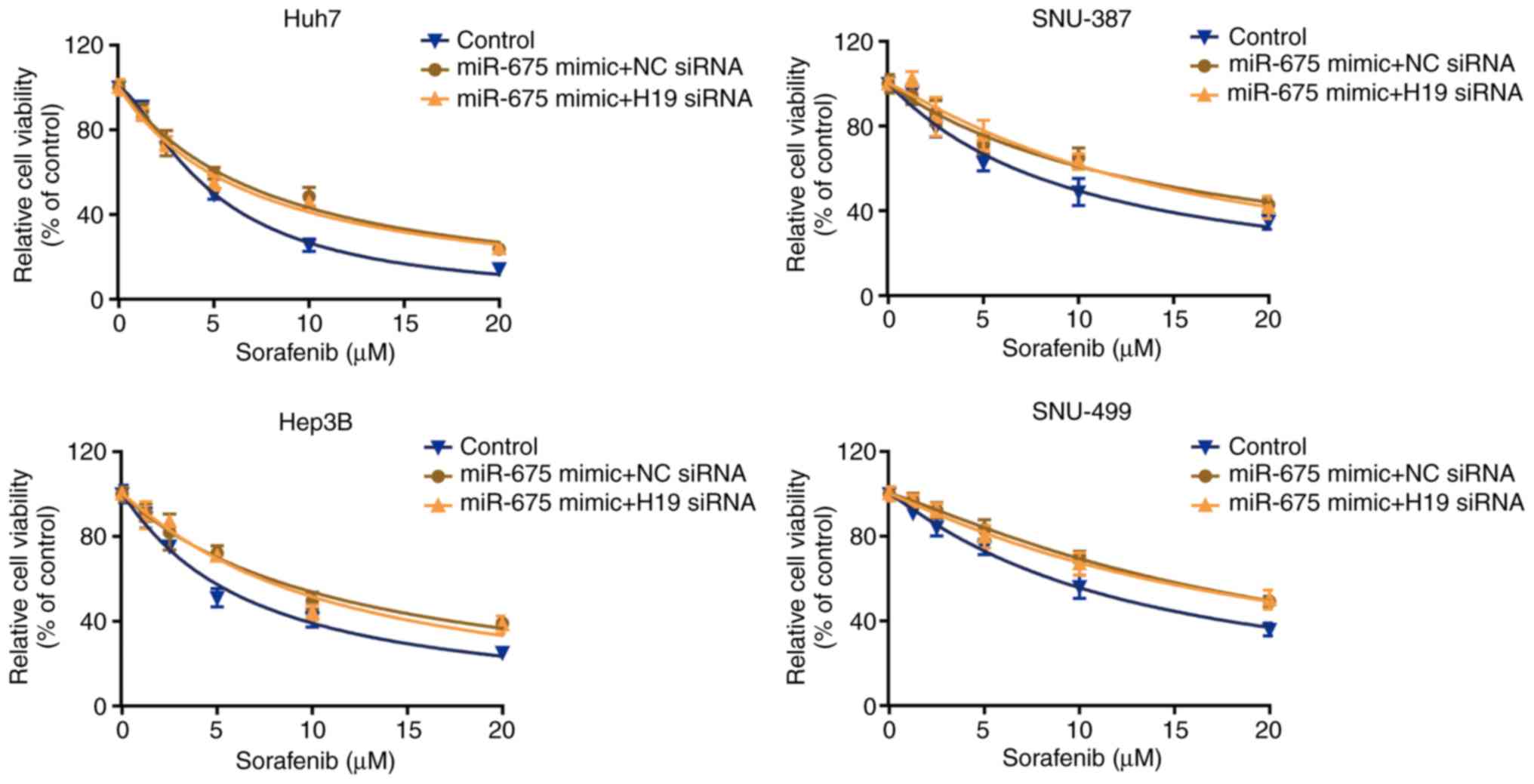
To examine whether miR-675 participates in the
regulatory effect of lncRNA H19 on sensitivity to sorafenib, we
transfected the four cell lines with miR-675 mimic, and then
treated them with H19 siRNA. We found that H19 siRNA did not change
the effect of miR-675 on sorafenib sensitivity, which was confirmed
by CCK-8 assay (Fig. 6). These data
indicate that miR-675 mediates the regulatory effect of lncRNA H19
on sorafenib sensitivity.
Discussion
Our findings revealed that H19 expression is
negatively related to sorafenib sensitivity in hepatocellular
carcinoma (HCC) cells, and that knockdown of H19 could sensitize
HCC cells to sorafenib. Mechanistically, the results demonstrated
that long non-coding RNA (lncRNA) H19 knockdown can suppress
epithelial-mesenchymal transition (EMT) to enhance the sensitivity
of HCC cells to sorafenib. Moreover, we found that knockdown of H19
was associated with an obvious reduction in the expression level of
miR-675 in HCC cells. miR-675 mimic transfection promoted sorafenib
resistance in the HCC cells by enhancing EMT, while the miR-675
inhibitor had the opposite effect. H19 knockdown did not influence
the effect of miR-675 on sorafenib sensitivity in HCC cells.
lncRNA H19 is located on chromosome 11 in humans and
has been characterized as an oncogenic lncRNA in a few types of
cancer due to its promotion of cell proliferation and
chemoresistance (15–17). In the present study, we first found
that H19 expression is negatively related to sorafenib sensitivity
in HCC cells based on CCK-8 and EdU assays. After transfection of
H19 siRNA, we observed that the sensitivity of the liver cancer
cells to sorafenib was significantly increased. These data suggest
that H19 is an oncogene.
Epithelial-mesenchymal transition (EMT) is a
biologic process whereby polarized epithelial cells lose epithelial
characteristics and develop a mesenchymal phenotype. In EMT, cell
adhesion molecules (such as E-cadherin) are lost while mesenchymal
markers (such as vimentin) are induced, leading to the loss of
polarity and enhancement of tumor cell migration and invasion. As a
result, tumor cells become more insensitive to antitumor drugs
(2,18,19).
Many reports have shown that tumor drug resistance is frequently
accompanied by EMT in different types of cancer, including lung
cancer (20), pancreatic cancer
(21), gastric cancer (22), and breast cancer (23). In HCC, long-term exposure of liver
cancer cells to sorafenib can induce epithelial-mesenchymal
transition to develop resistance, increase the risk of invasive and
rebound growth (24). PDCD2
sensitizes HepG2 cells to sorafenib by inhibiting mesenchymal
transformation of epithelial cells (25). Therefore, we hypothesized that
knockdown of H19 would sensitize HCC cells to sorafenib via EMT. To
test this hypothesis, we detected the levels of EMT-related
proteins after silencing of H19. The results showed that E-cadherin
expression was increased and vimentin expression was decreased by
both immunofluorescence and western blot analysis, indicating that
EMT was suppressed.
MicroRNAs (miRNAs) are a class of single-stranded
non-coding RNAs of 18–22 nucleotides that regulate gene expression
at the post-transcriptional level by targeting and degrading
specific mRNAs (26,27). In many cancers, miRNAs can serve as
oncogenes or tumor-suppressor genes and participate in
chemosensitivity. For example, microRNA-31-5p regulates
chemosensitivity by blocking the nuclear location of PARP1 in
hepatocellular carcinoma (27).
miR-20a-5p regulates gemcitabine chemosensitivity by targeting RRM2
in pancreatic cancer cells and serves as a predictor for
gemcitabine-based chemotherapy (28). miR-494 was found to increase the
chemosensitivity to doxorubicin of gastric cancer cells by
targeting phosphodiesterase 4D (29).
Accumulating evidence suggests that H19 may function
as a reservoir of miR-675. Li et al found that miR-675 is
embedded in the first exon of the H19 transcript (30). Smits et al demonstrated that
H19 is the primary miRNA precursor of miR-675 in both humans and
mice (31). Similarly, it was also
reported that transfection with H19 complementary DNA containing
the pri-miR-675 hairpin increased the expression of mature miR-675
in human kidney 293T cells (32).
Therefore, we performed RT-qPCR analysis to detect the level of
miR-675 after H19 silencing, and found that H19 knockdown decreased
the expression of miR-675. We then examined whether miR-675
regulates the sensitivity of HCC cells to sorafenib. Transfection
of miR-675 mimic made HCC cells insensitive to sorafenib, as shown
by CCK-8 and EdU assays. In contrast, miR-675 inhibitor sensitized
HCC cells to sorafenib. Mechanistically, miR-675 alters
chemosensitivity via EMT. Finally, to confirm that H19 regulates
sorafenib resistance through miR-675, we transfected four HCC cell
lines with miR-675 mimic and then treated them with H19 siRNA. The
results showed that there was no significant difference between the
miR-675 group and the H19+miR-675 group.
There are some limitations in the present study.
Firstly, H19 overexpression experiments were previously attempted
but the plasmid was not constructed successfully. Secondly, in
vivo experiments were attempted but it was not possible to
successfully establish the model. We think the limitation of the
technical condition may be the main reason. In addition, we doubt
that the cell viability could be insufficient to initiate
tumorigenesis in vivo. Third, considering there were many
studies to report that H19 is the precursor of miR-675, we simply
verified these findings in HCC but did not investigate a direct
regulatory relationship between H19 and miR-675. Further
investigations may resolve these issues.
In conclusion, this study showed that knockdown of
H19 sensitized HCC cells to sorafenib by downregulating miR-675,
thereby preventing EMT. Thus, these findings provide evidence to
define H19 as a potential therapeutic target for hepatocellular
carcinoma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Natural Science Foundation of Henan Province (nos. 162300410274 and
182300410298), and the National Natural Science Foundation of China
(no. 81273260).
Availability of data and material
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
Conceptualization of the research study was carried
out by FX and WC. The research methodology was designed by JS and
JM. Software analysis of the data was carried out by XL. Validation
of the results was carried out by LY. Data curation was conducted
by ZL and HL. Writing of the original draft preparation was carried
out by YX and writing, review and editing of the manuscript was
done by YL. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Henan Provincial People's Hospital and written
informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu YJ, Zheng B, Wang HY and Chen L: New
knowledge of the mechanisms of sorafenib resistance in liver
cancer. Acta Pharmacol Sin. 38:614–622. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giannini EG, Farinati F, Ciccarese F,
Pecorelli A, Rapaccini GL, Di Marco M, Benvegnù L, Caturelli E,
Zoli M, Borzio F, et al: Prognosis of untreated hepatocellular
carcinoma. Hepatology. 61:184–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lai HH, Li CW, Hong CC, Sun HY, Chiu CF,
Ou DL and Chen PS: TARBP2-mediated destabilization of Nanog
overcomes sorafenib resistance in hepatocellular carcinoma. Mol
Oncol. 13:928–945. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma H, Yuan L, Li W, Xu K and Yang L: The
LncRNA H19/miR-193a-3p axis modifies the radio-resistance and
chemotherapeutic tolerance of hepatocellular carcinoma cells by
targeting PSEN1. J Cell Biochem. 119:8325–8335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang J, Lv B, Su Y, Wang X, Bu J and Yao
L: Exosome-mediated transfer of lncRNA HOTTIP promotes cisplatin
resistance in gastric cancer cells by regulating HMGA1/miR-218
axis. Onco Targets Ther. 12:11325–11338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang M, Han D, Yuan Z, Hu H, Zhao Z, Yang
R, Jin Y, Zou C, Chen Y, Wang G, et al: Long non-coding RNA H19
confers 5-Fu resistance in colorectal cancer by promoting
SIRT1-mediated autophagy. Cell Death Dis. 9:11492018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yu Y, Zhang X, Tian H, Zhang Z and Tian Y:
Knockdown of long non-coding RNA HOTAIR increases cisplatin
sensitivity in ovarian cancer by inhibiting cisplatin-induced
autophagy. J BUON. 23:1396–1401. 2018.PubMed/NCBI
|
|
9
|
Gao H, Hao G, Sun Y, Li L and Wang Y: Long
noncoding RNA H19 mediated the chemosensitivity of breast cancer
cells via Wnt pathway and EMT process. Onco Targets Ther.
11:8001–8012. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Z, Yu D, Li H, Lv Y and Li S: Long
non-coding RNA UCA1 confers tamoxifen resistance in breast cancer
endocrinotherapy through regulation of the EZH2/p21 axis and the
PI3K/AKT signaling pathway. Int J Oncol. 54:1033–1042.
2019.PubMed/NCBI
|
|
11
|
Xiong H, Ni Z, He J, Jiang S, Li X, He J,
Gong W, Zheng L, Chen S, Li B, et al: LncRNA HULC triggers
autophagy via stabilizing Sirt1 and attenuates the chemosensitivity
of HCC cells. Oncogene. 36:3528–3540. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pachnis V, Belayew A and Tilghman SM:
Locus unlinked to alpha-fetoprotein under the control of the murine
raf and Rif genes. Proc Natl Acad Sci USA. 81:5523–5527. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Müller V, Oliveira-Ferrer L, Steinbach B,
Pantel K and Schwarzenbach H: Interplay of lncRNA H19/miR-675 and
lncRNA NEAT1/miR-204 in breast cancer. Mol Oncol. 13:1137–1149.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding D, Li C, Zhao T, Li D, Yang L and
Zhang B: LncRNA H19/miR-29b-3p/PGRN axis promoted
epithelial-mesenchymal transition of colorectal cancer cells by
acting on Wnt signaling. Mol Cells. 41:423–435. 2018.PubMed/NCBI
|
|
16
|
Lei Y, Guo W, Chen B, Chen L, Gong J and
Li W: Tumor-released lncRNA H19 promotes gefitinib resistance via
packaging into exosomes in non-small cell lung cancer. Oncol Rep.
40:3438–3446. 2018.PubMed/NCBI
|
|
17
|
Duan S, Li M, Wang Z, Wang L and Liu Y:
H19 induced by oxidative stress confers temozolomide resistance in
human glioma cells via activating NF-κB signaling. OncoTargets
Ther. 11:6395–6404. 2018. View Article : Google Scholar
|
|
18
|
Maheswaran T and Rushbrook SM:
Epithelial-mesenchymal transition and the liver: Role in
hepatocellular carcinoma and liver fibrosis. J Gastroenterol
Hepatol. 27:418–420. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukuda K, Takeuchi S, Arai S, Katayama R,
Nanjo S, Tanimoto A, Nishiyama A, Nakagawa T, Taniguchi H, Suzuki
T, et al: Epithelial-to-mesenchymal transition is a mechanism of
ALK inhibitor resistance in lung cancer independent of ALK mutation
status. Cancer Res. 79:1658–1670. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gaianigo N, Melisi D and Carbone C: EMT
and treatment resistance in pancreatic cancer. Cancers (Basel).
9:E1222017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun J, Xu Z, Lv H, Wang Y, Wang L, Ni Y,
Wang X, Hu C, Chen S, Teng F, et al: eIF5A2 regulates the
resistance of gastric cancer cells to cisplatin via induction of
EMT. Am J Transl Res. 10:4269–4279. 2018.PubMed/NCBI
|
|
23
|
Dong H, Hu J, Zou K, Ye M, Chen Y, Wu C,
Chen X and Han M: Activation of LncRNA TINCR by H3K27 acetylation
promotes trastuzumab resistance and epithelial-mesenchymal
transition by targeting MicroRNA-125b in breast cancer. Mol Cancer.
18:32019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van Malenstein H, Dekervel J, Verslype C,
Van Cutsem E, Windmolders P, Nevens F and van Pelt J: Long-term
exposure to sorafenib of liver cancer cells induces resistance with
epithelial-to-mesenchymal transition, increased invasion and risk
of rebound growth. Cancer Lett. 329:74–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu H, Wang M, Liang N and Guan L: PDCD2
sensitizes HepG2 cells to sorafenib by suppressing epithelial
mesenchymal transition. Mol Med Rep. 19:2173–2179. 2019.PubMed/NCBI
|
|
26
|
Huang XH, Wang Q, Chen JS, Fu XH, Chen XL,
Chen LZ, Li W, Bi J, Zhang LJ, Fu Q, et al: Bead-based microarray
analysis of microRNA expression in hepatocellular carcinoma:
MiR-338 is downregulated. Hepatol Res. 39:786–794. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Que KT, Zhou Y, You Y, Zhang Z, Zhao XP,
Gong JP and Liu ZJ: MicroRNA-31-5p regulates chemosensitivity by
preventing the nuclear location of PARP1 in hepatocellular
carcinoma. J Exp Clin Cancer Res. 37:2682018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu H, Lu S, Yang D, Zhang L, Ye J, Li M
and Hu W: MiR-20a-5p regulates gemcitabine chemosensitivity by
targeting RRM2 in pancreatic cancer cells and serves as a predictor
for gemcitabine-based chemotherapy. Biosci Rep. 39:BSR201813742019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng QP, Du DB, Ming Q, Hu F, Wu ZB and
Qiu S: MicroRNA 494 increases chemosensitivity to doxorubicin in
gastric cancer cells by targeting phosphodiesterases 4D. Cell Mol
Biol (Noisy-le-grand). 64:62–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li DY, Busch A, Jin H, Chernogubova E,
Pelisek J, Karlsson J, Sennblad B, Liu S, Lao S, Hofmann P, et al:
H19 induces abdominal aortic aneurysm development and progression.
Circulation. 138:1551–1568. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smits G, Mungall AJ, Griffiths-Jones S,
Smith P, Beury D, Matthews L, Rogers J, Pask AJ, Shaw G, VandeBerg
JL, et al: Conservation of the H19 noncoding RNA and H19-IGF2
imprinting mechanism in therians. Nat Genet. 40:971–976. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cai X and Cullen BR: The imprinted H19
noncoding RNA is a primary microRNA precursor. RNA. 13:313–316.
2007. View Article : Google Scholar : PubMed/NCBI
|