Cancer metastasis has always been a challenge in the
clinic, and is largely responsible for treatment failure and high
mortality. It is known that invasive tumors undergo the EMT process
where cells fail to maintain an epithelial phenotype and acquire a
mesenchymal phenotype, thus transmitting from the primary tumor to
other locations and forming secondary growths (1). To solve this problem, scientists have
dedicated themselves to exploring the molecular mechanisms of this
process. After four decades of work, various signaling pathways are
known to participate in EMT including the transforming growth
factor-β (TGF-β), Wnt/β-catenin, Hedgehog (Hh), Notch, fibroblast
growth factor receptors (FGFRs), and nuclear factor kappa B (NF-κB)
signaling pathways (2). Of these,
the connections of cell adhesion, the Wnt/β-catenin pathway and EMT
are more clearly studied.
The formation of adherens junctions needs cell-cell
adhesion molecules, such as the cadherin superfamily, and nectins
(3). Cadherins and nectins bind to
their anchoring proteins, catenins and afadin, respectively, to
form functional modules that affect the actin cytoskeleton
(4). The nectin family, which is
composed of four members, namely, nectin-1, −2, −3, and −4
(5), is involved in cell-cell
adhesion in various cell types by forming a nectin-afadin complex
(6,7). Previous findings have shown the
significant role of the E-cadherin/β-catenin complex in maintaining
stabilized cell-cell junctions (8).
E-cadherin is regarded as the key component of the adherens
junction complex (9), and invasive
tumor is characterized by a marked decrease in E-cadherin
expression (10). Several
transcription factors such as Twist, Snail and zinc finger
E-box-binding homeobox 1/2 (ZEB1/2) trigger EMT by directly binding
to the E-box sequences of E-cadherin promoter, thus repressing its
transcription (11). For example,
Snail1 and Snail2 bind to CDH1 (gene of E-cadherin)
promoter-based E-box DNA sequences and summon the polycomb
repressive complex 2 (PRC2), resulting in CDH1 histone methylation
and acetylation (12). Activation
of these transcription factors is attributed to the translocation
of β-catenin from the cytoplasm to the nucleus, which is considered
to be the central event in EMT (13). β-catenin has demonstrated its
crucial role in Wnt signal transduction (14). In the presence of Wnt signals, the
phosphorylation of β-catenin by glycogen synthase kinase 3β (GSK3β)
is inhibited, followed by β-catenin disassembly from the
destruction complex and accumulation in the cytoplasm (15). Therefore, current research primarily
focuses on the canonical Wnt signaling (β-catenin dependent)
pathway in which the mechanisms are more clearly established.
MiRNAs are small noncoding molecules with 19–25
nucleotides, which regulate gene expression at the
post-transcriptional level by inhibiting mRNA translation or
facilitating mRNA degradation (16). Previous findings suggested that EMT
is regulated by miRNAs through alteration of the Wnt/β-catenin
pathway (17). However, the complex
role of miRNA as an EMT inhibitor or promoter and its underlying
mechanism need further clarification.
In this review, we focused on the interaction
between miRNAs and the Wnt signaling pathway. Through literature
retrieval, we summarized the distinct effects of miRNAs on Wnt
signaling in the regulation of cancer metastasis, aiming to
identify the mechanism of EMT regulation by miRNAs and potential
therapeutic targets in invasive tumor treatment.
EMT comprises an essential biological process during
which cells fail to maintain epithelial cell polarity and acquire
the mesenchymal phenotype, thus increasing cell motility and
invasion (18). EMT was reported to
be involved in numerous biological activities such as embryogenesis
(19), heart-valve (20) and neural crest formation (21). Scientists categorize EMT into three
types including embryonic development and organ formation, wound
healing and organ fibrosis, and cancer progression (22). The critical role of EMT in cancer
has been extensively studied in recent years. It is generally
accepted that EMT facilitates the invasion and metastasis of early
stage tumors and contributes to cancer progression (23). The latest studies reveal that
EMT-induced tumor progression is not only mediated by phenotypic
change but also related to stemness (24), immune evasion (25), metabolic reprogramming (26), and therapeutic resistance (27) of cancer cells.
EMT is characterized by decreased expression of
epithelial markers such as E-cadherin, γ-catenin and increased
expression of mesenchymal markers such as N-cadherin, vimentin,
Snail, Twist and ZEB (28).
E-cadherin, a pivotal transmembrane adhesion protein in maintaining
cell-cell junctions and polarity, was confirmed to stabilize cell
junctions through forming the E-cadherin/β-catenin complex
(8). The loss of E-cadherin, which
contributes to the mesenchymal phenotype of cancer cells, is a
basic event in tumor metastasis (10). As a result, the newly transformed
mesenchymal cells detach from the primary tumor, invade into the
circulation, and reform into epithelial cells through MET (29), thus leading to tumor formation at a
distant secondary site (30).
EMT is regulated by various signaling pathways such
as the TGF-β, Wnt/β-catenin, Hedgehog and Notch signaling pathway
(18). These pathways trigger EMT
by stimulating transcription factors including Snail, Twist, and
ZEB1/2, which directly bind to the promoter-based E-box DNA
sequences of E-cadherin and repress its transcription. In addition,
Snail also facilitates the transcription of mesenchymal markers
such as vimentin and N-cadherin (31). Among all the signaling pathways, the
Wnt/β-catenin pathway shows its pivotal role in the regulation of
EMT.
Wnt (wingless and Int-1) signals are evolutionarily
conserved consisting of secreted Wnt ligands, Frizzled (FZD)
receptors and coreceptors, intracellular adaptors, and scaffolding
proteins (32). The foremost roles
of the Wnt signaling pathway in cell proliferation,
differentiation, adhesion, invasion, migration, and stem cell
self-renewal have been well established (17). Abnormal Wnt signaling is commonly
correlated with multiple types of disease such as neural tube
defects (33), rheumatoid arthritis
(34), hepatic fibrosis (35), and cardiovascular disease (36). The Wnt signaling pathway is divided
into two categories, a canonical pathway (β-catenin-dependent) and
noncanonical pathway (β-catenin-independent), both of which are
closely related to EMT (37).
β-catenin is regarded as a key protein in Wnt signaling, since
accumulation of β-catenin in the cytoplasm gives rise to its
translocation and activation in the nucleus (15), further initiating the transcription
of EMT-related genes (38).
When the Wnt signal is absent, β-catenin forms a
destruction complex with Axin, adenomatous polyposis coli (APC),
casein kinase I α (CKIα) and GSK3β (39). In this stage, β-catenin is
phosphorylated by GSK3β and CKIα, forming β-catenin degradation by
ubiquitination (40). In addition,
Wnt signaling inhibitory factors such as Dickkopf (DKK) family,
secreted frizzled-related protein (SFRP) family and Wnt inhibitory
factor 1 (WIF1) contribute to the inactive status of β-catenin
(41). Dkk, a small family of
secreted glycoproteins, is comprised of four members, Dkk1-4. Dkk1
and Dkk2 bring about Wnt signal inhibition by binding to
low-density lipoprotein receptor-related protein (LRP) 5/6 with
high affinity (42). However, Dkk2
plays a dual role as an inhibitor or activator of the Wnt signaling
pathway, depending on the cellular context (43,44).
Dkk3 was reported to be different from other members of the DKK
family as it does not bind LRP6 and does not affect Wnt signaling
(45). In addition, there are some
studies demonstrating the inhibitory effects of Xenopus
Cerberus and Wise proteins on Wnt (46). Xenopus Cerberus binds to Wnt
proteins via independent sites to inhibit the Wnt signaling pathway
(47), whereas Wise may inhibit or
activate Wnt signaling in a context-dependent manner (48). In addition to Wnt signaling
inhibitory factors, E-cadherin also suppresses β-catenin by forming
the E-cadherin/β-catenin complex to prevent nuclear translocation
of β-catenin (49). When receptors
receive Wnt ligands such as Wnt1 and Wnt3 binding to the FZD and
LRP 5/6, LRP 5/6 and FZD form a complex to affect the stabilization
of β-catenin and prevent its degradation, resulting in β-catenin
accumulation in cytoplasm (50). As
a consequence, β-catenin translocates to the nucleus and forms a
complex with the T-cell factor/lymphoid enhancer factor (TCF/LEF),
which promotes transcription of Wnt target genes including
Twist, Snail and other oncogenes such as Cyclin D1,
matrix metalloproteniase-7 (MMP-7) and cellular
myelocytomatosis oncogene (c-Myc) (51), thus facilitating EMT (52).
MicroRNAs are small noncoding molecules with 19–25
nucleotides that play fundamental roles in almost every cellular
process such as cell differentiation and homeostasis (53) by regulating gene expression at the
post-transcriptional level (54).
MiRNA genes are transcribed into primary miRNA (pri-miRNA) by RNA
polymerase II (55). Exportin 5
recognizes the 2-nucleotide overhang of the pre-miRNA and
transports it to the cytoplasm, then pre-miRNA undergoes multistep
biogenesis to become mature miRNAs (56). MiRNAs bind to the 3′-untranslated
region (UTR) of target mRNAs to suppress their translation or
accelerate degradation. It is reported that approximately 10–40% of
mRNAs are regulated by miRNAs in humans (57) and dysregulation of miRNA may result
in tumor metastasis (58). Research
has increasingly focused on the interaction between miRNA and EMT
as miRNAs affect multiple EMT-related signaling pathways such as
Wnt/β-catenin, North, TGF-β pathway and their target genes
(59). The role of miRNA as a tumor
suppressor or oncogene during EMT has attracted much attention.
Elucidating miRNA functions in the regulation of EMT may contribute
to the finding of potential therapeutic targets.
A large number of miRNAs were found to be
downregulated in a wide spectrum of cancers (60,61),
indicating their inhibitory effect on tumorigenesis and
development. Furthermore, clinicopathological analysis also
revealed that tumor migration and invasion was negatively
correlated with a number of miRNAs (62). Numerous studies reported that miRNAs
may function as EMT inhibitors by targeting the Wnt signaling
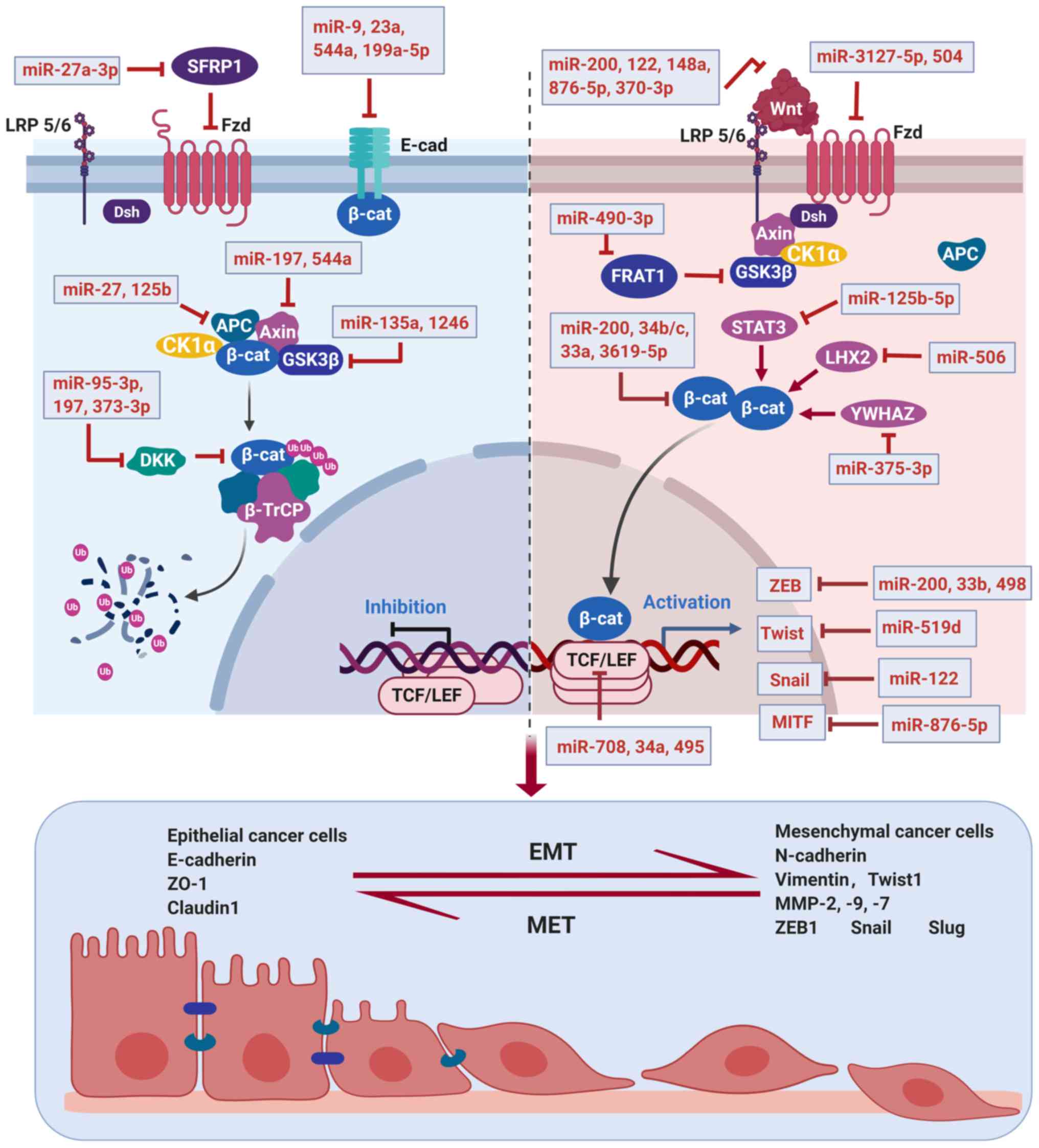
pathway or its downstream transcription factors (Table I, Fig.
1); thus, overexpression of these miRNAs may be a therapeutic
method to reverse EMT.
The EMT-induction transcription factors, most of
which are downstream of the Wnt signaling pathway, have been
studied extensively, including E-cadherin suppressors such as ZEB,
Twist and Snail, which are considered to be regulated by miRNAs in
various types of cancer (10).
The miR-200 family, comprising of 5 miRNA sequences
(miR-200a, miR-200b, miR-200c, miR-141 and miR-429), is believed to
play a significant role in EMT (63). The EMT initiated in several types of
cancer has been shown to be correlated with the underexpression of
the miR-200 family such as bladder cancer (64), breast cancer (65), melanoma (66), ovarian cancer (67), gastric cancer (68), and prostate cancer (69). Gregory et al found the levels
of the miR-200 family were significantly reduced following
TGF-β-mediated EMT in invasive breast cancer since the low level of
miR-200 leads to the absence of E-cadherin, indicating miR-200 as a
negative regulator of EMT (70). A
mechanism study demonstrated that miR-200 inhibits Wnt signaling by
targeting transcription factors ZEB1/2 and binding to β-catenin
mRNA to suppress its translation (71). The ZEB family, containing two
members ZEB1 and ZEB2, binds to the promoter-based E-box DNA
sequences of E-cadherin thus repressing its transcription and
facilitating the activation of mesenchymal genes (72). The inhibitory effects of miR-200 on
β-catenin and ZEB1/2 were further confirmed in gastric
adenocarcinoma (73,74), colonic adenocarcinoma (75), and hepatocellular carcinoma (HCC)
(76). Overexpression of miR-200
results in E-cadherin upregulation by targeting ZEB1 and ZEB2,
thereby inhibiting EMT and restoring the epithelial phenotype of
cancer cells (77). However,
components of Wnt signaling can inversely affect the miR-200
family. Tian et al reported that a downstream target of Wnt
signaling, Achaete scute-like 2 (Ascl2) negatively regulates
miR-200 family expression, thus inhibition of Ascl2 obviously
restores miR-200 expression and suppresses EMT, making Ascl2 a
promising target to reverse EMT (75). In addition to EMT inhibition,
restoring the level of miR-200 can also induce cancer cell
apoptosis and increase the sensitivity of cancer cells to
chemotherapeutic drugs. For instance, niclosamide potentially
induces the apoptosis of colon cancer cells by upregulating the
miR-200 family members (78).
Another study demonstrated that the overexpression of miR-200b
could inhibit cell proliferation and enhance apoptosis and then
reverse docetaxel chemoresistance of lung adenocarcinoma (LAD)
cells by directly targeting E2F transcription factor 3 (E2F3),
which were also verified in tissues of LAD patients (79).
In addition to the miR-200 family, miR-33b binds to
3′-UTR of ZEB1 and inhibits ZEB1 expression in lung adenocarcinoma
cells, thus blocking Wnt/β-catenin signaling and suppressing tumor
growth and EMT in vitro and in vivo (80). Research by Zhang et al
identified miR-498 which was downregulated in liver cancer, and
suppressed the growth and metastasis of liver cancer cells partly
by directly targeting ZEB2, making miR-498 a potential biomarker
for diagnosis and a promising therapeutic target for liver cancer
treatment (81). Snail and Twist
are also targeted by miRNAs in the regulation of EMT. Yue et
al demonstrated that miR-519d directly binds to 3′-UTR of
Twist1 to facilitate its degradation in gastric cancer cells,
suggesting miR-519d as a potential therapeutic target for gastric
cancer treatment (82). Jin et
al reported that miR-122 inhibits EMT in HCC by targeting
Snail1 and Snail2 to suppress the Wnt/β-catenin signaling pathway
(83).
The translocation of β-catenin in the nucleus is
followed by the activation of TCF/LEF-Legless-Pygo DNA binding
proteins, which triggers transcription of many oncogenes such as
extracellular matrix receptor III (CD44), c-Myc,
MMP-7, and cyclin D1 (52). LEF1, a pivotal transcription factor
in the Wnt signaling pathway, was reported to be a target of
miR-708 and miR-34a by directly binding to 3′-UTR of LEF1,
suggesting miR-708 and miR-34a as EMT inhibitors in melanoma and
prostate cancer, respectively (84,85).
Pygopus2 (Pygo2) is regarded as a tumor promoter in various types
of cancer due to its combination with free β-catenin to cause
abnormal activation of downstream oncogenes (86). A study demonstrated that miR-516a-3p
inhibits breast cancer cell growth, metastasis and EMT by binding
to 3′-UTR of Pygo2 mRNA, resulting in blockage of the Pygo2/Wnt
pathway (87). In addition,
transcription factor 4 (TCF4) has been found to promote the
occurrence and development of several cancers by recognizing
β-catenin to initiate the transcription of Wnt target genes
(38). MiR-495 was reported to bind
to the 3′-UTR of TCF4, thereby suppressing the migration, invasion,
and proliferation of non-small-cell lung cancer (NSCLC) by
inactivating the Wnt/β-catenin pathway (88).
Some other transcription factors were also reported
to be affected by miRNA in the regulation of Wnt/β-catenin
signaling. Melanogenesis-associated transcription factor (MITF), as
one of the representative target genes of β-catenin, plays a
carcinogenic role in gastric cancer. A study reported that
miR-876-5p was able to bind to 3′-UTR of MITF and downregulate its
expression, thus suppressing viability and migration of gastric
cancer cells, and inducing cell apoptosis (89). Octamer-binding protein 4 (Oct4), an
octamer motif-binding transcription factor, has been confirmed to
exhibit an oncogenic effect in several types of cancer (90,91). A
study by Ling et al demonstrated that miR-145 suppresses EMT
in lung cancer cells by targeting Oct4 to inactivate the
Wnt/β-catenin signaling pathway (92).
Accumulation of β-catenin in cytoplasm is the
central event in Wnt signaling activation; thus, miRNAs targeting
β-catenin may act as EMT inhibitors. In addition to the miR-200
family mentioned above, miR-34b/c suppress β-catenin mRNA
expression by targeting the 3′-UTR of β-catenin in prostate cancer
(93). MiR-3619-5p directly binds
to 3′-UTR of β-catenin and causes its downregulation in bladder
carcinoma (94). Similarly, miR-33a
targets the 3′-UTR of β-catenin to block EMT in human pancreatic
cancer cells (95).
It is generally recognized that Wnt ligands are
regulated by various miRNAs. For example, Wnt1 is a direct target
gene of miR-122 in HCC HepG2 and Huh7 cell lines, thus
downregulation of miR-122 facilitates EMT in HCC cells by
activating Wnt signaling (96).
Another study confirmed that Wnt1 is a target gene of miR-148a in
HCC cells, suggesting miR-148a acts as an HCC metastasis suppressor
by blocking the Wnt signaling pathway (97). In addition, Peng et al
reported that miR-148a suppresses EMT and invasion of pancreatic
cancer cells by targeting Wnt10b and inhibiting the Wnt signaling
pathway, making miR-148a a novel therapeutic target for pancreatic
cancer treatment (98). Wnt5A was
found to be targeted by miR-876-5p, which suppresses the viability
and migration of gastric cancer cells and induces cell apoptosis
(89). Moreover, Wnt7a, which
activates Wnt signaling to promote EMT of bladder cancer, can be
inhibited by miR-370-3p (99).
Wnt ligands transduce signals by binding to several
receptors such as FZD and LRP5/6, this process was described to be
regulated by miRNAs. When Wnt ligands bind to receptors, FZD and
LRP5/6 form a complex on the surface of the cell membrane. Then,
Dsh protein is recruited and constitutes a complex with Axin, which
binds GSK3β and CKIα to release β-catenin, thus forming β-catenin
accumulation in the cytoplasm (50). MiR-3127-5p was reported to block
Wnt/β-catenin signaling by directly targeting FZD4 in NSCLC
(100). Moreover, miR-504
negatively regulates the Wnt/β-catenin pathway by directly
targeting FZD7, thus suppressing the mesenchymal phenotype of
glioblastoma (101).
The phosphorylation of β-catenin by GSK3 is
necessary for β-catenin degradation when the Wnt signal is absent.
Researchers found that proto-oncogene frequently rearranged in
advanced T-cell lymphomas 1 (FRAT1) belongs to the GSK3-binding
protein family, which inhibits GSK3-mediated phosphorylation of
β-catenin and positively regulates the Wnt signaling pathway
(102). MiR-490-3p is identified
to directly target FRAT1, suggesting its tumor suppressive effects
(103). SLC39A7 (ZIP7), a zinc
transporter essential for the activation of tyrosine kinase, is
considered to be a potential target of Wnt/β-catenin. A study by
Cui et al suggested that miR-15a-3p suppresses prostate
cancer by targeting SLC39A7 to inhibit the Wnt/β-catenin signaling
pathway (104). In addition,
Nimmanon et al reported that activation of SLC39A7 drives
the PI3K/Akt pathway (105). Thus,
targeting SLC39A7 by miR-15a-3p to suppress cancer cell progression
may also result from inhibition of the PI3K/Akt pathway.
MiRNAs may regulate Wnt signal transduction by
crosstalk with other signaling pathways (106). Signal transducer and activator of
transcription 3 (STAT3) has been confirmed to be associated with
EMT via regulation of β-catenin (107). Guo et al reported that
miR-125b-5p targeting STAT3 results in β-catenin phosphorylation
and degradation in HCC cells, indicating the inhibitory effect of
miR-125b-5p on β-catenin-mediated EMT (62). Cyclin-dependent kinase 2 (CDK2), a
member of the Ser/Thr protein kinase family, plays a crucial role
in cancer proliferation and metastasis (108). Zhang et al found
miR-3619-5p directly targets CDK2 and β-catenin to suppress bladder
carcinoma progression, while further studies revealed that
miR-3619-5p inhibits Wnt signaling partly through the induction of
p21 following CDK2 and β-catenin inhibition (94). Cullin 4B (CUL4B), a scaffold protein
assembling the cullin-RING-based E3 ubiquitin-protein ligase
complexes, plays a critical role in proteolysis and tumorigenesis
(109). Zhang et al
reported that CUL4B is a direct target of miR-300 in pancreatic
cancer cells, and downregulation of CUL4B by miR-300 results in
inhibition of the Wnt signaling pathway and EMT (110). Similarly, Cullin 4A (CUL4A), also
known as a core component of multiple cullin-RING-based E3
ubiquitin-protein ligase complexes, is negatively regulated by
miR-377, indicating the inhibitory effect of miR-377 on the Wnt
signaling pathway (111). A member
of the receptor tyrosine kinases (RTKs) family, EPH receptor A2
(EphA2) is highly expressed in solid tumors, suggesting its
important role in tumor initiation, progression, and invasion
(112). MiR-302b and miR-338 serve
as EphA2 inhibitors to suppress gastric cancer tumorigenesis and
metastasis by inactivating the Wnt/β-catenin pathway (113,114). LIM Homeobox 2 (LHX2), a member of
the LIM homeobox family, is involved in elevated β-catenin level
and cell proliferation in pancreatic ductal adenocarcinoma
(115). Liang et al
revealed that miR-506 targets LHX2 to repress EMT and lymph node
metastasis in nasopharyngeal carcinoma. They also found decreased
TCF4 following LHX2 inhibition is responsible for Wnt/β-catenin
signaling inactivation (116).
Moreover, protein regulator of cytokinesis 1 (PRC1) was reported to
mediate early HCC formation, transfer, stemness and development
through Wnt/β-catenin signaling (117) and miR-194 could target PRC1 to
suppress EMT in HCC cells by inactivating the Wnt/β-catenin
signaling pathway (118).
Additionally, Chen et al found that YWHAZ (14-3-3ζ)
regulates the EMT process by interaction with β-catenin in NSCLC
(119). On this basis, Guo et
al demonstrated miR-375-3p targets YWHAZ to inhibit migration,
invasion, and the EMT processes of gastric cancer cells by blocking
the Wnt/β-catenin signaling pathway (120).
Although miRNAs may regulate Wnt signaling by
affecting other signaling pathways, the underlying mechanism on how
they interact has not been clearly defined. For example, miR-22
targeting formin-like 2 (FMNL2) (121), miR-136 targeting premelanosome
protein (PMEL) (122), miR-29c
targeting protein tyrosine phosphatase 4A2 (PTP4A2) and G protein
subunit alpha 13 (GNA13) (123),
and miR-378 targeting SDAD1 (60)
all participate in Wnt/β-catenin signaling inhibition; however, the
relationship between these targets and Wnt signaling needs further
exploration. Therefore, the study of miRNAs targeting Wnt/β-catenin
signaling, not only reveals the complex process of EMT, but also
gives us better understanding of the crosstalk between Wnt
signaling and other signaling pathways. For instance, Zhang et
al found that miR-770 functions as a tumor suppressor by
directly targeting the Jumonji domain containing 6 (JMJD6) 3′-UTR
and inhibiting the Wnt/β-catenin pathway in NSCLC, suggesting
Wnt/β-catenin as the downstream signal of JMJD6 in NSCLC cells
(124).
MiRNAs upregulated in various types of cancer
display their carcinogenic role in tumor progression, migration,
and invasion (125,126). There is a smaller quantity of
miRNAs as EMT promoters compared with EMT inhibitors by targeting
the Wnt signaling pathway (Table
II, Fig. 1), but exploration of
these miRNAs as potential therapeutic targets is also meaningful in
combating EMT (127).
E-cadherin is an important intercellular adhesion
molecule in maintaining cell-cell junctions and polarity. It is
known that suppression of E-cadherin may result in cell detachment,
invasion, and metastasis (128).
Therefore, miRNAs which target E-cadherin are involved in EMT
initiation. According to research, miR-9 (129), miR-23a (130), miR-544a (125) and miR-199a-5p (131) suppress E-cadherin to trigger EMT
in particular cancer types, indicating these miRNAs are potential
targets in cancer therapy.
Axin, APC, and GSK3β are β-catenin suppressors that
act by forming a destructive complex to anchor β-catenin thus
making it degrade. Therefore, miRNAs which target Axin, APC, and
GSK3β activate Wnt signaling to trigger EMT by stabilization of
β-catenin in the nucleus (132). A
study by Mao et al demonstrated that miR-135a activates the
Wnt/β-catenin signaling pathway by directly targeting GSK3β to
accelerate the EMT, invasion, and migration of bladder cancer cells
(133). In addition, miR-1246
facilitates the Wnt/β-catenin pathway through targeting GSK3β,
which partly contributes to lung cancer metastasis (134). However, there is another study
demonstrating different effects and mechanisms of miRNA on GSK3β.
GSK3β modulates the NF-κB signaling pathway as it facilitates NF-κB
function through post-transcriptional regulation of the NF-κB
complex (135). Liu et al
found that GSK3β is a direct target of miR-377-3p and is
upregulated by miR-377-3p. Consequently, miR-377-3p promotes cell
proliferation and EMT by upregulating GSK-3β expression and
activating the NF-κB pathway in human colorectal cancer (136). In addition, miR-197 was reported
to directly target Axin2 in HCC, leading to activation of
Wnt/β-catenin signaling and EMT (126). Similarly, miR-544a plays an
oncogenic role by directly targeting Axin2 to trigger EMT of
gastric cancer (125). Moreover,
APC was identified as the direct and functional target of miR-27
(132) and miR-125b (137) in gastric cancer and breast cancer,
respectively, making miR-27 and miR-125b promising therapeutic
targets for invasive cancer treatment.
The Wnt/β-catenin signaling pathway could be
negatively regulated by antagonist molecules, therefore miRNAs
targeting antagonists of the Wnt signaling have been regarded as
EMT drivers. The DKK gene family, composed of DKK1-4 (138), was found to inhibit tumor invasion
and migration by negative regulation of β-catenin (139). Some studies focused on the DKK
family and found that miR-95-3p targeting DKK3 in prostatic cancer
(140), miR-197 targeting DKK2 in
HCC (126), and miR-373-3p
targeting DKK1 in tongue squamous cell carcinoma (141) are responsible for the activation
of Wnt/β-catenin signaling and EMT. Secreted frizzled-related
protein 1 (SFRP1) acts as an antagonist of Wnt signaling by binding
to Wnt proteins through its CRD domain against the transmembrane
frizzled receptor (142).
MiR-27a-3p was confirmed to promote EMT in oral squamous carcinoma
stem cells by targeting SFRP1 (143). Zinc and ring finger 3 (ZNRF3)
belongs to the E3 ubiquitin ligase family, which negatively
regulates Wnt/β-catenin signaling by promoting the turnover of FZD
and LRP6 (144). Qiao et al
found that miR-106b-3p promotes cell proliferation and invasion by
directly targeting ZNRF3, thus triggering EMT of esophageal
squamous cell carcinoma (ESCC) (145). In addition, miR-146b-5p induces
EMT in thyroid cancer by silencing of ZNRF3 (146). KLF4 (Kruppel-like factor 4),
highly expressed in the adult intestine, is another negative
regulator of Wnt signaling by interacting with β-catenin (147). Chen et al showed that
miR-92a acts as an oncogene by directly targeting KLF4, thus
affecting Wnt/β-catenin pathway and participating in colorectal
cancer progression (148). A study
by Parenti et al also demonstrated that Mesalazine treatment
suppresses the expression of miR-130a and miR-135b, which target
KLF4 mRNA, to mediate β-catenin inhibition in colon cancer
(149). Furthermore, it was
identified that miR-374a activates Wnt/β-catenin signaling to
promote breast cancer metastasis by targeting multiple negative
regulators of Wnt including WIF1, PTEN, and Wnt5A (15). Similarly, downregulation of PTEN and
Wnt5A by miR-26b also results in colorectal cancer metastasis
(150). However, the effect of the
miR-29 family on WIF1 in NSCLC is completely opposite, as miR-29
positively regulates WIF1 expression by inhibiting the methylation
of its promoter, thus inhibiting the Wnt signaling pathway
(151).
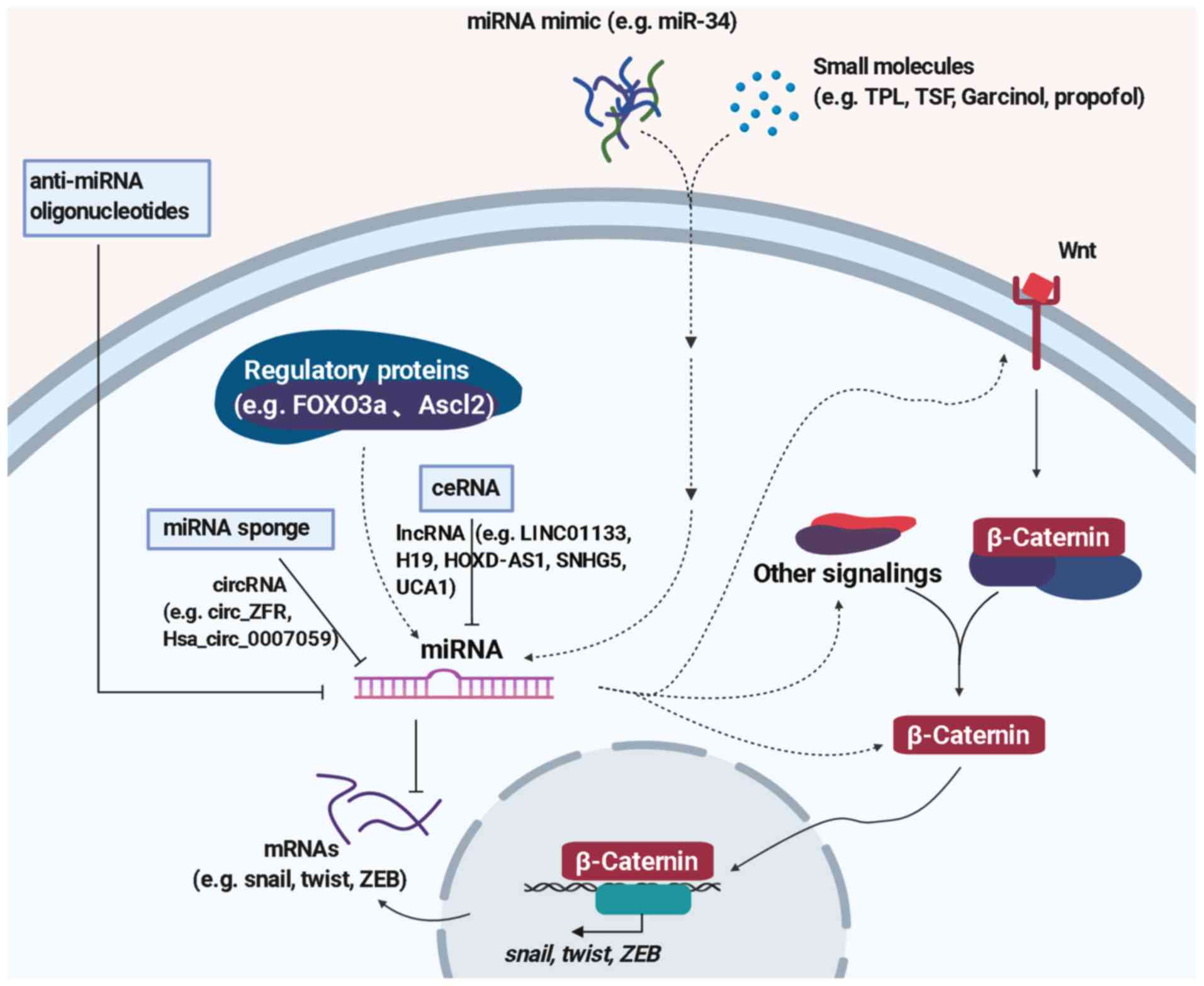
Since miRNAs play important roles in the regulation
of EMT by activating or inhibiting the Wnt signaling pathway,
miRNA-based therapies including those inhibiting miRNA function or
restoring miRNA expression have been suggested as efficient
strategies in cancer treatment (152) (Fig.
2). Delivering miRNA mimics contributes to the restoration of
tumor-suppressive miRNA, while miRNA sponges, anti-miRNA
oligonucleotides, small molecule inhibitors are useful approaches
to block tumor promotive miRNA (153).
The first miRNA-based therapy for cancer is MRX34,
which was designed to deliver miR-34 mimic to cancer cells. MiR-34,
which exerts a suppressive effect on Wnt signaling and tumor
metastasis, is downregulated in various types of cancer including
colon cancer, liver cancer, NSCLC, and cervical cancer (154). Several preclinical studies
demonstrated that delivery of miR-34 mimic has promising effects
against liver cancer (155), lung
cancer (156), and prostate cancer
(157). MRX34 encapsulated in
lipid is under clinical testing (NCT01829971) in several solid and
haematological malignancies (158). In addition, miravirsen, a locked
nucleic acid (LNA)-based antisense oligonucleotide targeting
miR-122, reached phase II trials for treating hepatitis (127). Recently, LNA-modified miR-92a
inhibitor MRG-110 and miR-29 mimic MRG-201 are under phase I
clinical trials by miRagen Therapeutics, Inc. (159). RGLS5579, which targets miR-10b,
demonstrated statistically significant improvements in survival in
an orthotopic glioblastoma multiforme animal model, and the
addition of a single dose of RGLS5579 combined with temozolomide
led to a >2-fold improvement in survival compared to TMZ alone
(https://www.sec.gov/Archives/edgar/data/1505512/000162828020003483/rgls20191231-10k.htm).
Moreover, replenishing tumor-suppressive miRNAs such as miR-200,
miR-26a, miR-506, miR-520, miR-15/16 and inhibiting
tumor-stimulating miRNAs such as miR-10b, miR-221, miR-155, miR-630
have also been included in preclinical studies (160).
Efforts have been made to explore small molecular
compounds targeting EMT-related miRNAs. A natural compound isolated
from Tripterygium wifordii Hook F, namely, Triptolide (TPL),
was reported to exert anti-colorectal cancer properties by
downregulating miR-191, thus blocking NF-κB and Wnt/β-catenin
signaling activation (161).
Toosendanin (TSN), a triterpenoid extracted from the bark or fruits
of Melia toosendan Sieb et Zucc, suppresses gastric cancer
proliferation, invasion, and migration by targeting miR-200a to
downregulate β-catenin (162). In
addition, another study demonstrated that Garcinol exerts
antineoplastic effects on aggressive breast cancer due to reversal
of the mesenchymal phenotype, which is mediated by miR-200s and
let-7s targeting NF-κB and Wnt signaling (163). Moreover, Du et al showed
that propofol can inhibit the proliferation and EMT of MCF-7 cells
by targeting miR-21 to regulate the PI3K/Akt and Wnt/β-catenin
pathway (164). These findings not
only provide promising compounds against EMT but also reveal the
mechanism of miRNAs as targets in Wnt signaling regulation.
There are numerous studies demonstrating the
upstream regulators of miRNA including key proteins, circular RNA
(circRNA), and long non-coding RNA (lncRNA) (Fig. 2). For example, Forkhead box-O 3a
(FOXO3a) inhibits β-catenin through transactivating miR-34b/c
(93). As mentioned above, Ascl2
negatively regulates the miR-200 family which belongs to tumor
suppressors, making Ascl2 a potential target to reverse EMT
(75). Hsa_circ_0007059 blocks the
Wnt/β-catenin and ERK1/2 pathways by targeting miR-378 in A549 and
H1975 cells (172). MiR-106a-3p is
a direct target of lncRNA LINC01133 which suppresses gastric cancer
metastasis by acting as a competitive endogenous RNA (ceRNA) for
miR-106a-3p to regulate APC in Wnt/β-catenin signaling (173). Furthermore, LncRNA H19 (174), lncRNA HOXD-AS1 (175), lncRNA SNHG5 (176) and lncRNA UCA1 (177) were confirmed to regulate the Wnt
signaling pathway by targeting miRNAs. All the evidence indicate
that multiple miRNA-mediated signal transductions participate in
the regulation of EMT. Revealing the connections of miRNAs and
their upstream regulators may give us new prospective therapeutic
strategies for cancer treatment.
Since miRNA has established its role in EMT, the
strategy of utilizing miRNA to overcome cancer metastasis has
increasingly gained attention. Although the complex mechanism of
EMT regulation by miRNA has not been fully defined, miRNAs are
still regarded as potential therapeutic implements in cancer
(Fig. 2). On the one hand, various
methods of directly switching the level of miRNA by miRNA mimics,
miRNA sponges, or anti-miRNA oligonucleotides, which are under
study for different phases, have been shown to be effective. On the
other hand, indirect regulation of miRNAs by affecting upstream
regulators (protein, circRNA, lncRNA) or crosstalk with other
signaling pathways are also useful approaches to inhibit EMT.
Currently, a number of miRNA-based therapies are in clinical trials
to treat cancer or other diseases. However, safety concerns
regarding miRNA therapy always exist. Off-target side-effects,
toxicity, and carcinogenicity of miRNA are important challenges in
the development of miRNA therapy. Seeking effective delivery
systems for miRNA is also a dilemma, so further research may focus
on these issues to improve the utilization value of miRNA
therapy.
In conclusion, miRNAs regulate Wnt/β-catenin
signaling through targeting transcription factors and key proteins
of Wnt signaling or crosstalk with other signaling pathways.
However, the complicated role of miRNA as either a tumor suppressor
or an oncogene and its underlying mechanism need further
exploration. This review, not only provides potential applications
of miRNAs as molecular targets in invasive tumor treatment, but
also helps us gain a better understanding of the complexity of the
EMT process and crosstalk between Wnt/β-catenin and other signaling
pathways.
The authors would like to thank Professor Dong-Mei
Zhang (College of Pharmacy, Jinan University) and Jun-Shan Liu
(Traditional Chinese Medicine, Southern Medical University) for
their guidance.
This review was supported by the National Natural
Science Foundation of China (grant nos. 81803790, 81573918, and
81703975), National Natural Science Foundation of Guangdong (grant
no. 2020A1515011090), Project of Administration of Traditional
Chinese Medicine of Guangdong Province of China (grant no.
20181069), the Fundamental Research Funds for the Central
Universities (grant no. 21618336), and Public Health Research
Projects of Futian District, Shenzhen (grant no. FTWS2019064)
Not applicable.
EXZ and LJD designed the study and revised the
manuscript. YHL, LC and GZ searched the literature and drafted the
manuscript. AYS, BL, JYS and CFZ were also involved in the
conception of the study. JW, XL, CFY and YYC assisted with the
critical revision of the manuscript. All authors have read and
approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Pasquier J, Abu-Kaoud N, Al Thani H and
Rafii A: Epithelial to mesenchymal transition in a clinical
perspective. J Oncol. 2015:7921822015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Okumura N, Kagami T, Fujii K, Nakahara M
and Koizumi N: Involvement of nectin-afadin in the adherens
junctions of the corneal endothelium. Cornea. 37:633–640. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsukita S, Furuse M and Itoh M:
Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol.
2:285–293. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takai Y, Miyoshi J, Ikeda W and Ogita H:
Nectins and nectin-like molecules: Roles in contact inhibition of
cell movement and proliferation. Nat Rev Mol Cell Biol. 9:603–615.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Inagaki M, Irie K, Ishizaki H,
Tanaka-Okamoto M, Miyoshi J and Takai Y: Role of cell adhesion
molecule nectin-3 in spermatid development. Genes Cells.
11:1125–1132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okabe N, Shimizu K, Ozaki-Kuroda K,
Nakanishi H, Morimoto K, Takeuchi M, Katsumaru H, Murakami F and
Takai Y: Contacts between the commissural axons and the floor plate
cells are mediated by nectins. Dev Biol. 273:244–256. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heuberger J and Birchmeier W: Interplay of
cadherin-mediated cell adhesion and canonical Wnt signaling. Cold
Spring Harb Perspect Biol. 2:a0029152010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Coopman P and Djiane A: Adherens Junction
and E-Cadherin complex regulation by epithelial polarity. Cell Mol
Life Sci. 73:3535–3553. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo F, Parker Kerrigan BC, Yang D, Hu L,
Shmulevich I, Sood AK, Xue F and Zhang W: Post-transcriptional
regulatory network of epithelial-to-mesenchymal and
mesenchymal-to-epithelial transitions. J Hematol Oncol. 7:192014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong C, Wu Y, Wang Y, Wang C, Kang T,
Rychahou PG, Chi YI, Evers BM and Zhou BP: Interaction with Suv39H1
is critical for Snail-mediated E-cadherin repression in breast
cancer. Oncogene. 32:1351–1362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo Q and Qin W: DKK3 blocked
translocation of β-catenin/EMT induced by hypoxia and improved
gemcitabine therapeutic effect in pancreatic cancer Bxpc-3 cell. J
Cell Mol Med. 19:2832–2841. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao JH, Luo Y, Jiang YG, He DL and Wu CT:
Knockdown of β-Catenin through shRNA cause a reversal of EMT and
metastatic phenotypes induced by HIF-1α. Cancer Invest. 29:377–382.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
16
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu C, Zhuang Y, Jiang S, Liu S, Zhou J, Wu
J, Teng Y, Xia B, Wang R and Zou X: Interaction between
Wnt/beta-catenin pathway and microRNAs regulates
epithelial-mesenchymal transition in gastric cancer (Review). Int J
Oncol. 48:2236–2246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zaravinos A: The Regulatory Role of
MicroRNAs in EMT and Cancer. J Oncol. 2015:8658162015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim DH, Xing T, Yang Z, Dudek R, Lu Q and
Chen YH: Epithelial Mesenchymal Transition in Embryonic
Development, Tissue Repair and Cancer: A Comprehensive Overview. J
Clin Med. 7:12017. View Article : Google Scholar
|
|
20
|
Zhang RR, Gui YH and Wang X: Role of the
canonical Wnt signaling pathway in heart valve development.
Zhongguo Dang Dai Er Ke Za Zhi. 17:757–762. 2015.(In Chinese).
PubMed/NCBI
|
|
21
|
Ahsan K, Singh N, Rocha M, Huang C and
Prince VE: Prickle1 is required for EMT and migration of zebrafish
cranial neural crest. Dev Biol. 448:16–35. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Paolillo M, Serra M and Schinelli S:
Integrins in glioblastoma: Still an attractive target? Pharmacol
Res. 113:55–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Santoro R, Zanotto M, Carbone C, Piro G,
Tortora G and Melisi D: MEKK3 sustains EMT and stemness in
pancreatic cancer by regulating YAP and TAZ transcriptional
activity. Anticancer Res. 38:1937–1946. 2018.PubMed/NCBI
|
|
25
|
Hu B, Tian X and Li Y, Yang T, Han Z, An
J, Kong L and Li Y: Epithelial-mesenchymal transition may be
involved in the immune evasion of circulating gastric tumor cells
via downregulation of ULBP1. Cancer Med. 9:2686–2697. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kang H, Kim H, Lee S, Youn H and Youn B:
Role of Metabolic Reprogramming in Epithelial-Mesenchymal
Transition (EMT). Int J Mol Sci. 20:20422019. View Article : Google Scholar
|
|
27
|
Garg M: Epithelial plasticity, autophagy
and metastasis: Potential modifiers of the crosstalk to overcome
therapeutic resistance. Stem Cell Rev Rep. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen L, Mai W, Chen M, Hu J, Zhuo Z, Lei
X, Deng L, Liu J, Yao N, Huang M, et al: Arenobufagin inhibits
prostate cancer epithelial-mesenchymal transition and metastasis by
down-regulating β-catenin. Pharmacol Res. 123:130–142. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Derynck R, Muthusamy BP and Saeteurn KY:
Signaling pathway cooperation in TGF-β-induced
epithelial-mesenchymal transition. Curr Opin Cell Biol. 31:56–66.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Creighton CJ, Li X, Landis M, Dixon JM,
Neumeister VM, Sjolund A, Rimm DL, Wong H, Rodriguez A,
Herschkowitz JI, et al: Residual breast cancers after conventional
therapy display mesenchymal as well as tumor-initiating features.
Proc Natl Acad Sci USA. 106:13820–13825. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: An alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Astudillo P: Wnt5a Signaling in Gastric
Cancer. Front Cell Dev Biol. 8:1102020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang M, Marco P, Capra V and Kibar Z:
Update on the Role of the Non-Canonical Wnt/Planar Cell Polarity
Pathway in Neural Tube Defects. Cells. 8:11982019. View Article : Google Scholar
|
|
34
|
Cici D, Corrado A, Rotondo C and Cantatore
FP: Wnt signaling and biological therapy in rheumatoid arthritis
and spondyloarthritis. Int J Mol Sci. 20:55522019. View Article : Google Scholar
|
|
35
|
Huang GR, Wei SJ, Huang YQ, Xing W, Wang
LY and Liang LL: Mechanism of combined use of vitamin D and
puerarin in anti-hepatic fibrosis by regulating the Wnt/β-catenin
signalling pathway. World J Gastroenterol. 24:4178–4185. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gay A and Towler DA: Wnt signaling in
cardiovascular disease: Opportunities and challenges. Curr Opin
Lipidol. 28:387–396. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Villarroel A, Del Valle-Perez B, Fuertes
G, Curto J, Ontiveros N, Garcia de Herreros A and Duñach M: Src and
Fyn define a new signaling cascade activated by canonical and
non-canonical Wnt ligands and required for gene transcription and
cell invasion. Cell Mol Life Sci. 77:919–935. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rao TP and Kuhl M: An updated overview on
Wnt signaling pathways: A prelude for more. Circ Res.
106:1798–1806. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Clevers H and Nusse R: Wnt/beta-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kawano Y and Kypta R: Secreted antagonists
of the Wnt signalling pathway. J Cell Sci. 116:2627–2634. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bafico A, Liu G, Yaniv A, Gazit A and
Aaronson SA: Novel mechanism of Wnt signalling inhibition mediated
by Dickkopf-1 interaction with LRP6/Arrow. Nat Cell Biol.
3:683–686. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brott BK and Sokol SY: Regulation of
Wnt/LRP signaling by distinct domains of Dickkopf proteins. Mol
Cell Biol. 22:6100–6110. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu W, Glinka A, Delius H and Niehrs C:
Mutual antagonism between dickkopf1 and dickkopf2 regulates
Wnt/beta-catenin signalling. Curr Biol. 10:1611–1614. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mao B and Niehrs C: Kremen2 modulates
Dickkopf2 activity during Wnt/LRP6 signaling. Gene. 302:179–183.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cruciat CM and Niehrs C: Secreted and
transmembrane wnt inhibitors and activators. Cold Spring Harb
Perspect Biol. 5:a0150812013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Piccolo S, Agius E, Leyns L, Bhattacharyya
S, Grunz H, Bouwmeester T and De Robertis EM: The head inducer
Cerberus is a multifunctional antagonist of Nodal, BMP and Wnt
signals. Nature. 397:707–710. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
48
|
Itasaki N, Jones CM, Mercurio S, Rowe A,
Domingos PM, Smith JC and Krumlauf R: Wise, a context-dependent
activator and inhibitor of Wnt signalling. Development.
130:4295–4305. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, beta-catenin, and ZEB1 in malignant progression of
cancer. Cancer Metastasis Rev. 28:151–166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Talbot LJ, Bhattacharya SD and Kuo PC:
Epithelial-mesenchymal transition, the tumor microenvironment, and
metastatic behavior of epithelial malignancies. Int J Biochem Mol
Biol. 3:117–136. 2012.PubMed/NCBI
|
|
51
|
Ghahhari NM and Babashah S: Interplay
between microRNAs and WNT/beta-catenin signalling pathway regulates
epithelial-mesenchymal transition in cancer. Eur J Cancer.
51:1638–1649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guo Y, Xiao L, Sun L and Liu F:
Wnt/beta-catenin signaling: A promising new target for fibrosis
diseases. Physiol Res. 61:337–346. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gebert LFR and MacRae IJ: Regulation of
microRNA function in animals. Nat Rev Mol Cell Biol. 20:21–37.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek
SH and Kim VN: MicroRNA genes are transcribed by RNA polymerase II.
EMBO J. 23:4051–4060. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Okada C, Yamashita E, Lee SJ, Shibata S,
Katahira J, Nakagawa A, Yoneda Y and Tsukihara T: A high-resolution
structure of the pre-microRNA nuclear export machinery. Science.
326:1275–1279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dalmay T: Mechanism of miRNA-mediated
repression of mRNA translation. Essays Biochem. 54:29–38. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Thomson JM, Newman M, Parker JS,
Morin-Kensicki EM, Wright T and Hammond SM: Extensive
post-transcriptional regulation of microRNAs and its implications
for cancer. Genes Dev. 20:2202–2207. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Moyret-Lalle C, Ruiz E and Puisieux A:
Epithelial-mesenchymal transition transcription factors and miRNAs:
‘Plastic surgeons’ of breast cancer. World J Clin Oncol. 5:311–322.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zeng M, Zhu L, Li L and Kang C: miR-378
suppresses the proliferation, migration and invasion of colon
cancer cells by inhibiting SDAD1. Cell Mol Biol Lett. 22:122017.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tian L, Zhao Z, Xie L and Zhu J:
MiR-361-5p inhibits the mobility of gastric cancer cells through
suppressing epithelial-mesenchymal transition via the Wnt/β-catenin
pathway. Gene. 675:102–109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Guo R, Wu Z, Wang J, Li Q, Shen S, Wang W,
Zhou L, Wang W, Cao Z and Guo Y: Development of a
Non-Coding-RNA-based EMT/CSC Inhibitory Nanomedicine for In Vivo
Treatment and Monitoring of HCC. Adv Sci (Weinh). 6:18018852019.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Teague EM, Print CG and Hull ML: The role
of microRNAs in endometriosis and associated reproductive
conditions. Hum Reprod Update. 16:142–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wiklund ED, Bramsen JB, Hulf T, Dyrskjøt
L, Ramanathan R, Hansen TB, Villadsen SB, Gao S, Ostenfeld MS,
Borre M, et al: Coordinated epigenetic repression of the miR-200
family and miR-205 in invasive bladder cancer. Int J Cancer.
128:1327–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tryndyak VP, Beland FA and Pogribny IP:
E-cadherin transcriptional down-regulation by epigenetic and
microRNA-200 family alterations is related to mesenchymal and
drug-resistant phenotypes in human breast cancer cells. Int J
Cancer. 126:2575–2583. 2010.PubMed/NCBI
|
|
66
|
Elson-Schwab I, Lorentzen A and Marshall
CJ: MicroRNA-200 family members differentially regulate
morphological plasticity and mode of melanoma cell invasion. PLoS
One. 5:e131762010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hu X, Macdonald DM, Huettner PC, Feng Z,
El Naqa IM, Schwarz JK, Mutch DG, Grigsby PW, Powell SN and Wang X:
A miR-200 microRNA cluster as prognostic marker in advanced ovarian
cancer. Gynecol Oncol. 114:457–464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Shinozaki A, Sakatani T, Ushiku T, Hino R,
Isogai M, Ishikawa S, Uozaki H, Takada K and Fukayama M:
Downregulation of microRNA-200 in EBV-associated gastric carcinoma.
Cancer Res. 70:4719–4727. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kong D, Li Y, Wang Z, Banerjee S, Ahmad A,
Kim HR and Sarkar FH: miR-200 regulates PDGF-D-mediated
epithelial-mesenchymal transition, adhesion, and invasion of
prostate cancer cells. Stem Cells. 27:1712–1721. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Saydam O, Shen Y, Wurdinger T, Senol O,
Boke E, James MF, TannousB A, Stemmer-Rachamimov AO, Yi M, Stephens
RM, et al: Downregulated microRNA-200a in meningiomas promotes
tumor growth by reducing E-cadherin and activating the
Wnt/beta-catenin signaling pathway. Mol Cell Biol. 29:5923–5940.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sanchez-Tillo E, Lazaro A, Torrent R,
Cuatrecasas M, Vaquero EC, Castells A, Engel P and Postigo A: ZEB1
represses E-cadherin and induces an EMT by recruiting the SWI/SNF
chromatin-remodeling protein BRG1. Oncogene. 29:3490–3500. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cong N, Du P, Zhang A, Shen F, Su J, Pu P,
Wang T, Zjang J, Kang C and Zhang Q: Downregulated microRNA-200a
promotes EMT and tumor growth through the wnt/β-catenin pathway by
targeting the E-cadherin repressors ZEB1/ZEB2 in gastric
adenocarcinoma. Oncol Rep. 29:1579–1587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Su J, Zhang A, Shi Z, Ma F, Pu P, Wang T,
Zhang J, Kang C and Zhang Q: MicroRNA-200a suppresses the
Wnt/β-catenin signaling pathway by interacting with β-catenin. Int
J Oncol. 40:1162–1170. 2012.PubMed/NCBI
|
|
75
|
Tian Y, Pan Q, Shang Y, Zhu R, Ye J, Liu
Y, Zhong X, Li S, He Y, Chen L, et al: MicroRNA-200 (miR-200)
cluster regulation by achaete scute-like 2 (Ascl2): Impact on the
epithelial-mesenchymal transition in colon cancer cells. J Biol
Chem. 289:36101–36115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liu J, Ruan B, You N, Huang Q, Liu W, Dang
Z, Xu W, Zhou T, Ji R, Cao Y, et al: Downregulation of miR-200a
induces EMT phenotypes and CSC-like signatures through targeting
the beta-catenin pathway in hepatic oval cells. PLoS One.
8:e794092013. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Suliman MA, Zhang Z, Na H, Ribeiro AL,
Zhang Y, Niang B, Hamid AS, Zhang H, Xu L and Zuo Y: Niclosamide
inhibits colon cancer progression through downregulation of the
Notch pathway and upregulation of the tumor suppressor miR-200
family. Int J Mol Med. 38:776–784. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Feng B, Wang R, Song HZ and Chen LB:
MicroRNA-200b reverses chemoresistance of docetaxel-resistant human
lung adenocarcinoma cells by targeting E2F3. Cancer. 118:3365–3376.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Qu J, Li M, An J, Zhao B, Zhong W, Gu Q,
Cao L, Yang H and Hu C: MicroRNA-33b inhibits lung adenocarcinoma
cell growth, invasion, and epithelial-mesenchymal transition by
suppressing Wnt/β-catenin/ZEB1 signaling. Int J Oncol.
47:2141–2152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang X, Xu X, Ge G, Zang X, Shao M, Zou
S, Zhang Y, Mao Z, Zhang J, Mao F, et al: miR498 inhibits the
growth and metastasis of liver cancer by targeting ZEB2. Oncol Rep.
41:1638–1648. 2019.PubMed/NCBI
|
|
82
|
Yue H, Tang B, Zhao Y, Niu Y, Yin P, Yang
W, Zhang Z and Yu P: MIR-519d suppresses the gastric cancer
epithelial-mesenchymal transition via Twist1 and inhibits
Wnt/β-catenin signaling pathway. Am J Transl Res. 9:3654–3664.
2017.PubMed/NCBI
|
|
83
|
Jin Y, Wang J, Han J, Luo D and Sun Z:
MiR-122 inhibits epithelial-mesenchymal transition in
hepatocellular carcinoma by targeting Snail1 and Snail2 and
suppressing WNT/β-cadherin signaling pathway. Exp Cell Res.
360:210–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Song XF, Wang QH and Huo R: Effects of
microRNA-708 on epithelial-mesenchymal transition, cell
proliferation and apoptosis in melanoma cells by targeting lef1
through the wnt signaling pathway. Pathol Oncol Res. 25:377–389.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Liang J, Li Y, Daniels G, Sfanos K, De
Marzo A, Wei J, Li X, Chen W, Wang J, Zhong X, et al: LEF1
Targeting EMT in Prostate Cancer Invasion Is Regulated by miR-34a.
Mol Cancer Res. 13:681–688. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Chen J, Luo Q, Yuan Y, Huang X, Cai W, Li
C, Wei T, Zhang L, Yang M, Liu Q, et al: Pygo2 associates with MLL2
histone methyltransferase and GCN5 histone acetyltransferase
complexes to augment Wnt target gene expression and breast cancer
stem-like cell expansion. Mol Cell Biol. 30:5621–5635. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Chi Y, Wang F, Zhang T, Xu H, Zhang Y,
Shan Z, Wu S, Fan Q and Sun Y: miR-516a-3p inhibits breast cancer
cell growth and EMT by blocking the Pygo2/Wnt signalling pathway. J
Cell Mol Med. 23:6295–6307. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zheng HE, Wang G, Song J, Liu Y, Li YM and
Du WP: MicroRNA-495 inhibits the progression of non-small-cell lung
cancer by targeting TCF4 and inactivating Wnt/beta-catenin pathway.
Eur Rev Med Pharmacol Sci. 22:7750–7759. 2018.PubMed/NCBI
|
|
89
|
Xu Z, Yu Z, Tan Q, Wei C, Tang Q, Wang L
and Hong Y: MiR-876-5p regulates gastric cancer cell proliferation,
apoptosis and migration through targeting WNT5A and MITF. Biosci
Rep. 39:BSR201900662019. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zhang Y, Zhang X, Wang X, Gan L, Yu G,
Chen Y, Liu K, Li P, Pan J, Wang J and Qin S: Inhibition of LDH-A
by lentivirus-mediated small interfering RNA suppresses
intestinal-type gastric cancer tumorigenicity through the
downregulation of Oct4. Cancer Lett. 321:45–54. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Iida H, Suzuki M, Goitsuka R and Ueno H:
Hypoxia induces CD133 expression in human lung cancer cells by
up-regulation of OCT3/4 and SOX2. Int J Oncol. 40:71–79.
2012.PubMed/NCBI
|
|
92
|
Ling DJ, Chen ZS, Zhang YD, Liao QD, Feng
JX, Zhang XY and Shi TS: MicroRNA-145 inhibits lung cancer cell
metastasis. Mol Med Rep. 11:3108–3114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Liu H, Yin J, Wang H, Jiang G, Deng M,
Zhang G, Bu X, Cai S, Du J and He Z: FOXO3a modulates
WNT/beta-catenin signaling and suppresses epithelial-to-mesenchymal
transition in prostate cancer cells. Cell Signal. 27:510–518. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang Q, Miao S, Han X, Li C, Zhang M, Cui
K, Xiong T, Chen Z, Wang C and Xu H: MicroRNA-3619-5p suppresses
bladder carcinoma progression by directly targeting β-catenin and
CDK2 and activating p21. Cell Death Dis. 9:9602018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Liang C, Wang Z, Li YY, Yu BH, Zhang F and
Li HY: miR-33a suppresses the nuclear translocation of beta-catenin
to enhance gemcitabine sensitivity in human pancreatic cancer
cells. Tumour Biol. 36:9395–9403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wang N, Wang Q, Shen D, Sun X, Cao X and
Wu D: Downregulation of microRNA-122 promotes proliferation,
migration, and invasion of human hepatocellular carcinoma cells by
activating epithelial-mesenchymal transition. Onco Targets Ther.
9:2035–2047. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yan H, Dong X, Zhong X, Ye J, Zhou Y, Yang
X, Shen J and Zhang J: Inhibitions of epithelial to mesenchymal
transition and cancer stem cells-like properties are involved in
miR-148a-mediated anti-metastasis of hepatocellular carcinoma. Mol
Carcinog. 53:960–969. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Peng L, Liu Z, Xiao J, Tu Y, Wan Z, Xiong
H, Li Y and Xiao W: MicroRNA-148a suppresses epithelial-mesenchymal
transition and invasion of pancreatic cancer cells by targeting
Wnt10b and inhibiting the Wnt/beta-catenin signaling pathway. Oncol
Rep. 38:301–308. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Huang X, Zhu H, Gao Z, Li J, Zhuang J,
Dong Y, Shen B, Li M, Zhou H, Guo H, Huang R and Yan J: Wnt7a
activates canonical Wnt signaling, promotes bladder cancer cell
invasion, and is suppressed by miR-370-3p. J Biol Chem.
293:6693–6706. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Yang Y, Sun Y, Wu Y, Tang D, Ding X, Xu W,
Su B and Gao W: Downregulation of miR-3127-5p promotes
epithelial-mesenchymal transition via FZD4 regulation of
Wnt/β-catenin signaling in non-small-cell lung cancer. Mol
Carcinog. 57:842–853. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Liu Q, Guan Y, Li Z and Wang Y, Liu Y, Cui
R and Wang Y: miR-504 suppresses mesenchymal phenotype of
glioblastoma by directly targeting the FZD7-mediated Wnt-β-catenin
pathway. J Exp Clin Cancer Res. 38:3582019. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Jin R, Liu W, Menezes S, Yue F, Zheng M,
Kovacevic Z and Richardson DR: The metastasis suppressor NDRG1
modulates the phosphorylation and nuclear translocation of
β-catenin through mechanisms involving FRAT1 and PAK4. J Cell Sci.
127:3116–3130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zheng K, Zhou X, Yu J, Li Q, Wang H, Li M,
Shao Z, Zhang F, Luo Y, Shen Z, et al: Epigenetic silencing of
miR-490-3p promotes development of an aggressive colorectal cancer
phenotype through activation of the Wnt/β-catenin signaling
pathway. Cancer Lett. 376:178–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cui Y, Yang Y, Ren L, Yang J, Wang B, Xing
T, Chen H and Chen M: miR-15a-3p Suppresses Prostate Cancer Cell
Proliferation and Invasion by Targeting SLC39A7 Via Downregulating
Wnt/β-Catenin Signaling Pathway. Cancer Biother Radiopharm.
34:472–479. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Nimmanon T, Ziliotto S, Morris S, Flanagan
L and Taylor KM: Phosphorylation of zinc channel ZIP7 drives MAPK,
PI3K and mTOR growth and proliferation signalling. Metallomics.
9:471–481. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhao X, Lu Y, Nie Y and Fan D: MicroRNAs
as critical regulators involved in regulating epithelial-
mesenchymal transition. Curr Cancer Drug Targets. 13:935–944. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Huang H, Wang C, Liu F, Li HZ, Peng G, Gao
X, Dong KQ, Wang HR, Kong DP, Qu M, et al: Reciprocal network
between cancer stem-like cells and macrophages facilitates the
progression and androgen deprivation therapy resistance of prostate
cancer. Clin Cancer Res. 24:4612–4626. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Arai K, Eguchi T, Rahman MM, Sakamoto R,
Masuda N, Nakatsura T, Calderwood SK, Kozaki K and Itoh M: A Novel
high-throughput 3D screening system for EMT inhibitors: A pilot
screening discovered the EMT inhibitory activity of CDK2 Inhibitor
SU9516. PLoS One. 11:e01623942016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wang X and Chen Z: Knockdown of CUL4B
Suppresses the Proliferation and Invasion in Non-Small Cell Lung
Cancer Cells. Oncol Res. 24:271–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhang JQ, Chen S, Gu JN, Zhu Y, Zhan Q,
Cheng DF, Chen H, Deng XX, Shen BY and Peng CH: MicroRNA-300
promotes apoptosis and inhibits proliferation, migration, invasion
and epithelial-mesenchymal transition via the Wnt/β-catenin
signaling pathway by targeting CUL4B in pancreatic cancer cells. J
Cell Biochem. 119:1027–1040. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yu R, Cai L, Chi Y, Ding X and Wu X:
miR377 targets CUL4A and regulates metastatic capability in ovarian
cancer. Int J Mol Med. 41:3147–3156. 2018.PubMed/NCBI
|
|
112
|
Paraiso KH, Das Thakur M, Fang B, Koomen
JM, Fedorenko IV, John JK, Tsao H, Flaherty KT, Sondak VK, Messina
JL, et al: Ligand-independent EPHA2 signaling drives the adoption
of a targeted therapy-mediated metastatic melanoma phenotype.
Cancer Discov. 5:264–273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Huang J, He Y, McLeod HL, Xie Y, Xiao D,
Hu H, Chen P, Shen L, Zeng S, Yin X, et al: miR-302b inhibits
tumorigenesis by targeting EphA2 via Wnt/β-catenin/EMT signaling
cascade in gastric cancer. BMC Cancer. 17:8862017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Song B, Lin HX, Dong LL, Ma JJ and Jiang
ZG: MicroRNA-338 inhibits proliferation, migration, and invasion of
gastric cancer cells by the Wnt/β-catenin signaling pathway. Eur
Rev Med Pharmacol Sci. 22:1290–1296. 2018.PubMed/NCBI
|
|
115
|
Zhou F, Gou S, Xiong J, Wu H, Wang C and
Liu T: Oncogenicity of LHX2 in pancreatic ductal adenocarcinoma.
Mol Biol Rep. 41:8163–8167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Liang TS, Zheng YJ, Wang J, Zhao JY, Yang
DK and Liu ZS: MicroRNA-506 inhibits tumor growth and metastasis in
nasopharyngeal carcinoma through the inactivation of the
Wnt/β-catenin signaling pathway by down-regulating LHX2. J Exp Clin
Cancer Res. 38:972019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Chen J, Rajasekaran M, Xia H, Zhang X,
Kong SN, Sekar K, Seshachalam VP, Deivasigamani A, Goh BK, Ooi LL,
et al: The microtubule-associated protein PRC1 promotes early
recurrence of hepatocellular carcinoma in association with the
Wnt/β-catenin signalling pathway. Gut. 65:1522–1534. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Tang H, Zhao H, Yu ZY, Feng X, Fu BS, Qiu
CH and Zhang JW: MicroRNA-194 inhibits cell invasion and migration
in hepatocellular carcinoma through PRC1-mediated inhibition of
Wnt/β-catenin signaling pathway. Dig Liver Dis. 51:1314–1322. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Chen CH, Chuang SM, Yang MF, Liao JW, Yu
SL and Chen JJ: A novel function of YWHAZ/beta-catenin axis in
promoting epithelial-mesenchymal transition and lung cancer
metastasis. Mol Cancer Res. 10:1319–1331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Guo F, Gao Y, Sui G, Jiao D, Sun L, Fu Q
and Jin C: miR-375-3p/YWHAZ/β-catenin axis regulates migration,
invasion, EMT in gastric cancer cells. Clin Exp Pharmacol Physiol.
46:144–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Shi L, Huo JW, Chen SS, Xue JX, Gao WY, Li
XY, Song YH, Xu HT, Zhu XW and Chen K: MicroRNA-22 targets FMNL2 to
inhibit melanoma progression via the regulation of the
Wnt/β-catenin signaling pathway and epithelial-mesenchymal
transition. Eur Rev Med Pharmacol Sci. 23:5332–5342.
2019.PubMed/NCBI
|
|
122
|
Wang JJ, Li ZF, Li XJ, Han Z, Zhang L and
Liu ZJ: Effects of microRNA-136 on melanoma cell proliferation,
apoptosis, and epithelial-mesenchymal transition by targetting PMEL
through the Wnt signaling pathway. Biosci Rep. 37:BSR201707432017.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zhang JX, Mai SJ, Huang XX, Wang FW, Liao
YJ, Lin MC, Kung HF, Zeng YX and Xie D: MiR-29c mediates
epithelial-to-mesenchymal transition in human colorectal carcinoma
metastasis via PTP4A and GNA13 regulation of β-catenin signaling.
Ann Oncol. 25:2196–2204. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Zhang Z, Yang Y and Zhang X: MiR-770
inhibits tumorigenesis and EMT by targeting JMJD6 and regulating
WNT/β-catenin pathway in non-small cell lung cancer. Life Sci.
188:163–171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki K
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hu Z, Wang P, Lin J, Zheng X, Yang F,
Zhang G, Chen D, Xie J, Gao Z, Peng L and Xie C: MicroRNA-197
Promotes Metastasis of Hepatocellular Carcinoma by Activating
Wnt/β-Catenin Signaling. Cell Physiol Biochem. 51:470–486. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Gebert LF, Rebhan MA, Crivelli SE, Denzler
R, Stoffel M and Hall J: Miravirsen (SPC3649) can inhibit the
biogenesis of miR-122. Nucleic Acids Res. 42:609–621. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Petrova YI, Schecterson L and Gumbiner BM:
Roles for E-cadherin cell surface regulation in cancer. Mol Biol
Cell. 27:3233–3244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Xu XZ, Li XA, Luo Y, Liu JF, Wu HW and
Huang G: MiR-9 promotes synovial sarcoma cell migration and
invasion by directly targeting CDH1. Int J Biochem Cell Biol.
112:61–71. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Ma F, Li W, Liu C, Li W, Yu H, Lei B, Ren
Y, Li Z, Pang D and Qian C: MiR-23a promotes TGF-β1-induced EMT and
tumor metastasis in breast cancer cells by directly targeting CDH1
and activating Wnt/β-catenin signaling. Oncotarget. 8:69538–69550.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhao X, He L, Li T, Lu Y, Miao Y, Liang S,
Guo H, Bai M, Xie H, Luo G, et al: SRF expedites metastasis and
modulates the epithelial to mesenchymal transition by regulating
miR-199a-5p expression in human gastric cancer. Cell Death Differ.
21:1900–1913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Zhang Z, Liu S, Shi R and Zhao G: miR-27
promotes human gastric cancer cell metastasis by inducing
epithelial-to-mesenchymal transition. Cancer Genet. 204:486–491.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Mao XW, Xiao JQ, Li ZY, Zheng YC and Zhang
N: Effects of microRNA-135a on the epithelial-mesenchymal
transition, migration and invasion of bladder cancer cells by
targeting GSK3β through the Wnt/β-catenin signaling pathway. Exp
Mol Med. 50:e4292018. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Yang F, Xiong H, Duan L, Li Q, Li X and
Zhou Y: MiR-1246 Promotes Metastasis and Invasion of A549 cells by
Targeting GSK-3betaMediated Wnt/β-Catenin Pathway. Cancer Res
Treat. 51:1420–1429. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Hoeflich KP, Luo J, Rubie EA, Tsao MS, Jin
O and Woodgett JR: Requirement for glycogen synthase kinase-3beta
in cell survival and NF-kappaB activation. Nature. 406:86–90. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Liu WY, Yang Z, Sun Q, Yang X, Hu Y, Xie
H, Gao HJ, Guo LM, Yi JY, Liu M and Tang H: miR-377-3p drives
malignancy characteristics via upregulating GSK-3β expression and
activating NF-κB pathway in hCRC cells. J Cell Biochem.
119:2124–2134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Nie J, Jiang HC, Zhou YC, Jiang B, He WJ,
Wang YF and Dong J: MiR-125b regulates the proliferation and
metastasis of triple negative breast cancer cells via the
Wnt/β-catenin pathway and EMT. Biosci Biotechnol Biochem.
83:1062–1071. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Barrantes Idel B, Montero-Pedrazuela A,
Guadano-Ferraz A, Obregon MJ, Martinez de Mena R, Gailus-Durner V,
Fuchs H, Franz TJ, Kalaydjiev S, Klempt M, et al: Generation and
characterization of dickkopf3 mutant mice. Mol Cell Biol.
26:2317–2326. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Hoang BH, Kubo T, Healey JH, Yang R,
Nathan SS, Kolb EA, Mazza B, Meyers PA and Gorlick R: Dickkopf 3
inhibits invasion and motility of Saos-2 osteosarcoma cells by
modulating the Wnt-beta-catenin pathway. Cancer Res. 64:2734–2739.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Xi M, Cheng L, Hua W, Zhou YL, Gao QL,
Yang JX and Qi SY: MicroRNA-95-3p promoted the development of
prostatic cancer via regulating DKK3 and activating Wnt/β-catenin
pathway. Eur Rev Med Pharmacol Sci. 23:1002–1011. 2019.PubMed/NCBI
|
|
141
|
Weng J, Zhang H, Wang C, Liang J, Chen G,
Li W, Tang H and Hou J: miR-373-3p Targets DKK1 to promote
EMT-induced metastasis via the Wnt/β-catenin pathway in tongue
squamous cell carcinoma. Biomed Res Int. 2017:60109262017.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Elzi DJ, Song M, Hakala K, Weintraub ST
and Shiio Y: Wnt antagonist SFRP1 functions as a secreted mediator
of senescence. Mol Cell Biol. 32:4388–4399. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Qiao B, He BX, Cai JH, Tao Q and King-Yin
Lam A: MicroRNA-27a-3p modulates the Wnt/β-catenin signaling
pathway to promote epithelial-mesenchymal transition in oral
squamous carcinoma stem cells by targeting SFRP1. Sci Rep.
7:446882017. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Zebisch M, Xu Y, Krastev C, MacDonald BT,
Chen M, Gilbert RJ, He X and Jones EY: Structural and molecular
basis of ZNRF3/RNF43 transmembrane ubiquitin ligase inhibition by
the Wnt agonist R-spondin. Nat Commun. 4:27872013. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Qiao G, Dai C, He Y, Shi J and Xu C:
Effects of miR106b3p on cell proliferation and
epithelialmesenchymal transition, and targeting of ZNRF3 in
esophageal squamous cell carcinoma. Int J Mol Med. 43:1817–1829.
2019.PubMed/NCBI
|
|
146
|
Deng X, Wu B, Xiao K, Kang J, Xie J, Zhang
X and Fan Y: MiR-146b-5p promotes metastasis and induces
epithelial-mesenchymal transition in thyroid cancer by targeting
ZNRF3. Cell Physiol Biochem. 35:71–82. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Zhang W, Chen X, Kato Y, Evans PM, Yuan S,
Yang J, Rychahou PG, Yang VW, He X, Evers BM and Liu C: Novel cross
talk of Kruppel-like factor 4 and beta-catenin regulates normal
intestinal homeostasis and tumor repression. Mol Cell Biol.
26:2055–2064. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Chen E, Li Q, Wang H, Yang F, Min L and
Yang J: MiR-92a promotes tumorigenesis of colorectal cancer, a
transcriptomic and functional based study. Biomed Pharmacother.
106:1370–1377. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Parenti S, Montorsi L, Fantini S, Mammoli
F, Gemelli C, Atene CG, Losi L, Frassineti C, Calabretta B,
Tagliafico E, et al: KLF4 Mediates the Effect of 5-ASA on the
β-Catenin Pathway in Colon Cancer Cells. Cancer Prev Res (Phila).
11:503–510. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Fan D, Lin X, Zhang F, Zhong W, Hu J, Chen
Y, Cai Z, Zou Y, He X, Chen X, et al: MicroRNA 26b promotes
colorectal cancer metastasis by downregulating phosphatase and
tensin homolog and wingless-type MMTV integration site family
member 5A. Cancer Sci. 109:354–362. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Tan M, Wu J and Cai Y: Suppression of Wnt
signaling by the miR-29 family is mediated by demethylation of
WIF-1 in non-small-cell lung cancer. Biochem Biophys Res Commun.
438:673–679. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Tokarz P and Blasiak J: The role of
microRNA in metastatic colorectal cancer and its significance in
cancer prognosis and treatment. Acta Biochim Pol. 59:467–474. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Bader AG: miR-34-a microRNA replacement
therapy is headed to the clinic. Front Genet. 3:1202012. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Slaby O, Laga R and Sedlacek O:
Therapeutic targeting of non-coding RNAs in cancer. Biochem J.
474:4219–4251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Trang P, Wiggins JF, Daige CL, Cho C,
Omotola M, Brown D, Weidhaas JB, Bader AG and Slack FJ: Systemic
delivery of tumor suppressor microRNA mimics using a neutral lipid
emulsion inhibits lung tumors in mice. Mol Ther. 19:1116–1122.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, et
al: The microRNA miR-34a inhibits prostate cancer stem cells and
metastasis by directly repressing CD44. Nat Med. 17:211–215. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Zhang L, Liao Y and Tang L: MicroRNA-34
family: A potential tumor suppressor and therapeutic candidate in
cancer. J Exp Clin Cancer Res. 38:532019. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Gallant-Behm CL, Piper J, Dickinson BA,
Dalby CM, Pestano LA and Jackson AL: A synthetic microRNA-92a
inhibitor (MRG-110) accelerates angiogenesis and wound healing in
diabetic and nondiabetic wounds. Wound Repair Regen. 26:311–323.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Qi Y and Li J: Triptolide inhibits the
growth and migration of colon carcinoma cells by down-regulation of
miR-191. Exp Mol Pathol. 107:23–31. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Wang G, Huang YX, Zhang R, Hou LD, Liu H,
Chen XY, Zhu JS and Zhang J: Toosendanin suppresses oncogenic
phenotypes of human gastric carcinoma SGC7901 cells partly via
miR200amediated downregulation of β-catenin pathway. Int J Oncol.
51:1563–1573. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Ahmad A, Sarkar SH, Bitar B, Ali S,
Aboukameel A, Sethi S, Li Y, Bao B, Kong D, Banerjee S, et al:
Garcinol regulates EMT and Wnt signaling pathways in vitro and in
vivo, leading to anticancer activity against breast cancer cells.
Mol Cancer Ther. 11:2193–2201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Du Q, Zhang X, Zhang X, Wei M, Xu H and
Wang S: Propofol inhibits proliferation and epithelial-mesenchymal
transition of MCF-7 cells by suppressing miR-21 expression. Artif
Cells Nanomed Biotechnol. 47:1265–1271. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Zhao M, Xu P, Liu Z, Zhen Y, Chen Y, Liu
Y, Fu Q, Deng X, Liang Z, Li Y, et al: Dual roles of miR-374a by
modulated c-Jun respectively targets CCND1-inducing PI3K/AKT signal
and PTEN-suppressing Wnt/β-catenin signaling in non-small-cell lung
cancer. Cell Death Dis. 9:782018. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Sathyanarayanan A, Chandrasekaran KS and
Karunagaran D: microRNA-145 downregulates SIP1-expression but
differentially regulates proliferation, migration, invasion and Wnt
signaling in SW480 and SW620 cells. J Cell Biochem. 119:2022–2035.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Li D, Tian B and Jin X: miR-630 Inhibits
Epithelial-to-Mesenchymal Transition (EMT) by Regulating the
Wnt/β-Catenin Pathway in Gastric Cancer Cells. Oncol Res. 27:9–17.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Chien AJ, Moore EC, Lonsdorf AS,
Kulikauskas RM, Rothberg BG, Berger AJ, Major MB, Hwang ST, Rimm DL
and Moon RT: Activated Wnt/beta-catenin signaling in melanoma is
associated with decreased proliferation in patient tumors and a
murine melanoma model. Proc Natl Acad Sci USA. 106:1193–1198. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Ha TY: MicroRNAs in human diseases: From
cancer to cardiovascular disease. Immune Netw. 11:135–154. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Wang HY, Liu YN, Wu SG, Hsu CL, Chang TH,
Tsai MF, Lin YT and Shih JY: MiR-200c-3p suppression is associated
with development of acquired resistance to epidermal growth factor
receptor (EGFR) tyrosine kinase inhibitors in EGFR mutant non-small
cell lung cancer via a mediating epithelial-to-mesenchymal
transition (EMT) process. Cancer Biomark. 28:351–363. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Cochrane DR, Spoelstra NS, Howe EN,
Nordeen SK and Richer JK: MicroRNA-200c mitigates invasiveness and
restores sensitivity to microtubule-targeting chemotherapeutic
agents. Mol Cancer Ther. 8:1055–1066. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Gao S, Yu Y, Liu L, Meng J and Li G:
Circular RNA hsa_circ_0007059 restrains proliferation and
epithelial-mesenchymal transition in lung cancer cells via
inhibiting microRNA-378. Life Sci. 233:1166922019. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Yang XZ, Cheng TT, He QJ, Lei ZY, Chi J,
Tang Z, Liao QX, Zhang H, Zeng LS and Cui SZ: LINC01133 as ceRNA
inhibits gastric cancer progression by sponging miR-106a-3p to
regulate APC expression and the Wnt/beta-catenin pathway. Mol
Cancer. 17:1262018. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Ding D, Li C, Zhao T, Li D, Yang L and
Zhang B: LncRNA H19/miR-29b-3p/PGRN axis promoted
epithelial-mesenchymal transition of colorectal cancer cells by
acting on wnt signaling. Mol Cells. 41:423–435. 2018.PubMed/NCBI
|
|
175
|
Zhang Y, Dun Y, Zhou S and Huang XH:
LncRNA HOXD-AS1 promotes epithelial ovarian cancer cells
proliferation and invasion by targeting miR-133a-3p and activating
Wnt/β-catenin signaling pathway. Biomed Pharmacother. 96:1216–1221.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Li Y, Guo D, Zhao Y, Ren M, Lu G, Wang Y,
Zhang J, Mi C, He S and Lu X: Long non-coding RNA SNHG5 promotes
human hepatocellular carcinoma progression by regulating
miR-26a-5p/GSK3β signal pathway. Cell Death Dis. 9:8882018.
View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Chen X, Gao J, Yu Y, Zhao Z and Pan Y:
Long non-coding RNA UCA1 targets miR-185-5p and regulates cell
mobility by affecting epithelial-mesenchymal transition in melanoma
via Wnt/β-catenin signaling pathway. Gene. 676:298–305. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Taube JH, Malouf GG, Lu E, Sphyris N,
Vijay V, Ramachandran PP, Ueno KR, Gaur S, Nicoloso MS, Rossi S, et
al: Epigenetic silencing of microRNA-203 is required for EMT and
cancer stem cell properties. Sci Rep. 3:26872013. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Strillacci A, Valerii MC, Sansone P,
Caggiano C, Sgromo A, Vittori L, Fiorentino M, Poggioli G, Rizzello
F, Campieri M and Spisni E: Loss of miR-101 expression promotes
Wnt/β-catenin signalling pathway activation and malignancy in colon
cancer cells. J Pathol. 229:379–389. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Huang Z, Li Q, Luo K, Zhang Q, Geng J,
Zhou X, Xu Y, Qian M, Zhang JA, Ji L and Wu J: miR-340-FHL2 axis
inhibits cell growth and metastasis in ovarian cancer. Cell Death
Dis. 10:3722019. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Liu P, Chen B, Gu Y and Liu Q: PNMA1,
regulated by miR-33a-5p, promotes proliferation and EMT in
hepatocellular carcinoma by activating the Wnt/β-catenin pathway.
Biomed Pharmacother. 108:492–499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Han S, Cao C, Tang T, Lu C, Xu J, Wang S,
Xue L, Zhang X and Li M: ROBO3 promotes growth and metastasis of
pancreatic carcinoma. Cancer Lett. 366:61–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Li Y, Lv Z, He G, Wang J, Zhang X, Lu G,
Ren X, Wang F, Zhu X, Ding Y, et al: The SOX17/miR-371-5p/SOX2 axis
inhibits EMT, stem cell properties and metastasis in colorectal
cancer. Oncotarget. 6:9099–9112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Xu W, Ji J, Xu Y, Liu Y, Shi L, Liu Y, Lu
X, Zhao Y, Luo F, Wang B, et al: MicroRNA-191, by promoting the EMT
and increasing CSC-like properties, is involved in neoplastic and
metastatic properties of transformed human bronchial epithelial
cells. Mol Carcinog. 54 (Suppl 1):E148–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Li Y, Sun D, Gao J, Shi Z, Chi P, Meng Y,
Zou C and Wang Y: MicroRNA-373 promotes the development of
endometrial cancer by targeting LATS2 and activating the
Wnt/β-Catenin pathway. J Cell Biochem. 2018.
|
|
186
|
Yang Z, Wang XL, Bai R, Liu WY, Li X, Liu
M and Tang H: miR-23a promotes IKKα expression but suppresses ST7L
expression to contribute to the malignancy of epithelial ovarian
cancer cells. Br J Cancer. 115:731–740. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Wang C, Wang X, Su Z, Fei H, Liu X and Pan
Q: MiR-25 promotes hepatocellular carcinoma cell growth, migration
and invasion by inhibiting RhoGDI1. Oncotarget. 6:36231–36244.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Hao J, Jin X, Shi Y and Zhang H: miR-93-5p
enhance lacrimal gland adenoid cystic carcinoma cell tumorigenesis
by targeting BRMS1L. Cancer Cell Int. 18:722018. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Tang J, Li L, Huang W, Sui C, Yang Y, Lin
X, Hou G, Chen X, Fu J, Yuan S, et al: MiR-429 increases the
metastatic capability of HCC via regulating classic Wnt pathway
rather than epithelial-mesenchymal transition. Cancer Lett.
364:33–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Song Q, Xu Y, Yang C, Chen Z, Jia C, Chen
J, Zhang Y, Lai P, Fan X, Zhou X, et al: miR-483-5p promotes
invasion and metastasis of lung adenocarcinoma by targeting RhoGDI1
and ALCAM. Cancer Res. 74:3031–3042. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Wang H, Yan B, Zhang P, Liu S, Li Q, Yang
J, Yang F and Chen E: MiR-496 promotes migration and
epithelial-mesenchymal transition by targeting RASSF6 in colorectal
cancer. J Cell Physiol. 235:1469–1479. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Liu L, Tian YC, Mao G, Zhang YG and Han L:
MiR-675 is frequently overexpressed in gastric cancer and enhances
cell proliferation and invasion via targeting a potent anti-tumor
gene PITX1. Cell Signal. 62:1093522019. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Zhang X, Peng Y, Huang Y, Yang M, Yan R,
Zhao Y, Cheng Y, Liu X, Deng S, Feng X, et al: SMG-1 inhibition by
miR-192/-215 causes epithelial-mesenchymal transition in gastric
carcinogenesis via activation of Wnt signaling. Cancer Med.
7:146–156. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Pei YF, Yin XM and Liu XQ: TOP2A induces
malignant character of pancreatic cancer through activating
β-catenin signaling pathway. Biochim Biophys Acta Mol Basis Dis.
1864:197–207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Guo YH, Wang LQ, Li B, Xu H, Yang JH,
Zheng LS, Yu P, Zhou AD, Zhang Y, Xie SJ, et al: Wnt/β-catenin
pathway transactivates microRNA-150 that promotes EMT of colorectal
cancer cells by suppressing CREB signaling. Oncotarget.
7:42513–42526. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Liang H, Wang C, Gao K, Li J and Jia R:
MuicroRNA-421 promotes the progression of nonsmall cell lung cancer
by targeting HOPX and regulating the Wnt/β-catenin signaling
pathway. Mol Med Rep. 20:151–161. 2019.PubMed/NCBI
|
|
197
|
Listing H, Mardin WA, Wohlfromm S, Mees ST
and Haier J: MiR-23a/-24-induced gene silencing results in
mesothelial cell integration of pancreatic cancer. Br J Cancer.
112:131–139. 2015. View Article : Google Scholar : PubMed/NCBI
|