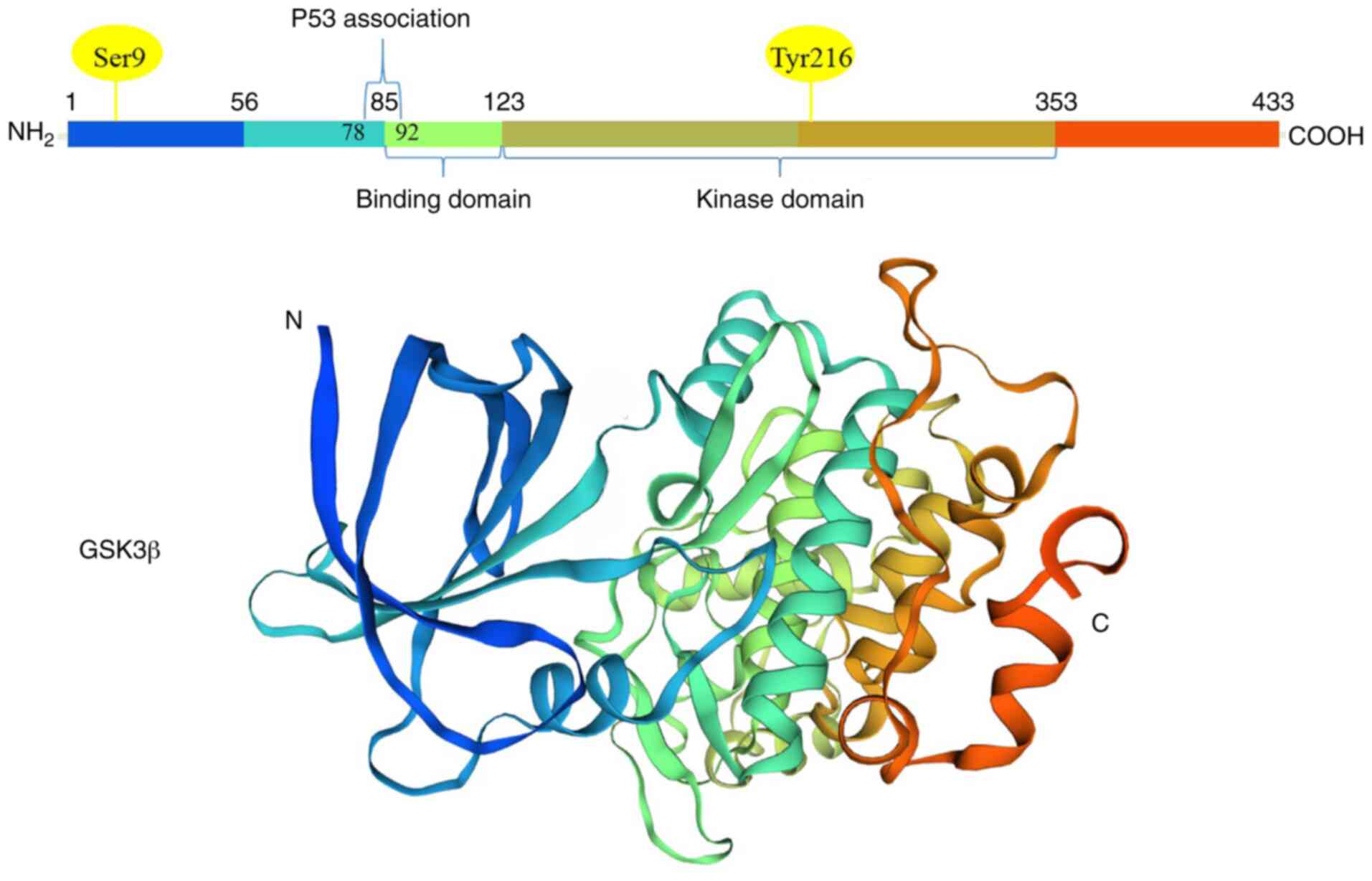
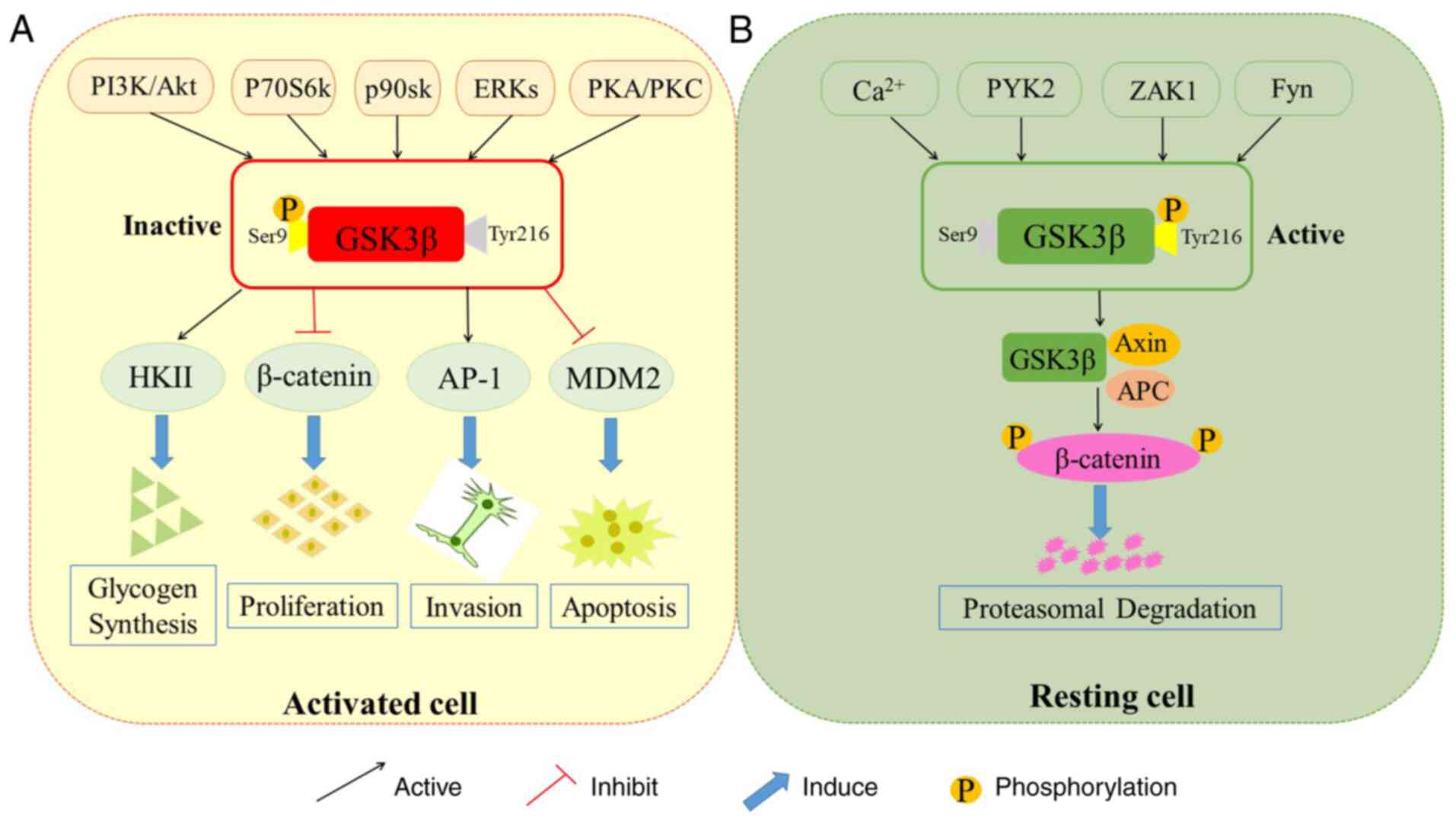
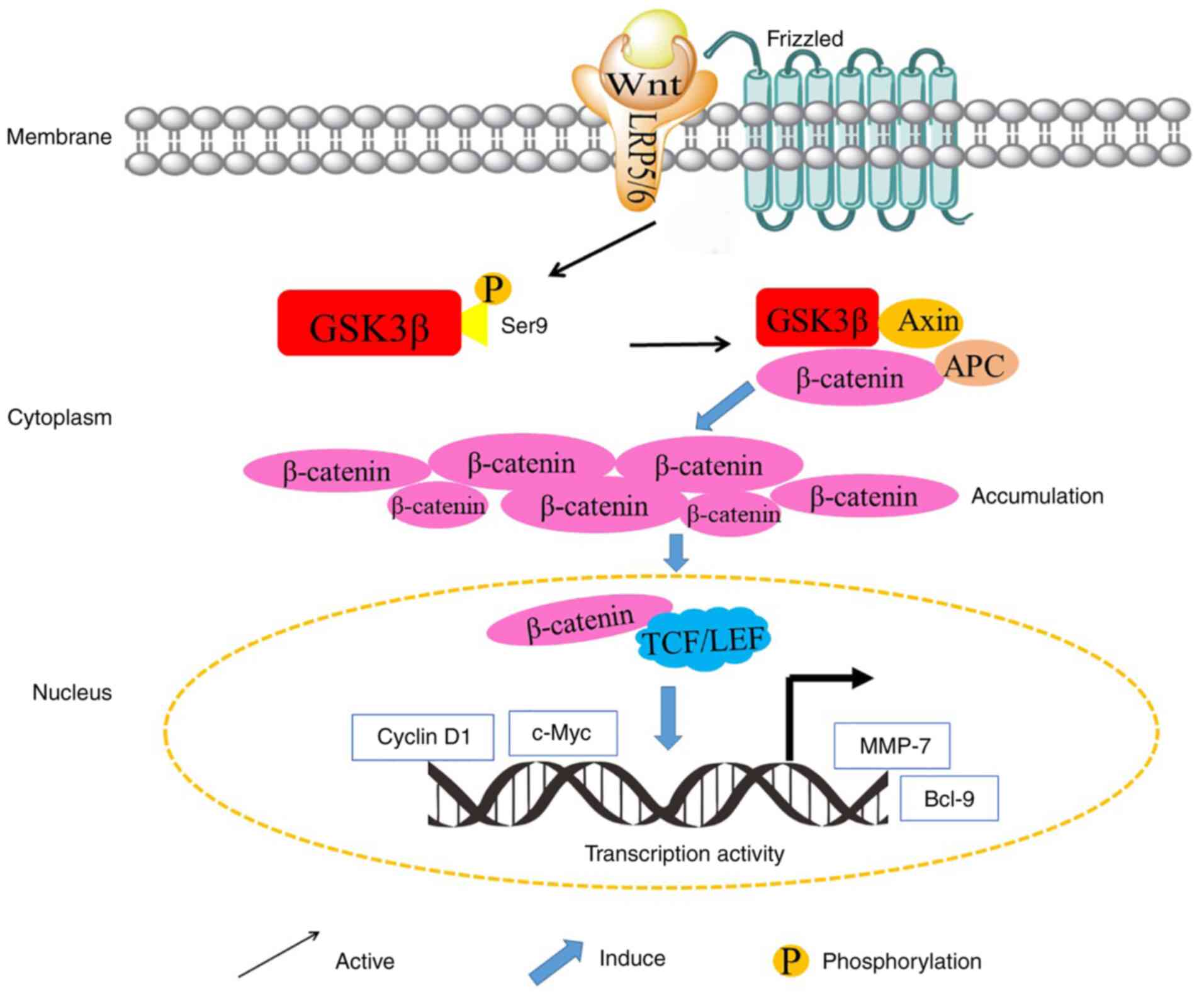
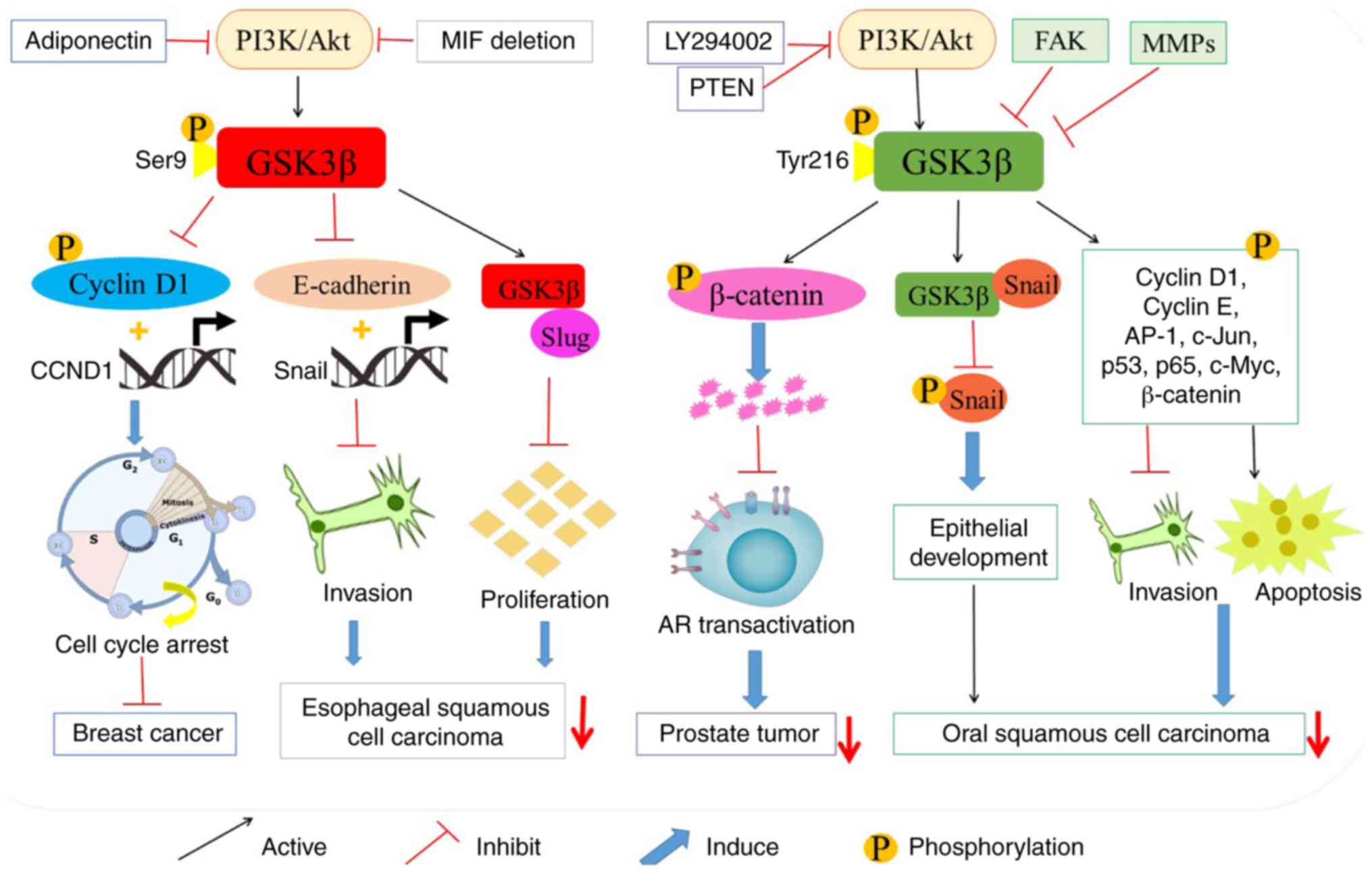
Glycogen synthase kinase 3 (GSK3), a member of the
mitogen-activated protein (MAP) kinase superfamily, has two highly
conserved homologous isoforms, GSK 3α (51 kDa) and GSK 3β (47 kDa).
Discovered in 1980, both of these two isoforms are multifunctional
serine and threonine protein kinases, and they are widely expressed
in all eukaryotes (1–3). Encoded by the human chromosome 19q13.2
and 3q13.33, respectively, GSK 3α and GSK 3β share 98% identical
sequences in their kinase domains (4,5).
Despite structural similarities, their functions are different by
phosphorylation at distinctive sites (5). For the activation of GSK 3α and GSK
3β, tyrosine 279 (Y279) and tyrosine 216 (Y216) are phosphorylated
by upstream signaling molecules respectively, and these two sites
are located at the T-loop of GSK 3 (activation domain) (Fig. 1). Notably, GSK 3α and GSK 3β are
inactivated by site-specific phosphorylation, which is tightly
controlled by diverse mechanisms (6). All these mechanisms for the
inactivation of GSK 3α at Ser21 and GSK 3β at Ser9 have been
revealed to be attributed to the phosphoinositide3 kinase
(PI3K)-dependent mechanism (7).
Activated by PI3 kinase, protein kinase A (PKA), protein kinase B
(PKB) (also termed as Akt), protein kinase C (PKC) and p90Rsk
contribute to the inactivation of GSK 3, which ultimately leads to
the dephosphorylation of GSK 3 substrates (8). Initially, the inactivation of GSK 3
was considered to be a critical mediator in glycogen metabolism and
insulin signaling, since it could give rise to the phosphorylation
of glycogen synthase and could promote glycogen synthesis (9). Currently, emerging evidence has
demonstrated that GSK 3, particularly GSK 3β, is well-established
as a vital component in signaling pathways of cell regulation,
involving cell proliferation, differentiation, motility, apoptosis
and the cell cycle (1,2,10). As
an attractive therapeutic target, GSK 3 has been revealed to be
implicated in the pathogenesis of numerous human diseases, such as
diabetes, inflammation, Alzheimer's disease, cardiovascular and
bipolar disease (11–13). In addition, GSK 3β has been
demonstrated to be a transcription factor regulating tumor
progression, invasion and metastasis (14,15).
In different types of cancers, GSK 3β plays a controversial role as
a tumor suppressor or a tumor promoter (16,17).
Furthermore, GSK 3β has been demonstrated to be a key regulator of
chemo-resistance and radio-resistance in tumor treatment (18,19).
Therefore, the regulative roles of GSK 3β in tumor development and
progression require elucidation. In addition, to date, it has been
revealed that GSK 3β inhibitors are potential therapeutic drugs in
cancer therapy, thus, pre-clinical and clinical studies of GSK 3β
inhibitors are also presented.
It has been revealed that the activity of GSK 3β
could be regulated in a substrate-specific manner, which comprises
four key mechanisms: Phosphorylation, subcellular localization, the
formation of protein complexes and the phosphorylation status of
GSK 3β substrates (12). Among
them, phosphorylation is deemed to be the most important regulatory
mechanism of GSK 3β, which would lead to the inactivation of GSK 3β
at Ser 9. The activated PI3K/Akt signaling pathway phosphorylates
GSK 3β at inhibitory serine residues and the phosphorylation may
result in the activation of transcription factors in response to
carcinogenesis, for instance activating protein-1 (AP-1), cyclic
adenosine monophosphate (cAMP)-response element binding protein
(CREB), β-catenin, c-Myc and nuclear factor-kappa (NF-κB) (1,20). In
addition, upstream proteins of GSK 3β, including P70S6 kinase,
extracellular signal-regulated kinases (ERKs), p90sk (also called
MAPKAP kinase-1), protein kinase C (PKC) and cAMP-dependent protein
kinase [protein kinase A, (PKA)], could lead to the inactivation of
GSK 3β (1,21,22).
Each regulatory kinase is influenced by GSK 3β distribution, while
scaffolding proteins could restrain the interaction between kinases
(6) (Fig. 2A). It has been established that the
phosphorylation of tyrosine at 216 site of GSK 3β (Tyr216) is
regulated by calcium-dependent tyrosine kinase, proline-rich
tyrosine kinase 2 (PYK2), cAMP-activated protein tyrosine kinase,
Zaphod kinase 1 (ZAK1) or Fyn (23,24).
Additionally, phosphorylation of GSK 3β at Tyr216 is regulated by
mitogen-activated protein kinase (MEK1/2) (25). Generally, GSK 3β is classified as an
active constituent enzyme in the cytoplasm of resting cells. When
activated, GSK 3β is accumulated in the nucleus and mitochondria
(26) (Fig. 2B). Notably, by activating the genes
of downstream transcription factors and recruiting the aggregation
of these transcription factors, nuclear translocation of GSK 3β
regulates a great number of transcription factors (27).
More than 40 proteins are downregulated by GSK 3β.
These proteins are involved in a broad range of cellular processes,
including protein synthesis, cell differentiation, proliferation
and apoptosis (2,10,14).
Substrates that are closely related to tumorigenesis and cancer
development tend to be dephosphorylated. In the canonical Wnt
signaling pathway, β-catenin, an important element of Wnt
signaling, is controlled by GSK 3β (28). With the absence of Wnt signaling,
activated GSK 3β forms a complex with adenomatous polyposis coli
(APC) and the transcriptional co-activator β-catenin, and therefore
the complex binds with the scaffolding protein Axin. This newly
formed complex enables GSK 3β to phosphorylate β-catenin and leads
to the degradation of β-catenin. Conversely, the Wnt signaling
pathway could inactivate GSK 3β and prevent the phosphorylation of
β-catenin by GSK 3β (29). In
addition, β-catenin binds to the T-cell factor (TCF)/lymphocyte
enhancer factor (LEF), and ultimately promotes the transcription of
TCF/LEF transcription factor family-related genes, including
proto-oncogenes, such as c-Myc, cyclin D1 and the vascular
endothelial growth factor (VEGF), as well as genes that regulate
cell invasion and migration, including matrix metalloproteinase-7
(MMP-7) (30–32). Therefore, as the upstream gene of
β-catenin, GSK 3β plays a pivotal role in cell proliferation,
migration, metastasis and differentiation by controlling Wnt
signaling. Additionally, other proto-oncogenic or tumor suppressing
transcription factors (such as p53) and translation factors are the
substrates of GSK 3β. With regard to DNA damage studies, the
increasingly activated GSK 3β in the nucleus and mitochondria
promotes the expression of the tumor suppressor protein p53, and
forms a complex with p53, which enhances the response of
p53-induced apoptosis (33,34). Furthermore, GSK 3β directly
phosphorylates Kruppel-like factor 6 (KLF6), a zinc-finger
transcription factor which is a tumor suppressor, inhibiting the
tumor growth by increasing KLF6-mediated growth suppression through
p53-independent transactivation of p21 (35) (Fig.
3).
For decades, accumulating research has been
exploring the role of GSK 3β in human oncology. Based on the
studies of the GSK 3β signaling mechanism, the regulation of GSK 3β
in cancer progression is intricate (14,36).
It has been demonstrated that GSK 3β acts as a tumor suppressor in
prostate cancer, by inhibiting androgen receptor-mediated cell
growth (37). Conversely, it has
been observed in colon cancer cell lines and colorectal cancer
patients that the expression levels and the enzyme activities of
GSK 3β were higher than normal cells, therefore GSK 3β appears to
be a tumor promoter (38). Herein,
in order to further elucidate GSK 3β as a potential therapeutic
target for various cancers, a comprehensive review about the
multiple roles of GSK 3β in tumorigenesis is provided.
Since GSK 3β downregulates numerous proto-oncogenic
proteins and cell cycle checkpoint proteins, GSK 3β has been
generally recognized as a tumor suppressor by inhibiting cell
proliferation (39). Previous
studies have demonstrated that PI3K inhibitor LY294002 and tumor
suppressor phosphatase and tension homolog deleted on chromosome 10
(PTEN) stimulate GSK 3β-mediated degradation of β-catenin by
inhibiting the PI3K/Akt pathway, thereby inhibiting the progression
of prostate tumors (40,41). In addition, GSK 3β is phosphorylated
and inactivated by the activation of PI3K/Akt pathway, and
therefore, inactivated GSK 3β prevents the phosphorylation of
cyclin D1, inhibits the expression of cyclin D1 gene (CCND1), and
accumulates cyclin D1 at the nucleus of breast cancer cells
(42). Moreover, it has been
revealed that adiponectin could prevent Akt-induced phosphorylation
of GSK 3β and decrease intracellular accumulation and nuclear
activity of β-catenin, subsequently reducing the expression of
cyclin D1 (43). This mechanism
leads to the apoptosis of breast cancer cells MDA-MB-231 and T47D,
and induces the cell cycle arrest of MDA-MB-231 cells at G0-G1
phase. Additionally, it has been revealed by in vivo
experiments that the adiponectin-mediated suppressive effect on the
GSK 3β/β-catenin signaling pathway could reduce mammary
tumorigenesis in female nude mice (43). Therefore, GSK 3β is perceived as a
tumor suppressor in the tumorigenesis and progression of breast
cancer (Fig. 4).
Emerging evidence has revealed that GSK 3β
phosphorylates various tumor factors (TFs), including cyclin D1,
cyclin E, AP-1, c-Jun, p53, p65, Snail, c-Myc and β-catenin, which
would facilitate their degradation and prevent them from entering
the nucleus. Thereby, their activities are downregulated in oral
squamous cell carcinoma (OSCC) (44–51).
It has been indicated that activated GSK 3β binds to the
mesenchymal Snail gene. Then it inhibits the phosphorylation,
degradation and cytoplasmic translocation of Snail, and
subsequently inhibits p-cadherin-induced development and
homeostasis of epithelial architecture. Consequently, GSK 3β may
reverse the p-cadherin-contributed oncogenesis in OSCC (52). In addition, it has been demonstrated
that the overexpression of focal adhesion kinase (FAK) may activate
NF-κB by inactivating GSK 3β, and therefore promotes cancer cell
migration. In addition, the inactivation of GSK 3β has also been
revealed to suppress cell apoptosis in pre-invasive and invasive
OSCC (51,53,54).
Furthermore, as the induction of intracellular reactive oxygen
species (ROS) stimulates cell apoptosis (55), constitutively activated GSK 3β has
been reported to increase the mitochondrial membrane potential and
to promote ROS (56,57). In addition, several matrix
metalloproteinases (MMPs) can also facilitate the migration of
cancer cells, thereby upregulating and activating tumor factors,
such as Snail, AP-1 and NF-κB in OSCC (58,59).
Collectively, GSK 3β could be a potential therapeutic target for
the treatment of OSCC by suppressing tumorigenesis (Fig. 4).
It has been indicated that the overexpression of
macrophage migration inhibitory factor (MIF) is associated with the
development and progression of esophageal squamous cell carcinoma
(ESCC) (60,61). Further study has demonstrated that
the deletion of MIF attenuates Akt-dependent GSK 3β phosphorylation
and restores tumor suppressor activity of GSK 3β (61). After MIF knockdown, the activation
of GSK 3β prevents the transcription and expression of Snail gene,
one of activators of epithelial-mesenchymal transition (EMT)
(62). Moreover, it has also been
revealed to inhibit the expression of E-cadherin in ESCC (61). GSK 3β not only inhibits cell
survival and proliferation, but also restrains tumor invasion and
metastasis. GSK 3β interacts with Slug, the zinc-finger-containing
transcriptional repressor, by suppressing E-cadherin expression and
promoting cancer cell migration, invasion and metastasis in the
non-small cell lung cancers (NSCLCs) (63). Notably, GSK 3β also reduces the
stability of Slug protein by promoting the C-terminus of
Hsc70-interacting protein (CHIP)-mediated Slug degradation.
Conversely, GSK 3β inhibitors could induce the accumulation of
non-degradable Slug, which increases the migratory and invasive
capabilities of lung cancer cells (63). Collectively, the activation of GSK
3β may offer a new prospect for inhibiting the tumorigenesis and
development of tumors (Fig. 4).
GSK 3β also plays a critical role in tumor cell
proliferation. Overexpression of GSK 3β has been observed in
various tumor types, including colon, liver, ovarian and pancreatic
tumors (14,64). It has been previously reported that
GSK 3β inhibitors (SB-216763 and AR-A014418) significantly suppress
the growth of patient-derived xenograft (PDX) colon cancer, and GSK
3β inhibitors are anticipated to be therapeutic agents for colon
cancer (64). The overexpression of
nuclear GSK 3β and the loss of membrane β-catenin are substantially
correlated with poor survival, distant metastasis and worse
prognosis of colon cancer patients (65). It is further discovered that the
formation of nuclear GSK 3β and β-catenin could decrease the
transcription of TCF/LEF target gene (66). In addition, it has been clarified
that troglitazone, an agonist of peroxisome proliferator activated
receptor γ (PPARγ), could suppress the activity of NF-κB by
inhibiting the activity of GSK 3β, reducing the expression of G0/G1
phase regulatory proteins including Cdk2, Cdk4 and cyclin B1, and
increasing the cleavage of apoptosis-associated proteins, such as
caspase-3 and caspase-9 of human colon cancer and prostate cancer
(67,68). A previous study has revealed that
p53+/+ colorectal cancer cells treated with GSK 3β
inhibitor or GSK 3β silencing RNAs could facilitate the increase of
p53-dependent apoptosis, instead of cell cycle arrest (69). Furthermore, the activation of
PI3K/Akt signaling would result in the inactivation of GSK 3β,
which promotes cell growth by inhibiting apoptosis-related proteins
including Bcl-2 family in 1, 2-dimethylhydrazine (DMH)-induced
colorectal cancer (70).
In addition, the overexpression of GSK 3β and the
nuclear transcription of NF-κB play critical roles in the survival
and proliferation of pancreatic cancer cells (71). However, GSK 3β knockdown has been
revealed to decrease the expression of Bcl-2 and VEGF, and
ultimately result in the growth arrest of tumors (72). In addition, by increasing the
expression of cyclin D1 and facilitating the proliferation of
ovarian cancer cells, activation of GSK 3β could promote the
activation of NF-κB and lead to cell cycle entering of tumor cells
into the S phase (73).
Accumulating evidence has revealed that GSK 3β could affect
intracellular glucose metabolism by restraining the p53-, NF-κB-
and c-Myc-mediated pathways, and thereafter promote the
proliferation and survival of glioblastoma cells and protect them
from apoptosis (20,74). Markedly, GSK 3β could transfer from
the cytoplasm to the nucleus in glioblastoma (GBM) when stimulated
by ionizing radiation (IR), and could bind to p53 binding protein 1
(53BP1) at Ser166. The binding of GSK 3β with 53BP1 results in the
increased DNA double-strand break (DSB) repair in GBM after IR
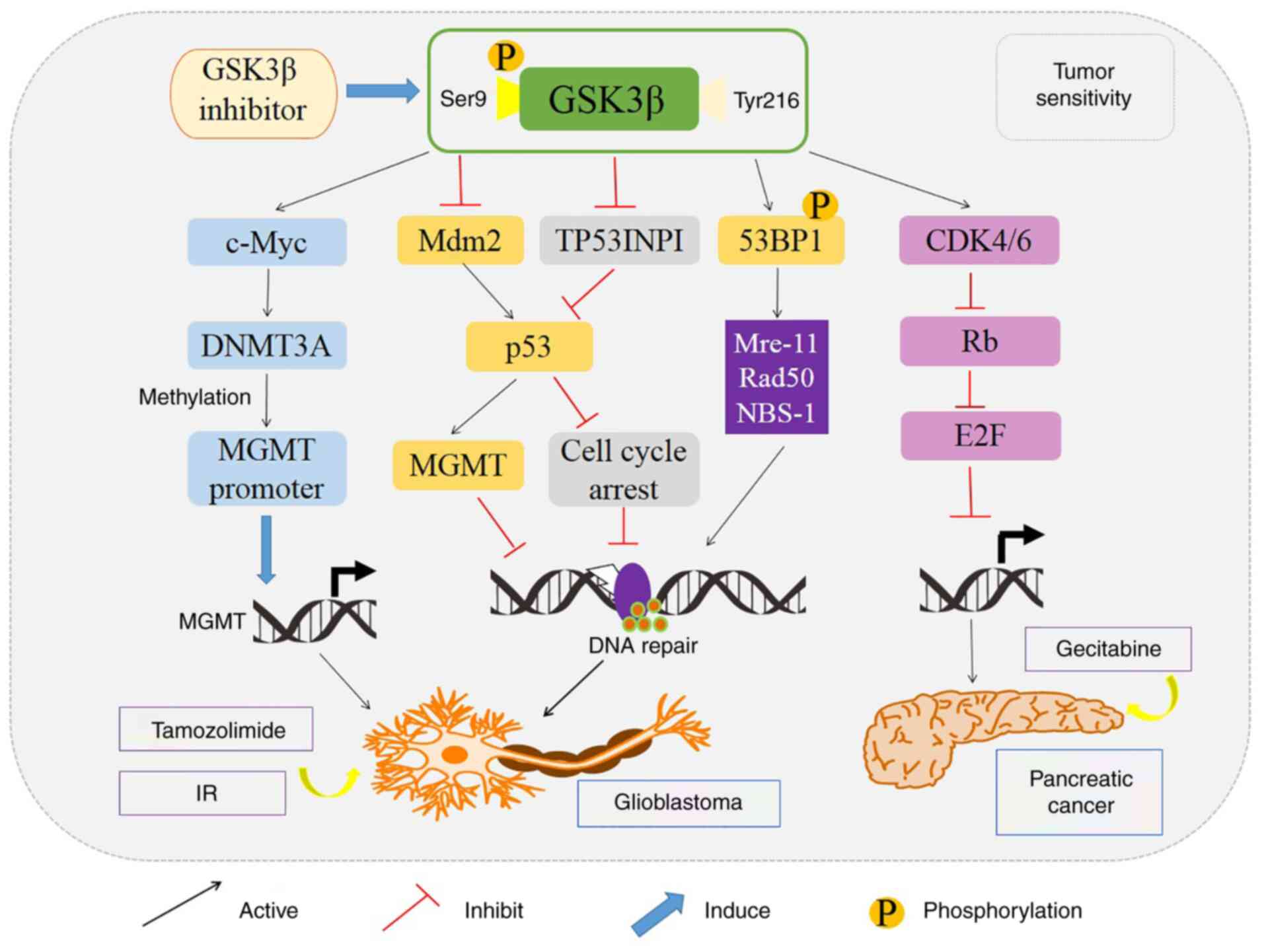
(75) (Fig. 5). Notably, GSK 3β inhibitor,
SB216763, comparatively decreases the proliferation of GBM cells
and induces apoptosis of GBM cells by halting DSB repair.
Similarly, it has been revealed that GSK 3β inhibitor could block
the NF-κB pathway and reduce the NF-κB-mediated transcription in
osteosarcoma (OSA) (76).
However, as the present inhibitors are not specific
but targeting both GSK 3α and GSK 3β, the oncogenic role of GSK 3β
has been explored by gain and loss-of-function approaches of
transgenic mice, including both knockout and knock-in animal
models. A study by Kerkela et al revealed that the GSK
3β−/− mice exhibited hypertrophic myopathy, which was
caused by cardiomyocyte hyperproliferation with increased
expression and nuclear localization of three important regulators
of proliferation (GATA4, cyclin D1, and c-Myc) (77). Furthermore, Hoeflich et al
reported that disruption of the GSK gene resulted in embryonic
lethality caused by severe liver degeneration during mid-gestation
with the activation of the transcription factor activation NF-κB,
and the apoptotic sensitivity of GSK 3β−/− fibroblasts
was a direct consequence of GSK 3β deficiency (78). Using GSK 3β knock-in mice, studies
have revealed that GSK-3 supports the maintenance of MLL leukemia
cells by promoting continuous degradation of the cyclin-dependent
kinase inhibitor p27Kip1 (79,80).
In addition, phosphorylation of mTOR, p70S6K, and 4E-BP1 were
impaired in GSK 3β knock-in mice (81). Therefore, GSK 3β can be perceived as
an oncogenic protein and GSK 3β inhibitors are potential new drugs
in cancer therapy.
GSK 3β is also considered to be a pivotal mediator
in regulating the sensitivity of tumor cells for chemotherapy and
radiotherapy. It has been observed that the expression of GSK 3β is
significantly increased in paclitaxel-resistant ovarian carcinoma
cell line (SKOV3) (82). Moreover,
downregulation of GSK 3β increased the therapeutic effect of
5-fluorouracil (5-FU), abolished cell viability and colony growth,
and reduced the xenograft tumor mass in various drug-resistant
p53-null colon cancer cell lines, which implies that GSK 3β
inhibitor, in combination with chemotherapy, may represent a novel
strategy for the treatment of chemotherapy-resistant tumors
(83). GSK 3β inhibition was
revealed to allow chemo-resistant carcinoma cells to become more
susceptible to the synthetic multi-kinase inhibitor (sorafenib)
which inhibits cell growth and angiogenesis (84). In addition, the increased expression
of GSK 3β phosphorylation at Ser9 has been revealed to promote the
chemo-sensitivity in cisplatin-resistant CP70 cells. The pGSK 3β
(Ser9) increased the cisplatin resistance of ovarian carcinomas by
reducing the GSK 3β regulated stabilization of p53 expression
(85). Furthermore, LY294002, a
PI3K inhibitor, has been revealed to enhance chemotherapy-induced
apoptosis and the cleavage of caspase-8 by promoting GSK 3β
dephosphorylation at Ser9 (86).
Similarly, in MCF-7 breast cancer cells, selective inhibition of
GSK 3β attenuated cytotoxicity induced by histone deacetylase
inhibitor trichostatin A (87).
Additionally, GSK 3β inhibition also enhanced the sensitivity of
glioblastoma cells to radiation (75). In glioblastoma, GSK 3β inhibition
improved the therapeutic effect of temozolomide by modulating the
expression of O6-methylguanine DNA methyl
transferase (MGMT) promoter via c-Myc signaling pathway (74,88).
Moreover, GSK 3β inhibitor enhanced the sensitivity of pancreatic
cancer to gemcitabine by decreasing the expression of tumor protein
53-induced-nuclear-protein 1 (TP53INP1), a pro-apoptotic
stress-induced or DSB repair-mediated p53 target gene, and by
downregulating the CDK4/6 complex-dependent phosphorylation of Rb
tumor suppressor protein (89,90)
(Fig. 5).
Collectively, GSK 3β regulates various responses to
chemotherapy among a variety of tumor cell types, which is not
entirely consistent with its role as a tumor suppressor or tumor
promoter. On the one hand, GSK 3β inhibitor acts as a tumor
promoter to attenuate the sensitivity of breast cancer cells to
chemotherapy. On the other hand, GSK 3β functions as a tumor
promoter for pancreatic cancer and GSK 3β inhibitor enhances the
sensitivity of pancreatic cancer cells to chemotherapy. Therefore,
the mechanisms of the GSK 3β signaling pathway in tumorigenesis and
drug responses are significantly diverse, and require further
investigations.
GSK 3β has been revealed to play a critical role in
the regulation of a variety of cellular functions, including cell
proliferation, differentiation, motility, apoptosis and the cell
cycle (1,2,10). In
addition, GSK3 inhibitors may be appropriate for the treatment of
certain diseases, including diabetes, bipolar disorder,
inflammation and certain types of cancer (91). Inhibitors of GSK 3β are categorized
as GSK 3β-selective or non-selective and ATP-competitive or
non-ATP-competitive (92–121).
Lithium, a well-documented inhibitor of GSK3, can
promote phosphorylation of GSK 3β at Ser9 and inhibit the activity
of GSK 3β by competing with magnesium (Mg2+) rather than
ATP or its substrate. It has been indicated that lithium has become
a gold standard for the treatment of bipolar mood and nervous
disorders (92,93). Additionally, lithium has an effect
on embryonic development by promoting the differentiation of
epithelial cells, increasi calcngium (Ca2+) storage in
the bones, and hindering the entering of the cell cycle (94–96).
Lithium has been revealed to induce apoptosis in both Jurkat cells
and differentiated immortalized hippocampal neurons by stimulating
the death domain-containing receptor Fas (97). Notably, the treatment with lithium
was revealed to increase the levels of p21WAF/Cip1 (a
protein with anti-apoptotic function) and survivin (a protein that
supports the growth of cells by suppressing apoptosis and promoting
cell proliferation) in human GBM cells (98). Furthermore, lithium was revealed to
inhibit cell motility and compromise the invasive phenotype of
v-Src-transformed cells, which was mediated by the activation of
phospho-tyrosine phosphatases via the regulation of the cell redox
status (99).
To date, several compounds targeting GSK 3 are still
under pre-clinical studies, and few of them are in clinical trials
(Table I). The mechanisms of GSK3
inhibition greatly vary. Both lithium and SB216763 have been
demonstrated to have effects on tumor growth by inhibiting cell
proliferation, however 9-ING-41 tends to induce apoptosis (100). SB216763, an ATP-competitive
inhibitor of GSK 3β, has been revealed to downregulate AR-mediated
prostate cancer cell growth in vitro and in vivo
(101,102). It has also been revealed to
potentiate the death ligand-induced apoptotic response in
pancreatic cancer cells (PANC1 and MIA PaCa-2) by activating the
c-Jun N-terminal kinase (JNK) pathway (103). Similarly, SB-415286, another
ATP-competitive inhibitor specific for GSK 3β, was demonstrated to
induce the expression of a β-catenin-LEF/TCF regulated reporter
gene (104). Its treatment
potentiated TRAIL- and CH-11-induced apoptosis in HepG2 cells, and
induced growth arrest and apoptosis in multiple myeloma (MM) cells
(105,106). Additionally, it decreased the
viability of Neuro-2A cells, and induced G2/M arrest (107). Non-ATP competitive GSK 3β
inhibitors, including LiCl, TDZD-8 and L803-mts, could also induce
significant inhibition of tumor growth (108). A previous study has revealed that
TWS119 could reverse the effects of tamoxifen on β-catenin and
Snail expression thereby inhibiting GSK-3β/β-catenin activation
(109). Furthermore, TWS119
exhibited different effects on CD4+ and CD8+
T cells in tumor-infiltrating lymphocytes (TILs) by stimulating the
expansion of naive T cell and CD8 stem cell-like memory T cells,
and inducing CD8+ effector T-cell proliferation in TILs
(110). The selective GSK 3β
inhibitor, manzamine A, can reduce the metastatic potential of
AsPC-1 pancreatic cancer cells and promote TRAIL-induced cell
apoptosis (111). Inhibition of
GSK 3β with AR-A014418 was also revealed to increase the
sensitivity of PANC1 cells to gemcitabine by regulating the Rb/E2F
pathway (93) (112).
Notably, 9-ING-41
[benzofuran-3-yl-(indol-3-yl)maleimide], a maleimide-based
ATP-competitive small molecule GSK-3β inhibitor, has been
demonstrated to be more selective for GSK 3β than for other related
kinases (113). In vitro
cell line studies and in vivo animal models have revealed
the antitumor effects of 9-ING-41 in various cancers, including
bladder cancer (114), renal
cancer (115), neuroblastoma
(116), B-cell lymphoma (117), breast cancer (118) and glioblastoma (119). A previous study on 9-ING-41
revealed that it upregulates the phosphorylation of the inhibitory
serine residue of GSK 3β in ovarian cancer cells, and subsequently
induces the apoptosis of ovarian cancer cells (120). A study from Ugolkov et al
demonstrated that 9-ING-41 leads to the regression of
patient-derived xenograft (PDX) tumors from metastatic pleural
effusions obtained from patients with progressive, chemo-refractory
breast cancer (118). Their group
also revealed that 9-ING-41 significantly enhanced
1-(2-chloroethyl)-3-cyclohexyl-1-nitrosourea (CCNU) antitumor
activity in chemo-resistant PDX models of GBM by decreasing tumor
size and increasing overall survival (119). In addition, 9-ING-41 could
attenuate the progression of pulmonary fibrosis by improving lung
functions and inhibition of myofibroblast differentiation in lung
fibroblasts ex vivo and pulmonary fibrosis in vivo
(121). With significant
pre-clinical antitumor activity, 9-ING-41 is under phase 1/2
clinical trials in patients with refractory hematologic
malignancies or solid Tumors (Clinical Trials ID: NCT03678883)
(https://clinicaltrials.gov/ct2/show/NCT03678883).
A previous study has revealed the immune-regulatory
role of GSK 3β via the phosphorylation of two important
transcription factors, NF-κB and CREB (122). Moreover, the phosphorylation of
these two transcription factors could result in proteasomal
degradation. In addition, the inactivation of GSK 3β has been
revealed to lead to the nuclear transfer of NF-κB and CREB, which
stimulate the secretion of inflammation-related cytokines (123,124).
For the innate immune system, the NK lymphocyte
plays a critical role, and its activation relies on specific
receptors including NKG2D/2B4 or NKG2D/DNAM-1. However, these
activators may inhibit the activity of GSK 3β via the ERK or Akt
signaling pathways (125,126). Studies on GSK 3β knockdown and its
inhibitors revealed that the inactivation or silencing of GSK 3β
leads to NK cell activation and enhanced function (127,128). A study on MM cells indicated that
the inhibition of GSK3 upregulated MICA transcription and
translation both in MM cell lines and in tumor cells isolated from
MM patients, without significant effects on the basal expression of
the MICB and DNAM-1 ligand poliovirus receptor/CD155. Moreover, GSK
3 inhibitors increased NK-mediated cytotoxicity of MM cells by
activating NK cell degranulation. In addition, combined with
lenalidomide or melphalan, treatment with GSK-3 inhibitors induced
MICA expression and increased NK cell-mediated tumor killing by
promoting NKG2D recognition in NK cells. Therefore, GSK-3
inhibitors could be novel therapeutic drugs targeting MICA
expression and improving immune response in MM patients (129). For the acute myelogenous leukemia
(AML) patients, NK cells expressed high levels of GSK 3β.
Therefore, treatment with the GSK 3 inhibitors or the genetic
inactivation of the GSK 3 led to the increased activity of NK cells
to kill AML cells. Furthermore, GSK3 inhibition promoted the
formation AML-NK cell conjugates by upregulating LFA expression on
NK cells and by inducing ICAM-1 expression on AML cells (130). Another study on GSK 3 revealed
that its inhibitor significantly upregulated transcription factors
associated with late-stage NK-cell maturation, such as E-box
binding homeobox 2 (ZEB2), PR/SET domain 1 (PRDM1), and T-box 21
(TBX21) and subsequently increased CD57 acquisition and maturation.
In addition, NK cells, which were expanded ex vivo in the
presence of GSK3 inhibitor, produced more TNF and IFN-γ, natural
cytotoxicity and antibody-dependent cellular cytotoxicity (131). Therefore, currently, there are
several ongoing clinical trials of adaptive NK cells treated with
GSK 3 inhibitors for relapsed AML, ovarian cancer as well as other
solid tumors.
Accumulating evidence has demonstrated the
significance of GSK 3 in T cells. Because it is constitutively
active in resting T cells, GSK 3 acts as a negative regulator of
T-cell response by inhibiting CD8+ T-cell proliferation
and IL-2 production (132). The
inactivation of GSK 3 can specifically downregulate PD-1 expression
by enhancing CD8+ CTL function and clearance of lymphoma
cells (133,134). Additionally, GSK-3 inhibition was
revealed to be as effective as anti-PD-1 and PD-L1 blocking
antibodies in suppressing the growth of melanoma and lymphoma in
mouse models (135). Therefore,
GSK-3 was revealed as a central regulator of PD-1 expression.
Recently, the same group revealed that GSK-3 also negatively
regulated lymphocyte activation gene-3 (LAG-3) expression on
CD4+ and CD8+ T cells. The combination of
GSK-3 inhibitor with LAG-3 blockade resulted in the suppression of
B16 melanoma growth and enhancement of tumor clearance, by
increasing the expression of the transcription factor T-bet and
binding with the LAG-3 promoter, and subsequently increasing
granzyme B and interferon-γ1 expression (136). GSK 3β inhibition has been reported
to increase the tumor cell cytotoxic capacity of CD8+
memory stem T cells in vitro against gastric cancer cells
(137). Moreover, the inactivation
of GSK 3β has been revealed to stimulate iTreg differentiation and
increase the suppressive activity via the activation of the
TGF-β/Smad3 signaling pathway (138). Furthermore, treatment of
GBM-specific IL-13 CAR-T cells with the GSK 3 inhibitor resulted in
reduced PD-1 expression, increased T cell survival and
proliferation. Therefore, GSK 3 inactivation has also been revealed
to contribute to the tumor treatment in CAR-T cell immunotherapy
(139). Because the polarized-M2
phenotype of macrophages affects tumor growth, invasiveness,
angiogenesis (140), and GSK 3β
has been reported to enhance the polarization of microglia toward
M2 (141), GSK 3β may be the
potential therapeutic target for cancer immunotherapy.
Collectively, these studies revealed that GSK 3 inhibition could be
a new immunotherapeutic strategy for oncotherapy.
It has been widely acknowledged that GSK 3β is
involved in tumorigenesis, cancer progression and metastasis.
However, there are still controversies about the role of GSK 3β as
a tumor suppressor or tumor promoter. GSK 3β may function as a
tumor suppressor which inhibits neoplastic growth in prostate, oral
and lung cancer when overexpressed or activated (51,142–144). Conversely, GSK 3β has also been
revealed to facilitate carcinogenesis, and is recognized as a
‘tumor promoter’ in colon, pancreatic and ovarian cancers (65,89,90,120,145). Therefore, based on its pivotal
role in tumorigenesis and tumor progression, GSK 3β is predicted to
be a clinical prognostic indicator for certain malignant cancers.
In conclusion, a more comprehensive understanding of GSK 3β would
be comparatively beneficial to disease therapy. Targeting GSK 3β
may be an ideal therapeutic strategy for malignant tumors that are
characterized by infinite proliferation, metastasis and
chemo-resistance, thus providing significant insight for the
further research of tumor treatment.
Not applicable.
The present study was supported by the National
Natural Science Foundation of China (grant no. 81802504), the
National Key Research and Development Program of China (grant no.
2020YFC2005500), the Key Research and Development Program of
Science and Technology of the Department of Sichuan Province (grant
no. 2019YFS0514) and the Sichuan Medical Association (grant no.
Q19037).
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the present
study.
YW conceived and designed the review. RH and SD
wrote the paper. TL and XX prepared the figures and table. YW
reviewed and edited the manuscript. All authors read and approved
the manuscript, and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
This review does not contain any studies with human
participants or animals performed by any of the authors.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Grimes CA and Jope RS: The multifaceted
roles of glycogen synthase kinase 3beta in cellular signaling. Prog
Neurobiol. 65:391–426. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Doble BW and Woodgett JR: GSK-3: Tricks of
the trade for a multi-tasking kinase. J Cell Sci. 116:1175–1186.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Embi N, Rylatt DB and Cohen P: Glycogen
synthase kinase-3 from rabbit skeletal muscle. Separation from
cyclic-AMP-dependent protein kinase and phosphorylase kinase. Eur J
Biochem. 107:519–527. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Woodgett JR: Molecular cloning and
expression of glycogen synthase kinase-3/factor A. EMBO J.
9:2431–2438. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thotala DK and Yazlovitskaya EM: GSK3B
(glycogen synthase kinase 3 beta). Atlas Genet Cytogenet Oncol
Haematol. 15:7–10. 2011.
|
|
6
|
Jope RS and Johnson GV: The glamour and
gloom of glycogen synthase kinase-3. Trends Biochem Sci. 29:95–102.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang X, Yu SX, Lu Y, Bast RC Jr, Woodgett
JR and Mills GB: Phosphorylation and inactivation of glycogen
synthase kinase 3 by protein kinase A. Proc Natl Acad Sci USA.
97:11960–11965. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cross DA, Alessi DR, Cohen P, Andjelkovich
M and Hemmings BA: Inhibition of glycogen synthase kinase-3 by
insulin mediated by protein kinase B. Nature. 378:785–789. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Summers SA, Kao AW, Kohn AD, Backus GS,
Roth RA, Pessin JE and Birnbaum MJ: The role of glycogen synthase
kinase 3beta in insulin-stimulated glucose metabolism. J Biol Chem.
274:17934–17940. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jacobs KM, Bhave SR, Ferraro DJ, Jaboin
JJ, Hallahan DE and Thotala D: GSK-3β: A bifunctional role in cell
death pathways. Int J Cell Biol. 2012:9307102012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Amar S, Belmaker RH and Agam G: The
possible involvement of glycogen synthase kinase-3 (GSK-3) in
diabetes, cancer and central nervous system diseases. Curr Pharm
Des. 17:2264–2277. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jope RS, Yuskaitis CJ and Beurel E:
Glycogen synthase kinase-3 (GSK3): Inflammation, diseases, and
therapeutics. Neurochem Res. 32:577–595. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo J: The role of GSK3beta in the
development of the central nervous system. Front Biol (Beijing).
7:212–220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo J: Glycogen synthase kinase 3beta
(GSK3beta) in tumorigenesis and cancer chemotherapy. Cancer Lett.
273:194–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiara F and Rasola A: GSK-3 and
mitochondria in cancer cells. Front Oncol. 3:162013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Naito S, Bilim V, Yuuki K, Ugolkov A,
Motoyama T, Nagaoka A, Kato T and Tomita Y: Glycogen synthase
kinase-3beta: A prognostic marker and a potential therapeutic
target in human bladder cancer. Clin Cancer Res. 16:5124–5132.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang QL, Xie XB, Wang J, Chen Q, Han AJ,
Zou CY, Yin JQ, Liu DW, Liang Y, Zhao ZQ, et al: Glycogen synthase
kinase-3β, NF-κB signaling, and tumorigenesis of human
osteosarcoma. J Natl Cancer Inst. 104:749–763. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Majewska E and Szeliga M: AKT/GSK3β
signaling in glioblastoma. Neurochem Res. 42:918–924. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Domoto T, Pyko IV, Furuta T, Miyashita K,
Uehara M, Shimasaki T, Nakada M and Minamoto T: Glycogen synthase
kinase-3β is a pivotal mediator of cancer invasion and resistance
to therapy. Cancer Sci. 107:1363–1372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kotliarova S, Pastorino S, Kovell LC,
Kotliarov Y, Song H, Zhang W, Bailey R, Maric D, Zenklusen JC, Lee
J and Fine HA: Glycogen synthase kinase-3 inhibition induces glioma
cell death through c-MYC, nuclear factor-kappaB, and glucose
regulation. Cancer Res. 68:6643–6651. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Najib S and Sánchez-Margalet V:
Homocysteine thiolactone inhibits insulin-stimulated DNA and
protein synthesis: Possible role of mitogen-activated protein
kinase (MAPK), glycogen synthase kinase-3 (GSK-3) and p70 S6K
phosphorylation. J Mol Endocrinol. 34:119–126. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eldar-Finkelman H, Seger R, Vandenheede JR
and Krebs EG: Inactivation of glycogen synthase kinase-3 by
epidermal growth factor is mediated by mitogen-activated protein
kinase/p90 ribosomal protein S6 kinase signaling pathway in NIH/3T3
cells. J Biol Chem. 270:987–990. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hartigan JA, Xiong WC and Johnson GV:
Glycogen synthase kinase 3beta is tyrosine phosphorylated by PYK2.
Biochem Biophys Res Commun. 284:485–489. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hartigan JA and Johnson GV: Transient
increases in intracellular calcium result in prolonged
site-selective increases in Tau phosphorylation through a glycogen
synthase kinase 3beta-dependent pathway. J Biol Chem.
274:21395–21401. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Takahashi-Yanaga F, Shiraishi F, Hirata M,
Miwa Y, Morimoto S and Sasaguri T: Glycogen synthase kinase-3beta
is tyrosine-phosphorylated by MEK1 in human skin fibroblasts.
Biochem Biophys Res Commun. 316:411–415. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bijur GN and Jope RS: Glycogen synthase
kinase-3 beta is highly activated in nuclei and mitochondria.
Neuroreport. 14:2415–2419. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Diehl JA, Cheng M, Roussel MF and Sherr
CJ: Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis
and subcellular localization. Genes Dev. 12:3499–3511. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Manoukian AS and Woodgett JR: Role of
glycogen synthase kinase-3 in cancer: Regulation by Wnts and other
signaling pathways. Adv Cancer Res. 84:203–229. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Valvezan AJ, Zhang F, Diehl JA and Klein
PS: Adenomatous polyposis coli (APC) regulates multiple signaling
pathways by enhancing glycogen synthase kinase-3 (GSK-3) activity.
J Biol Chem. 287:3823–3832. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peifer M and Polakis P: Wnt signaling in
oncogenesis and embryogenesis-a look outside the nucleus. Science.
287:1606–1609. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lustig B and Behrens J: The Wnt signaling
pathway and its role in tumor development. J Cancer Res Clin Oncol.
129:199–221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Giles RH, van Es JH and Clevers H: Caught
up in a Wnt storm: Wnt signaling in cancer. Biochim Biophys Acta.
1653:1–24. 2003.PubMed/NCBI
|
|
33
|
Watcharasit P, Bijur GN, Zmijewski JW,
Song L, Zmijewska A, Chen X, Johnson GV and Jope RS: Direct,
activating interaction between glycogen synthase kinase-3beta and
p53 after DNA damage. Proc Natl Acad Sci USA. 99:7951–7955. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Watcharasit P, Bijur GN, Song L, Zhu J,
Chen X and Jope RS: Glycogen synthase kinase-3beta (GSK3beta) binds
to and promotes the actions of p53. J Biol Chem. 278:48872–48879.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lang UE, Kocabayoglu P, Cheng GZ,
Ghiassi-Nejad Z, Muñoz U, Vetter D, Eckstein DA, Hannivoort RA,
Walsh MJ and Friedman SL: GSK3β phosphorylation of the KLF6 tumor
suppressor promotes its transactivation of p21. Oncogene.
32:4557–4564. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin J, Song T, Li C and Mao W: GSK-3β in
DNA repair, apoptosis, and resistance of chemotherapy, radiotherapy
of cancer. Biochim Biophys Acta Mol Cell Res. 1867:1186592020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang L, Lin HK, Hu YC, Xie S, Yang L and
Chang C: Suppression of androgen receptor-mediated transactivation
and cell growth by the glycogen synthase kinase 3 beta in prostate
cells. J Biol Chem. 279:32444–32452. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shakoori A, Ougolkov A, Yu ZW, Zhang B,
Modarressi MH, Billadeau DD, Mai M, Takahashi Y and Minamoto T:
Deregulated GSK3beta activity in colorectal cancer: Its association
with tumor cell survival and proliferation. Biochem Biophys Res
Commun. 334:1365–1373. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Takahashi-Yanaga F and Sasaguri T:
GSK-3beta regulates cyclin D1 expression: A new target for
chemotherapy. Cell Signal. 20:581–589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Verras M and Sun Z: Roles and regulation
of Wnt signaling and beta-catenin in prostate cancer. Cancer Lett.
237:22–32. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sharma M, Chuang WW and Sun Z:
Phosphatidylinositol 3-kinase/Akt stimulates androgen pathway
through GSK3beta inhibition and nuclear beta-catenin accumulation.
J Biol Chem. 277:30935–30941. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Quintayo MA, Munro AF, Thomas J, Kunkler
IH, Jack W, Kerr GR, Dixon JM, Chetty U and Bartlett JM: GSK3β and
cyclin D1 expression predicts outcome in early breast cancer
patients. Breast Cancer Res Treat. 136:161–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Y, Lam JB, Lam KS, Liu J, Lam MC, Hoo
RL, Wu D, Cooper GJ and Xu A: Adiponectin modulates the glycogen
synthase kinase-3beta/beta-catenin signaling pathway and attenuates
mammary tumorigenesis of MDA-MB-231 cells in nude mice. Cancer Res.
66:11462–11470. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Baral R, Patnaik S and Das BR:
Co-overexpression of p53 and c-myc proteins linked with advanced
stages of betel- and tobacco-related oral squamous cell carcinomas
from eastern India. Eur J Oral Sci. 106:907–913. 2010. View Article : Google Scholar
|
|
45
|
de Sousa SO, Mesquita RA, Pinto DS Jr and
Gutkind S: Immunolocalization of c-Fos and c-Jun in human oral
mucosa and in oral squamous cell carcinoma. J Oral Pathol Med.
31:78–81. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Franz M, Spiegel K, Umbreit C, Richter P,
Codina-Canet C, Berndt A, Altendorf-Hofmann A, Koscielny S, Hyckel
P, Kosmehl H, et al: Expression of Snail is associated with
myofibroblast phenotype development in oral squamous cell
carcinoma. Histochem Cell Biol. 131:651–660. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Iwai S, Katagiri W, Kong C, Amekawa S,
Nakazawa M and Yura Y: Mutations of the APC, beta-catenin, and axin
1 genes and cytoplasmic accumulation of beta-catenin in oral
squamous cell carcinoma. J Cancer Res Clin Oncol. 131:773–782.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Buss H, Dörrie A, Schmitz ML, Frank R,
Livingstone M, Resch K and Kracht M: Phosphorylation of serine 468
by GSK-3beta negatively regulates basal p65 NF-kappaB activity. J
Biol Chem. 279:49571–49574. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Goto H, Kawano K, Kobayashi I, Sakai H and
Yanagisawa S: Expression of cyclin D1 and GSK-3beta and their
predictive value of prognosis in squamous cell carcinomas of the
tongue. Oral Oncol. 38:549–556. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mishra A, Bharti AC, Saluja D and Das BC:
Transactivation and expression patterns of Jun and Fos/AP-1
super-family proteins in human oral cancer. Int J Cancer.
126:819–829. 2010.PubMed/NCBI
|
|
51
|
Mishra R: Glycogen synthase kinase 3 beta:
Can it be a target for oral cancer. Mol Cancer. 9:1442010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bauer K, Dowejko A, Bosserhoff AK,
Reichert TE and Bauer RJ: P-cadherin induces an epithelial-like
phenotype in oral squamous cell carcinoma by GSK-3beta-mediated
Snail phosphorylation. Carcinogenesis. 30:1781–1788. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kornberg LJ: Focal adhesion kinase
expression in oral cancers. Head Neck. 20:634–639. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ko BS, Chang TC, Chen CH, Liu CC, Kuo CC,
Hsu C, Shen YC, Shen TL, Golubovskaya VM, Chang CC, et al:
Bortezomib suppresses focal adhesion kinase expression via
interrupting nuclear factor-kappa B. Life Sci. 86:199–206. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Takeuchi H, Taoka R, Mmeje CO, Jinesh GG,
Safe S and Kamat AM: CDODA-Me decreases specificity protein
transcription factors and induces apoptosis in bladder cancer cells
through induction of reactive oxygen species. Urol Oncol.
34:337.e11–e18. 2016. View Article : Google Scholar
|
|
56
|
Miller WP, Toro AL, Barber AJ and Dennis
MD: REDD1 Activates a ROS-generating feedback loop in the retina of
diabetic mice. Invest Ophthalmol Vis Sci. 60:2369–2379. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Deng S, Dai G, Chen S, Nie Z, Zhou J, Fang
H and Peng H: Dexamethasone induces osteoblast apoptosis through
ROS-PI3K/AKT/GSK3β signaling pathway. Biomed Pharmacother.
110:602–608. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ziober BL, Silverman SS Jr and Kramer RH:
Adhesive mechanisms regulating invasion and metastasis in oral
cancer. Crit Rev Oral Biol Med. 12:499–510. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Erdem NF, Carlson ER, Gerard DA and Ichiki
AT: Characterization of 3 oral squamous cell carcinoma cell lines
with different invasion and/or metastatic potentials. J Oral
Maxillofac Surg. 65:1725–1733. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhao J and Liao K: Expression of
macrophage migration inhibitory factor in esophageal squamous cell
carcinoma and normal esophageal tissue. Acta Acad Med Mil Tertiae.
29:740–742. 2008.
|
|
61
|
Liu RM, Sun DN, Jiao YL, Wang P, Zhang J,
Wang M, Ma J, Sun M, Gu BL, Chen P, et al: Macrophage migration
inhibitory factor promotes tumor aggressiveness of esophageal
squamous cell carcinoma via activation of Akt and inactivation of
GSK3β. Cancer Lett. 412:289–296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang H, Wang HS, Zhou BH, Li CL, Zhang F,
Wang XF, Zhang G, Bu XZ, Cai SH and Du J: Epithelial-mesenchymal
transition (EMT) induced by TNF-α requires AKT/GSK-3β-mediated
stabilization of snail in colorectal cancer. PLoS One.
8:e566642013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kao SH, Wang WL, Chen CY, Chang YL, Wu YY,
Wang YT, Wang SP, Nesvizhskii AI, Chen YJ, Hong TM and Yang PC:
GSK3β controls epithelial-mesenchymal transition and tumor
metastasis by CHIP-mediated degradation of Slug. Oncogene.
33:3172–3182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Shakoori A, Mai W, Miyashita K, Yasumoto
K, Takahashi Y, Ooi A, Kawakami K and Minamoto T: Inhibition of
GSK-3 beta activity attenuates proliferation of human colon cancer
cells in rodents. Cancer Sci. 98:1388–1393. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Vidri RJ and Fitzgerald TL: GSK-3: An
important kinase in colon and pancreatic cancers. Biochim Biophys
Acta Mol Cell Res. 1867:1186262020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Huang W, Chang HY, Fei T, Wu H and Chen
YG: GSK3 beta mediates suppression of cyclin D2 expression by tumor
suppressor PTEN. Oncogene. 26:2471–2482. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ban JO, Oh JH, Son SM, Won D, Song HS, Han
SB, Moon DC, Kang KW, Song MJ and Hong JT: Troglitazone, a PPAR
agonist, inhibits human prostate cancer cell growth through
inactivation of NFΚB via suppression of GSK-3β expression. Cancer
Biol Ther. 12:288–296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ban JO, Kwak DH, Oh JH, Park EJ, Cho MC,
Song HS, Song MJ, Han SB, Moon DC, Kang KW and Hong JT: Suppression
of NF-kappaB and GSK-3beta is involved in colon cancer cell growth
inhibition by the PPAR agonist troglitazone. Chem Biol Interact.
188:75–85. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Ghosh JC and Altieri DC: Activation of
p53-dependent apoptosis by acute ablation of glycogen synthase
kinase-3beta in colorectal cancer cells. Clin Cancer Res.
11:4580–4588. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Perse M and Cerar A: Morphological and
molecular alterations in 1,2 dimethylhydrazine and azoxymethane
induced colon carcinogenesis in rats. J Biomed Biotechnol.
2011:4739642011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ougolkov AV, Fernandez-Zapico ME, Bilim
VN, Smyrk TC, Chari ST and Billadeau DD: Aberrant nuclear
accumulation of glycogen synthase kinase-3beta in human pancreatic
cancer: Association with kinase activity and tumor
dedifferentiation. Clin Cancer Res. 12:5074–5081. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhou W, Wang L, Gou SM, Wang TL, Zhang M,
Liu T and Wang CY: ShRNA silencing glycogen synthase kinase-3 beta
inhibits tumor growth and angiogenesis in pancreatic cancer. Cancer
Lett. 316:178–186. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cao Q, Lu X and Feng YJ: Glycogen synthase
kinase-3beta positively regulates the proliferation of human
ovarian cancer cells. Cell Res. 16:671–677. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Miyashita K, Kawakami K, Nakada M, Mai W,
Shakoori A, Fujisawa H, Hayashi Y, Hamada J and Minamoto T:
Potential therapeutic effect of glycogen synthase kinase 3beta
inhibition against human glioblastoma. Clin Cancer Res. 15:887–897.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yang Y, Lei T, Du S, Tong R, Wang H, Yang
J, Huang J, Sun M, Wang Y and Dong Z: Nuclear GSK3β induces DNA
double-strand break repair by phosphorylating 53BP1 in
glioblastoma. Int J Oncol. 52:709–720. 2018.PubMed/NCBI
|
|
76
|
Nishimura H, Nakamura O, Yamagami Y, Mori
M, Horie R, Fukuoka N and Yamamoto T: GSK-3 inhibitor inhibits cell
proliferation and induces apoptosis in human osteosarcoma cells.
Oncol Rep. 35:2348–2354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kerkela R, Kockeritz L, Macaulay K, Zhou
J, Doble BW, Beahm C, Greytak S, Woulfe K, Trivedi CM, Woodgett JR,
et al: Deletion of GSK-3beta in mice leads to hypertrophic
cardiomyopathy secondary to cardiomyoblast hyperproliferation. J
Clin Invest. 118:3609–3618. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Hoeflich KP, Luo J, Rubie EA, Tsao MS, Jin
O and Woodgett JR: Requirement for glycogen synthase kinase-3beta
in cell survival and NF-kappaB activation. Nature. 406:86–90. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
McManus EJ, Sakamoto K, Armit LJ,
Ronaldson L, Shpiro N, Marquez R and Alessi DR: Role that
phosphorylation of GSK3 plays in insulin and Wnt signalling defined
by knockin analysis. EMBO J. 24:1571–1583. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang Z, Smith KS, Murphy M, Piloto O,
Somervaille TC and Cleary ML: Glycogen synthase kinase 3 in MLL
leukaemia maintenance and targeted therapy. Nature. 455:1205–1209.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Vigneron F, Dos Santos P, Lemoine S,
Bonnet M, Tariosse L, Couffinhal T, Duplaà C and Jaspard-Vinassa B:
GSK-3β at the crossroads in the signalling of heart
preconditioning: Implication of mTOR and Wnt pathways. Cardiovasc
Res. 90:49–56. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Fu Y, Hu D, Qiu J, Xie X, Ye F and Lu WG:
Overexpression of glycogen synthase kinase-3 in ovarian carcinoma
cells with acquired paclitaxel resistance. Int J Gynecol Cancer.
21:439–444. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Grassilli E, Narloch R, Federzoni E,
Ianzano L, Pisano F, Giovannoni R, Romano G, Masiero L, Leone BE,
Bonin S, et al: Inhibition of GSK3B bypass drug resistance of
p53-null colon carcinomas by enabling necroptosis in response to
chemotherapy. Clin Cancer Res. 19:3820–3831. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Kawazoe H, Bilim VN, Ugolkov AV, Yuuki K,
Naito S, Nagaoka A, Kato T and Tomita Y: GSK-3 inhibition in vitro
and in vivo enhances antitumor effect of sorafenib in renal cell
carcinoma (RCC). Biochem Biophys Res Commun. 423:490–495. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Cai G, Wang J, Xin X, Ke Z and Luo J:
Phosphorylation of glycogen synthase kinase-3 beta at serine 9
confers cisplatin resistance in ovarian cancer cells. Int J Oncol.
31:657–662. 2007.PubMed/NCBI
|
|
86
|
Beurel E, Kornprobst M, Blivet-Van
Eggelpoël MJ, Cadoret A, Capeau J and Desbois-Mouthon C: GSK-3beta
reactivation with LY294002 sensitizes hepatoma cells to
chemotherapy-induced apoptosis. Int J Oncol. 27:215–222.
2005.PubMed/NCBI
|
|
87
|
Alao JP, Stavropoulou AV, Lam WF and
Coombes RC: Role of glycogen synthase kinase 3 beta (GSK3beta) in
mediating the cytotoxic effects of the histone deacetylase
inhibitor trichostatin A (TSA) in MCF-7 breast cancer cells. Mol
Cancer. 5:402006. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pyko IV, Nakada M, Sabit H, Teng L,
Furuyama N, Hayashi Y, Kawakami K, Minamoto T, Fedulau AS and
Hamada J: Glycogen synthase kinase 3β inhibition sensitizes human
glioblastoma cells to temozolomide by affecting O6-methylguanine
DNA methyltransferase promoter methylation via c-Myc signaling.
Carcinogenesis. 34:2206–2217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Shimasaki T, Ishigaki Y, Nakamura Y,
Takata T, Nakaya N, Nakajima H, Sato I, Zhao X, Kitano A, Kawakami
K, et al: Glycogen synthase kinase 3β inhibition sensitizes
pancreatic cancer cells to gemcitabine. J Gastroenterol.
47:321–333. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kitano A, Shimasaki T, Chikano Y, Nakada
M, Hirose M, Higashi T, Ishigaki Y, Endo Y, Takino T, Sato H, et
al: Aberrant glycogen synthase kinase 3β is involved in pancreatic
cancer cell invasion and resistance to therapy. PLoS One.
8:e552892013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Maqbool M and Hoda N: GSK3 inhibitors in
the therapeutic development of diabetes, cancer and
neurodegeneration: Past, present and future. Curr Pharm Des.
23:4332–4350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bowden CL: Efficacy of lithium in mania
and maintenance therapy of bipolar disorder. J Clin Psychiatry. 61
(Suppl 9):S35–S40. 2000.
|
|
93
|
Del Grande C, Muti M, Musetti L,
Pergentini I, Corsi M, Turri M, Soldani I, Corsini GU and Dell'Osso
L: Long-term treatment of bipolar disorder: How should we use
lithium salts? Riv Psichiatr. 47:515–526. 2012.(In Italian).
PubMed/NCBI
|
|
94
|
Clément-Lacroix P, Ai M, Morvan F,
Roman-Roman S, Vayssière B, Belleville C, Estrera K, Warman ML,
Baron R and Rawadi G: Lrp5-independent activation of Wnt signaling
by lithium chloride increases bone formation and bone mass in mice.
Proc Natl Acad Sci USA. 102:17406–17411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Stump RJ, Lovicu FJ, Ang SL, Pandey SK and
McAvoy JW: Lithium stabilizes the polarized lens epithelial
phenotype and inhibits proliferation, migration, and epithelial
mesenchymal transition. J Pathol. 210:249–257. 2010. View Article : Google Scholar
|
|
96
|
Sun A, Shanmugam I, Song J, Terranova PF,
Thrasher JB and Li B: Lithium suppresses cell proliferation by
interrupting E2F-DNA interaction and subsequently reducing S-phase
gene expression in prostate cancer. Prostate. 67:976–988. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Song L, Zhou T and Jope RS: Lithium
facilitates apoptotic signaling induced by activation of the Fas
death domain-containing receptor. BMC Neurosci. 5:202004.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Karlovic D, Jakopec S, Dubravcic K,
Batinic D, Buljan D and Osmak M: Lithium increases expression of
p21 WAF/Cip1 and survivin in human glioblastoma cells. Cell Biol
Toxicol. 23:83–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Neel BD, Lopez J, Chabadel A and Gillet G:
Lithium suppresses motility and invasivity of v-src-transformed
cells by glutathione-dependent activation of phosphotyrosine
phosphatases. Oncogene. 28:3246–3260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Hilliard T, Muehlbauer A, Gaisina I,
Gaisin A, Gallier F, Kozikowski A and Burdette J: Novel glycogen
synthase kinase 3beta inhibitors induce apoptosis in ovarian
cancer. Biol Reprod. 83 (Suppl 1):S6912010. View Article : Google Scholar
|
|
101
|
Rinnab L, Schütz SV, Diesch J, Schmid E,
Küfer R, Hautmann RE, Spindler KD and Cronauer MV: Inhibition of
glycogen synthase kinase-3 in androgen-responsive prostate cancer
cell lines: Are GSK inhibitors therapeutically useful? Neoplasia.
10:624–634. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Schütz SV, Schrader AJ, Zengerling F,
Genze F, Cronauer MV and Schrader M: Inhibition of glycogen
synthase kinase-3β counteracts ligand-independent activity of the
androgen receptor in castration resistant prostate cancer. PLoS
One. 6:e253412011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Marchand B, Tremblay I, Cagnol S and
Boucher MJ: Inhibition of glycogen synthase kinase-3 activity
triggers an apoptotic response in pancreatic cancer cells through
JNK-dependent mechanisms. Carcinogenesis. 33:529–537. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Coghlan MP, Culbert AA, Cross DA, Corcoran
SL, Yates JW, Pearce NJ, Rausch OL, Murphy GJ, Carter PS, Roxbee
Cox L, et al: Selective small molecule inhibitors of glycogen
synthase kinase-3 modulate glycogen metabolism and gene
transcription. Chem Biol. 7:793–803. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Beurel E, Blivet-Van Eggelpoel MJ,
Kornprobst M, Moritz S, Delelo R, Paye F, Housset C and
Desbois-Mouthon C: Glycogen synthase kinase-3 inhibitors augment
TRAIL-induced apoptotic death in human hepatoma cells. Biochem
Pharmacol. 77:54–65. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Piazza F, Manni S, Tubi LQ, Montini B,
Pavan L, Colpo A, Gnoato M, Cabrelle A, Adami F, Zambello R, et al:
Glycogen Synthase Kinase-3 regulates multiple myeloma cell growth
and bortezomib-induced cell death. BMC Cancer. 10:5262010.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Dickey A, Schleicher S, Leahy K, Hu R,
Hallahan D and Thotala DK: GSK-3β inhibition promotes cell death,
apoptosis, and in vivo tumor growth delay in neuroblastoma Neuro-2A
cell line. J Neurooncol. 104:145–153. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zhu Q, Yang J, Han S, Liu J, Holzbeierlein
J, Thrasher JB and Li B: Suppression of glycogen synthase kinase 3
activity reduces tumor growth of prostate cancer in vivo. Prostate.
71:835–845. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Yan P, Tang H, Chen X, Ji S, Jin W, Zhang
J, Shen J, Deng H, Zhao X, Shen Q and Huang H: Tamoxifen attenuates
dialysate-induced peritoneal fibrosis by inhibiting
GSK-3β/β-catenin axis activation. Biosci Rep. 38:BSR201802402018.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Tang YY, Sheng SY, Lu CG, Zhang YQ, Zou
JY, Lei YY, Gu Y and Hong H: Effects of glycogen synthase kinase-3β
inhibitor TWS119 on proliferation and cytokine production of TILs
from human lung cancer. J Immunother. 41:319–328. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Guzmán EA, Johnson JD, Linley PA,
Gunasekera SE and Wright AE: A novel activity from an old compound:
Manzamine A reduces the metastatic potential of AsPC-1 pancreatic
cancer cells and sensitizes them to TRAIL-induced apoptosis. Invest
New Drugs. 29:777–785. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Mamaghani S, Patel S and Hedley DW:
Glycogen synthase kinase-3 inhibition disrupts nuclear
factor-kappaB activity in pancreatic cancer, but fails to sensitize
to gemcitabine chemotherapy. BMC Cancer. 9:1322009. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Gaisina IN, Gallier F, Ougolkov AV, Kim
KH, Kurome T, Guo S, Holzle D, Luchini DN, Blond SY, Billadeau DD
and Kozikowski AP: From a natural product lead to the
identification of potent and selective
benzofuran-3-yl-(indol-3-yl)maleimides as glycogen synthase kinase
3beta inhibitors that suppress proliferation and survival of
pancreatic cancer cells. J Med Chem. 52:1853–1863. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kuroki H, Anraku T, Kazama A, Bilim V,
Tasaki M, Schmitt D, Mazar AP, Giles FJ, Ugolkov A and Tomita Y:
9-ING-41, a small molecule inhibitor of GSK-3beta, potentiates the
effects of anticancer therapeutics in bladder cancer. Sci Rep.
9:199772019. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Anraku T, Kuroki H, Kazama A, Bilim V,
Tasaki M, Schmitt D, Mazar A, Giles FJ, Ugolkov A and Tomita Y:
Clinically relevant GSK3β inhibitor 9-ING-41 is active as a single
agent and in combination with other antitumor therapies in human
renal cancer. Int J Mol Med. 45:315–323. 2020.PubMed/NCBI
|
|
116
|
Ugolkov AV, Bondarenko GI, Dubrovskyi O,
Berbegall AP, Navarro S, Noguera R, O'Halloran TV, Hendrix MJ,
Giles FJ and Mazar AP: 9-ING-41, a small-molecule glycogen synthase
kinase-3 inhibitor, is active in neuroblastoma. Anticancer Drugs.
29:717–724. 2018.PubMed/NCBI
|
|
117
|
Karmali R, Chukkapalli V, Gordon LI,
Borgia JA, Ugolkov A, Mazar AP and Giles FJ: GSK-3β inhibitor,
9-ING-41, reduces cell viability and halts proliferation of B-cell
lymphoma cell lines as a single agent and in combination with novel
agents. Oncotarget. 8:114924–114934. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Ugolkov A, Gaisina I, Zhang JS, Billadeau
DD, White K, Kozikowski A, Jain S, Cristofanilli M, Giles F,
O'Halloran T, et al: GSK-3 inhibition overcomes chemoresistance in
human breast cancer. Cancer Lett. 380:384–392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ugolkov A, Qiang W, Bondarenko G, Procissi
D, Gaisina I, James CD, Chandler J, Kozikowski A, Gunosewoyo H,
O'Halloran T, et al: Combination treatment with the GSK-3 Inhibitor
9-ING-41 and CCNU cures orthotopic chemoresistant glioblastoma in
patient-derived xenograft models. Transl Oncol. 10:669–678. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Hilliard TS, Gaisina IN, Muehlbauer AG,
Gaisin AM, Gallier F and Burdette JE: Glycogen synthase kinase 3β
inhibitors induce apoptosis in ovarian cancer cells and inhibit
in-vivo tumor growth. Anticancer Drugs. 22:978–985. 2011.PubMed/NCBI
|
|
121
|
Jeffers A, Qin W, Owens S, Koenig KB,
Komatsu S, Giles FJ, Schmitt DM, Idell S and Tucker TA: Glycogen
synthase kinase-3β inhibition with 9-ING-41 attenuates the
progression of pulmonary fibrosis. Sci Rep. 9:189252019. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Gotschel F, Kern C, Lang S, Sparna T,
Markmann C, Schwager J, McNelly S, von Weizsäcker F, Laufer S,
Hecht A and Merfort I: Inhibition of GSK3 differentially modulates
NF-kappaB, CREB, AP-1 and beta-catenin signaling in hepatocytes,
but fails to promote TNF-alpha-induced apoptosis. Exp Cell Res.
314:1351–1366. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Cheng Y, Pardo M, Armini RS, Martinez A,
Mouhsine H, Zagury JF, Jope RS and Beurel E: Stress-induced
neuroinflammation is mediated by GSK3-dependent TLR4 signaling that
promotes susceptibility to depression-like behavior. Brain Behav
Immun. 53:207–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Hoffmeister L, Diekmann M, Brand K and
Huber R: GSK3: A kinase balancing promotion and resolution of
inflammation. Cells. 9:8202020. View Article : Google Scholar
|
|
125
|
Teng L, Meng Q, Lu J, Xie J, Wang Z, Liu Y
and Wang D: Liquiritin modulates ERK and AKT/GSK-3β-dependent
pathways to protect against glutamate-induced cell damage in
differentiated PC12 cells. Mol Med Rep. 10:818–824. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Gerhardt D, Bertola G, Dietrich F,
Figueiró F, Zanotto-Filho A, Moreira Fonseca JC, Morrone FB,
Barrios CH, Battastini AM and Salbego CG: Boldine induces cell
cycle arrest and apoptosis in T24 human bladder cancer cell line
via regulation of ERK, AKT, and GSK-3β. Urol Oncol. 32:36.e1–e9.
2014. View Article : Google Scholar
|
|
127
|
Kwon HJ, Kwon SJ, Lee H, Park HR, Choi GE,
Kang SW, Kwon SW, Kim N, Lee SY, Ryu S, et al: NK cell function
triggered by multiple activating receptors is negatively regulated
by glycogen synthase kinase-3β. Cell Signal. 27:1731–1741. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Fionda C, Soriani A, Zingoni A, Santoni A
and Cippitelli M: NKG2D and DNAM-1 ligands: Molecular targets for
NK cell-mediated immunotherapeutic intervention in multiple
myeloma. Biomed Res Int. 2015:1786982015. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Fionda C, Malgarini G, Soriani A, Zingoni
A, Cecere F, Iannitto ML, Ricciardi MR, Federico V, Petrucci MT,
Santoni A and Cippitelli M: Inhibition of glycogen synthase
kinase-3 increases NKG2D ligand MICA expression and sensitivity to
NK cell-mediated cytotoxicity in multiple myeloma cells: Role of
STAT3. J Immunol. 190:6662–6672. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Parameswaran R, Ramakrishnan P, Moreton
SA, Xia Z, Hou Y, Lee DA, Gupta K, deLima M, Beck RC and Wald DN:
Repression of GSK3 restores NK cell cytotoxicity in AML patients.
Nat Commun. 7:111542016. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Cichocki F, Valamehr B, Bjordahl R, Zhang
B, Rezner B, Rogers P, Gaidarova S, Moreno S, Tuininga K, Dougherty
P, et al: GSK3 inhibition drives maturation of NK cells and
enhances their antitumor activity. Cancer Res. 77:5664–5675. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ohteki T, Parsons M, Zakarian A, Jones RG,
Nguyen LT, Woodgett JR and Ohashi PS: Negative regulation of T cell
proliferation and interleukin 2 production by the serine threonine
kinase GSK-3. J Exp Med. 192:99–104. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Taylor A and Rudd CE: Glycogen synthase
kinase 3 inactivation compensates for the lack of CD28 in the
priming of CD8+ cytotoxic T-cells: Implications for
anti-PD-1 immunotherapy. Front Immunol. 8:16532017. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Taylor A, Harker JA, Chanthong K,
Stevenson PG, Zuniga EI and Rudd CE: Glycogen synthase kinase 3
inactivation drives T-bet-mediated downregulation of Co-receptor
PD-1 to enhance CD8(+) cytolytic T cell responses. Immunity.
44:274–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Taylor A, Rothstein D and Rudd CE:
Small-molecule inhibition of PD-1 transcription is an effective
alternative to antibody blockade in cancer therapy. Cancer Res.
78:706–717. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Rudd CE, Chanthong K and Taylor A: Small
molecule inhibition of GSK-3 specifically inhibits the
transcription of inhibitory Co-receptor LAG-3 for enhanced
anti-tumor immunity. Cell Rep. 30:2075–2082.e4. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Zhang JY, Zhao YL, Lv YP, Cheng P, Chen W,
Duan M, Teng YS, Wang TT, Peng LS, Mao FY, et al: Modulation of
CD8+ memory stem T cell activity and glycogen synthase
kinase 3β inhibition enhances anti-tumoral immunity in gastric
cancer. Oncoimmunology. 7:e14129002018. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Xia Y, Zhuo H, Lu Y, Deng L, Jiang R,
Zhang L, Zhu Q, Pu L, Wang X and Lu L: Glycogen synthase kinase 3β
inhibition promotes human iTreg differentiation and suppressive
function. Immunol Res. 62:60–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Sengupta S, Katz SC, Sengupta S and
Sampath P: Glycogen synthase kinase 3 inhibition lowers PD-1
expression, promotes long-term survival and memory generation in
antigen-specific CAR-T cells. Cancer Lett. 433:131–139. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Takeuchi H, Tanaka M, Tanaka A, Tsunemi A
and Yamamoto H: Predominance of M2-polarized macrophages in bladder
cancer affects angiogenesis, tumor grade and invasiveness. Oncol
Lett. 11:3403–3408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wang G, Shi Y, Jiang X, Leak RK, Hu X, Wu
Y, Pu H, Li WW, Tang B, Wang Y, et al: HDAC inhibition prevents
white matter injury by modulating microglia/macrophage polarization
through the GSK3β/PTEN/Akt axis. Proc Natl Acad Sci USA.
112:2853–2858. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Mazor M, Kawano Y, Zhu H, Waxman J and
Kypta RM: Inhibition of glycogen synthase kinase-3 represses
androgen receptor activity and prostate cancer cell growth.
Oncogene. 23:7882–7892. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Goc A, Al-Husein B, Katsanevas K,
Steinbach A, Lou U, Sabbineni H, DeRemer DL and Somanath PR:
Targeting Src-mediated Tyr216 phosphorylation and activation of
GSK-3 in prostate cancer cells inhibit prostate cancer progression
in vitro and in vivo. Oncotarget. 5:775–787. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Yu XJ, Han QB, Wen ZS, Ma L, Gao J and
Zhou GB: Gambogenic acid induces G1 arrest via GSK3β-dependent
cyclin D1 degradation and triggers autophagy in lung cancer cells.
Cancer Lett. 322:185–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Kunnimalaiyaan S, Gamblin TC and
Kunnimalaiyaan M: Glycogen synthase kinase-3 inhibitor AR-A014418
suppresses pancreatic cancer cell growth via inhibition of
GSK-3-mediated Notch1 expression. HPB (Oxford). 17:770–776. 2015.
View Article : Google Scholar : PubMed/NCBI
|