Introduction
Osteosarcoma (OS) is the most prevalent primary
malignant bone tumor in young adolescents (1). According to an epidemiology study of
osteosarcoma in 2009, the incidence rate of OS is ~4 cases per
million individuals in young children and adolescents in the United
States (2). The prognosis of OS is
usually unfavorable due to its aggressive nature (3). There is a high rate of disability and
mortality among patients with OS (4). Due to the lack of effective biomarkers
for an early diagnosis of OS, the majority of patients with OS are
diagnosed at an advanced stage at their first visit (5,6).
Therefore, identification of novel targets for the molecular
treatment of OS is urgently required.
Circular (circ) RNAs are a type of
endogenously-expressed covalently closed RNA transcripts with
limited protein coding ability. Increasing evidence has indicated
that circRNAs are intensively associated with cancer progression
and metastasis (7,8). circRNA homeodomain interacting protein
kinase 3 (circHIPK3) was first identified by ribominus RNA
sequencing data from six human normal tissues and seven types of
human cancer (9). circHIPK3 acts as
an oncogene in various types of malignant tumor (10–13).
Zheng et al (9) reported
that circHIPK3 increases the expression levels of miR-7-targeting
proto-oncogenes (protein tyrosine kinase 2, insulin like growth
factor 1 receptor, EGFR and YY1 transcription factor), and promotes
colorectal cancer growth and metastasis. Chen et al
(14) revealed that circHIPK3
serves as a miR-124 sponge and regulates aquaporin 3 (Gill blood
group)-mediated proliferation and migration in hepatocellular
carcinoma. To the best of our knowledge, the role of circHIPK3 in
OS remains to be elucidated.
Histone deacetylase 4 (HDAC4) is a key member of
class IIa HDACs, and HDAC4 performs a wide variety of functions.
Additionally, HDAC4 is post-transcriptionally regulated by several
microRNAs (miRNAs/miRs), such as miR-1, miR-29, miR-140, mir-155,
miR-200a, miR-206 and miR-365, in various cells (15). Our previous study demonstrated that
HDAC4 is a target of miR-140-5p, and HDAC4 was closely associated
with proliferation and apoptosis in OS (16). Zeng et al (17) demonstrated that HDAC4 is upregulated
in esophageal carcinoma (EC), and HDAC4 promotes migration of EC
cells by enhancing the epithelial-to-mesenchymal transition.
Currently, to the best of our knowledge, research on circRNAs and
HDAC4 is limited.
The present study focused on the expression and
function of circHIPK3 in OS. It was revealed that circHIPK3 served
as an oncogene, and circHIPK3 promoted OS cell proliferation,
migration and invasion through modulation of HDAC4 via sponging of
microRNA 637 (miR-637).
Materials and methods
Patients and tissue samples
A total of 12 chondroma (8 males and 4 females; age
range, 17–49 years; mean age, 28.4 years) and 12 OS (6 males and 6
females; age range, 6–16 years; mean age, 11.5 years) samples were
obtained from Shengjing Hospital (Shenyang, China) during
tumorectomy between September 2014 and February 2019. All tissue
specimens were histologically diagnosed by two pathologists
according to the criteria defined by the World Health Organization
(18). Written informed signed
consent was obtained from each patient or the parents of the
patients who were minors for the use of tissue specimens in the
present study. The Institute Research Medical Ethics Committee of
Shengjing Hospital approved the present study.
Cell culture
The human osteoblast hFOB 1.19 cell line was
cultured in DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc.). The
four human OS cell lines (HOS, MG-63, U2OS and SJSA) were cultured
in DMEM (Gibco; Thermo Fisher Scientific, Inc.). All cell lines
were purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. Media were supplemented with 10% (v/v)
FBS (Invitrogen; Thermo Fisher Scientific, Inc.), 100 IU/ml
penicillin and 100 mg/ml streptomycin (Baomanbio). hFOB 1.19 cells
were maintained at 34°C with 5% CO2. OS cell lines were
maintained in a humidified atmosphere containing 5% CO2
at 37°C.
GEO database reanalysis and
bioinformatics RNA-RNA interaction prediction
The differentially expressed miRNA data in OS were
downloaded from the GEO database (https://www.ncbi.nlm.nih.gov/geo/) GSE70415 (19) and GSE28423 (20). Differentially expressed miRNAs
between 18 ccRCC tissues and 18 paired ccRCC normal tissues were
reanalyzed using the online software GEO2R (21). The expression levels of miR-508-3p
in 18 ccRCC tissues and 18 paired ccRCC normal tissues were also
reanalyzed in GSE116251 (22). To
predict the potential circRNA-miRNA and mRNA-miRNA interactions,
circBank (23), RegRNA (version
2.0) (24) and circularRNA
interactome (25) were used
according to the corresponding instructions.
RNase R treatment
The stability of circHIPK3 was determined by an
Rnase R assay as previously described (26). Briefly, 2 mg total RNA was incubated
for 30 min at 37°C with or without 5 U/µg RNase R (Epicentre;
Illumina, Inc.) and subsequently purified using an RNeasy MinElute
Cleaning kit (Qiagen GmBH), followed by analysis by reverse
transcription (RT)-PCR.
Actinomycin D assay
The actinomycin D assay was performed as previously
described (26). HOS and U2OS cells
were exposed to 2 µg/ml actinomycin D for 30 min (Sigma-Aldrich;
Merck KGaA) at 37°C at different time points (4, 8, 12 and 24 h).
Total RNA was extracted at different time points and the expression
levels of circHIPK3 and linear HIPK3 mRNA were determined by
RT-quantitative (q)PCR.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was performed as previously described
(27), and the 2−∆∆Cq
method was used for quantification (28). Total RNA from tissue specimens and
cultured cells was extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. RT reactions were performed at 44°C for 1
h followed by 92°C for 10 min to synthesize cDNA with the
PrimeScript RT Master Mix (Takara Bio, Inc.) from 500 ng RNA.
Stem-loop reverse transcription reactions were performed for
miR-637 using the TaqMan MicroRNA detection kit (Thermo Fisher
Scientific, Inc.). The qPCR analyses were performed using SYBR
Premix Ex Taq II (Takara Bio, Inc.). β-actin and U6 were employed
as endogenous controls for mRNAs and miR-637, respectively. PCR
amplification conditions were as follows: 2 min at 95°C for one
cycle, followed by denaturation for 15 sec at 95°C and extension
for 60 sec at 60°C for 38 cycles. Primer sequences are listed in
Table I.
 | Table I.Primer and oligonucleotide sequences
used in the present study. |
Table I.
Primer and oligonucleotide sequences
used in the present study.
|
Primers/oligonucleotides | Sequence
(5′-3′) |
|---|
| circHIPK3
forward |
TATGTTGGTGGATCCTGTTCGGCA |
| circHIPK3
reverse |
TGGTGGGTAGACCAAGACTTGTGA |
| HIPK3 forward |
TGGAGACTGGGGGAAGATGA |
| HIPK3 reverse |
CACACTAACTGGCTGAGGGG |
| HDAC4 forward |
TCAGACATCTTTGGGAAGGG |
| HDAC4 reverse |
CAACCTCCATCTTGCCTTGT |
| GAPDH forward |
GTCAAGGCTGAGAACGGGAA |
| GAPDH reverse |
AAATGAGCCCCAGCCTTCTC |
| miR-637
forward |
ACUGGGGGCUUUCGGGCUCUGCGU |
| miR-637
reverse |
ACGCAGAGCCCGAAAGCCCCCAGU |
| U6 forward
primer |
CTCGCTTCGGCAGCACA |
| U6 reverse
primer |
AACGCTTCACGAATTTGCGT |
| sicircHIPK3_1
sense |
UGGCCUCACAAGUCUUGGU |
| sicircHIPK3_1
antisense |
ACCAAGACUUGUGAGGCCAUA |
| sicircHIPK3_2
sense |
GGUACUACAGGUAUGGCCUTT |
| sicircHIPK3_2
antisense |
AGGCCAUACCUGUAGUACCGA |
| siHDAC4 sense |
GAAAAGGUUUUACAGCAAATT |
| siHDAC4
antisense |
UUUGCUGUAAAACCUUUUCTG |
| miR-637 mimic |
ACUGGGGGCUUUCGGGCUCUGCGU |
| miR-637
inhibitor |
ACGCAGAGCCCGAAAGCCCCCAGU |
RNA fluorescence in situ hybridization
(FISH)
FISH was performed as previously described (27). Specific probes targeting circHIPK3
or miR-637 were synthesized by Guangzhou RiboBio Co., Ltd. The RNA
FISH assay was performed using a FISH kit (Guangzhou RiboBio Co.,
Ltd.) according to the manufacturer's protocols. In brief, HOS and
U2OS cells were seeded onto glass coverslips (0.8×0.8 cm) and
cultured to 80–95% confluence. The coverslips were rinsed twice
with PBS, fixed with 4% paraformaldehyde for 10 min at room
temperature and blocked with 5% BSA (Sigma-Aldrich; Merck KGaA) for
1 h at room temperature. Then, the coverslips were permeabilized
with 0.3% Triton X-100 (Sigma-Aldrich; Merck KGaA) for 15 min and
incubated in a hybridization solution containing 1 µM Cy3-labeled
circHIPK3 probes and 1 µM Dig-labeled locked nucleic acid miR-637
probes (Guangzhou RiboBio Co., Ltd.) supplemented with 1% BSA in a
humid chamber at 37°C overnight. The next day, the coverslips were
rinsed with a solution of 0.1% Tween-20 (Sigma-Aldrich; Merck KGaA)
in 4X sodium citrate buffer (SSC; Sigma-Aldrich; Merck KGaA) for 5
min, a solution of 0.1% Tween-20 in 2X SSC for 5 min and a solution
of 0.1% Tween-20 in 1X SSC for 5 min at 42°C in dark. Lastly, the
coverslips were washed three times with 1X PBS for 5 min at room
temperature. The signals of circHIPK3 probes or miR-637 probes were
detected using Cy5-Streptavidin (Thermo Fisher Scientific, Inc.) or
a tyramide-conjugated Alexa 488 fluorochrome TSA kit (Thermo Fisher
Scientific, Inc.). Nuclei were counterstained with
4,6-diamidino-2-phenylindole at room temperature for 10 min. The
images were acquired on a Leica SP5 confocal microscope (Leica
Micosystems GmbH).
Oligonucleotide transfection
miR-637 mimics and negative control (NC mimic),
miR-637 inhibitors and negative control (NC inhibitor), effective
siRNA oligonucleotides that targeted the splicing sites of
circHIPK3 (designed using circPrimer; version 1.2.0.5) (29) or targeted HDAC4 and scrambled
control siRNAs were synthesized by Guangzhou RiboBio Co., Ltd. When
the OS cells reached 80% confluence, the oligonucleotides (100 nM)
were transfected using Lipofectamine® RNAiMax (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
After 48 h of transfection, the OS cells were used for further
detection. The sequences of the oligonucleotides are listed in
Table I.
Cell Counting Kit 8 (CCK8) assay
The CCK8 assay was performed as previously described
(30). HOS and U2OS cells were
seeded in 96-well plates (2×103 cells/well) in 200 µl
culture medium and conditioned at 37°C with 5% CO2. At
days 1, 2, 3, 4 and 5, 10 µl CCK8 solution (Dojindo Molecular
Technologies, Inc.) was added into each well and incubated at 37°C
for 2 h according to the manufacturer's protocol. The absorbance
was measured at an optical density of 450 nm using a microplate
reader (Bio-Rad Laboratories, Inc.). The experiments were performed
in triplicate.
Transwell assay
The transwell assay was performed as previously
described (31). First, upper
chambers (Corning Inc.) were pre-coated with (for the invasion
assay; precoating for 1 h at room temperature) or without (for the
migration assay) 20 µg Matrigel (BD Biosciences). HOS and U2OS
cells (at a density of 1×105) were then seeded into the
upper chambers. Culture medium without serum and medium with 10%
FBS was added into the upper and lower chambers, respectively.
Following incubation at 37°C for 24 h, the migrated or invaded OS
cells were fixed with 4% paraformaldehyde at room temperature for
30 min, stained with crystal violet at 25°C for 1 min and counted
under a light microscope (Olympus Corporation).
Wound healing assay
The wound healing assay was performed as previously
described (32). HOS and U2OS cells
were seeded and transfected with target plasmids in 6-well plates
supplemented with culture medium with 10% FBS, and allowed to reach
70–80% confluence. A 200-µl pipette tip was used to scratch the
artificial wound across the diameter of the wells and the culture
medium was changed to serum-free medium. Wound closure was observed
at 0 and 24 h, and images were captured under a phase contrast
light microscope (magnification, ×200) at 0 and 24 h after wound
incision. Each experiment was repeated three times.
Western blot analysis
The procedure was performed as previously described
(33). Total proteins were
extracted using RIPA lysis buffer (Sigma-Aldrich; Merck KGaA) and
subsequently quantified using a bicinchoninic acid protein assay
kit (Santa Cruz Biotechnology, Inc.). Samples (20 µg/lane) were
separated via 10% SDS-PAGE and transferred onto a PVDF membrane
(Amresco, LLC). Membranes were blocked with 5% BSA (Sigma-Aldrich;
Merck KGaA) for 1 h at room temperature. Primary antibodies against
HDAC4 (dilution, 1:500; cat. no. ab234084; Abcam) and GAPDH
(dilution, 1:500; cat. no. ab8245; Abcam) were incubated with the
membranes at 4°C overnight. The subsequent day, the membranes were
incubated with secondary antibody (goat anti-mouse immunoglobulin G
HRP-conjugated; cat. no. ab205719; dilution, 1:2,000; Abcam).
Signals of targeted proteins were detected using an ECL Western
Blotting Substrate kit (cat. no. ab65623; Abcam) and bands were
analyzed with ImageJ software version 2 (National Institutes of
Health).
Dual-luciferase reporter assay
The dual-luciferase reporter assay was performed as
previously described (16).
Wild-type (WT) and mutant (MUT) circHIPK3 (LUC-circHIPK3-WT and
LUC-circHIPK3-MUT) or HDAC4 (LUC-HDAC4-WT and LUC-HDAC4-MUT)
reporter plasmids that contained WT or MUT miR-637 seed sequences
were synthesized by Shanghai GenePharma Co., Ltd. The constructed
reporter plasmids and miR-637 mimic were co-transfected into HOS
and U2OS cells using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.). After 48 h, luciferase activity
was measured and compared with Renilla luciferase activity
using the Dual-Luciferase Reporter Assay system (Promega
Corporation) according to the manufacturer's protocol.
Statistical analysis
All experiments were performed in triplicate and all
data from three independent experiments are presented as the mean ±
standard deviation. Statistical analyses were performed using
GraphPad Prism 5 software (GraphPad Software, Inc.). Survival
curves were estimated by the Kaplan-Meier method and a log-rank
test was performed to assess overall survival rate of patients with
OS. A receiver operating characteristic curve (ROC) analysis was
used to evaluate the clinical value of circHIPK3. The relationship
between circHIPK3 and miR-637 was assessed by Pearson's correlation
analysis. For comparisons, unpaired Student's t-test, Mann-Whitney
U test and one-way ANOVA followed by Dunnett's post hoc test were
performed, as appropriate. P<0.05 was considered to indicate a
statistically significant difference.
Results
circHIPK3 is relatively highly
expressed in OS cells, and predominantly localized in the
cytoplasm
First, the present study demonstrated that circHIPK3
was derived from exon 2 of HIPK mRNA, as previously reported
(26) (Fig. 1A and Fig. S1). The expression levels of
circHIPK3 were then detected in 12 OS tissues and 12 paired
chondroma tissues using RT-qPCR. As the data presented in Fig. 1B shows, circHIPK3 expression was
significantly upregulated in OS tissues. In addition, the present
study measured circHIPK3 expression in OS cells. RT-qPCR (Fig. 1C) demonstrated that circHIPK3
expression was upregulated in four OS cell lines (HOS, MG63, U2OS
and SJSA) compared with in the normal osteoblast hFOB1.19 cell
line. Furthermore, the present study revealed that circHIPK3 was
mainly located in the cytoplasm of HOS and U2OS cells (Fig. 1D). A previous study considered
circRNAs to be stable nucleic acids (34). Therefore, an RNase R assay was
performed in the present study to evaluate the stability of
circHIPK3. As shown in Fig. 1E and
F, compared with the Mock group, RNase R led to a significant
decrease in HIPK3 mRNA, while the expression of circHIPK3 was not
markedly changed. Additionally, an actinomycin D assay indicated
that circHIPK3 was more stable than the linear HIPK3 mRNA (Fig. 1G and H). Finally, the present study
analyzed the clinical value of circHIPK3 in the collected cases.
ROC curve and Kaplan-Meier analyses demonstrated that circHIPK3 had
clinical value in OS (area under the curve, 0.8750; P<0.0001;
Fig. 1I) and high circHIPK3
expression was closely associated with shorter overall survival of
patients with OS (Fig. 1J).
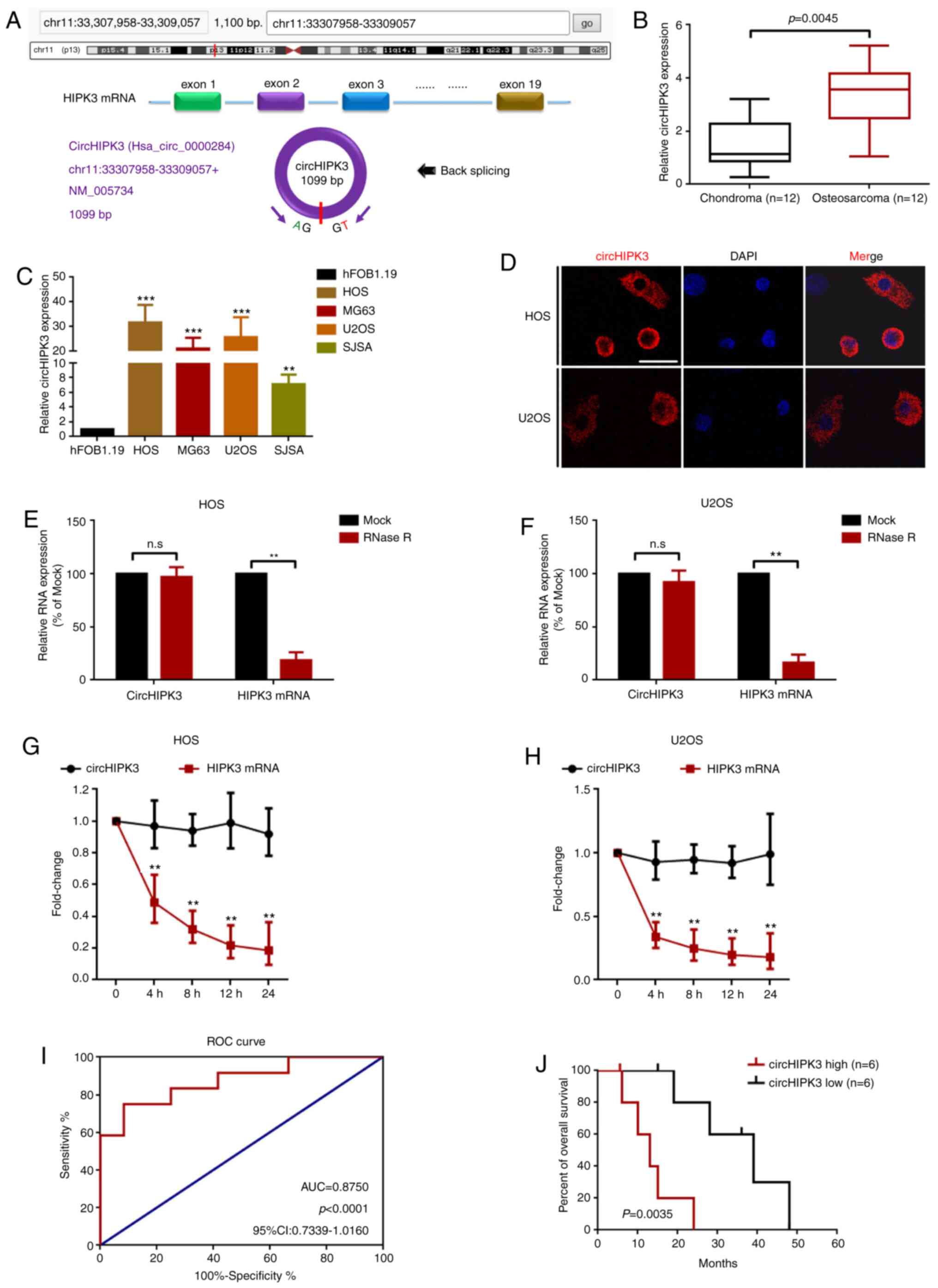 | Figure 1.circHIPK3 is relatively highly
expressed in OS cells, and predominantly localized in the
cytoplasm. (A) circHIPK3 was derived from exon 2 of linear HIPK3
mRNA as illustrated. (B) circHIPK3 expression in 12 pairs of OS and
chondroma tissue samples was determined using RT-qPCR. (C)
circHIPK3 expression in four OS cell lines and in the normal
osteoblast hFOB1.19 cell line was measured using RT-qPCR.
**P<0.01 and ***P<0.001 vs. hFOB1.19. (D) circHIPK3 was
mainly located in the cytoplasm according to a fluorescence in
situ hybridization assay. Magnification, ×400; scale bar, 20
µm. An RNase R assay was performed to determine the stability of
circHIPK3 in (E) HOS and (F) U2OS cells. **P<0.01. An
actinomycin D assay was performed to check the stability of
circHIPK3 in (G) HOS and (H) U2OS cells. **P<0.01. (I) Clinical
values of circHIPK3 in OS were analyzed using an ROC curve (AUC,
0.8750; P<0.0001). (J) High expression levels of circHIPK3 were
associated with shorter overall survival of patients with OS. n=6
for each group. P=0.0035. All data are presented as the mean ± SD
of three independent experiments. AUC, area under the curve; circ,
circular RNA; HIPK3, homeodomain interacting protein kinase 3; n.s,
not significant (P>0.05); OS, osteosarcoma; ROC, receiver
operating characteristic; RT-qPCR, reverse
transcription-quantitative PCR. |
Knockdown of circHIPK3 suppresses
proliferation, migration and invasion in OS cells
In order to investigate the role of circHIPK3 in OS,
the present study performed loss-of-function assays. Two specific
siRNAs that targeted the junction sites were designed to knock down
circHIPK3 in OS cells. As demonstrated by RT-qPCR (Fig. 2A and B), sicircHIPK3#1 presented
higher silencing efficacy. Therefore, this was selected as the RNA
interference (RNAi) tool in the following RNAi experiments.
Subsequently, the present study used a CCK8 assay to determine the
proliferation ability of OS cells. As the data presented in
Fig. 2C and D show, knockdown of
circHIPK3 suppressed proliferation in HOS and U2OS cells. Finally,
the present study performed a Transwell migration/invasion assay
and a wound healing assay to detect the role of circHIPK3 in OS
cell metastasis. As shown in the representative images in Fig. 2E and F, knockdown of circHIPK3
inhibited OS cell metastasis.
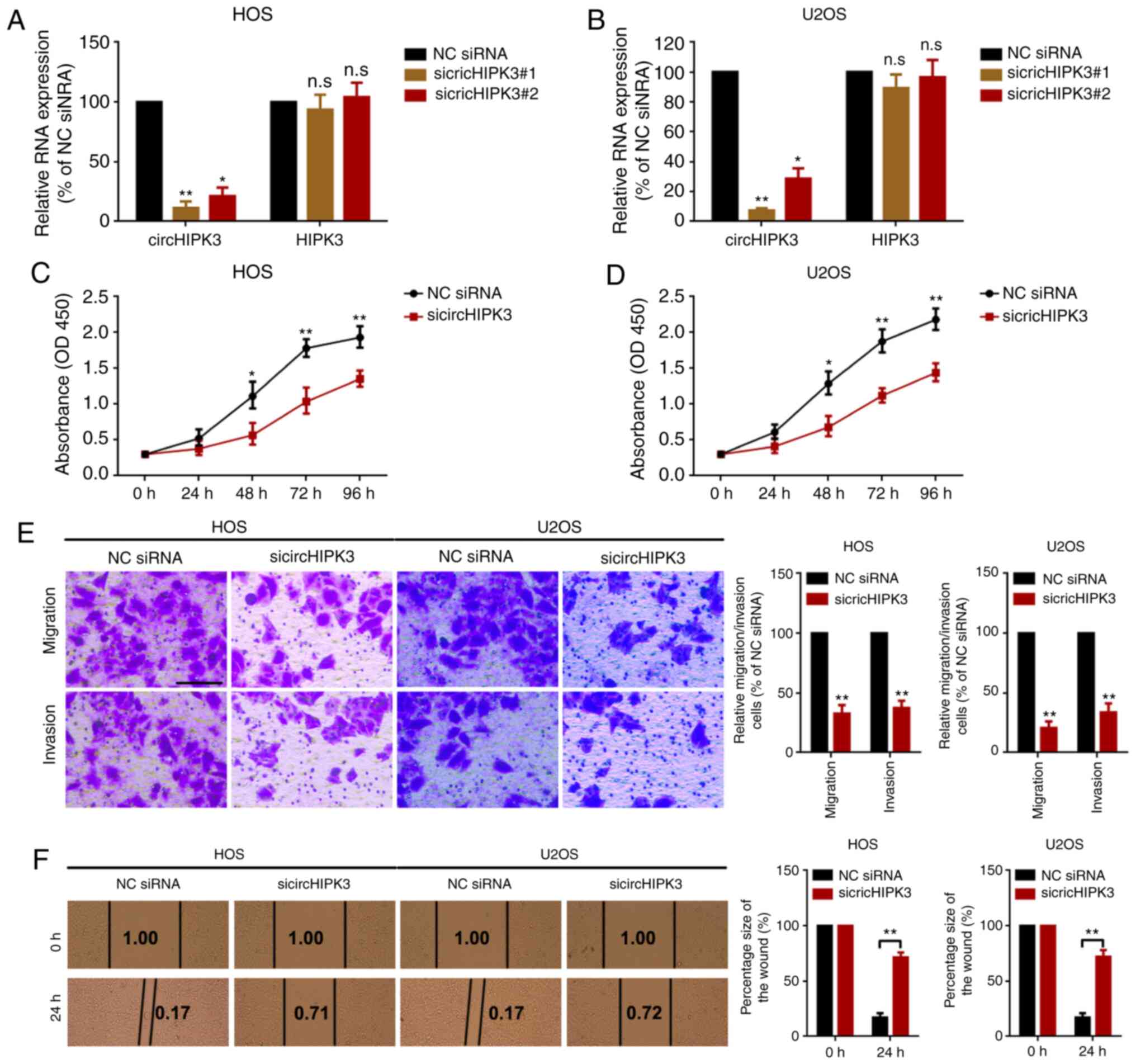 | Figure 2.Knockdown of circHIPK3 suppresses
proliferation, migration and invasion in OS cells. Expression
levels of circHIPK3 and HIPK3 mRNA in (A) HOS and (B) U2OS cells
after transfection of specific circHIPK3 siRNAs were measured by
reverse transcription-quantitative PCR. A Cell Counting Kit 8 assay
was performed to investigate the proliferation ability of (C) HOS
and (D) U2OS cells. (E) A Transwell assay was used to evaluate the
metastatic ability of HOS and U2OS cells. Magnification, ×20; scale
bar, 200 µm. (F) A wound healing assay was performed to determine
the migration ability of HOS and U2OS cells. Magnification, ×4;
scale bar, 500 µm. *P<0.05 and **P<0.01 vs. NC siRNA group.
All data are presented as the mean ± SD of three independent
experiments. circ, circular RNA; HIPK3, homeodomain interacting
protein kinase 3; NC, negative control; n.s, not significant
(P>0.05); OD, optical density; siRNA, small interfering RNA. |
circHIPK3 sponges miR-637 in OS
cells
Accumulating evidence has indicated that circRNAs
serve as miRNA sponges by harboring multiple miRNAs, and thereby
function as miRNA inhibitors (7,35). The
present study demonstrated that circHIPK3 was mainly located in the
cytoplasm (Fig. 1D). Therefore, it
was assumed that circHIPK2 may work via a similar mechanism in OS.
The present study screened the miRNAs that may interact with
circHIPK3 using the online tools circBank, RegRNA and circularRNA
interactome. As demonstrated by a Venn diagram (Fig. 3A), only miR-637 was identified in
all three databases. Therefore, the present study further
investigated the expression levels of miR-637 in OS. By analyzing
Gene Expression Omnibus (GEO) datasets GSE70415, it was revealed
that miR-637 was expressed at low levels in OS cells (Fig. 3B). Using another non-coding RNA
profiling array dataset (GSE28423), it was identified that miR-637
was expressed at low levels in 9 OS cell line compared with in 4
normal human bone samples (Fig.
3C). In the present study, it was revealed that miR-637 was
downregulated in OS cell lines compared with in hFOB1.19 cells
(Fig. 3D). Furthermore, using a
FISH assay, it was revealed that both miR-637 and circHIPK3 were
co-located in the cytoplasm (Fig.
3E). To further investigate the association between miR-637 and
circHIPK3, the present study performed Spearman's correlation
analysis. As shown in Fig. 3F,
circHIPK3 expression was negatively correlated with miR-637
expression. In addition, it was demonstrated that upregulation and
downregulation of miR-637 inversely regulated circHIPK3 expression
(Fig. 3G and H). Conversely,
miR-637 expression was inversely regulated by circHIPK3
upregulation and downregulation (Fig.
3I and J). These results indicated that miR-637 interacted with
circHIPK3 in a reciprocal suppressive manner. Finally, a luciferase
assay revealed that miR-637 targeted circHIPK3 via direct binding
(Fig. 3K-M).
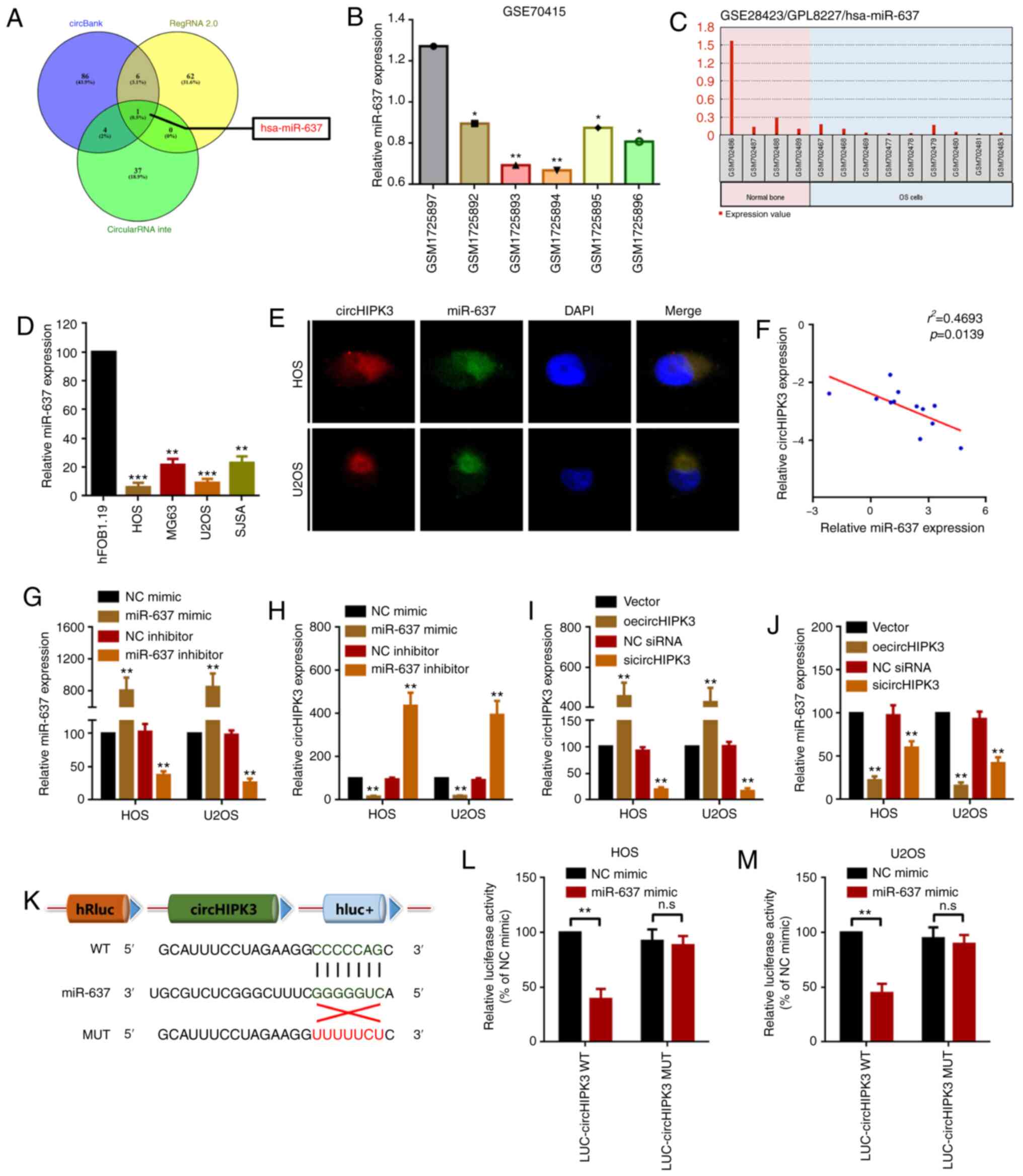 | Figure 3.circHIPK3 sponges miR-637 in OS
cells. (A) Potential miRNAs that may interact with circHIPK3 were
predicted by circBank (http://www.circbank.cn/), RegRNA 2.0 (http://regrna2.mbc.nctu.edu.tw/) and circularRNA
interactome (https://circinteractome.nia.nih.gov/). miR-637
expression in GEO datasets (B) GSE70415 and (C) GSE28423 was
analyzed using GEO2R. *P<0.05 and **P<0.01 vs. GSM1725897
group. (D) An RT-qPCR assay was used to determine the expression
levels of miR-637 in OS cell lines. **P<0.01 and ***P<0.001
vs. hFOB1.19 group. (E) circHIPK3 and miR-637 were colocalized in
the cytoplasm of HOS and U2OS cells as demonstrated by a
fluorescence in situ hybridization assay. Magnification,
×400; scale bar, 20 µm. (F) Spearman's correlation analysis
revealed that miR-637 expression was inversely correlated with
circHIPK3 expression. (G) miR-637 and (H) circHIPK3 expression
after different miR-637 interventions was investigated by RT-qPCR.
(I) circHIPK3 and (J) miR-637 expression after different circHIPK3
interventions was measured by RT-qPCR. (K) Diagram of constructed
luciferase reporter plasmids. miR-637 could directly target
circHIPK3 in (L) HOS and (M) U2OS cells, as demonstrated by a
luciferase assay. n.sP>0.05 and **P<0.01 vs. NC
mimic group. All data are presented as the mean ± SD of three
independent experiments. circ, circular RNA; GEO, Gene Expression
Omnibus; HIPK3, homeodomain interacting protein kinase 3; LUC,
luciferase; miR-637, microRNA-637; MUT, mutant; NC, negative
control; n.s, not significant (P>0.05); oe, overexpression; OS,
osteosarcoma; RT-qPCR, reverse transcription-quantitative PCR;
siRNA, small interfering RNA; WT, wild-type. |
miR-637 suppresses proliferation,
migration and invasion partially via targeting of HDAC4 in OS
cells
miRNAs are well-known to regulate their downstream
genes via direct targeting. Our previous study indicated that HDAC4
serves as an oncogene in OS (16).
The present study hypothesized that HDAC4 was also a target of
miR-637. First, it was revealed that miR-637 had limited effects on
the mRNA expression levels of HDAC4 (Fig. 4A). Furthermore, miR-637 inversely
regulated HDAC4 protein expression (Fig. 4B), indicating that miR-637 regulated
HDAC4 at the post-transcriptional level. HDAC4-specific small
interfering RNAs (siHDAC4) and overexpression plasmids (oeHDAC4)
were used to determine the role of HDAC4 in miR-637-mediated
proliferation and motility changes. The silencing and
overexpression efficacy of siHDAC4 and oeHDAC4 were demonstrated
(Fig. S2). Secondly, the present
study demonstrated that upregulation of miR-637 suppressed OS cell
proliferation, migration and invasion, and the suppressive effect
was reversed by upregulation of HDAC4 (transfection of HDAC4
overexpression plasmid oeHDAC4; Fig.
4C). Conversely, knockdown of miR-637 promoted OS cell
proliferation, migration and invasion, and the effect was
attenuated by knockdown of HDAC4 (transfection with siHDAC4;
Fig. 4D). Subsequently, the present
study demonstrated that HDAC4 was a downstream target of miR-637
using a luciferase assay (Fig. 4E and
F). The aforementioned results indicated that miR-637
suppressed OS cell proliferation, migration and invasion partially
by directly targeting HDAC4.
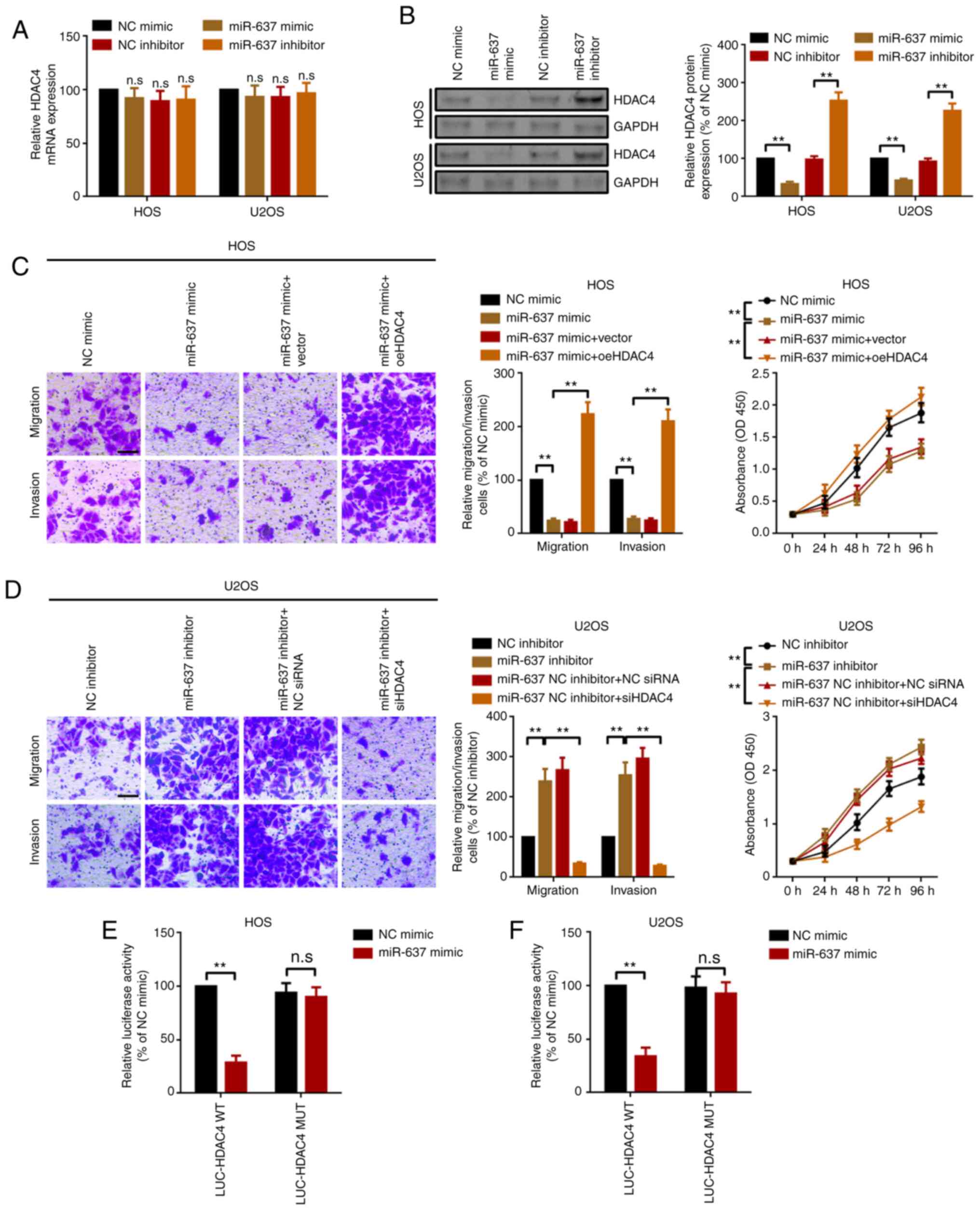 | Figure 4.miR-637 suppresses proliferation,
migration and invasion partially via HDAC4 modulation in
osteosarcoma cells. (A) HDAC4 mRNA expression in HOS and U2OS cells
was measured by reverse transcription-quantitative PCR. (B) HDAC4
protein expression was investigated by western blotting. Metastatic
and proliferation abilities of (C) HOS and (D) U2OS cells were
determined by Transwell and Cell Counting Kit 8 assays.
Magnification, ×20; scale bar, 200 µm. A luciferase assay was used
to demonstrate the targeted binding effect between miR-637 and
HDAC4 in (E) HOS and (F) U2OS cells. **P<0.01. All data are
presented as the mean ± SD of three independent experiments. HDAC4,
histone deacetylase 4; LUC, luciferase; miR-637, microRNA-637; MUT,
mutant; NC, negative control; n.s, not significant (P>0.05); OD,
optical density; oe, overexpression; siRNA, small interfering RNA;
WT, wild-type. |
circHIPK3 promotes proliferation,
migration and invasion by modulating the miR-637/HDAC4 axis in
OS
The present study investigated the association among
circHIPK3, miR-637 and HDAC4. Transwell and CCK8 assays were
performed to investigate the role of miR-637 in circHIPK3-induced
proliferation, migration and invasion. As shown in Fig. 5A, overexpression of circHIPK3
promoted OS cell proliferation, migration and invasion. The
facilitative effect was reversed by miR-637 mimics (upregulation of
miR-637). Conversely, inhibition of circHIPK3 suppressed OS cell
proliferation, migration and invasion, and the suppressive effect
was attenuated by inhibition of miR-637 (transfection of miR-637
inhibitor; Fig. 5B). In addition,
similar to the proliferation and metastatic assays, overexpression
of circHIPK3 promoted HDAC4 protein expression, and the
facilitative effect was attenuated by miR-637 (Fig. 5C). On the other hand, suppression of
cirHIPK3 inhibited HDAC4 expression, while the suppressive effect
was reversed by knockdown of miR-637 (Fig. 5D). In summary, all findings
suggested that circHIPK3 promoted proliferation, migration and
invasion via modulation of the miR-637/HDAC4 axis in OS.
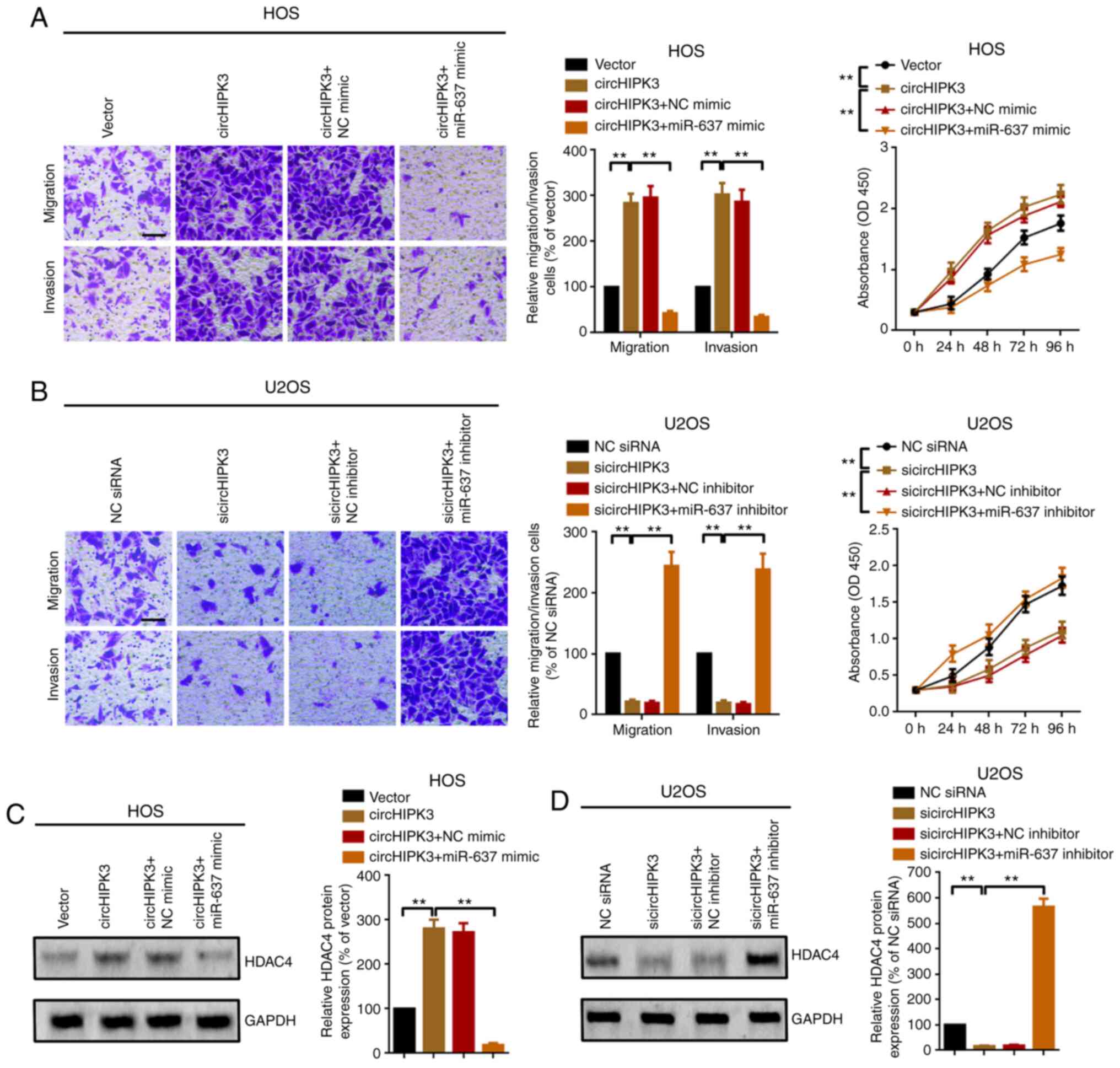 | Figure 5.circHIPK3 promotes proliferation and
metastasis via modulation of the miR-637/HDAC4 axis in
osteosarcoma. Metastatic and proliferation abilities of (A) HOS and
(B) U2OS cells were investigated using Transwell and Cell Counting
Kit 8 assays. Magnification, ×20; scale bar, 200 µm. (C) HDAC4
protein expression after different circHIPK3 and miR-637
interventions was investigated by western blotting. **P<0.01.
(D) Knockdown of circHIPK3 suppressed HDAC4 protein expression,
while the suppressive effect was abolished by a further
downregulation of miR-637, as determined by a western blotting
assay. **P<0.01. All data are presented as the mean ± SD of
three independent experiments. circ, circular RNA; HDAC4, histone
deacetylase 4; HIPK3, homeodomain interacting protein kinase 3;
miR-637, microRNA-637; NC, negative control; OD, optical density;
n.s, not significant (P>0.05); siRNA, small interfering RNA. |
Discussion
circRNAs are a type of RNA molecule with closed
single-stranded structure. The formation of circRNAs primarily
occurs through back-splicing of precursor mRNA or skipping events
of thousands of genes in eukaryotes as covalently closed continuous
loops (36). circRNAs function via
multiple mechanisms, including modulation of transcription, miRNA
sponging and associating with protein binding (37). circHIPK3 was first identified by
ribominus RNA sequencing data from six human normal tissues and
seven types of human cancer (9).
circHIPK3 serves as an oncogene in various types of cancer,
including gastric cancer, CRC, lung cancer and prostate cancer
(38–41). In the present study, the expression
and function of circHIPK3 in OS were determined. Using RT-qPCR, it
was revealed that circHIPK3 expression was upregulated in OS tissue
samples and in OS cells. In addition, the present study
demonstrated that high levels of circHIPK3 were closely associated
with shorter overall survival of patients with OS. Previous studies
have demonstrated that circRNAs are abundantly enriched in the
cytoplasm and serve as miRNAs sponges (42–44).
Therefore, the present study performed a localization FISH assay to
demonstrate the distribution of circHIPK3. A specific probe was
designed to target circHIPK3, and it was revealed that circHIPK3
was prominently located in OS cell cytoplasm. Functionally, the
present study revealed that circHIPK3 promoted OS cell
proliferation, migration and invasion using loss-of-function CCK8
and Transwell assays.
circRNAs serve as miRNAs sponges by acting as
competing endogenous RNAs (42).
Since circHIPK3 was primarily located in the OS cell cytoplasm, the
present study inferred that circHIPK3 may interact with certain
miRNAs. miR-637 has been reported as a tumor suppressor in several
types of cancer (45,46). In a melanoma-associated study, Zhang
et al (47) revealed that
overexpression of miR-637 suppresses proliferation and cell cycle
G1-S transition, and induces apoptosis in melanoma
cells. Du et al (45)
reported that miR-637 is expressed at low levels in hepatoma, and
miR-637 inhibits proliferation and invasion of hepatoma cells by
targeted degradation of AKT1. By analyzing GEO datasets GSE70415
and GSE28423 and RT-qPCR, the present study demonstrated that
miR-637 expression was downregulated in OS. Furthermore, it was
revealed that both miR-637 and circHIPK3 were co-localized in the
cytoplasm, and they interacted with each other in a reciprocal
suppressive manner. These results demonstrated that miR-637 and
circHIPK3 may interact on a biological basis. Furthermore, the
results of the luciferase assay indicated that miR-637 could bind
to circHIPK3 directly. Finally, the present study investigated the
association between miR-637 and HDAC4, a well-known oncogene in
various malignancies, including OS (15). In our previous study (16), the oncogenic role of HDAC4 in OS was
observed. In the present study, it was revealed that HDAC4 was
inversely regulated by miR-637 at the post-transcriptional level.
In addition, the present study demonstrated that miR-637 targeted
HDAC4 via direct binding. The functional Transwell and CCK8 assays
suggested that miR-637 inhibited OS cell proliferation, migration
and invasion partially via the HDAC4 signaling pathway.
In summary, the results of the present study
suggested that circHIPK3 exerted a regulatory function by sponging
miR-637 and promoting OS cell proliferation, migration and invasion
partially via the HDAC4 signaling pathway. circHIPK3/miR-637/HDAC4
signaling may be a novel target in the treatment of OS.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Key R&D
Program of Liaoning Province (grant no. 20180550251).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GW conceived the experiments. YW, BL, MH and YS
performed the experiments. ST analyzed the data. GW wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study and the associated experimental
protocols (both human and animal experiments) were performed in
compliance with ethical guidelines and approved by the Institute
Research Medical Ethics Committee of the Shengjing Hospital of
China Medical University, Shenyang, China (approval no.
2020PS051K). All chondroma and OS tissues were also used in
accordance with the Declaration of Helsinki. Written informed
consent was provided by each patient before the study. All patients
agreed that the data from their samples could be used for
experimental studies and paper presentations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sweetnam R: Osteosarcoma. Ann R Coll Surg
Engl. 44:38–58. 1969.
|
|
2
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar
|
|
3
|
Berner K, Johannesen TB, Berner A,
Haugland HK, Bjerkehagen B, Bøhler PJ and Bruland ØS: Time-trends
on incidence and survival in a nationwide and unselected cohort of
patients with skeletal osteosarcoma. Acta Oncol. 54:25–33. 2015.
View Article : Google Scholar
|
|
4
|
Friebele JC, Peck J, Pan X, Abdel-Rasoul M
and Mayerson JL: Osteosarcoma: A meta-analysis and review of the
literature. Am J Orthop (Belle Mead NJ). 44:547–553. 2015.
|
|
5
|
Bishop MW, Janeway KA and Gorlick R:
Future directions in the treatment of osteosarcoma. Curr Opin
Pediatr. 28:26–33. 2016. View Article : Google Scholar
|
|
6
|
Zamborsky R, Kokavec M, Harsanyi S and
Danisovic L: Identification of prognostic and predictive
osteosarcoma biomarkers. Med Sci (Basel). 7:282019.
|
|
7
|
Patop IL and Kadener S: circRNAs in
cancer. Curr Opin Genet Dev. 48:121–127. 2018. View Article : Google Scholar
|
|
8
|
Kristensen LS, Hansen TB, Venø MT and
Kjems J: Circular RNAs in cancer: Opportunities and challenges in
the field. Oncogene. 37:555–565. 2018. View Article : Google Scholar
|
|
9
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar
|
|
10
|
Liu N, Zhang J, Zhang LY and Wang L:
CircHIPK3 is upregulated and predicts a poor prognosis in
epithelial ovarian cancer. Eur Rev Med Pharmacol Sci. 22:3713–3718.
2018.
|
|
11
|
Ke Z, Xie F, Zheng C and Chen D: CircHIPK3
promotes proliferation and invasion in nasopharyngeal carcinoma by
abrogating miR-4288-induced ELF3 inhibition. J Cell Physiol.
234:1699–1706. 2019. View Article : Google Scholar
|
|
12
|
Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang
C, Liu D, Wang M, Wang L, Zeng F and Jiang G: CircHIPK3 sponges
miR-558 to suppress heparanase expression in bladder cancer cells.
EMBO Rep. 18:1646–1659. 2017. View Article : Google Scholar
|
|
13
|
Yu H, Chen Y and Jiang P: Circular RNA
HIPK3 exerts oncogenic properties through suppression of miR-124 in
lung cancer. Biochem Biophys Res Commun. 506:455–462. 2018.
View Article : Google Scholar
|
|
14
|
Chen G, Shi Y, Liu M and Sun J: circHIPK3
regulates cell proliferation and migration by sponging miR-124 and
regulating AQP3 expression in hepatocellular carcinoma. Cell Death
Dis. 9:1752018. View Article : Google Scholar
|
|
15
|
Wang Z, Qin G and Zhao TC: HDAC4:
Mechanism of regulation and biological functions. Epigenomics.
6:139–150. 2014. View Article : Google Scholar
|
|
16
|
Sun Y and Qin B: Long noncoding RNA MALAT1
regulates HDAC4-mediated proliferation and apoptosis via decoying
of miR-140-5p in osteosarcoma cells. Cancer Medicine. 7:4584–4597.
2018. View Article : Google Scholar
|
|
17
|
Zeng LS, Yang XZ, Wen YF, Mail SJ, Wang
MH, Zhang MY, Zheng XFS and Wang HY: Overexpressed HDAC4 is
associated with poor survival and promotes tumor progression in
esophageal carcinoma. Aging (Albany NY). 8:1236–1249. 2016.
View Article : Google Scholar
|
|
18
|
Sissons HA: The WHO classification of bone
tumors. Recent Results Cancer Res. 104–108. 1976.
|
|
19
|
Kawano M, Tanaka K, Itonaga I, Ikeda S,
Iwasaki T and Tsumura H: microRNA-93 promotes cell proliferation
via targeting of PTEN in osteosarcoma cells. J Exp Clin Cancer Res.
34:762015. View Article : Google Scholar
|
|
20
|
Namløs HM, Meza-Zepeda LA, Barøy T,
Østensen IHG, Kresse SH, Kuijjer ML, Serra M, Bürger H,
Cleton-Jansen AM and Myklebost O: Modulation of the osteosarcoma
expression phenotype by microRNAs. PLoS One. 7:e480862012.
View Article : Google Scholar
|
|
21
|
Davis S and Meltzer PS: GEOquery: A bridge
between the gene expression omnibus (GEO) and bioconductor.
Bioinformatics. 23:1846–1847. 2007. View Article : Google Scholar
|
|
22
|
Zhang J, Ye Y, Chang DW, Lin SH, Huang M,
Tannir NM, Matin S, Karam JA, Wood CG, Chen ZN and Wu X: Global and
targeted miRNA expression profiling in clear cell renal cell
carcinoma tissues potentially links miR-155-5p and miR-210-3p to
both tumorigenesis and recurrence. Am J Pathol. 188:2487–2496.
2018. View Article : Google Scholar
|
|
23
|
Glažar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar
|
|
24
|
Chang TH, Huang HY, Hsu JB, Weng SL, Horng
JT and Huang HD: An enhanced computational platform for
investigating the roles of regulatory RNA and for identifying
functional RNA motifs. BMC Bioinformatics. 14 (Suppl 2):S42013.
View Article : Google Scholar
|
|
25
|
Dudekula DB, Panda AC, Grammatikakis I, De
S, Abdelmohsen K and Gorospe M: CircInteractome: A web tool for
exploring circular RNAs and their interacting proteins and
microRNAs. RNA Biol. 13:34–42. 2016. View Article : Google Scholar
|
|
26
|
Zeng K, Chen X, Xu M, Liu X, Hu X, Xu T,
Sun H, Pan Y, He B and Wang S: CircHIPK3 promotes colorectal cancer
growth and metastasis by sponging miR-7. Cell Death Dis. 9:4172018.
View Article : Google Scholar
|
|
27
|
Cheng Z, Yu C, Cui S, Wang H, Jin H, Wang
C, Li B, Qin M, Yang C, He J, et al: circTP63 functions as a ceRNA
to promote lung squamous cell carcinoma progression by upregulating
FOXM1. Nat Commun. 10:32002019. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Zhong S, Wang J, Zhang Q, Xu H and Feng J:
CircPrimer: A software for annotating circRNAs and determining the
specificity of circRNA primers. BMC Bioinformatics. 19:2922018.
View Article : Google Scholar
|
|
30
|
Wang Y, Zeng X, Wang N, Zhao W, Zhang X,
Teng S, Zhang Y and Lu Z: Long noncoding RNA DANCR, working as a
competitive endogenous RNA, promotes ROCK1-mediated proliferation
and metastasis via decoying of miR-335-5p and miR-1972 in
osteosarcoma. Mol Cancer. 17:892018. View Article : Google Scholar
|
|
31
|
Wang Y, Lu Z, Wang N, Feng J, Zhang J,
Luan L, Zhao W and Zeng X: Long noncoding RNA DANCR promotes
colorectal cancer proliferation and metastasis via miR-577
sponging. Exp Mol Med. 50:1–17. 2018. View Article : Google Scholar
|
|
32
|
Wang Y, Zhang Y, Yang T, Zhao W, Wang N,
Li P, Zeng X and Zhang W: Long non-coding RNA MALAT1 for promoting
metastasis and proliferation by acting as a ceRNA of miR-144-3p in
osteosarcoma cells. Oncotarget. 8:59417–59434. 2017. View Article : Google Scholar
|
|
33
|
Yan Y, Wang Z and Qin B: A novel long
noncoding RNA, LINC00483 promotes proliferation and metastasis via
modulating of FMNL2 in CRC. Biochem Biophys Res Commun.
509:441–447. 2019. View Article : Google Scholar
|
|
34
|
Li X, Yang L and Chen LL: The biogenesis,
functions, and challenges of circular RNAs. Mol Cell. 71:428–442.
2018. View Article : Google Scholar
|
|
35
|
Arnaiz E, Sole C, Manterola L,
Iparraguirre L, Otaegui D and Lawrie CH: CircRNAs and cancer:
Biomarkers and master regulators. Semin Cancer Biol. 58:90–99.
2019. View Article : Google Scholar
|
|
36
|
Chen LL: The biogenesis and emerging roles
of circular RNAs. Nat Rev Mol Cell Biol. 17:205–211. 2016.
View Article : Google Scholar
|
|
37
|
Bach DH, Lee SK and Sood AK: Circular RNAs
in cancer. Mol Ther Nucleic Acids. 16:118–129. 2019. View Article : Google Scholar
|
|
38
|
Cai C, Zhi Y, Wang K, Zhang P, Ji Z, Xie C
and Sun F: CircHIPK3 overexpression accelerates the proliferation
and invasion of prostate cancer cells through regulating
miRNA-338-3p. OncoTargets Ther. 12:3363–3372. 2019. View Article : Google Scholar
|
|
39
|
Zhang Y, Li C, Liu X, Wang Y, Zhao R, Yang
Y, Zheng X, Zhang Y and Zhang X: circHIPK3 promotes
oxaliplatin-resistance in colorectal cancer through autophagy by
sponging miR-637. EBioMedicine. 48:277–288. 2019. View Article : Google Scholar
|
|
40
|
Chen X, Mao R, Su W, Yang X, Geng Q, Guo
C, Wang Z, Wang J, Kresty LA, Beer DG, et al: Circular RNA
circHIPK3 modulates autophagy via MIR124-3p-STAT3-PRKAA/AMPKα
signaling in STK11 mutant lung cancer. Autophagy. 16:659–671. 2020.
View Article : Google Scholar
|
|
41
|
Liu WG and Xu Q: Upregulation of circHIPK3
promotes the progression of gastric cancer via Wnt/β-catenin
pathway and indicates a poor prognosis. Eur Rev Med Pharmacol Sci.
23:7905–7912. 2019.
|
|
42
|
Zhong Y, Du Y, Yang X, Mo Y, Fan C, Xiong
F, Ren D, Ye X, Li C, Wang Y, et al: Circular RNAs function as
ceRNAs to regulate and control human cancer progression. Mol
Cancer. 17:792018. View Article : Google Scholar
|
|
43
|
Cui X, Wang J, Guo Z, Li M, Li M, Liu S,
Liu H, Li W, Yin X, Tao J and Xu W: Emerging function and potential
diagnostic value of circular RNAs in cancer. Mol Cancer.
17:1232018. View Article : Google Scholar
|
|
44
|
Han TS, Hur K, Cho HS and Ban HS:
Epigenetic associations between lncRNA/circRNA and miRNA in
hepatocellular carcinoma. Cancers (Basel). 12:E26222020. View Article : Google Scholar
|
|
45
|
Du YM and Wang YB: MiR-637 inhibits
proliferation and invasion of hepatoma cells by targeted
degradation of AKT1. Eur Rev Med Pharmacol Sci. 23:567–575.
2019.
|
|
46
|
Li JX, Ding XM, Han S, Wang K, Jiao CY and
Li XC: mir-637 inhibits the proliferation of cholangiocarcinoma
cell QBC939 through interfering CTSB expression. Eur Rev Med
Pharmacol Sci. 22:1265–1276. 2018.
|
|
47
|
Zhang J, Liu WL, Zhang L, Ge R, He F, Gao
TY, Tian Q, Mu X, Chen LH, Chen W and Li X: MiR-637 suppresses
melanoma progression through directly targeting P-REX2a and
inhibiting PTEN/AKT signaling pathway. Cell Mol Biol
(Noisy-le-Grand). 64:50–57. 2018. View Article : Google Scholar
|














