Introduction
Hepatocellular carcinoma (HCC) is listed as one of
the top 10 malignant tumors by the World Health Organization
(1). With an increasing morbidity
rate, HCC contributes to ~600,000 deaths annually worldwide
(2). Despite the significant
progress in the diagnosis and surgical treatment of the disease,
patients with HCC have a low overall survival rate, with dismal
prognosis (2,3). Therefore, an improved understanding
and further determination of the regulatory mechanisms underlying
HCC initiation and progression are required to identify potential
diagnostic markers for the treatment of patients with HCC.
According to the results of genomic and
transcriptomic sequencing, a large proportion of the genome is
transcribed into non-coding RNAs (ncRNAs), while only a small
percentage of the human genome is transcribed into protein-coding
mRNAs (4,5). Long ncRNAs (lncRNAs) are members of
the ncRNA family, which are characterized by being >200
nucleotides in length (6).
Increasing evidence has identified that lncRNAs are frequently
aberrantly expressed in numerous types of disease, particularly in
several types of cancer (7,8). Despite lacking the ability to encode
proteins, lncRNAs are capable of regulating gene expression via
several different mechanisms, including mRNA transportation, and
both transcriptional and post-transcriptional modulation (9–11).
Notably, emerging studies have proposed a competitive endogenous
RNA (ceRNA) mechanism, by which lncRNAs sponge microRNAs
(miRNAs/miRs) to modulate gene expression at a post-transcriptional
level (12,13). For instance, the lncRNA LOXL1-AS1
was discovered to upregulate USF1 expression levels by sponging
miR-708-5p in gastric carcinoma (14). In addition, lncRNA NONHSAT101069 was
found to promote breast cancer by downregulating the expression
levels of miR-129-5p and upregulating the expression levels of
Twist1 (15). Nevertheless, the
mechanism of action of ST8SIA6-AS1 in HCC remains poorly
understood.
miRNAs are another category of ncRNAs, which are
short, single-stranded ncRNAs of ~22 nucleotides in length
(16). miRNAs bind with the 3′
untranslated region (3′UTR) of mRNAs to degrade mRNA and/or inhibit
the translation of target genes, which has been identified to
regulate tumor occurrence and development (16,17).
For example, miR-98-5p was found to regulate the cell cycle via
targeting CDC25A in osteosarcoma (18), while the overexpression of
miR-367-3p was found to inhibit cell proliferation and invasion
through the Wnt/β-catenin signaling pathway by targeting SPAG5 in
cervical cancer (19). However,
although miR-338 has been reported to regulate HCC progression, the
interaction between ST8SIA6-AS1 and miR-338 remains to be fully
elucidated in HCC.
The present study aimed to investigate the function
and mechanism of action of ST8SIA6-AS1 in HCC. The results revealed
that ST8SIA6-AS1 promoted the migration and invasion of
hypoxia-induced HCC cells via the miR-338/MEPCE axis, providing a
potential diagnostic or therapeutic marker for the treatment of
HCC.
Materials and methods
Patient studies
HCC tissues (n=22) and adjacent normal tissues
(n=22) were obtained from HCC patients with a mean age of 58 years
(range, 32–76 years) between September 2015 and November 2017 at
The Affiliated Hospital of Xuzhou Medical University (Xuzhou,
China); all tissue specimens were maintained at −80°C. The patients
had not received any other anticancer treatment before surgery. The
study protocol was approved by the Ethics Committee of the
Affiliated Hospital of Xuzhou Medical University and written
informed consent was obtained from all patients.
Cell culture and treatment
HCC cell lines (HCCLM6, Hep3B and Huh-7) and a human
normal liver cell line (THLE-3) were purchased from The Cell Bank
of Type Culture Collection of the Chinese Academy of Sciences. All
cells were cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.), supplemented with 10% FBS and 1% penicillin/streptomycin
stock solution (Sigma-Aldrich; Merck KGaA). The cell lines were
maintained in a humidified atmosphere containing 5% CO2
at 37°C. Oxygen concentrations were maintained at 1% to induce
hypoxia and hypoxic conditions were established using a humidified
variable aerobic workstation with 5% CO2 at 37°C.
Cell transfection
The miR-338 mimic and negative control (NC) mimic,
and the miR-338 inhibitor and NC inhibitor, were used to
overexpress and inhibit miR-338, respectively. Short hairpin RNA
(shRNA/sh) targeting ST8SIA6-AS1 (sh-ST8SIA6-AS1#1 and
sh-ST8SIA6-AS1#2) and the NC (sh-NC) were used to knock down the
expression levels of ST8SIA6-AS1. Full length methylphosphate
capping enzyme (MEPCE) was subcloned into the pcDNA3.1 vector to
overexpress MEPCE; an empty pcDNA3.1 vector was used as the
control. All the plasmids were purchased from Shanghai GenePharma
Co., Ltd. and transfected into Hep3B or Huh-7 cells using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from HCC tissues or cell
lines using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). An miRNeasy Mini kit (Qiagen GmbH) and the
TaqMan® MicroRNA Reverse Transcription kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) were used to isolate
miRNAs and reverse transcribe miRNA into cDNA, respectively. For
the quantification of mRNAs, a Prime Script™ RT reagent kit (Takara
Bio, Inc.) was used for RT. qPCR was subsequently performed using a
TaqMan® Universal PCR Master mix II (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The relative expression levels of
RNAs (normalized to GAPDH or U6) were quantified using the
2−ΔΔCq method.
Nuclear-cytoplasmic fractionation
The distribution of ST8SIA6-AS1 in the cytoplasm or
nucleus of HCC cells was determined using a PARIS kit (Thermo
Fisher Scientific, Inc.). Briefly, Hep3B or Huh-7 cells were
harvested and lysed on ice, and the supernatant was collected
following centrifugation. The extracted RNAs were analyzed using
RT-qPCR, and GAPDH and U6 were used as the cytoplasmic and nucleic
controls, respectively.
Wound healing assay
A total of 8×104 transfected Hep3B or
Huh-7 cells/ml were cultured in RPMI-1640 medium, supplemented with
10% FBS, until 100% confluence. A sterile pipette tip was used to
generate single-line scratches and the cell monolayer was
subsequently washed twice with PBS. The wound closure was imaged at
0 and 24 h using a light microscope (magnification, ×200).
Transwell Matrigel assay
A total of 1×105 Hep3B or Huh-7 cells
suspended in 500 µl serum-free medium were plated into the upper
champers of Transwell plates (Corning Inc.). The upper chambers
were precoated with Matrigel. Medium supplemented with 5% FBS was
plated into the lower chambers. Following incubation for 24 h, the
invasive Hep3B or Huh-7 cells were fixed with methanol and stained
with 0.1% crystal violet for 20 min at room temperature. The
invasive cells were visualized under a light microscope
(magnification, ×200; Olympus Corporation).
Dual-luciferase reporter assay
Full-length MEPCE 3′UTR and ST8SIA6-AS1 were
amplified and cloned into pmirGLO vectors (Promega Corp.) to
construct the pmirGLO-MEPCE-WT reporter gene plasmid
(5′-CAGCAAGGCUGGCUGGUGCUGGA-3′) and pmirGLO-ST8SIA6-AS1-WT reporter
gene plasmid (5′-GAACAGAAUCGCUAAUAUGCUGG-3′), respectively.
Mutations within the miR-338 binding site were created using the
QuikChange II Site-Directed Mutagenesis kit (Stratagene; Agilent
Technologies, Inc.), to generate the pmirGLO-MEPCE-MUT
(5′-CAGCAAGGCUGGCUGGCAACAAC-3′) reporter gene plasmid and
pmirGLO-ST8SIA6-AS1-MUT (5′-GAACAGAAUCGCUAAACCAACAA-3′) reporter
gene plasmid. Hep3B and Huh-7 cells were co-transfected with
miR-338 mimics or NC mimics and ST8SIA6-AS1-WT (or MEPCE-WT) or
ST8SIA6-AS1- Mut (or MEPCE-Mut) using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.). Following 48 h
of transfection, the relative luciferase activity was analyzed
using a Dual-Luciferase Reporter assay system. Firefly luciferase
activity was normalized to Renilla luciferase activity
(Promega Corp.).
RNA immunoprecipitation (RIP)
assay
A RIP assay was performed using the Magna
RNA-binding protein immunoprecipitation kit (EMD Millipore).
Briefly, the cell lysate (Hep3B or Huh-7 cells) was incubated in
RIP buffer containing magnetic beads conjugated to human anti-Ago2
antibody. IgG served as the endogenous control. The
immunoprecipitated RNA was isolated, purified and analyzed using
RT-qPCR.
Statistical analysis
SPSS 20.0 software (IBM Corp.) was used for
statistical analysis and data are presented as the mean ± SD of
three independent repeats. Clinicopathological characteristics were
evaluated using the χ2 test. Comparison between HCC and
adjacent normal tissues was performed using a paired Student's
t-test, while comparisons between the experimental and control
groups was performed using an unpaired Student's t-test. Comparison
among multiple groups was performed using one-way ANOVA, followed
by Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
ST8SIA6-AS1 expression levels are
upregulated in HCC and are associated with a poor prognosis
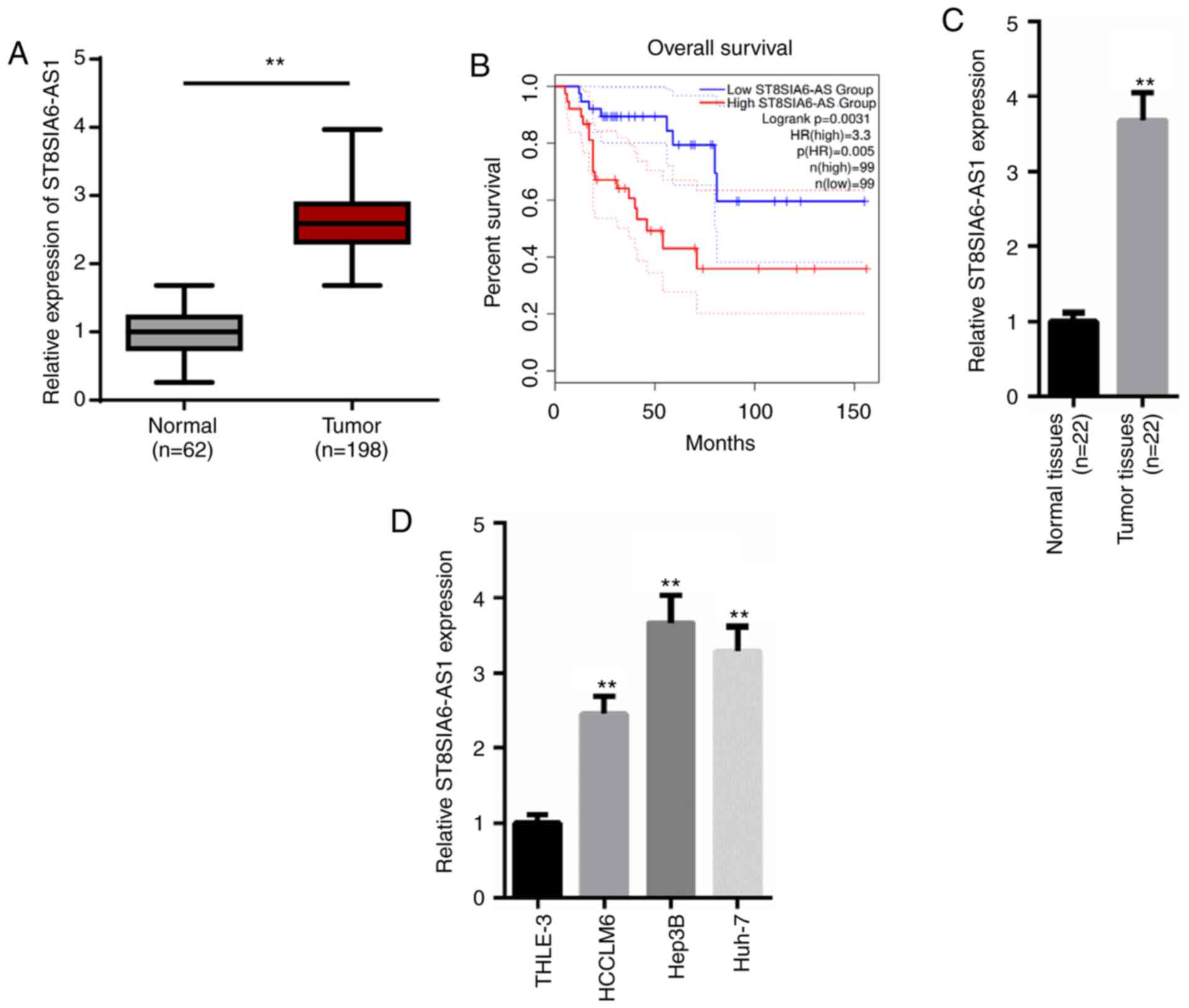
According to the TCGA database (https://tcga-data.nci.nih.gov/tcga/), ST8SIA6-AS1
expression levels were significantly upregulated in liver
hepatocellular carcinoma (LIHC) tissues (n=198) compared with
normal tissues (n=62; Fig. 1A).
Additionally, patients with HCC with higher levels of ST8SIA6-AS1
had a lower survival rate compared with patients with low
expression levels of ST8SIA6-AS1 (Fig.
1B). Similarly, RT-qPCR analysis revealed that expression
levels of ST8SIA6-AS1 were significantly upregulated in HCC tissues
(n=22) or cell lines (HCCLM6, Hep3B and Huh-7) compared with the
adjacent normal tissues (n=22) or the human normal liver cell line
(THLE-3; Fig. 1C and D). Moreover,
the clinical data indicated that the ST8SIA6-AS1 expression levels
were positively correlated with the histological grade, TNM stage
and vein invasion (Table I).
Altogether, these results suggest that ST8SIA6-AS1 expression
levels may be upregulated in HCC tissues and cell lines, and that
the upregulation of ST8SIA6-AS1 expression levels may predict the
poor prognosis of HCC.
 | Table I.Association between ST8SIA6-AS1
expression and clinicopathologic features in patients with HCC. |
Table I.
Association between ST8SIA6-AS1
expression and clinicopathologic features in patients with HCC.
|
|
|
| ST8SIA6-AS1 |
|
|---|
|
|
|
|
|
|
|---|
| Parameters | Group | Total | Low | High | P-value |
|---|
| Sex | Male | 13 | 6 | 7 | 0.496 |
|
| Female | 9 | 5 | 4 |
|
| Age (years) | <50 | 10 | 4 | 6 | 0.272 |
|
| ≥50 | 12 | 7 | 5 |
|
| Tumor size (cm) | <5 | 8 | 4 | 4 | 0.807 |
|
| ≥5 | 14 | 8 | 6 |
|
| Histological
grade | Low | 7 | 5 | 2 | <0.001 |
|
| High | 15 | 4 | 11 |
|
| TNM stage | I–II | 8 | 5 | 3 | 0.006 |
|
| III–IV | 14 | 5 | 9 |
|
| Vein invasion | Absence | 7 | 6 | 1 | <0.001 |
|
| Presence | 15 | 5 | 10 |
|
| Cirrhosis | Negative | 16 | 8 | 8 | 0.805 |
|
| Positive | 6 | 3 | 3 |
|
| Hepatitis B | Negative | 7 | 4 | 3 | 0.818 |
|
| Positive | 15 | 7 | 8 |
|
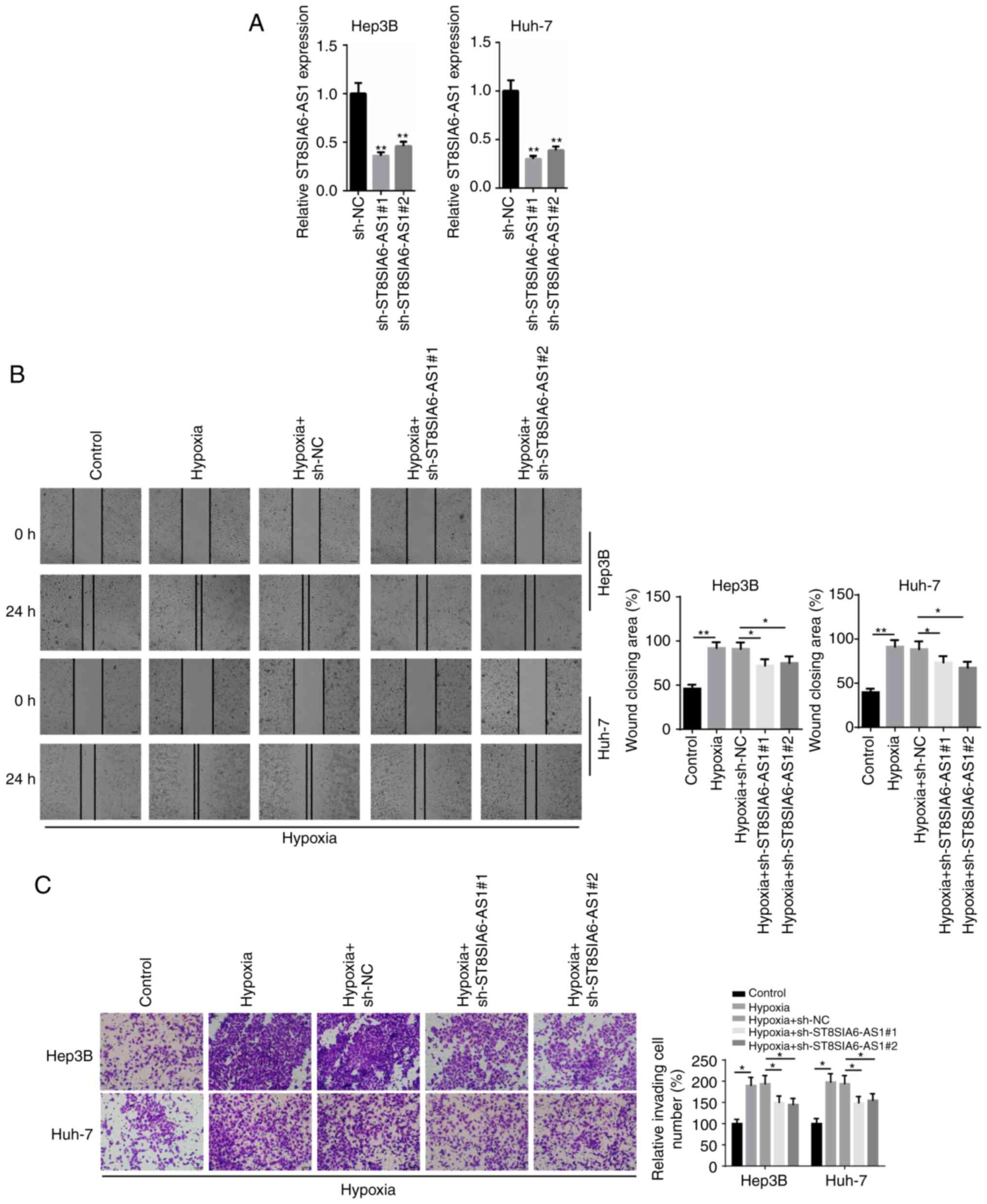
Knockdown of ST8SIA6-AS1 inhibits
hypoxia-induced HCC cell progression
To investigate the biological role of ST8SIA6-AS1 in
HCC, loss-of-function assays were performed. The expression levels
of ST8SIA6-AS1 were markedly downregulated following the
transfection of sh-ST8SIA6-AS1#1 or sh-ST8SIA6-AS1#2 into Hep3B and
Huh-7 cells (Fig. 2A).
Subsequently, a wound healing assay demonstrated that the migratory
ability of HCC cells was significantly increased following hypoxia
induction compared with the control group, while the migratory rate
was significantly hindered following the transfection with
sh-ST8SIA6-AS1#1 or sh-ST8SIA6-AS1#2 (Fig. 2B). Similarly, the Transwell Matrigel
assay revealed that the invasive capacity of HCC cells was enhanced
by hypoxia treatment, while the invasive capacity was significantly
inhibited by the knockdown of ST8SIA6-AS1 (Fig. 2C). Together, these results indicated
that the genetic knockdown of ST8SIA6-AS1 may inhibit the migration
and invasion of hypoxia-induced HCC cells.
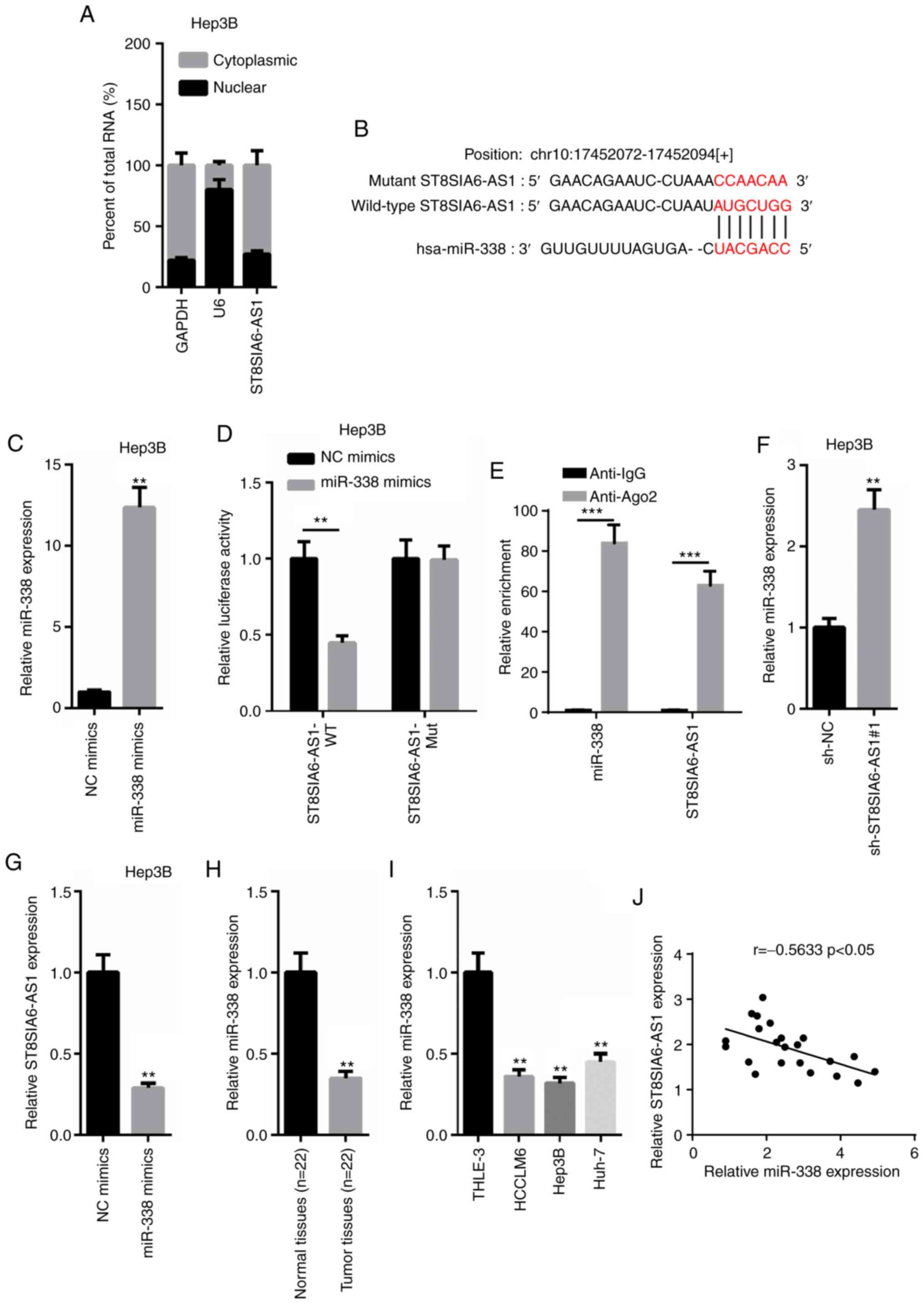
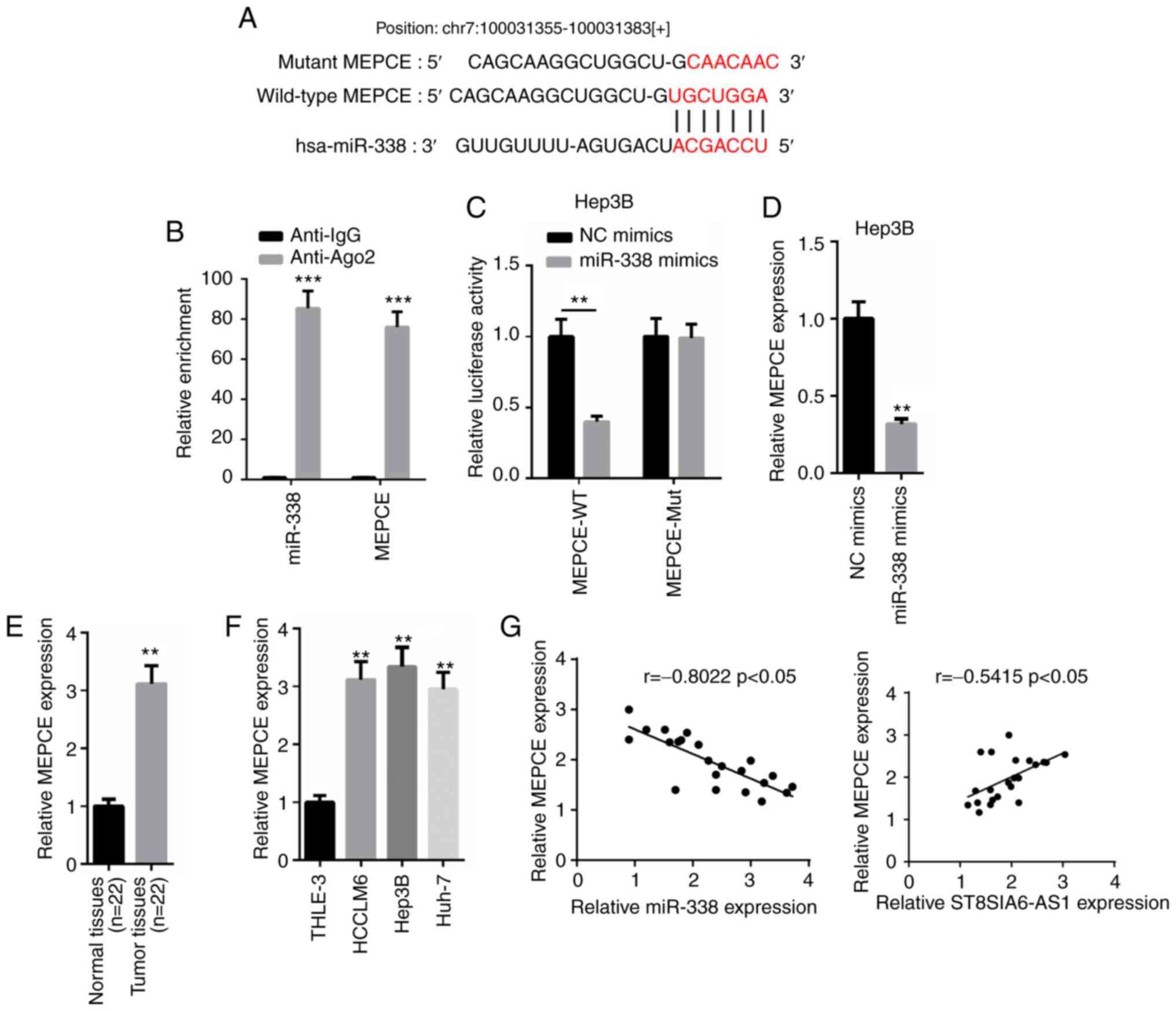
ST8SIA6-AS1 directly binds with
miR-338 to inhibit its expression levels
According to the results obtained from
nuclear-cytoplasmic fractionation, the majority of ST8SIA6-AS1 was
observed to be distributed in the cytoplasm of Hep3B cells
(Fig. 3A). Thus, it was
hypothesized that ST8SIA6-AS1 may form a ceRNA network in HCC.
miR-338 was predicted to contain a binding site on ST8SIA6-AS1
using starBase (http://starbase.sysu.edu.cn; Fig. 3B). Thus, the expression levels of
miR-338 were overexpressed by transfecting miR-338 mimics into
Hep3B cells (Fig. 3C). A
dual-luciferase reporter assay revealed that miR-338 overexpression
induced a marked decrease in pmirGLO-ST8SIA6-AS1-WT relative
luciferase activity in Hep3B cells, whereas no differences were
observed in pmirGLO-ST8SIA6-AS1-Mut-transfected cells (Fig. 3D). Subsequently, a RIP assay
revealed that both ST8SIA6-AS1 and miR-338 expression levels were
enriched in anti-Ago2 cells, suggesting that ST8SIA6-AS1 and
miR-338 may co-exist in RNA-induced silencing complexes (RISC;
Fig. 3E). In addition, ST8SIA6-AS1
knockdown significantly upregulated miR-338 expression levels,
while miR-338 overexpression significantly downregulated
ST8SIA6-AS1 expression levels (Fig. 3F
and G). Furthermore, miR-338 expression levels were revealed to
be downregulated in HCC tissues or cells compared with the adjacent
normal tissues or THLE-3 cells (Fig. 3H
and I). Moreover, the expression levels of ST8SIA6-AS1 and
miR-338 in HCC tissues were negatively correlated with other
(Fig. 3J). In conclusion, these
findings suggest that ST8SIA6-AS1 directly interacts with miR-338
to suppress its expression levels in HCC.
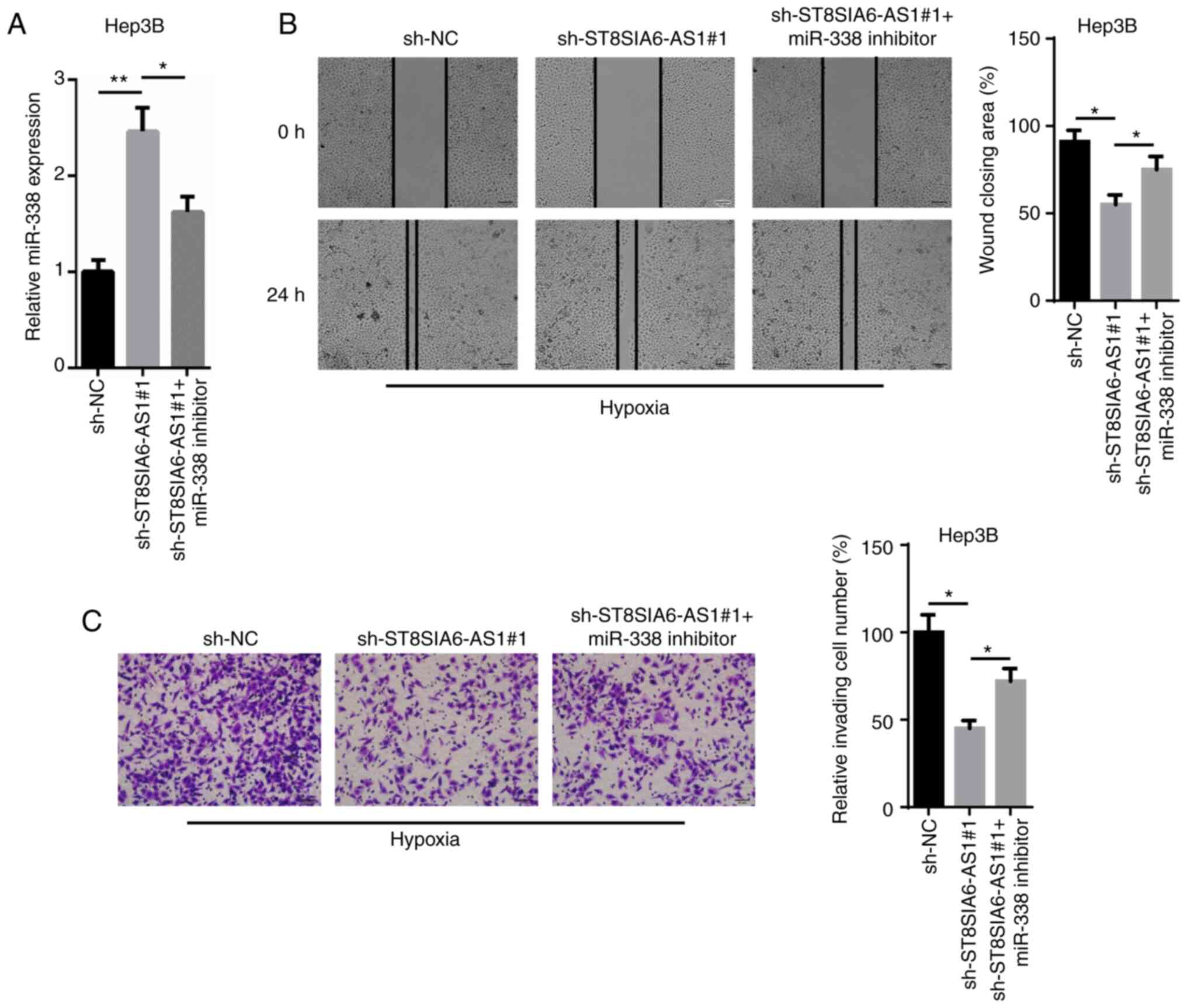
miR-338 partially counteracts the
effect of ST8SIA6-AS1 in hypoxia-treated HCC cell progression
To determine whether miR-338 is an important
effector for ST8SIA6-AS1-mediated carcinogenesis and the
development of hypoxia-treated HCC cells, an miR-338 inhibitor was
transfected into sh-ST8SIA6-AS1-expressing Hep3B cells. The
upregulation of miR-338 mediated by ST8SIA6-AS1 silencing was
reversed following the transfection of the miR-338 inhibitor
(Fig. 4A). Moreover, the wound
healing and Transwell assays indicated that the inhibitory effects
caused by the genetic knockdown of ST8SIA6-AS1 on cell migration
and invasion were abolished following miR-338 inhibition (Fig. 4B and C). Taken together, these
findings suggest that miR-338 may partially counteract the effects
of ST8SIA6-AS1 in regards to hypoxia-induced HCC progression.
MEPCE is directly targeted and
downregulated by miR-338
According to the prediction using the TargetScan
website (http://www.targetscan.org), MEPCE was
discovered to contain a complementary binding site for miR-338
(Fig. 5A). The RIP assay revealed
that both miR-338 and MEPCE were abundantly expressed in the Ago2
group, but not in the IgG group (Fig.
5B). A dual-luciferase reporter assay also discovered that the
overexpression of miR-338 decreased the relative luciferase
activity of the MEPCE-WT reporter in Hep3B cells, while the
relative luciferase activity in cells transfected with the
MEPCE-Mut reporter remained the same (Fig. 5C). Moreover, MEPCE mRNA expression
levels were suppressed following the transfection of miR-338 mimics
into Hep3B cells (Fig. 5D). RT-qPCR
analysis demonstrated that the expression levels of MEPCE were
significantly upregulated in HCC tissues and cells compared with
normal tissues and cells (Fig. 5E and
F). In addition, a negative correlation was identified between
the expression levels of MEPCE and miR-338 in HCC tissues, whereas
a positive correlation was revealed between the expression levels
of MEPCE and ST8SIA6-AS1 (Fig. 5G).
Altogether, these findings indicate that MEPCE expression levels
may be upregulated in HCC and may directly interact with
miR-338.
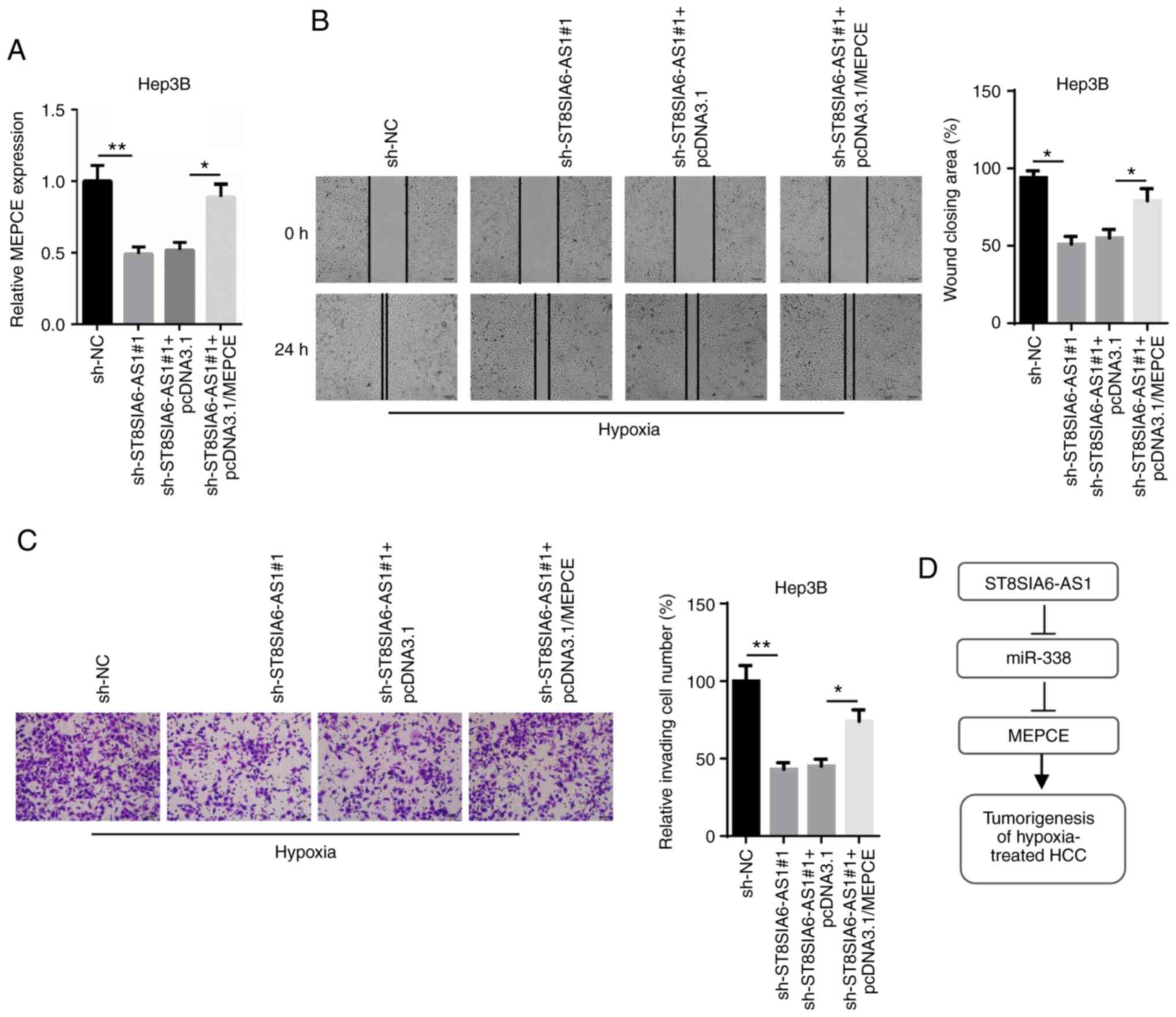
ST8SIA6-AS1 regulates the migration
and invasion of hypoxia-treated HCC cells by targeting MEPCE
To validate that MEPCE mediates the oncogenic role
of ST8SIA6-AS1, rescue assays were performed. The downregulation of
MEPCE mRNA expression levels induced by ST8SIA6-AS1 inhibition were
reversed following the transfection with pcDNA3.1/MEPCE (Fig. 6A). Additionally, both wound healing
and Transwell Matrigel assays revealed that the suppressive effect
of ST8SIA6-AS1 knockdown on cell migration and invasion was
reversed following MEPCE overexpression in hypoxia-treated Hep3B
cells (Fig. 6B and C). These
findings suggest that ST8SIA6-AS1 may affect the migration and
invasion of HCC cells under hypoxia by targeting MEPCE.
Discussion
Hepatocellular carcinoma (HCC) has contributed to
considerable mortality rates globally, demonstrating a high
incidence rate and an unfavorable prognosis (20). Modern medical treatments, such as
surgical resection, radiotherapy, biotherapy and chemotherapy have
been adopted to treat HCC; however, the overall survival rate of
patients with HCC remains low due to the high recurrence rates and
distant metastasis (21,22). Previous studies have identified
several molecular mechanisms of different long non-coding RNAs
(lncRNAs), including SNHG16, MIAT and HANR, in HCC (23–25).
However, the role of ST8SIA6-AS1 in HCC remains to be
determined.
lncRNAs have been discovered to modulate several
cellular processes, including cell proliferation, apoptosis,
differentiation, migration and invasion in numerous types of
cancer. For instance, the lncRNA C5orf66-AS1 was found to
accelerate the proliferation and epithelial-mesenchymal transition
(EMT) of cervical cancer cells by regulating the miR-637/RING1 axis
(26), while the lncRNA ZEB2-AS1
was found to promote cell proliferation, but prevent cell
apoptosis, by targeting the miR-143-5p/HIF-1α axis in gastric
cancer (27). Similarly, in the
present study, ST8SIA6-AS1 expression levels were revealed to be
upregulated in HCC tissues and cell lines, and the upregulated
expression levels of ST8SIA6-AS1 were closely associated with a
poor prognosis. Additionally, hypoxia has been demonstrated to be
strongly related with malignant progression, tumor metastasis and
drug resistance (28). However,
although hypoxia has been identified to be closely associated with
cellular processes in HCC (21),
the mechanism by which this occurs remains unknown. In the present
study, the migratory and invasive abilities of HCC cells were
promoted following hypoxia induction, while these effects were
hindered following the genetic knockdown of ST8SIA6-AS1.
Mechanistically, it has been widely established that
lncRNAs form a competitive endogenous RNA (ceRNA) network to
regulate the downstream target genes of miRNAs (29). For example, SPRY4-IT1 was proposed
to function as a ceRNA to sponge miR-101-3p and regulate EZH2
expression levels in bladder cancer (30). In addition, the lncRNA H19 was found
to modulate gastric cancer cell growth and metastasis through the
miR-22-3p/Snail1 axis (31). In the
present study, the majority of ST8SIA6-AS1 was observed to be
distributed in the cytoplasm of HCC cells, which may resemble the
ceRNA pattern of lncRNAs. miR-338, of which its expression levels
were found to be downregulated in HCC cells and tissues, was
predicted and revealed to bind with ST8SIA6-AS1, and the inhibition
of miR-338 partially counteracted the suppressive influence of
ST8SIA6-AS1 silencing on cell migration and invasion.
Methylphosphate capping enzyme (MEPCE) is an RNA
methyltransferase that methylates and stabilizes 7SK in the
nucleosol (32). Several previous
studies have reported that MEPCE is often overexpressed in human
breast cancer cells, and the upregulation of MEPCE is associated
with the tumorigenic phenotype and a poor prognosis in breast
cancer (33,34). However, to the best of our
knowledge, the biological functions of MEPCE remain unknown in HCC.
In the present study, MEPCE was revealed to be directly targeted by
miR-338. Additionally, MEPCE expression levels were upregulated and
negatively correlated with miR-338 expression levels in HCC.
Finally, rescue assays revealed that the overexpression of MEPCE
reversed the inhibitory effects of ST8SIA6-AS1 inhibition on
cellular processes.
In conclusion, the results of the present study
suggest that ST8SIA6-AS1 may promote the migration and invasion of
hypoxia-induced HCC cells via the miR-338/MEPCE axis (Fig. 6D). Thus, these findings suggest that
ST8SIA6-AS1 may represent a potential therapeutic target for
patients with HCC.
Acknowledgements
Not applicable.
Funding
This work was supported by the foundations of
Jiangsu Provincial Commission of Health and Family Planning
(H2018037, BJ18010); the Basic Research Program of Jiangsu Province
(BK20191153); and the Foundation of Xuzhou Institute of Technology
(KC19035).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BZ and RW designed the present study. ZL, KC and JL
performed all of the experiments. WS and QW analyzed the data and
prepared the figures. BZ and RW drafted the initial manuscript. BZ
reviewed and revised the manuscript. All authors approved the final
version of the manuscript.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of the Affiliated Hospital of Xuzhou Medical University
and written informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Banaudha KK and Verma M: Epigenetic
biomarkers in liver cancer. Methods Mol Biol. 1238:65–76. 2015.
View Article : Google Scholar
|
|
2
|
Raza A and Sood GK: Hepatocellular
carcinoma review: Current treatment, and evidence-based medicine.
World J Gastroenterol. 20:4115–4127. 2014. View Article : Google Scholar
|
|
3
|
Reig M, Mariño Z, Perelló C, Iñarrairaegui
M, Ribeiro A, Lens S, Díaz A, Vilana R, Darnell A, Varela M, et al:
Unexpected high rate of early tumor recurrence in patients with
HCV-related HCC undergoing interferon-free therapy. J Hepatol.
65:719–726. 2016. View Article : Google Scholar
|
|
4
|
Jarroux J, Morillon A and Pinskaya M:
History, discovery, and classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar
|
|
5
|
Kim S, Jeong H, Kim EY, Kim JF, Lee SY and
Yoon SH: Genomic and transcriptomic landscape of Escherichia coli
BL21(DE3). Nucleic Acids Res. 45:5285–5293. 2017. View Article : Google Scholar
|
|
6
|
Xu J, Bai J, Zhang X, Lv Y, Gong Y, Liu L,
Zhao H, Yu F, Ping Y, Zhang G, et al: A comprehensive overview of
lncRNA annotation resources. Brief Bioinform. 18:236–249. 2017.
|
|
7
|
Tian JB, Cao L and Dong GL: Long noncoding
RNA DDX11-AS1 induced by YY1 accelerates colorectal cancer
progression through targeting miR-873/CLDN7 axis. Eur Rev Med
Pharmacol Sci. 23:5714–5729. 2019.
|
|
8
|
Xie Q, Lin S, Zheng M, Cai Q and Tu Y:
Long noncoding RNA NEAT1 promotes the growth of cervical cancer
cells via sponging miR-9-5p. Biochem Cell Biol. 97:100–108. 2019.
View Article : Google Scholar
|
|
9
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar
|
|
10
|
Alvarez-Dominguez JR and Lodish HF:
Emerging mechanisms of long noncoding RNA function during normal
and malignant hematopoiesis. Blood. 130:1965–1975. 2017. View Article : Google Scholar
|
|
11
|
Zhu J, Fu H, Wu Y and Zheng X: Function of
lncRNAs and approaches to lncRNA-protein interactions. Sci China
Life Sci. 56:876–885. 2013. View Article : Google Scholar
|
|
12
|
Qi X, Zhang DH, Wu N, Xiao JH, Wang X and
Ma W: ceRNA in cancer: Possible functions and clinical
implications. J Med Genet. 52:710–718. 2015. View Article : Google Scholar
|
|
13
|
Chiu HS, Rodríguez Martínez M, Bansal M,
Subramanian A, Golub TR, Yang X, Sumazin P and Califano A:
High-throughput validation of ceRNA regulatory networks. BMC
Genomics. 18:4182017. View Article : Google Scholar
|
|
14
|
Sun Q, Li J, Li F, Li H, Bei S, Zhang X
and Feng L: lncRNA LOXL1-AS1 facilitates the tumorigenesis and
stemness of gastric carcinoma via regulation of miR-708-5p/USF1
pathway. Cell Prolif. 52:e126872019. View Article : Google Scholar
|
|
15
|
Yao N, Fu Y, Chen L, Liu Z, He J, Zhu Y,
Xia T and Wang S: Long non-coding RNA NONHSAT101069 promotes
epirubicin resistance, migration, and invasion of breast cancer
cells through NONHSAT101069/miR-129-5p/Twist1 axis. Oncogene.
38:7216–7233. 2019. View Article : Google Scholar
|
|
16
|
Wojczakowski W, Kobylarek D, Lindner J,
Limphaibool N and Kaczmarek M: MicroRNAs-novel biomarkers for
malignant pleural effusions. Contemp Oncol (Pozn). 23:133–140.
2019.
|
|
17
|
Chhatriya B, Mukherjee M, Ray S, Sarkar P,
Chatterjee S, Nath D, Das K and Goswami S: Comparison of tumour and
serum specific microRNA changes dissecting their role in pancreatic
ductal adenocarcinoma: A meta-analysis. BMC Cancer. 19:11752019.
View Article : Google Scholar
|
|
18
|
Liu X and Cui M: miRNA-98-5p inhibits the
progression of osteosarcoma by regulating cell cycle via targeting
CDC25A expression. Eur Rev Med Pharmacol Sci. 23:9793–9802.
2019.
|
|
19
|
Yang T, Tian S, Wang L, Wang Y and Zhao J:
MicroRNA-367-3p overexpression represses the proliferation and
invasion of cervical cancer cells through downregulation of
SPAG5-mediated Wnt/β-catenin signaling. Clin Exp Pharmacol Physiol.
47:687–695. 2019. View Article : Google Scholar
|
|
20
|
Chen DH, Wu QW, Li XD, Wang SJ and Zhang
ZM: SYPL1 overexpression predicts poor prognosis of hepatocellular
carcinoma and associates with epithelial-mesenchymal transition.
Oncol Rep. 38:1533–1542. 2017. View Article : Google Scholar
|
|
21
|
Zhao ZB, Chen F and Bai XF: Long noncoding
RNA MALAT1 regulates hepatocellular carcinoma growth under hypoxia
via sponging microRNA-200a. Yonsei Med J. 60:727–734. 2019.
View Article : Google Scholar
|
|
22
|
Liang R, Lin Y, Ye JZ, Yan XX, Liu ZH, Li
YQ, Luo XL and Ye HH: High expression of RBM8A predicts poor
patient prognosis and promotes tumor progression in hepatocellular
carcinoma. Oncol Rep. 37:2167–2176. 2017. View Article : Google Scholar
|
|
23
|
Xiao J, Lv Y, Jin F, Liu Y, Ma Y, Xiong Y,
Liu L, Zhang S, Sun Y, Tipoe GL, et al: lncRNA HANR promotes
tumorigenesis and increase of chemoresistance in hepatocellular
carcinoma. Cell Physiol Biochem. 43:1926–1938. 2017. View Article : Google Scholar
|
|
24
|
Huang X, Gao Y, Qin J and Lu S: lncRNA
MIAT promotes proliferation and invasion of HCC cells via sponging
miR-214. Am J Physiol Gastrointest Liver Physiol. 314:G559–G565.
2018. View Article : Google Scholar
|
|
25
|
Xu F, Zha G, Wu Y, Cai W and Ao J:
Overexpressing lncRNA SNHG16 inhibited HCC proliferation and
chemoresistance by functionally sponging hsa-miR-93. Onco Targets
Ther. 11:8855–8863. 2018. View Article : Google Scholar
|
|
26
|
Rui X, Xu Y, Jiang X, Ye W, Huang Y and
Jiang J: Long non-coding RNA C5orf66-AS1 promotes cell
proliferation in cervical cancer by targeting miR-637/RING1 axis.
Cell Death Dis. 9:11752018. View Article : Google Scholar
|
|
27
|
Wu F, Gao H, Liu K, Gao B, Ren H, Li Z and
Liu F: The lncRNA ZEB2-AS1 is upregulated in gastric cancer and
affects cell proliferation and invasion via miR-143-5p/HIF-1α axis.
Onco Targets Ther. 12:657–667. 2019. View Article : Google Scholar
|
|
28
|
Tatum JL, Kelloff GJ, Gillies RJ, Arbeit
JM, Brown JM, Chao KSC, Chapman JD, Eckelman WC, Fyles AW, Giaccia
AJ, et al: Hypoxia: Importance in tumor biology, noninvasive
measurement by imaging, and value of its measurement in the
management of cancer therapy. Int J Radiat Biol. 82:699–757. 2006.
View Article : Google Scholar
|
|
29
|
Zhang X, Hamblin MH and Yin KJ: The long
noncoding RNA Malat1: Its physiological and pathophysiological
functions. RNA Biol. 14:1705–1714. 2017. View Article : Google Scholar
|
|
30
|
Liu D, Li Y, Luo G, Xiao X, Tao D, Wu X,
Wang M, Huang C, Wang L, Zeng F and Jiang G: lncRNA SPRY4-IT1
sponges miR-101-3p to promote proliferation and metastasis of
bladder cancer cells through up-regulating EZH2. Cancer Lett.
388:281–291. 2017. View Article : Google Scholar
|
|
31
|
Gan L, Lv L and Liao S: Long noncoding RNA
H19 regulates cell growth and metastasis via the miR223p/Snail1
axis in gastric cancer. Int J Oncol. 54:2157–2168. 2019.
|
|
32
|
Shelton SB, Shah NM, Abell NS, Devanathan
SK, Mercado M and Xhemalçe B: Crosstalk between the RNA methylation
and histone-binding activities of MePCE regulates P-TEFb activation
on chromatin. Cell Rep. 22:1374–1383. 2018. View Article : Google Scholar
|
|
33
|
Liu R, Wang X, Chen GY, Dalerba P, Gurney
A, Hoey T, Sherlock G, Lewicki J, Shedden K and Clarke MF: The
prognostic role of a gene signature from tumorigenic breast-cancer
cells. N Engl J Med. 356:217–226. 2007. View Article : Google Scholar
|
|
34
|
Yao L, Chi Y, Hu X, Li S, Qiao F, Wu J and
Shao ZM: Elevated expression of RNA methyltransferase BCDIN3D
predicts poor prognosis in breast cancer. Oncotarget.
7:53895–53902. 2016. View Article : Google Scholar
|