Hepatocellular carcinoma (HCC) is the sixth most
common type of cancer worldwide and the third leading cause of
cancer-associated death (1). Most
patients cannot undergo radical surgery due to the presence of
intrahepatic or distant organ metastases, and at present, the
primary treatment methods for HCC include surgery, local ablation
therapy and radiation intervention (2). These methods allow for effective
treatment and management of patients with HCC during the early
stages, with 5-year survival rates as high as 70% (3). Despite the continuous development of
traditional treatment methods, the issue of recurrence and
metastasis of HCC, causing adverse effects to the survival and
prognosis of patients, remains a notable challenge. HCC is a
heterogeneous and chameleonic disease, with involvement of the
tumor microenvironment. HCC cells detach from the primary site,
migrate and invade the extracellular matrix, transfer to the target
organs and form metastatic lesions (4). Over 100 years ago, Paget (5) proposed the ‘seed and soil theory’ to
explain the directional migration of cancer cells and invasion to
specific organs. During tumor initiation, liver progenitor cells
and hepatocytes are hypothesized to undergo genetic and epigenetic
changes, which, together with disorders of the microenvironment,
eventually give rise to a distinct subpopulation of cancer stem
cells (CSCs) that have stem-like properties (6). Liver CSCs are considered to be
responsible for HCC tumorigenesis, progression, metastasis and
recurrence (7). Although
significant progress has been made in the exploration of the
mechanisms underlying the detachment of HCC cells, and therefore in
migration and invasion, preventing HCC metastasis remains a
poignant challenge for clinicians.
It has been found that chemokine networks may serve
pivotal roles in inducing organ-specific metastasis (8). Chemokines are a class of small
molecular proteins with similar structures, functions and
chemotactic properties, and their molecular weights are ~10 kDa,
and chemokines represent the largest member of the cytokine family
(9). At present, >50 chemokines
have been identified, which can be divided into four families: CXC,
CX3C, CC and XC, based on the different positions of the conserved
N-terminal cysteine residues (9).
Chemokines bind to a variety of different receptors, which belong
to the G-protein-binding receptor family, and there are ~23 types
of chemokine receptors that have been discovered (10). The receptors are composed of seven
transmembrane domains connected by extracellular and intracellular
loops and transfer signals by heterotrimeric Gi proteins (10). Chemokines and their receptors were
initially thought to allow for an interaction between immune cells
and the inflammatory sites (11).
After binding to the receptors, chemokines primarily serve a role
in migration of leukocytes, such as monocytes, eosinophils and
dendritic cells (DCs) (11).
Additionally, chemokines can directly act on non-immune cells, such
as tumor cells and vascular endothelial cells, to regulate the
proliferation, invasion and metastasis of tumor cells to promote
the progression of cancer in the tumor microenvironment (9). In addition, with the action of
specific chemokines, chemotaxis of different immune cell subgroups
to the tumor microenvironment is observed, which allows further
regulation of the tumor immune response (12). In addition to typical chemokine
receptors, chemokines can bind to the atypical chemokine receptor
(ACKR) subfamily, which is a key regulator of the chemokine
network, and is primarily expressed by stromal cells and
endothelial cells (13). At
present, ACKRs have not been shown to transmit signals, but instead
function to degrade chemokines, thus controlling the extent of
signal transduction (12). ACKR1
binds to >20 different CC and CXC chemokines (13). ACKR2 can bind to and degrade all CC
family chemokines, and ACKR3 is the scavenging receptor of CXC
motif chemokine ligand 12 (CXCL12) and CXCL11, which serves a key
role in tumor development (14). In
addition, ACKR4 primarily scavenges CC motif chemokine ligand 19
(CCL19), CCL21 and CCL25 (15,16).
Therefore, chemokines and their receptors can directly or
indirectly shape the tumor cell microenvironment, and then affect
the proliferation, invasion and metastasis of tumor cells. The
association between the chemokine regulatory network and the tumor
is complex and tissue-dependent. Some chemokines serve a role in
promoting cancer, whereas others have a cancer-suppressing effect;
for example, CCL2, CXCL10 and CX3C motif chemokine ligand 1
(CX3CL1)/CX3C motif chemokine receptor 1 (CX3CR1) can serve as
favorable or unfavorable prognostic factors depending on the
specific type of cancer (17). With
the in-depth study of chemokines, increasing attention has been
paid to the biological role and mechanism of chemokines in the
occurrence and development of HCC, particularly in the process of
metastasis (12,17). In addition, chemokines on the
exosomal surface may affect the chemotaxis of exosomes in the tumor
microenvironment. The present review provides a summary of the
relevant literature surrounding this topic, and discusses the
association between HCC and chemokines.
The morbidity (>20 cases/100,000 population in
East Asia) and mortality (95%) rates of HCC remain high, and there
are ~782,000 new cases and 600,000 deaths worldwide per year
(18). And the liver, one of the
most common metastatic organs of tumor cells, contains multiple
cellular components, such as hepatic stellate cells, macrophages
and sinusoidal endothelial cells, as well as acellular components,
such as matrix metalloproteinases (MMPs) (4,19).
Development of HCC may be associated with inflammation. Chronic
liver inflammation causes continuous damage and regeneration of
hepatocytes, which promotes the occurrence and development of HCC
(19). Additionally,
tumor-associated chronic inflammation involved the accumulation of
immune cells to the tumor (20,21),
and chemokines serve a key role in the recruitment and activation
of immune cells, and participate in tumor progression, invasion and
metastasis (22,23). HCC cells express a large number of
chemokines and their receptors. Han et al (24) injected human hepatocellular
carcinoma CBRH-7919 cells subcutaneously into nude mice, then
compared the expression levels of genes involved in tumor
progression between liver cancer and paracancerous tissues using
chemokine microarray analysis. The results revealed that 50
chemokine-associated genes were upregulated and another 3 genes
were downregulated in HCC, and western blotting confirmed the
changes of CXCL1, CXCL2, CXCL3 and CXC motif chemokine receptor 1
(CXCR1) expression in a CBRH-7919 mouse model (24). Additionally, silencing of the CXCL1
gene could inhibit the growth of CBRH-7919 tumors and significantly
decrease the protein expression levels of CXCL2, CXCL3 and IL-1β
(24). Dagouassat et al
(25) demonstrated that CCL2
derived from liver stromal cells mediated the migration and
invasion of HCC cells. Zhang et al (26) found that liver stromal cells
activated by tumors exhibited increased chemokine secretion,
including CCL1-6, CCL9, CCL12, CCL25, CXCL2, CXCL10 and CXCL12, and
that chemotaxis was associated with various signaling pathways,
including activation of the JAK-STAT signaling pathway and T-cell
receptor signaling pathway. The aforementioned studies demonstrate
that chemokines and their receptors, the expression of which is
frequently observed in HCC, may serve as novel therapeutic targets.
Table I shows the chemokines and
their receptors associated with HCC.
Unlike CXCL12 and CXCL1, CXCL16 expression in the
liver decreases when a tumor metastasizes to the liver (27). Typically, CXCL16 in the liver leads
to the migration of M1 macrophages to the liver, which results in
apoptosis of cancer cells by secreting TNF-α (28). In addition, the interaction between
CXCR6 and CXCL16 leads to an increase in IL-4 secretion, which
activates natural killer (NK) T cells to inhibit the proliferation
of malignant cells (27). A study
on gliomas revealed that CXCL16 and CX3CL1 downregulate the
expression levels of vascular endothelial growth factor C (29), suggesting that they may inhibit
tumor growth by decreasing angiogenesis. However, Takiguchi et
al (30) found that soluble
CXCL16 produced by mesenchymal stem cells (MSCs) via Wnt5a-Ror2
increased proliferation of human gastric cancer MKN45 cells
expressing CXCR6. In vivo and in vitro experiments
have demonstrated that by sequestrating CCL19 and CCL21, ACKR4
(also known as CCRL1) inhibits β-catenin from entering the nucleus,
blocking the Akt-GSK3β signaling pathway (31). Subsequently, the expression levels
of cancer-promoting factors cyclin D1 and c-Myc are decreased,
achieving the effect of inhibiting HCC (31). Shi et al (32) revealed that the increase of
CCR7+ mononuclear cells was significantly associated
with worse survival and increased recurrence, and confirmed that
ACKR4 had the potential to hinder CCR7+ cell chemotaxis
to HCC tissues by inhibiting CCL19 and CCL21. The aforementioned
studies suggest that chemokines have different effects in the tumor
microenvironment, including HCC. However, the specific roles of
chemokines in different cellular microenvironments and the
underlying mechanisms require further study.
As the only member of the CX3C chemokine subfamily,
CX3CL1 is a transmembrane glycoprotein that exists in two forms:
Membrane-bound and secretory. In addition, the mucin-like domain of
CX3CL1 contains a cleavage site that allows metalloproteinases
(such as ADAM10) to cleave and release proteins in a soluble form
(33). CXCR3 is the only receptor
of CX3CL1, and CX3CL1/CX3CR1 regulates the chemotaxis of
inflammatory cells, as well as the proliferation, migration and
invasion of cancer cells (34,35).
It has been reported that CX3CL1 is closely associated with the
metastasis of prostate (36) and
breast cancer (37). Additionally,
these studies confirmed that inhibition of CX3CL1 was an effective
strategy to prevent spinal metastasis of breast and prostate cancer
(36,37). Sun et al (34) revealed that CX3CL1/CX3CR1 expression
in HCC spinal metastases was upregulated. Bone marrow endothelial
cells promote the migration and invasion of Hep3B and MHCC97H cells
to the spine by secreting soluble CX3CL1 and this is inhibited
following neutralization of CX3CL1 (34). In CX3CR1 gene-deficient mice, Zheng
et al (38) found that the
metastasis of colon cancer cells to the liver was significantly
suppressed, and the upregulation of CX3CR1 gene expression in
tumor-associated cells was associated with worse prognosis.
Additionally, it was confirmed that CX3CR1 deficiency inhibited
macrophage aggregation and promoted macrophage apoptosis in
metastatic tumors, thus serving an antitumor effect (38). Considering the dual functions of
CX3CL1 as a chemoattractant for leukocytes and adhesion molecules
for tumor cells, CX3CL1 may have dual effects, promoting tumor
progression or antitumor effects (35). Chen et al (39) found that the increase of CX3CL1
expression levels were associated with liver metastasis and poor
prognosis after hepatectomy. Additionally, it was shown that
miR-561-5p downregulated CX3CL1 mRNA expression, leading to a
decrease in chemotaxis, functional regulation and infiltration of
CX3CR1+ NK cells, thus promoting HCC growth and lung
metastasis (39). Miao et al
(40) demonstrated that members of
the CX3CR1/Syk/PI3K signaling pathway were essential for
CX3CL1-induced platelet migration, which induced apoptosis of HCC
cells in vitro, thus inhibiting HCC growth. These studies
suggest that CX3CL1/CX3CR1 are potential therapeutic targets
against HCC growth and metastasis. Interference with CX3CL1/CX3CR1
may have future clinical implications for HCC prevention and
treatment.
Among the four types of chemokines, there are two
highly homologous XC chemokines: XC motif chemokine ligand 1 (XCL1)
and XCL2, whose genes (SCYC1 and SCYC2, respectively) are located
on chromosome 1. There are only two amino acid differences between
them, and XCL1 is the variant with the predominant effect (41). Unlike CXC, CC and CX3C chemokines
with two disulfide bonds and four cysteine residues at the
N-terminal, XCL1 has only one disulfide bond and two cysteine
residues (42). XC motif chemokine
receptor 1 (XCR1) is the receptor of XCL1, and human XCR1 is
primarily expressed in BDCA3+/CD141+ DCs,
which serve an important role in antigen presentation and enhancing
cytotoxic T-cell response (43).
Similar to other family members, XCR1 serves an important role in
human tumors. Several studies have revealed that XCR1 expression is
upregulated in ovarian, oral and breast cancer, where it promotes
cancer cell proliferation, adhesion, migration and invasion
(44–46). For example, Kim et al
(46) found that XCR1 was expressed
in both primary and metastatic human epithelial ovarian cancer
specimens and cell lines, but not in normal ovaries or ovarian
surface epithelial cells, highlighting the association between XCR1
and cell carcinogenesis and migration. Wang et al (47) revealed that XCL1/XCR1 significantly
promoted the proliferation and migration of lung cancer cells,
whereas the role of XCL1 in cell proliferation and migration was
abrogated following XCR1-knockdown using small interfering RNA.
However, not all studies support this conclusion. Another study
assessed XCR1 expression in an HCC cell line and on the
proliferation, metastasis and invasive ability of HCC (48). In vitro, it was found that
XCR1 silencing promoted the migration and invasion of HCC, whereas
the overexpression of XCR1 had the opposite effect, and the
mechanism may be associated with the inhibition of
epithelial-mesenchymal transition (EMT) (48). Additionally, overexpression of XCR1
in HCCLM3 cells decreased tumor growth, partially due to the
inhibition of the MAPK and PI3K/Akt signaling pathways (48). The aforementioned studies indicate
that XCR1 may have different roles in different tumors and thus
warrants further study.
The CC chemokine subfamily consists of >20
members, and the N-terminal of the members contain two adjacent
cysteine residues (9). CC
chemokines serve indispensable roles in HCC. It has been
demonstrated that CCL2 promotes HCC invasion and the effect of EMT,
accompanied by the activation of the Hedgehog signaling pathway
(56). Additionally, CCL2 can
stimulate angiogenesis and further promote the progression of liver
cancer (56). The combination of
CCL2 with CCR4+ regulatory T cells (Tregs) and
CCR2+Ly-6C+ myeloid-derived suppressor cells
(MDSCs) affects the glioma microenvironment and causes
immunosuppression, thus promoting tumor development, which
indicates that CCL2/CCR2 and CCL2/CCR4 serve a crucial role in
inducing the migration of monocytic-MDSCs to precancerous lesions
(57,58). Qi et al (59) confirmed that CCL7 and its receptor
CCR3 were key mediators of invasion and metastasis of lung and
colon tumor cells. CCL7 modulates signal transmission by binding to
CCR1, CCR2 and CCR3, the expression levels of which are upregulated
in liver metastases, with CCR3 exhibiting higher expression levels
than CCR1 or CCR2 (59).
Furthermore, it was revealed that CCL7 regulated the transport of
MMP-9 to invasive cells through its receptor CCR3, thus promoting
the formation of functional invasive cells, degrading collagen and
invading extracellular matrix, a key condition for HCC migration
(59). Via targeted gene expression
microarray screening alterations in G-protein coupled receptor
family gene expression, Wu et al (60) found that CCR10 was significantly
upregulated in inflammation-driven HCC tumors and paracancerous
tissue hepatocytes. Additionally, CCR10, secreted by hepatocytes,
activates the PI3K/Akt signaling pathway via Akt phosphorylation,
and inhibits apoptosis and promotes compensatory proliferation to
drive the occurrence of HCC (60).
CCR6 is another member of the CC chemokine receptor family, and its
ligand CCL20 is also known as macrophage regulatory protein 3a
(20). It has been reported that
CCR6 is highly expressed in a variety of cancer cells and
participates in different tumor-associated behaviors (61). Several studies have demonstrated
that CCL20/CCR6 promotes the proliferation, adhesion and
chemotactic migration of HCC cells (62,63).
CCL20 mediates the chemotaxis of Tregs to the tumor
microenvironment and this process is achieved via the activation of
STAT3 (64). Furthermore, the
expression of epidermal growth factor receptor/Ras-induced CCL20 in
the tumor microenvironment stimulates tumor microangiogenesis,
leading to rapid tumor growth and progression (64). Blocking the activity of CCR6 in the
tumor microenvironment may inhibit tumor neovascularization and
enhance traditional antitumor therapy (61). CCR6, the specific receptor of CCL20,
may be a promising anticancer target. Other CC members, such as
CCL28, promote the infiltration of Tregs into the tumor
environment, induce angiogenesis and facilitate tumor cell escape
from immune surveillance (65). Gao
et al (66) reported that
CCL15 secreted by HCC cells can promote chemotaxis to cancer foci
by binding to CCR1 on the membrane of MSCs. Although unmodified
MSCs have no significant effect on the progression of HCC tumors,
they are considered to be a promising gene delivery vector in tumor
therapy (66). In addition, due to
the similar phenotypes of CCR7+ monocytes and Tregs,
they can both promote tumor progression by producing TGF-β1
(32). The aforementioned studies
highlight novel therapeutic targets for the treatment of HCC.
CCL5 can bind to CCR1 and CCR3, but the activity of
CCL5 is exerted by binding to CCR5 (63,67).
CCL5/CCR5 expression in HCC tissues is significantly higher
compared with in non-neoplastic liver tissues, and CCL5 binding to
CCR5 activates the PI3K/Akt/mTOR signaling pathway, which is
associated with the occurrence and development of HCC, and the CCR5
antagonist, maraviroc, reverses this effect (67). Additionally, CCL5/CCR5 regulates
EMT, promoting HCC metastasis (68). Furthermore, macrophages exposed to
hepatitis C virus exhibit increased secretion of CCL5, which
further activates hepatic stellate cells to induce hepatitis and
liver fibrosis (69), highlighting
the biological and clinical significance of CCL5/CCR5 in the
development of HCC. As CCR5 exists in immune cells and cancer
cells, it can exert dual effects, both antitumor and
tumor-promoting effects (70).
Thus, the function of CCL5/CCR5 remains controversial. However, in
general, CCL5/CCR5 serves a major role in promoting tumors
(68). These findings suggest that
CCL5/CCR5 may be a potential target for the treatment of HCC.
The CC subfamily is also associated with tumor drug
resistance. In lung cancer, CCL2 is associated with docetaxel
resistance via activation of the PI3K/Akt signaling pathway where
it inhibits caspase3-dependent apoptosis (71). In addition, stroma-derived CCL2 and
CCL5 induce tumor cells to release IL-6 by activating the PYK2
signaling pathway (located upstream of the JAK1/STAT3 signaling
pathway), thereby resulting in carboplatin resistance (72). Similarly, the regulation of CCL20
expression via ATP-binding cassette subfamily B member 1 membrane
transporter is associated with doxorubicin resistance (73). Vaquero et al (74) examined 109 genes that may be
associated with chemotherapeutic drug resistance in HCC and found
that CCL14 and CCL15 mRNA expression was upregulated following
activation of nuclear receptor FXR; this upregulation was involved
in drug resistance and the mechanism of chemoresistance (MOC) may
be associated with changes in the balance of apoptosis-promoting
(MOC-5a) and survival-promoting (MOC-5b) genes. By detecting the
expression levels of CC chemokines associated with drug resistance
and by performing targeted blocking, the efficacy of chemotherapy
drugs may be improved.
In summary, CC subfamily chemokines are increasingly
considered to be involved in a variety of tumor biological
processes, and are closely associated with the development,
metastasis and regulation of the tumor microenvironment of HCC.
However, further studies and novel techniques/therapeutics are
required to improve the diagnosis and treatment of HCC effectively
via regulation of CC subfamily chemokines.
In normal liver tissues, CXC chemokines are widely
involved in the repair of hepatocyte injury by affecting hepatocyte
proliferation and regeneration (79), and they serve an important role in
the occurrence and development of HCC and the tumor
microenvironment. Although the ligands and receptors have high
sequence homology (~80%), the ligand selectivity of receptors
varies, and CXC chemokines are divided into two groups according to
the presence or absence of Glu-Leu-Arg (ELR motif) at the amino
terminal, which exerts a vital effect on tumorigenesis and
development, particularly in tumor angiogenesis (80). Members containing the ELR motif
(including CXCL1, 2, 3, 5, 6, 7 and 8) bind to CXCR2 and exert
angiogenic effects, whereas most CXC chemokines without the ELR
motif bind to CXCR3 and lead to vascular inhibition (80). For example, the CXCL5/CXCR2 axis is
involved in EMT in HCC by activating the PI3K/Akt/GSK-3 β/Snail
signaling pathway and significantly enhances the proliferative,
migratory and invasive abilities of HCC cells (81,82).
CXCL8, also known as IL-8, is usually secreted by tumor cells and
inflammatory cells (12). In HCC,
the levels of CXCL8 are significantly increased compared with in
normal liver tissue, leading to progression and metastasis by
activating the AKT/mTOR/STAT3 signaling pathway (83).
Studies have shown that neutrophils expressing CXCR2
chemotactically enter the liver by binding to CXCL1 (84), releasing MMP8 and MMP9 to induce
angiogenesis (85). Additionally,
CXCL8 increases immune infiltration of Tregs into the tumor
microenvironment (22). Therefore,
evaluating the expression levels of chemokines in the liver may be
a prognostic factor for follow-ups in patients with HCC. CXCL10
expression is positively associated with increased serum
α-fetoprotein levels, tumor size, tumor number and TNM stage
(86). Kaplan-Meier survival curve
analyses revealed that the overall survival rate and disease-free
survival rate of patients with high CXCL10 expression was low;
additionally, univariate/multivariate analyses demonstrated that
CXCL10 was an independent prognostic factor for patients with HCC
(86). By measuring the levels of
CXCL13 in patients with HCC, Li et al (87) found that CXCL13 levels in patients
with tumors >5 cm were significantly higher compared with those
in patients with tumors <5 cm in size, and the levels of CXCL13
were positively associated with Child-Pugh score and negatively
correlated with albumin and cholinesterase levels. It has been
suggested that CXCL13 may be used as an index to evaluate liver
function (87). Song et al
(88) reported that CXCL2
expression was upregulated in the blood of patients with HCC and
promoted the proliferation and metastasis of HepG2 and PG5 cells.
Conversely, Ding et al (89)
overexpressed CXCL2 in MHCC97H and HCCLM3 cell lines using
lentiviral transfection, and revealed that the exogenous expression
of CXCL2 inhibited cell proliferation by promoting apoptosis and
cell cycle arrest at the G1 phase. Additionally, Subat
et al (90) found that CXCL2
levels were significantly downregulated in tumor tissues compared
with in normal liver tissues, but high CXCL2 expression was
positively associated with the number of HCC tumors. This may be
due to different molecular mechanisms of occurrence and development
in different tumor cell lines.
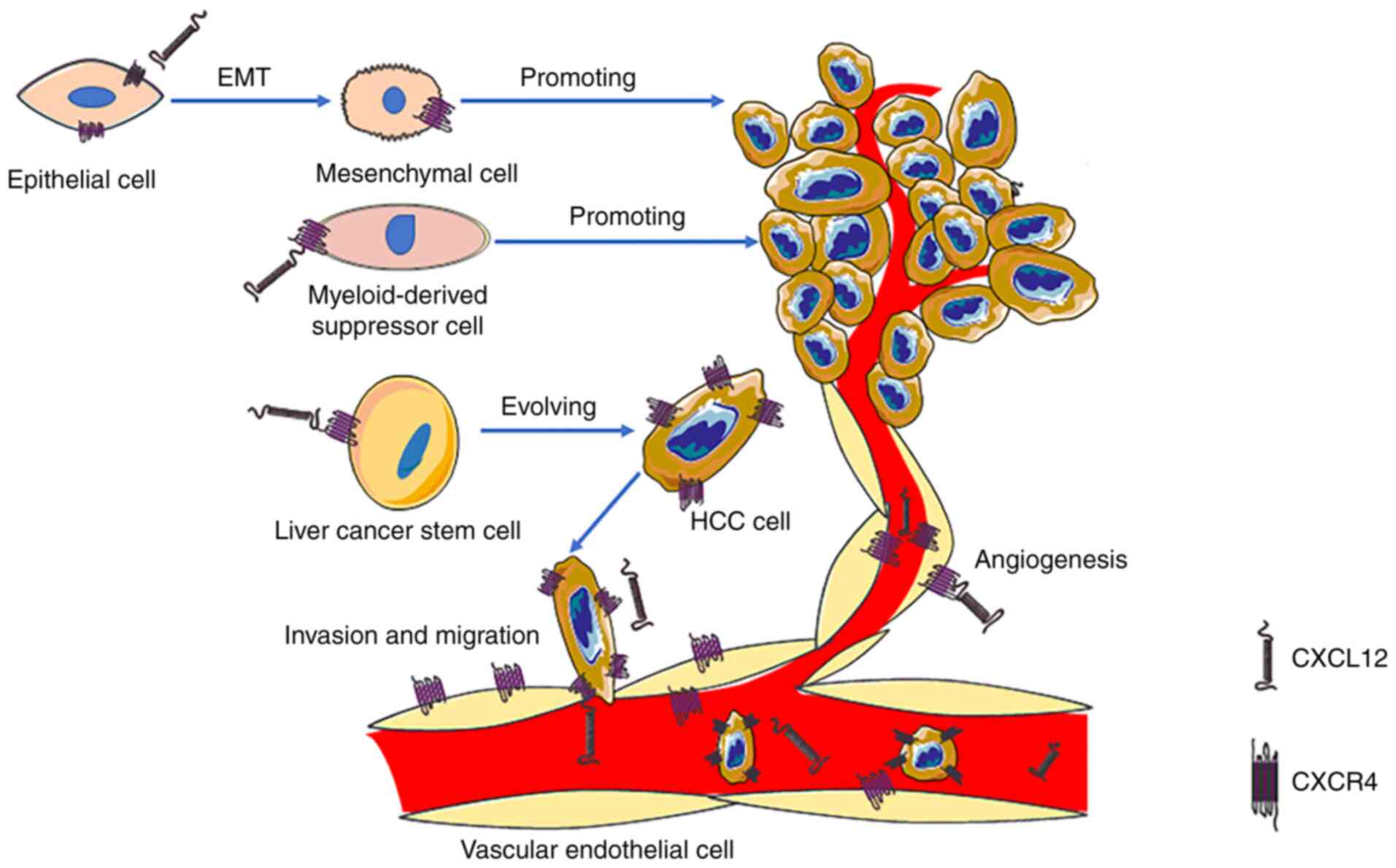
The role of CXCL12 is very important in the
development of HCC. In the past, it was considered that CXCR4 was
the only receptor of CXCL12. CXCL12/CXCR4 binds specifically and is
commonly involved in various pathophysiological processes, such as
anti-apoptotic and angiogenesis processes (29). Additionally, CXCL12/CXCR4 serves a
key role in cell migration to multiple organs, including the liver,
bone marrow and lungs (91).
Additionally, CXCL12/CXCR4 can promote EMT (92), decrease apoptosis and allow for
self-renewal of cancer cells by activating the anti-apoptotic
PI3K/Akt signaling pathway (93),
increase the expression levels of hypoxia-inducible factor and
activate MMPs, thereby jointly promoting angiogenesis and cancer
metastasis (94,95). Wang et al (96) found that the hepatitis B virus (HBV)
X protein (HBx) enhanced MDM2 expression by directly binding to
MDM2 and inhibiting its ubiquitin degradation, resulting in
enhanced transcriptional activity and CXCL12 and CXCR4 expression,
which in turn activated the Wnt/β-catenin signaling pathway and
enhanced the activity of OV6+ CSCs. In addition, the
aforementioned study indicated that the expression levels of any
two markers of the HBx/MDM2/CXCR4/OV6 axis in HCC biopsies could be
used to predict the prognosis of patients with HBV-associated HCC
(96). Another study demonstrated
that the occurrence rate of CXCR4 in the blood vessels of liver
cancer was ~50%, and that high CXCR4 expression indicated a poor
prognosis (97). These results
suggest that CXCR4 may be a prognostic factor and a potential
therapeutic target for HCC (Fig.
1). Knocking down CXCR4 gene expression using small interfering
RNA results in decreased invasiveness of uveal melanoma cells
exposed to soluble factors produced by the human liver (98). The U.S. Food and Drug Administration
has approved the CXCR4 antagonist plerixafor (Mozobil, AMD3100) for
peripheral blood stem cell mobilization following an autologous
transplantation (99). In addition,
Collins et al (100)
hypothesized that CXCL14 may be a positive allosteric modulator of
CXCR4, enhancing the potency of CXCR4. CXCR7 is another high
affinity receptor of CXCL12, which activates the Akt signaling
pathway to induce angiogenesis in HCC (101) and stimulates MAPK signal
transduction to promote HCC growth and invasion (102). These studies suggest that CXCL12
and its receptors serve a role that cannot be ignored in the
development of HCC.
CXCR3 is expressed in almost all cells, is
upregulated in several types of primary and metastatic tumors (such
as renal cancer, colon cancer and pulmonary metastasis tumor), and
is considered to be essential for migration of cancer cells
(12,103). Three variants of CXCR3 (CXCR3-A,
CXCR3-B and CXCR3-alt) have been identified, and two of these,
CXCR3-A and CXCR3-B, induce opposing physiological functions
(103). Overall, CXCR3-A promotes
cell proliferation, survival, chemotaxis, invasion and mediates
tumor metastasis, whereas CXCR3-B inhibits tumor growth and
promotes apoptosis and angiogenesis (104). Therefore, abnormal expression
levels of CXCR3-A and CXCR3-B can affect the progression of a
tumor. Compared with CXCR3-A, CXCR3-alt possesses a substantially
different C-terminal protein sequence, which differs from all known
chemokine receptors (104).
However, there are few studies on CXCR3-alt (104). At the cellular level, it has been
shown that knocking down CXCR3 significantly inhibits the
proliferation, adhesion, migration and invasion of HCC cells
(86). Gao et al (105) reported that high CXCR6 expression
promotes the invasion of HCC and the generation of an inflammatory
environment, and is associated with a poor prognosis in patients
with HCC. These results support the view that inhibition of the
CXCL16/CXCR6 pathway may improve the prognosis of HCC treatment.
CXCR7 is highly expressed in tumor endothelial cells, enhancing HCC
migration and invasion by affecting the phosphorylation of the
STAT3 signaling pathway (106).
The aforementioned studies suggest that targeted therapy for
chemokine receptors may be a potential direction for the diagnosis
and treatment of HCC.
MDSCs regulate the immunosuppressive network and
participate in all aspects of tumor progression, and their
migration is associated with autocrine or paracrine chemokines
(58). Xu et al (107) reported that hematopoietic stem
cells (HSCs) induce an increase in the number of MDSCs in HCC
lesions. Further animal experiments revealed that activated HSCs
secrete CCL2, CXCL1, CXCL5 and CXCL12 by binding to the MDSC
membrane surface receptors CCR2, CXCR2 and CXCR4 to mediate MDSC
migration (108). Additionally,
Zhang et al (26) found that
CCL2 and CXCL12 produced by tumor-associated liver stromal cells
contribute to the accumulation of MDSCs and promote HCC growth.
This highlights a potentially effective means of altering the tumor
microenvironment by regulating the secretion of chemokines from
liver mesenchymal cells.
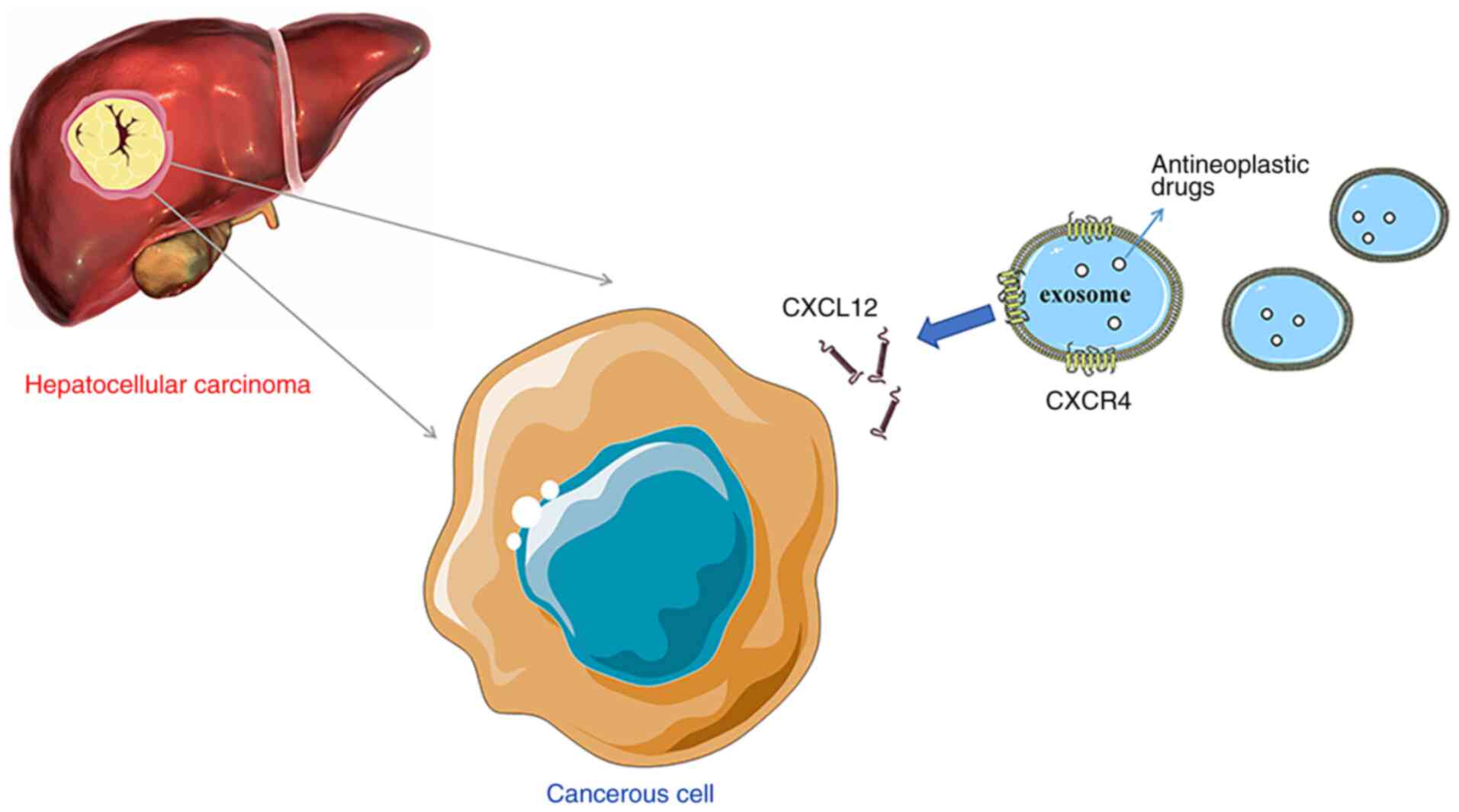
Whether chemokines are expressed on the surface of
exosomes, and whether they promote the chemotaxis of exosomes to
HCC has been investigated. Viñas et al (130) observed that CXCL12 expression in
kidneys was increased and that exosomes expressing CXCR4 were
enriched in mice with renal ischemia/reperfusion injury. Further
experiments revealed that hypoxia promoted CXCL12 expression in
human umbilical vein endothelial cells (HUVECs) (130). Compared with normal cells, HUVECs
exposed to hypoxia exhibited increased uptake of CXCR4-expressing
exosomes, and was significantly inhibited by the CXCR4 inhibitor
plerixafor (130). The
aforementioned study revealed the effect of CXCL12/CXCR4 in the
chemotaxis of exosomes to kidney endothelium. Additionally, Ciullo
et al (131) found that in
a mouse model of cardiac ischemia-reperfusion, compared with the
exosomes that did not express CXCR4, the exosomes expressing CXCR4
migrated more effectively using chemotaxis to the CXCL12-expressing
cardiomyocytes and exerted improved cardioprotective effects. Gao
et al (132) found that the
accumulation of exosomes produced by mature DCs in the spleen is
regulated by CCR7 on the exosomal membrane surface. A decrease in
CCR7 expression decreases the accumulation of exosomes in the
spleen and decreases the inflammatory reaction (132). In HCC, CXCL12 expression is
significantly increased compared with in normal liver tissues
(107). Theoretically, the
exosomes expressing CXCR4 may more effectively induce chemotaxis to
HCC and thus serve a role in targeted drug delivery. Thus, it was
hypothesized that the surface of modified HCC exosomes
overexpressing chemokine receptors, such as CXCR4, and loaded with
antitumor drugs, may enhance the chemotaxis of exosomes to HCC and
achieve the goal of targeted therapy (Fig. 2). Although exosomes have received
widespread attention, studies on this subject remain limited, and
the complexity of exosomes in the treatment of liver cancer is not
fully understood. In future studies, the association between
exocrine chemotaxis and chemokines should be further determined to
lay the foundation for the study of exosome-directed drug delivery
to the liver.
There is an urgent need to identify novel
therapeutic strategies for the management of HCC due to the poor
prognosis of patients. In recent years, immunotherapy and molecular
targeted therapy have become increasingly popular. However, the
unique immune response in the liver favors tolerance, which
represents a substantial challenge for conventional immunotherapy
in patients with HCC (133).
Altering the tumor microenvironment and preventing immune tolerance
are the goals of novel immune-based methods. HCC typically develops
alongside or following various other liver diseases (such as
hepatitis and cirrhosis), in which the constant inflammation
process enhances the formation and growth of the tumor (134). In a review by Gnoni et al
(4), it was suggested that the
cells present in the microenvironment, such as macrophages,
activated stellate cells and blood (platelets) cells, contribute to
stimulate and enhance tumorigenesis, angiogenesis and metastases of
HCC. Chemokines and their receptors not only affect the progression
of HCC directly or indirectly, but also serve an important
biological role in the migration of these cells, broadening the
research field to include host and tumor interactions. For example,
CCL15 recruits CCR1+CD14+ monocytes to the
HCC invasive margin, and these monocytes in turn suppress the
antitumor immune response, promote angiogenesis and accelerate HCC
metastasis (135). Blockage of the
CCL15-CCR1 axis decreases HCC growth and metastasis in vivo,
representing a promising therapeutic approach (135). Based on the involvement of
tumor-associated macrophages in cancer survival and proliferation,
targeting the chemokines and their receptors may result in a
valuable prognostic impact and may assist in the identification of
novel immunity-based cancer treatments. It is hypothesized that
patients with HCC may obtain more benefits when
chemokines/receptors are regulated.
In addition, exosomes, which mediate the transport
of various proteins, DNA and RNA molecules, may be used as drug
carriers; exosomes expressing chemokines can more effectively
induce chemotaxis to specific lesions (131–133). Thus, there is significant
potential for exosomes modified by chemokines to deliver drugs
directly, thus inhibiting the occurrence and development of HCC,
and this highlights the future direction for further research.
Not applicable.
The present review was supported by the Hebei
Provincial Government Clinical Medicine Talents Training and Basic
Research Project (grant no. 361003).
Not applicable.
DX and YZ wrote and reviewed the manuscript. DX, YL
and JH designed the figures. DX, JH and JW edited the manuscript.
DX, YZ and JW assessed all the raw data. YP and HT critically
revised the manuscript. YL, YP and HT are responsible for
confirming the authenticity of the data. All authors read and
approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Dong G, Zhang S, Shen S, Sun L, Wang X,
Wang H, Wu J, Liu T, Wang C, Wang H, et al: SPATS2, negatively
regulated by miR-145-5p, promotes hepatocellular carcinoma
progression through regulating cell cycle. Cell Death Dis.
11:8372020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang FP, Huang YP, Luo WX, Deng WY, Liu
CQ, Xu LB and Liu C: Construction of a risk score prognosis model
based on hepatocellular carcinoma microenvironment. World J
Gastroenterol. 26:134–153. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang XF, Yang X, Jia HL, Zhu WW, Lu L,
Shi W, Zhang H, Chen JH, Tao YF, Wang ZX, et al: Bcl-2 expression
is a poor predictor for hepatocellular carcinoma prognosis of
andropause-age patients. Cancer Biol Med. 13:459–468. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gnoni A, Santini D, Scartozzi M, Russo A,
Licchetta A, Palmieri V, Lupo L, Faloppi L, Palasciano G, Memeo V,
et al: Hepatocellular carcinoma treatment over sorafenib:
Epigenetics, microRNAs and microenvironment. Is there a light at
the end of the tunnel? Expert Opin Ther Targets. 19:1623–1635.
2015. View Article : Google Scholar
|
|
5
|
Paget S: The distribution of secondary
growth in cancer. Lancet. 1:571–573. 1889. View Article : Google Scholar
|
|
6
|
Sainz B and Heeschen C: Standing out from
the crowd: Cancer stem cells in hepatocellular carcinoma. Cancer
Cell. 23:431–433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li J and Zhu Y: Recent advances in liver
cancer stem cells: Non-coding RNAs, oncogenes and oncoproteins.
Front Cell Dev Biol. 8:5483352020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kakinuma T and Hwang ST: Chemokines,
chemokine receptors, and cancer metastasis. J Leukoc Biol.
79:639–651. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagarsheth N, Wicha MS and Zou W:
Chemokines in the cancer microenvironment and their relevance in
cancer immunotherapy. Nat Rev Immunol. 17:559–572. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Griffith JW, Sokol CL and Luster AD:
Chemokines and chemokine receptors: Positioning cells for host
defense and immunity. Annu Rev Immunol. 32:659–702. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakai N, Yoshidome H, Shida T, Kimura F,
Shimizu H, Ohtsuka M, Takeuchi D, Sakakibara M and Miyazaki M:
CXCR4/CXCL12 expression profile is associated with tumor
microenvironment and clinical outcome of liver metastases of
colorectal cancer. Clin Exp Metastasis. 29:101–110. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rezaeeyan H, Shirzad R, McKee TD and Saki
N: Role of chemokines in metastatic niche: New insights along with
a diagnostic and prognostic approach. APMIS. 126:359–370. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Raffaella B and Graham GJ: Atypical
chemokine receptors and their roles in the resolution of the
inflammatory response. Front Immunol. 7:2242016.PubMed/NCBI
|
|
14
|
Saaber F, Schütz D, Miess E, Abe P,
Desikan S, Ashok Kumar P, Balk S, Huang K, Beaulieu JM, Schulz S
and Stumm R: ACKR3 regulation of neuronal migration requires ACKR3
phosphorylation, but not β-Arrestin. Cell Rep. 26:1473–1488.e9.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Purvanov V, Matti C, Samson GPB, Kindinger
I and Legler DF: Fluorescently tagged CCL19 and CCL21 to monitor
CCR7 and ACKR4 functions. Int J Mol Sci. 19:38762018. View Article : Google Scholar
|
|
16
|
Matti C, D'Uonnolo G, Artinger M, Melgrati
S, Salnikov A, Thelen S, Purvanov V, Strobel TD, Spannagel L,
Thelen M and Legler DF: CCL20 is a novel ligand for the scavenging
atypical chemokine receptor 4. J Leukoc Biol. 107:1137–1154. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Do HTT, Lee CH and Cho J: Chemokines and
their receptors: Multifaceted roles in cancer progression and
potential value as cancer prognostic markers. Cancers (Basel).
12:2872020. View Article : Google Scholar
|
|
18
|
Ozakyol A: Global epidemiology of
hepatocellular carcinoma (HCC Epidemiology). J Gastrointest Cancer.
48:238–240. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Giannelli G, Rani B, Dituri F, Cao Y and
Palasciano G: Moving towards personalised therapy in patients with
hepatocellular carcinoma: The role of the microenvironment. Gut.
63:1668–1676. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Marra F and Tacke F: Roles for chemokines
in liver disease. Gastroenterology. 147:577–594.e1. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chiu DK, Xu IM, Lai RK, Tse AP, Wei LL,
Koh HY, Li LL, Lee D, Lo RC, Wong CM, et al: Hypoxia induces
myeloid-derived suppressor cell recruitment to hepatocellular
carcinoma through chemokine (C-C motif) ligand 26. Hepatology.
64:797–813. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kryczek I, Wang L, Wu K, Li W, Zhao E, Cui
T, Wei S, Liu Y, Wang Y, Vatan L, et al: Inflammatory regulatory T
cells in the microenvironments of ulcerative colitis and colon
carcinoma. Oncoimmunology. 5:e11054302016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kryczek I, Wu K, Zhao E, Wei S, Vatan L,
Szeliga W, Huang E, Greenson J, Chang A, Roliński J, et al:
IL-17+ regulatory T cells in the microenvironments of
chronic inflammation and cancer. J Immunol. 186:4388–4395. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han KQ, He XQ, Ma MY, Guo XD, Zhang XM,
Chen J, Han H, Zhang WW, Zhu QG, Nian H and Ma LJ: Inflammatory
microenvironment and expression of chemokines in hepatocellular
carcinoma. World J Gastroenterol. 21:4864–4874. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dagouassat M, Suffee N, Hlawaty H, Haddad
O, Charni F, Laguillier C, Vassy R, Martin L, Schischmanoff PO,
Gattegno L, et al: Monocyte chemoattractant protein-1 (MCP-1)/CCL2
secreted by hepatic myofibroblasts promotes migration and invasion
of human hepatoma cells. Int J Cancer. 126:1095–1108.
2010.PubMed/NCBI
|
|
26
|
Zhang H, He G, Kong Y, Chen Y, Wang B, Sun
X, Jia B, Xie X, Wang X, Chen D, et al: Tumour-activated liver
stromal cells regulate myeloid-derived suppressor cells
accumulation in the liver. Clin Exp Immunol. 188:96–108. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kee JY, Ito A, Hojo S, Hashimoto I,
Igarashi Y, Tsukada K, Irimura T, Shibahara N, Nakayama T, Yoshie
O, et al: Chemokine CXCL16 suppresses liver metastasis of
colorectal cancer via augmentation of tumor-infiltrating natural
killer T cells in a murine model. Oncol Rep. 29:975–982. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kee JY, Ito A, Hojo S, Hashimoto I,
Igarashi Y, Tsuneyama K, Tsukada K, Irimura T, Shibahara N,
Takasaki I, et al: CXCL16 suppresses liver metastasis of colorectal
cancer by promoting TNF-α-induced apoptosis by tumor-associated
macrophages. BMC Cancer. 14:9492014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Adamski V, Hattermann K, Kubelt C, Cohrs
G, Lucius R, Synowitz M, Sebens S and Held-Feindt J: Entry and exit
of chemotherapeutically-promoted cellular dormancy in glioblastoma
cells is differentially affected by the chemokines CXCL12, CXCL16,
and CX3CL1. Oncogene. 39:4421–4435. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Takiguchi G, Nishita M, Kurita K, Kakeji Y
and Minami Y: Wnt5a-Ror2 signaling in mesenchymal stem cells
promotes proliferation of gastric cancer cells by activating
CXCL16-CXCR6 axis. Cancer Sci. 107:290–297. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi JY, Yang LX, Wang ZC, Wang LY, Zhou J,
Wang XY, Shi GM, Ding ZB, Ke AW, Dai Z, et al: CC chemokine
receptor-like 1 functions as a tumour suppressor by impairing
CCR7-related chemotaxis in hepatocellular carcinoma. J Pathol.
235:546–558. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi JY, Duan M, Sun QM, Yang L, Wang ZC,
Mynbaev OA, He YF, Wang LY, Zhou J, Tang QQ, et al: Naive Treg-like
CCR7+ mononuclear cells indicate unfavorable prognosis
in hepatocellular carcinoma. Tumour Biol. 37:9909–9917. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wong HS, Jaumouillé V, Heit B, Doodnauth
SA, Patel S, Huang YW, Grinstein S and Robinson LA: Cytoskeletal
confinement of CX3CL1 limits its susceptibility to proteolytic
cleavage by ADAM10. Mol Biol Cell. 25:3884–3899. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun C, Hu A, Wang S, Tian B, Jiang L,
Liang Y, Wang H and Dong J: ADAM17-regulated CX3CL1 expression
produced by bone marrow endothelial cells promotes spinal
metastasis from hepatocellular carcinoma. Int J Oncol. 57:249–263.
2020.PubMed/NCBI
|
|
35
|
Liu W, Jiang L, Bian C, Liang Y, Xing R,
Yishakea M and Dong J: Role of CX3CL1 in diseases. Arch Immunol
Ther Exp (Warsz). 64:371–383. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu P, Liang Y, Jiang L, Wang H, Wang S
and Dong J: CX3CL1/fractalkine enhances prostate cancer spinal
metastasis by activating the Src/FAK pathway. Int J Oncol.
53:1544–1556. 2018.PubMed/NCBI
|
|
37
|
Liang Y, Yi L, Liu P, Jiang L, Wang H, Hu
A, Sun C and Dong J: CX3CL1 involves in breast cancer metastasizing
to the spine via the Src/FAK signaling pathway. J Cancer.
9:3603–3612. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zheng J, Yang M, Shao J, Miao Y, Han J and
Du J: Chemokine receptor CX3CR1 contributes to macrophage survival
in tumor metastasis. Mol Cancer. 12:1412013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen EB, Zhou ZJ, Xiao K, Zhu GQ, Yang Y,
Wang B, Zhou SL, Chen Q, Yin D, Wang Z, et al: The
miR-561-5p/CX3CL1 signaling axis regulates pulmonary
metastasis in hepatocellular carcinoma involving
CX3CR1+ natural killer cells infiltration.
Theranostics. 9:4779–4794. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miao S, Lu M, Liu Y, Shu D, Zhu Y, Song W,
Ma Y, Ma R, Zhang B, Fang C and Ming ZY: Platelets are recruited to
hepatocellular carcinoma tissues in a CX3CL1-CX3CR1 dependent
manner and induce tumour cell apoptosis. Mol Oncol. 14:2546–2559.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yoshida T, Imai T, Takagi S, Nishimura M,
Ishikawa I, Yaoi T and Yoshie O: Structure and expression of two
highly related genes encoding SCM-1/human lymphotactin. FEBS Lett.
395:82–88. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lei Y and Takahama Y: XCL1 and XCR1 in the
immune system. Microbes Infect. 14:262–267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yamazaki C, Sugiyama M, Ohta T, Hemmi H,
Hamada E, Sasaki I, Fukuda Y, Yano T, Nobuoka M, Hirashima T, et
al: Critical roles of a dendritic cell subset expressing a
chemokine receptor, XCR1. J Immunol. 190:6071–6082. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Khurram SA, Whawell SA, Bingle L, Murdoch
C, McCabe BM and Farthing PM: Functional expression of the
chemokine receptor XCR1 on oral epithelial cells. J Pathol.
221:153–163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gantsev SK, Umezawa K, Islamgulov DV,
Khusnutdinova EK, Ishmuratova RS, Frolova VY and Kzyrgalin SR: The
role of inflammatory chemokines in lymphoid neoorganogenesis in
breast cancer. Biomed Pharmacother. 67:363–366. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim M, Rooper L, Xie J, Rayahin J,
Burdette JE, Kajdacsy-Balla AA and Barbolina MV: The lymphotactin
receptor is expressed in epithelial ovarian carcinoma and
contributes to cell migration and proliferation. Mol Cancer Res.
10:1419–1429. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang T, Han S, Wu Z, Han Z, Yan W, Liu T,
Wei H, Song D, Zhou W, Yang X and Xiao J: XCR1 promotes cell growth
and migration and is correlated with bone metastasis in non-small
cell lung cancer. Biochem Biophys Res Commun. 464:635–641. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yanru W, Zhenyu B, Zhengchuan N, Qi Q,
Chunmin L and Weiqiang Y: Transcriptomic analyses of chemokines
reveal that down-regulation of XCR1 is associated with advanced
hepatocellular carcinoma. Biochem Biophys Res Commun.
496:1314–1321. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li X: The inducers of immunogenic cell
death for tumor immunotherapy. Tumori. 104:1–8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mizumoto Y, Hemmi H, Katsuda M, Miyazawa
M, Kitahata Y, Miyamoto A, Nakamori M, Ojima T, Matsuda K, Nakamura
M, et al: Anticancer effects of chemokine-directed antigen delivery
to a cross-presenting dendritic cell subset with immune checkpoint
blockade. Br J Cancer. 122:1185–1193. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Spranger S and Gajewski TF: A new paradigm
for tumor immune escape: β-catenin-driven immune exclusion. J
Immunother Cancer. 3:432015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Botelho NK, Tschumi BO, Hubbell JA, Swartz
MA, Donda A and Romero P: Combination of synthetic long peptides
and XCL1 fusion proteins results in superior tumor control. Front
Immunol. 10:2942019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen K, Wu Z, Zhao H, Wang Y, Ge Y, Wang
D, Li Z, An C, Liu Y, Wang F, et al: XCL1/Glypican-3 fusion gene
immunization generates potent antitumor cellular immunity and
enhances Anti-PD-1 efficacy. Cancer Immunol Res. 8:81–93. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Audsley KM, McDonnell AM and Waithman J:
Cross-presenting XCR1+ dendritic cells as targets for
cancer immunotherapy. Cells. 9:5652020. View Article : Google Scholar
|
|
55
|
Wylie B, Read J, Buzzai AC, Wagner T, Troy
N, Syn G, Stone SR, Foley B, Bosco A, Cruickshank MN, et al:
CD8+XCR1neg dendritic cells express high
levels of toll-like receptor 5 and a unique complement of endocytic
receptors. Front Immunol. 9:29902019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Qin CJ, Zhao LH, Zhou X, Zhang HL, Wen W,
Tang L, Zeng M, Wang MD, Fu GB, Huang S, et al: Inhibition of
dipeptidyl peptidase IV prevents high fat diet-induced liver cancer
angiogenesis by downregulating chemokine ligand 2. Cancer Lett.
420:26–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Giles AJ, Reid CM, Evans JD, Murgai M,
Vicioso Y, Highfill SL, Kasai M, Vahdat L, Mackall CL, Lyden D, et
al: Activation of hematopoietic stem/progenitor cells promotes
immunosuppression within the pre-metastatic niche. Cancer Res.
76:1335–1347. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chang AL, Miska J, Wainwright DA, Dey M,
Rivetta CV, Yu D, Kanojia D, Pituch KC, Qiao J, Pytel P, et al:
CCL2 produced by the glioma microenvironment is essential for the
recruitment of regulatory T cells and myeloid-derived suppressor
cells. Cancer Res. 76:5671–5682. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Qi S, Perrino S, Miao X, Lamarche-Vane N
and Brodt P: The chemokine CCL7 regulates invadopodia maturation
and MMP-9 mediated collagen degradation in liver-metastatic
carcinoma cells. Cancer Lett. 483:98–113. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wu Q, Chen JX, Chen Y, Cai LL, Wang XZ,
Guo WH and Zheng JF: The chemokine receptor CCR10 promotes
inflammation-driven hepatocarcinogenesis via PI3K/Akt pathway
activation. Cell Death Dis. 9:2322018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hippe A, Braun SA, Oláh P, Gerber PA,
Schorr A, Seeliger S, Holtz S, Jannasch K, Pivarcsi A, Buhren B, et
al: EGFR/Ras-induced CCL20 production modulates the tumour
microenvironment. Br J Cancer. 123:942–954. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Du D, Liu Y, Qian H, Zhang B, Tang X,
Zhang T and Liu W: The effects of the CCR6/CCL20 biological axis on
the invasion and metastasis of hepatocellular carcinoma. Int J Mol
Sci. 15:6441–6452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Huang F and Geng XP: Chemokines and
hepatocellular carcinoma. World J Gastroenterol. 16:1832–1836.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tan H, Wang S and Zhao L: A
tumour-promoting role of Th9 cells in hepatocellular carcinoma
through CCL20 and STAT3 pathways. Clin Exp Pharmacol Physiol.
44:213–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Facciabene A, Peng X, Hagemann IS, Balint
K, Barchetti A, Wang LP, Gimotty PA, Gilks CB, Lal P, Zhang L and
Coukos G: Tumour hypoxia promotes tolerance and angiogenesis via
CCL28 and T(reg) cells. Nature. 475:226–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gao Y, Zhou Z, Lu S, Huang X, Zhang C,
Jiang R, Yao A, Sun B and Wang X: Chemokine CCL15 mediates
migration of human bone marrow-derived mesenchymal stem cells
toward hepatocellular carcinoma. Stem Cells. 34:1112–1122. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Singh SK, Mishra MK, Eltoum IA, Bae S,
Lillard JW Jr and Singh R: CCR5/CCL5 axis interaction promotes
migratory and invasiveness of pancreatic cancer cells. Sci Rep.
8:13232018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Singh SK, Mishra MK, Rivers BM, Gordetsky
JB, Bae S and Singh R: Biological and clinical significance of the
CCR5/CCL5 axis in hepatocellular carcinoma. Cancers (Basel).
12:8832020. View Article : Google Scholar
|
|
69
|
Sasaki R, Devhare PB, Steele R, Ray R and
Ray RB: Hepatitis C virus-induced CCL5 secretion from macrophages
activates hepatic stellate cells. Hepatology. 66:746–757. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
González-Martín A, Mira E and Mañes S:
CCR5 in cancer immunotherapy: More than an ‘attractive’ receptor
for T cells. Oncoimmunology. 1:106–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wang T, Zhan Q, Peng X, Qiu Z and Zhao T:
CCL2 influences the sensitivity of lung cancer A549 cells to
docetaxel. Oncol Lett. 16:1267–1274. 2018.PubMed/NCBI
|
|
72
|
Pasquier J, Gosset M, Geyl C,
Hoarau-Véchot J, Chevrot A, Pocard M, Mirshahi M, Lis R, Rafii A
and Touboul C: CCL2/CCL5 secreted by the stroma induce IL-6/PYK2
dependent chemoresistance in ovarian cancer. Mol Cancer. 17:472018.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Su S, Sun X, Zhang Q, Zhang Z and Chen J:
CCL20 promotes ovarian cancer chemotherapy resistance by regulating
ABCB1 expression. Cell Struct Funct. 44:21–28. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Vaquero J, Briz O, Herraez E, Muntané J
and Marin JJ: Activation of the nuclear receptor FXR enhances
hepatocyte chemoprotection and liver tumor chemoresistance against
genotoxic compounds. Biochim Biophys Acta. 1833:2212–2219. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gu Y, Li X, Bi Y, Zheng Y, Wang J, Li X,
Huang Z, Chen L and Huang Y and Huang Y: CCL14 is a prognostic
biomarker and correlates with immune infiltrates in hepatocellular
carcinoma. Aging (Albany NY). 12:784–807. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rodríguez-Perea AL, Rojas M and
Velilla-Hernández PA: High concentrations of atorvastatin reduce
in-vitro function of conventional T and regulatory T cells. Clin
Exp Immunol. 196:237–248. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhu M, Xu W, Wei C, Huang J, Xu J, Zhang
Y, Zhao Y, Chen J, Dong S, Liu B and Liang C: CCL14 serves as a
novel prognostic factor and tumor suppressor of HCC by modulating
cell cycle and promoting apoptosis. Cell Death Dis. 10:7962019.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang X, Wan JX, Ke ZP, Wang F, Chai HX
and Liu JQ: TMEM88, CCL14 and CLEC3B as prognostic biomarkers for
prognosis and palindromia of human hepatocellular carcinoma. Tumour
Biol. 39:10104283177089002017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wilson GC, Kuboki S, Freeman CM, Nojima H,
Schuster RM, Edwards MJ and Lentsch AB: CXC chemokines function as
a rheostat for hepatocyte proliferation and liver regeneration.
PLoS One. 10:e01200922015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Vandercappellen J, Van Damme J and Struyf
S: The role of CXC chemokines and their receptors in cancer. Cancer
Lett. 267:226–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu G, Yang ZF, Zhou PY, Zhou C, Guan RY,
Sun BY, Fan J, Zhou J, Yi Y and Qiu SJ: ROR-α-1 inhibits the
proliferation, invasion, and migration of hepatocellular carcinoma
MHCC97H via downregulation of chemokine CXCL5. Cytokine.
129:1550042020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhou SL, Zhou ZJ, Hu ZQ, Li X, Huang XW,
Wang Z, Fan J, Dai Z and Zhou J: CXCR2/CXCL5 axis contributes to
epithelial-mesenchymal transition of HCC cells through activating
PI3K/Akt/GSK-3β/Snail signaling. Cancer Lettr. 358:124–135. 2015.
View Article : Google Scholar
|
|
83
|
Li XP, Yang XY, Biskup E, Zhou J, Li HL,
Wu YF, Chen ML and Xu F: Co-expression of CXCL8 and HIF-1α is
associated with metastasis and poor prognosis in hepatocellular
carcinoma. Oncotarget. 6:22880–22889. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Yamamoto M, Kikuchi H, Ohta M, Kawabata T,
Hiramatsu Y, Kondo K, Baba M, Kamiya K, Tanaka T, Kitagawa M and
Konno H: TSU68 prevents liver metastasis of colon cancer xenografts
by modulating the premetastatic niche. Cancer Res. 68:9754–9762.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Van den Eynden GG, Majeed AW, Illemann M,
Vermeulen PB, Bird NC, Høyer-Hansen G, Eefsen RL, Reynolds AR and
Brodt P: The multifaceted role of the microenvironment in liver
metastasis: Biology and clinical implications. Cancer Res.
73:2031–2043. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Li L, Zhu YH, Li Y and Guan XY:
Identification of chemokine CXCL10 in tumor microenvironment by
antibody array as a prognostic marker in hepatocellular carcinoma.
Neoplasma. 64:778–786. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li B, Su H, Cao J and Zhang L: CXCL13
rather than IL-31 is a potential indicator in patients with
hepatocellular carcinoma. Cytokine. 89:91–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Song X, Wang Z, Jin Y, Wang Y and Duan W:
Loss of miR-532-5p in vitro promotes cell proliferation and
metastasis by influencing CXCL2 expression in HCC. Am J Transl Res.
7:2254–2261. 2015.PubMed/NCBI
|
|
89
|
Ding J, Xu K, Zhang J, Lin B, Wang Y, Yin
S, Xie H, Zhou L and Zheng S: Overexpression of CXCL2 inhibits cell
proliferation and promotes apoptosis in hepatocellular carcinoma.
BMB Rep. 51:630–635. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Subat S, Mogushi K, Yasen M, Kohda T,
Ishikawa Y and Tanaka H: Identification of genes and pathways,
including the CXCL2 axis, altered by DNA methylation in
hepatocellular carcinoma. J Cancer Res Clin Oncol. 145:675–684.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Shi A, Shi H, Dong L, Xu S, Jia M, Guo X
and Wang T: CXCR7 as a chemokine receptor for SDF-1 promotes
gastric cancer progression via MAPK pathways. Scand J
Gastroenterol. 52:745–753. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li D, Qu C, Ning Z, Wang H, Zang K, Zhuang
L, Chen L, Wang P and Meng Z: Radiation promotes
epithelial-to-mesenchymal transition and invasion of pancreatic
cancer cell by activating carcinoma-associated fibroblasts. Am J
Cancer Res. 6:2192–2206. 2016.PubMed/NCBI
|
|
93
|
Croker AK and Allan AL: Cancer stem cells:
Implications for the progression and treatment of metastatic
disease. J Cell Mol Med. 12:374–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Jahanban-Esfahlan R, de la Guardia M,
Ahmadi D and Yousefi B: Modulating tumor hypoxia by nanomedicine
for effective cancer therapy. J Cell Physiol. 233:2019–2031. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Teng F, Tian WY, Wang YM, Zhang YF, Guo F,
Zhao J, Gao C and Xue FX: Cancer-associated fibroblasts promote the
progression of endometrial cancer via the SDF-1/CXCR4 axis. J
Hematol Oncol. 9:82016. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wang C, Wang MD, Cheng P, Huang H, Dong W,
Zhang WW, Li PP, Lin C, Pan ZY, Wu MC and Zhou WP: Hepatitis B
virus X protein promotes the stem-like properties of
OV6+ cancer cells in hepatocellular carcinoma. Cell
Death Dis. 8:e25602017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kaemmerer D, Schindler R, Mußbach F,
Dahmen U, Altendorf-Hofmann A, Dirsch O, Sänger J, Schulz S and
Lupp A: Somatostatin and CXCR4 chemokine receptor expression in
hepatocellular and cholangiocellular carcinomas: Tumor capillaries
as promising targets. BMC Cancer. 17:8962017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Li H, Yang W, Chen PW, Alizadeh H and
Niederkorn JY: Inhibition of chemokine receptor expression on uveal
melanomas by CXCR4 siRNA and its effect on uveal melanoma liver
metastases. Invest Ophthalmol Vis Sci. 50:5522–5528. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Deol A, Abrams J, Masood A, Al-Kadhimi Z,
Abidi MH, Ayash L, Lum LG, Ratanatharathorn V and Uberti JP:
Long-term follow up of patients proceeding to transplant using
plerixafor mobilized stem cells and incidence of secondary
myelodysplastic syndrome/AML. Bone Marrow Transplant. 48:1112–1116.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Collins PJ, McCully ML, Martínez-Muñoz L,
Santiago C, Wheeldon J, Caucheteux S, Thelen S, Cecchinato V,
Laufer JM, Purvanov V, et al: Epithelial chemokine CXCL14
synergizes with CXCL12 via allosteric modulation of CXCR4. FASEB J.
31:3084–3097. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Chen Y, Teng F, Wang G and Nie Z:
Overexpression of CXCR7 induces angiogenic capacity of human
hepatocellular carcinoma cells via the AKT signaling pathway. Oncol
Rep. 36:2275–2281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Lin L, Han MM, Wang F, Xu LL, Yu HX and
Yang PY: CXCR7 stimulates MAPK signaling to regulate hepatocellular
carcinoma progression. Cell Death Dis. 5:e14882014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Billottet C, Quemener C and Bikfalvi A:
CXCR3, a double-edged sword in tumor progression and angiogenesis.
Biochim Biophys Acta. 1836:287–295. 2013.PubMed/NCBI
|
|
104
|
Ma B, Khazali A and Wells A: CXCR3 in
carcinoma progression. Histol Histopathol. 30:781–792.
2015.PubMed/NCBI
|
|
105
|
Gao Q, Zhao YJ, Wang XY, Qiu SJ, Shi YH,
Sun J, Yi Y, Shi JY, Shi GM, Ding ZB, et al: CXCR6 upregulation
contributes to a proinflammatory tumor microenvironment that drives
metastasis and poor patient outcomes in hepatocellular carcinoma.
Cancer Res. 72:3546–3556. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wu Y, Tian L, Xu Y, Zhang M, Xiang S, Zhao
J and Wang Z: CXCR7 silencing inhibits the migration and invasion
of human tumor endothelial cells derived from hepatocellular
carcinoma by suppressing STAT3. Mol Med Rep. 18:1644–1650.
2018.PubMed/NCBI
|
|
107
|
Xu Y, Fang F, Jiao H, Zheng X, Huang L, Yi
X and Zhao W: Activated hepatic stellate cells regulate MDSC
migration through the SDF-1/CXCR4 axis in an orthotopic mouse model
of hepatocellular carcinoma. Cancer Immunol Immunother.
68:1959–1969. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tian H, Huang P, Zhao Z, Tang W and Xia J:
HIF-1α plays a role in the chemotactic migration of hepatocarcinoma
cells through the modulation of CXCL6 expression. Cell Physiol
Biochem. 34:1536–1546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Shen H, Yao X, Li H, Li X, Zhang T, Sun Q,
Ji C and Chen G: Role of exosomes derived from miR-133b modified
MSCs in an experimental rat model of intracerebral hemorrhage. J
Mol Neurosci. 64:421–430. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Halvaei S, Daryani S, Eslami-S Z, Samadi
T, Jafarbeik-Iravani N, Bakhshayesh TO, Majidzadeh-A K and Esmaeili
R: Exosomes in cancer liquid biopsy: A focus on breast cancer. Mol
Ther Nucleic Acids. 10:131–141. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Anel A, Gallego-Lleyda A, de Miguel D,
Naval J and Martínez-Lostao L: Role of exosomes in the regulation
of T-cell mediated immune responses and in autoimmune disease.
Cells. 8:1542019. View Article : Google Scholar
|
|
112
|
Almeida VH, Rondon AMR, Gomes T and
Monteiro RQ: Novel aspects of extracellular vesicles as mediators
of cancer-associated thrombosis. Cells. 8:7162019. View Article : Google Scholar
|
|
113
|
Rao PSS, O'Connell K and Finnerty TK:
Potential role of extracellular vesicles in the pathophysiology of
drug addiction. Mol Neurobiol. 55:6906–6913. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kohama I, Kosaka N, Chikuda H and Ochiya
T: An insight into the roles of MicroRNAs and exosomes in sarcoma.
Cancers (Basel). 11:4282019. View Article : Google Scholar
|
|
115
|
Mashouri L, Yousefi H, Aref AR, Ahadi AM,
Molaei F and Alahari SK: Exosomes: Composition, biogenesis, and
mechanisms in cancer metastasis and drug resistance. Mol Cancer.
18:752019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Sun JF, Zhang D, Gao CJ, Zhang YW and Dai
QS: Exosome-mediated MiR-155 transfer contributes to hepatocellular
carcinoma cell proliferation by targeting PTEN. Med Sci Monit Basic
Res. 25:218–228. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lee JY and Kim HS: Extracellular vesicles
in neurodegenerative diseases: A double-edged sword. Tissue Eng
Regen Med. 14:667–678. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Johnsen KB, Gudbergsson JM, Skov MN,
Pilgaard L, Moos T and Duroux M: A comprehensive overview of
exosomes as drug delivery vehicles-endogenous nanocarriers for
targeted cancer therapy. Biochim Biophys Acta. 1846:75–87.
2014.PubMed/NCBI
|
|
119
|
Yi YW, Lee JH, Kim SY, Pack CG, Ha DH,
Park SR, Youn J and Cho BS: Advances in analysis of biodistribution
of exosomes by molecular imaging. Int J Mol Sci. 21:6652020.
View Article : Google Scholar
|
|
120
|
Wu P, Zhang B, Ocansey DKW, Xu W and Qian
H: Extracellular vesicles: A bright star of nanomedicine.
Biomaterials. 6:1204672020. View Article : Google Scholar
|
|
121
|
Kim MS, Haney MJ, Zhao Y, Mahajan V,
Deygen I, Klyachko NL, Inskoe E, Piroyan A, Sokolsky M, Okolie O,
et al: Development of exosome-encapsulated paclitaxel to overcome
MDR in cancer cells. Nanomedicine. 12:655–664. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Si Y, Kim S, Zhang E, Tang Y,
Jaskula-Sztul R, Markert JM, Chen H, Zhou L and Liu XM: Targeted
exosomes for drug delivery: Biomanufacturing, surface tagging, and
validation. Biotechnol J. 15:e19001632020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Prada I and Meldolesi J: Binding and
fusion of extracellular vesicles to the plasma membrane of their
cell targets. Int J Mol Sci. 17:12962016. View Article : Google Scholar
|
|
124
|
Nakase I and Futaki S: Combined treatment
with a pH-sensitive fusogenic peptide and cationic lipids achieves
enhanced cytosolic delivery of exosomes. Sci Rep. 5:101122015.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Kooijmans SA, Aleza CG, Roffler SR, van
Solinge WW, Vader P and Schiffelers RM: Display of GPI-anchored
anti-EGFR nanobodies on extracellular vesicles promotes tumour cell
targeting. J Extracell Vesicles. 5:310532016. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hashimoto K, Ochi H, Sunamura S, Kosaka N,
Mabuchi Y, Fukuda T, Yao K, Kanda H, Ae K, Okawa A, et al:
Cancer-secreted hsa-miR-940 induces an osteoblastic phenotype in
the bone metastatic microenvironment via targeting ARHGAP1 and
FAM134A. Proc Natl Acad Sci USA. 115:2204–2209. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Luis-Ravelo D, Antón I, Zandueta C,
Valencia K, Ormazábal C, Martínez-Canarias S, Guruceaga E, Perurena
N, Vicent S, De Las Rivas J and Lecanda F: A gene signature of bone
metastatic colonization sensitizes for tumor-induced osteolysis and
predicts survival in lung cancer. Oncogene. 33:5090–5099. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Li H, Yang C, Shi Y and Zhao L: Exosomes
derived from siRNA against GRP78 modified bone-marrow-derived
mesenchymal stem cells suppress Sorafenib resistance in
hepatocellular carcinoma. J Nanobiotechnology. 16:1032018.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Viñas JL, Spence M, Gutsol A, Knoll W,
Burger D, Zimpelmann J, Allan DS and Burns KD: Receptor-ligand
interaction mediates targeting of endothelial colony forming
cell-derived exosomes to the kidney after ischemic injury. Sci Rep.
8:163202018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Ciullo A, Biemmi V, Milano G, Bolis S,
Cervio E, Fertig ET, Gherghiceanu M, Moccetti T, Camici GG,
Vassalli G and Barile L: Exosomal expression of CXCR4 targets
cardioprotective vesicles to myocardial infarction and improves
outcome after systemic administration. Int J Mol Sci. 20:4682019.
View Article : Google Scholar
|
|
132
|
Wei G, Jie Y, Haibo L, Chaoneng W, Dong H,
Jianbing Z, Junjie G, Leilei M, Hongtao S, Yunzeng Z and Junbo G:
Dendritic cells derived exosomes migration to spleen and induction
of inflammation are regulated by CCR7. Sci Rep. 7:429962017.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Longo V, Gnoni A, Casadei Gardini A,
Pisconti S, Licchetta A, Scartozzi M, Memeo R, Palmieri VO, Aprile
G, Santini D, et al: Immunotherapeutic approaches for
hepatocellular carcinoma. Oncotarget. 8:33897–33910. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Liu YC, Yeh CT and Lin KH: Cancer stem
cell functions in hepatocellular carcinoma and comprehensive
therapeutic strategies. Cells. 9:13312020. View Article : Google Scholar
|
|
135
|
Liu LZ, Zhang Z, Zheng BH, Shi Y, Duan M,
Ma LJ, Wang ZC, Dong LQ, Dong PP, Shi JY, et al: CCL15 recruits
suppressive monocytes to facilitate Immune escape and disease
progression in hepatocellular carcinoma. Hepatology. 69:143–159.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Sun F, Wang J, Sun Q, Li F, Gao H, Xu L,
Zhang J, Sun X, Tian Y, Zhao Q, et al: Interleukin-8 promotes
integrin β3 upregulation and cell invasion through PI3K/Akt pathway
in hepatocellular carcinoma. J Exp Clin Cancer Res. 38:4492019.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Li L, Xu L, Yan J, Zhen ZJ, Ji Y, Liu CQ,
Lau WY, Zheng L and Xu J: CXCR2-CXCL1 axis is correlated with
neutrophil infiltration and predicts a poor prognosis in
hepatocellular carcinoma. J Exp Clin Cancer Res. 34:1292015.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Lu Y, Li S, Ma L, Li Y, Zhang X, Peng Q,
Mo C, Huang L, Qin X and Liu Y: Type conversion of secretomes in a
3D TAM2 and HCC cell co-culture system and functional importance of
CXCL2 in HCC. Sci Rep. 6:245582016. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Zhang L, Zhang L, Li H, Ge C, Zhao F, Tian
H, Chen T, Jiang G, Xie H, Cui Y, et al: CXCL3 contributes to
CD133+ CSCs maintenance and forms a positive feedback
regulation loop with CD133 in HCC via Erk1/2 phosphorylation. Sci
Rep. 6:274262016. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Zhou SL, Dai Z, Zhou ZJ, Wang XY, Yang GH,
Wang Z, Huang XW, Fan J and Zhou J: Overexpression of CXCL5
mediates neutrophil infiltration and indicates poor prognosis for
hepatocellular carcinoma. Hepatology. 56:2242–2254. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ding Q, Xia Y, Ding S, Lu P, Sun L and Liu
M: An alternatively spliced variant of CXCR3 mediates the
metastasis of CD133+ liver cancer cells induced by CXCL9.
Oncotarget. 7:14405–14414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Ouyang Y, Liu K, Hao M, Zheng R, Zhang C,
Wu Y, Zhang X, Li N, Zheng J and Chen D: Radiofrequency
ablation-increased CXCL10 is associated with earlier recurrence of
hepatocellular carcinoma by promoting stemness. Tumour Biol.
37:3697–3704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Ling CC, Ng KT, Shao Y, Geng W, Xiao JW,
Liu H, Li CX, Liu XB, Ma YY, Yeung WH, et al: Post-transplant
endothelial progenitor cell mobilization via CXCL10/CXCR3 signaling
promotes liver tumor growth. J Hepatol. 60:103–109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Zhang Y, Zhao W, Li S, Lv M, Yang X, Li M
and Zhang Z: CXCL11 promotes self-renewal and tumorigenicity of
α2δ1+ liver tumor-initiating cells through CXCR3/ERK1/2
signaling. Cancer Lett. 449:163–171. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Li C, Kang D, Sun X, Liu Y, Wang J and Gao
P: The effect of C-X-C motif chemokine 13 on hepatocellular
carcinoma associates with Wnt signaling. Biomed Res Int.
2015:3454132015.PubMed/NCBI
|
|
146
|
Deng L, Chen N, Li Y, Zheng H and Lei Q:
CXCR6/CXCL16 functions as a regulator in metastasis and progression
of cancer. Biochim Biophys Acta. 1806:42–49. 2010.PubMed/NCBI
|
|
147
|
Yuan Y, Liu J, Liu Z, He Y, Zhang Z, Jiang
C and Qian Q: Chemokine CCL3 facilitates the migration of hepatoma
cells by changing the concentration intracellular Ca. Hepatol Res.
40:424–431. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Bai H, Weng Y, Bai S, Jiang Y, Li B, He F,
Zhang R, Yan S, Deng F, Wang J and Shi Q: CCL5 secreted from bone
marrow stromal cells stimulates the migration and invasion of Huh7
hepatocellular carcinoma cells via the PI3K-Akt pathway. Int J
Oncol. 45:333–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Liao WC, Yen HR, Liao CK, Tseng TJ, Lan CT
and Liu CH: DSE regulates the malignant characters of
hepatocellular carcinoma cells by modulating CCL5/CCR1 axis. Am J
Cancer Res. 9:347–362. 2019.PubMed/NCBI
|
|
150
|
Li Y, Wu J and Zhang P: CCL15/CCR1 axis is
involved in hepatocellular carcinoma cells migration and invasion.
Tumour Biol. 37:4501–4507. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Lu J, Yang Y, Yu P, Tao H, Lu X, Wang L,
Liu D, Chen Y and Chen C: Bioinformatics analysis and significance
of expression of CC chemokine ligand 23 (CCL23) in hepatocellular
carcinoma. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 35:903–909.
2019.(In Chinese). PubMed/NCBI
|
|
152
|
Zhuang H, Cao G, Kou C and Liu T:
CCL2/CCR2 axis induces hepatocellular carcinoma invasion and
epithelial-mesenchymal transition in vitro through
activation of the Hedgehog pathway. Oncol Rep. 39:21–30.
2018.PubMed/NCBI
|
|
153
|
Jin L, Liu WR, Tian MX, Jiang XF, Wang H,
Zhou PY, Ding ZB, Peng YF, Dai Z, Qiu SJ, et al: CCL24 contributes
to HCC malignancy via RhoB-VEGFA-VEGFR2 angiogenesis pathway and
indicates poor prognosis. Oncotarget. 8:5135–5148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Zhu F, Li X, Chen S, Zeng Q, Zhao Y and
Luo F: Tumor-associated macrophage or chemokine ligand CCL17
positively regulates the tumorigenesis of hepatocellular carcinoma.
Med Oncol. 33:172016. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Wiedemann GM, Röhrle N, Makeschin MC,
Fesseler J, Endres S, Mayr D and Anz D: Peritumoural CCL1 and CCL22
expressing cells in hepatocellular carcinomas shape the tumour
immune infiltrate. Pathology. 51:586–592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Cheng X, Wu H, Jin ZJ, Ma D, Yuen S, Jing
XQ, Shi MM, Shen BY, Peng CH, Zhao R and Qiu WH: Up-regulation of
chemokine receptor CCR4 is associated with Human Hepatocellular
Carcinoma malignant behavior. Sci Rep. 7:123622017. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Mukaida N, Sasaki SI and Baba T: CCL4
signaling in the tumor microenvironment. Adv Exp Med Biol.
1231:23–32. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Luo KQ, Shi YN and Peng JC: The effect of
chemokine CC motif ligand 19 on the proliferation and migration of
hepatocellular carcinoma. Tumour Biol. 35:12575–12581. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Yang L, Chang Y and Cao P: CCR7
preservation via histone deacetylase inhibition promotes
epithelial-mesenchymal transition of hepatocellular carcinoma
cells. Exp Cell Res. 371:231–237. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Hou PP, Luo LJ, Chen HZ, Chen QT, Bian XL,
Wu SF, Zhou JX, Zhao WX, Liu JM, Wang XM, et al: Ectosomal PKM2
promotes HCC by inducing macrophage differentiation and remodeling
the tumor microenvironment. Mol Cell. 78:1192–1206.e10. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Xu B, Deng C, Wu X, Ji T, Zhao L, Han Y,
Yang W, Qi Y, Wang Z, Yang Z and Yang Y: CCR9 and CCL25: A review
of their roles in tumor promotion. J Cell Physiol. 235:9121–9132.
2020. View Article : Google Scholar : PubMed/NCBI
|