Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumors of the digestive tract worldwide, as the third
most frequent malignancy in men and the second in women, and the
fourth leading cause of cancer-related death (1). The incidence of CRC is increasing in
younger age groups (2). Current
treatment strategies for CRC include surgery, radiation and
chemotherapy; however, the 5-year survival rate of patients with
metastatic CRC is ~10% (3).
Chemotherapy serves an essential role in the treatment of CRC, but
CRC is heterogeneous in response to adjuvant chemotherapy (4). Therefore, developing more effective
drugs for the treatment of CRC is important.
Lycorine is a natural Amaryllidaceae alkaloid that
is extracted from medicinal plants of the Amaryllidaceae family
(5). In addition to its analgesic,
antibacterial, antifungal, antimalarial and antiviral effects,
lycorine also displays antitumor effects against various types of
cancer, including multiple myeloma (6), bladder cancer (7) and breast cancer (8), with an IC50 value <10
µmol/l. Moreover, previous studies have reported that lycorine may
display relatively lower toxicity against healthy cells (7–9). For
example, the IC50 of lycorine in human normal
fibroblasts (Wi38, WS1, NHDF cell lines) was >100 µmol/l
(9). The aforementioned studies
suggest that lycorine may serve as a potentially effective and safe
drug candidate for cancer treatment.
In the present study, the effect of lycorine on
human CRC cell proliferation, migration, invasion, apoptosis and
cell cycle distribution was investigated, and the underlying
molecular mechanism was also explored. The results of the present
study indicated that due to in vitro cytostatic effects,
lycorine might serve as a potential therapeutic for CRC, and the
underlying mechanism might be associated with activation of ROS/p38
and AKT signaling, although further investigation is required.
Materials and methods
Cell lines and cell culture
Human CRC cell lines (LoVo, HCT116 and SW480) were
provided by the Key Laboratory of Clinical Laboratory Medical
Diagnostics (Ministry of Education, Chongqing Medical University).
Cells were cultured in DMEM (HyClone; GE Healthcare Life Sciences)
supplemented with 10% FBS (Shanghai ExCell Biology, Inc.) and 1%
penicillin/streptomycin (HyClone; GE Healthcare Life Sciences) at
37°C with 5% CO2. Lycorine (purity ≥98%; Chengdu
Ruifensi Biotechnology Co., Ltd.) was dissolved in DMSO
(Sigma-Aldrich; Merck KGaA) to a final concentration of 20 mM and
stored at −80°C.
Crystal violet staining
Cells were seeded into 24-well plates and cultured
overnight. At 50% confluence, cells were treated with different
concentrations (0, 1, 2, 4 or 8 µmol/l) of lycorine at 37°C for 24,
48 or 72 h. The control group was untreated (0 µmol/l lycorine) and
a 0.4% DMSO group (treated at 37°C for 24, 48 or 72 h) was also
established. At the indicated time point, cells were fixed with 4%
paraformaldehyde at 37°C for 20 min and stained with crystal violet
(Beyotime Institute of Biotechnology) at room temperature for 5
min. Stained cells were visualized using an Epson Perfection V200
Photo (Epson).
MTT assay
Cells were seeded (3×103 cells/well) into
96-well plates overnight. Subsequently, cells were treated with
different concentrations (0, 1, 2, 4 or 8 µmol/l) of lycorine at
37°C for 24, 48 or 72 h. At the indicated time point, cells were
incubated with 5 mg/ml MTT solution (Sigma-Aldrich; Merck KGaA) at
37°C for 4 h. The formazan crystals were dissolved with DMSO (150
µl/well). The absorbance was measured at a wavelength of 490 nm
using a spectrophotometer (Gene Company, Ltd.). Cell viability (%)
was calculated using the following formula: (Dtreatment
group/Dcontrol group) ×100%, where D
represents optical density.
Wound healing assay
Cells were seeded into 6-well plates. At 80–90%
confluence, the cell monolayer was scratched with a 10 µl pipette
tip and treated with different concentrations (0, 1, 2, 4 or 8
µmol/l) of lycorine and cultured in DMEM supplemented with 5% FBS
for 0, 12 or 24 h at 37°C. The wounds were observed in three
randomly selected fields of view using a light microscope
(magnification, ×100). The wound healing rate (%) was calculated
using the following formula: [(0 h wound width −12 or 24 h wound
width)/0 h wound width] ×100.
Transwell assay
For cell invasion, the 24-well upper chamber (EMD
Millipore) was precoated with Matrigel at 37°C for 60 min (BD
Biosciences). Subsequently, cells were seeded (5×104
cells/well) into the upper chamber with serum-free DMEM containing
different concentrations (0, 1, 2, 4 or 8 µmol/l) of lycorine. The
lower chamber was filled with DMEM supplemented with 10% FBS.
Following incubation at 37°C for 24 h, non-invading cells on the
upper surface of the membrane were removed. Invading cells were
fixed with 4% paraformaldehyde at 37°C for 20 min and stained with
crystal violet at room temperature for 5 min. Stained cells were
visualized using a light microscope (magnification, ×100). To
assess cell migration, the aforementioned protocol was performed,
but the Transwell chambers were not precoated with Matrigel.
Hoechst 33258 staining
Cells were seeded into 24-well plates. At 50%
confluence, cells were treated with different concentrations (0, 1,
2, 4 or 8 µmol/l) of lycorine at 37°C for 24 h. Following fixation
with 4% paraformaldehyde at 37°C for 20 min, cells were stained
with Hoechst 33258 solution (Beijing Solarbio Science &
Technology Co., Ltd.) for 5 min at room temperature. Alterations in
nuclear morphology were visualized using a fluorescence microscope
(magnification, ×200). When apoptosis occurs, cell nuclei display
enhanced fluorescence intensity due to nuclear fragmentation and
chromatin condensation.
Flow cytometry
Cells were plated into 60-mm dishes. At 50%
confluence, cells were treated with different concentrations (0, 1,
2, 4 or 8 µmol/l) of lycorine at 37°C for 24 h. To analyse cell
apoptosis including early and late apoptosis, cells were harvested,
washed twice with PBS, stained using the Annexin V-FITC/PI kit
(Beijing Solarbio Science & Technology Co., Ltd.), according to
the manufacturer's protocol, and analyzed using a flow cytometer
(BD Biosciences). To assess cell cycle distribution, cells were
re-suspended in 70% ethanol overnight at 4°C for fixation and
permeabilisation and stained with PI/RNase Staining Buffer kit (BD
Biosciences) according to the manufacturer's protocol. Both cell
apoptosis and cell cycle distribution were detected using a
CytoFLEX flow cytometer (Beckman Coulter, Inc.) and analysed with
FlowJo software (version 10.4.0; BD Biosciences).
Intracellular reactive oxygen species
(ROS) measurement
Cells were seeded in 6-well plates. At 50%
confluence, cells were treated with 8 µmol/l lycorine at 37°C for 4
or 24 h. Subsequently, cells were incubated with
2′,7′-dichlorofluorescein-diacetate (DCFH-DA; 10 µmol/l) reagent
(Beyotime Institute of Biotechnology) in serum-free DMEM for 30 min
at 37°C. Following washing three times with PBS, the fluorescence
intensity was measured at an excitation wavelength of 488 nm and an
emission wavelength of 525 nm using a fluorescence plate reader
(BioTek Instruments, Inc.). Moreover, following fixation with 4%
paraformaldehyde at 37°C for 15 min, cells were stained with DAPI
at room temperature for 10 min. Intracellular ROS accumulation,
which was assessed using DCFH-DA as aforementioned and nuclear DAPI
staining, was visualized using a confocal microscope
(magnification, ×200).
Western blotting
Cells were seeded into 10-cm dishes. At 50%
confluence, cells were treated with different concentrations of
lycorine (1, 4 or 8 µmol/l) at 37°C for 24 h. Total protein was
extracted from cells using RIPA lysate (Beyotime Institute of
Biotechnology) supplemented with 1% protease inhibitors and
phosphatase inhibitors (Roche Applied Science). Protein
concentrations were determined using a BCA assay (Beyotime
Institute of Biotechnology). Proteins (35 µg) were separated via
10% SDS-PAGE (Beyotime Institute of Biotechnology) and subsequently
transferred to PVDF membranes (EMD Millipore). Following blocking
with 5% BSA (Beijing Solarbio Science & Technology Co., Ltd.)
at 37°C for 2 h, the membranes were incubated overnight at 4°C with
primary antibodies, followed by incubation with the corresponding
HRP-conjugated secondary antibodies at 37°C for 1 h. Protein bands
were visualized using an ChemiDoc XRS+ enhanced
chemiluminescence detection system (Bio-Rad Laboratories, Inc.).
Protein expression levels were semi-quantified using ImageLab
software (version 5.2.1; Bio-Rad Laboratories, Inc.). The primary
and secondary antibodies used for western blotting are listed in
Table SI.
Statistical analysis
Data are presented as the mean ± SD of three
independent experiments. Comparisons among groups were analyzed
using one-way ANOVA followed by Bonferroni's post hoc test.
Statistical analyses were performed using SPSS software (version
21.0; IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Lycorine inhibits CRC cell
proliferation
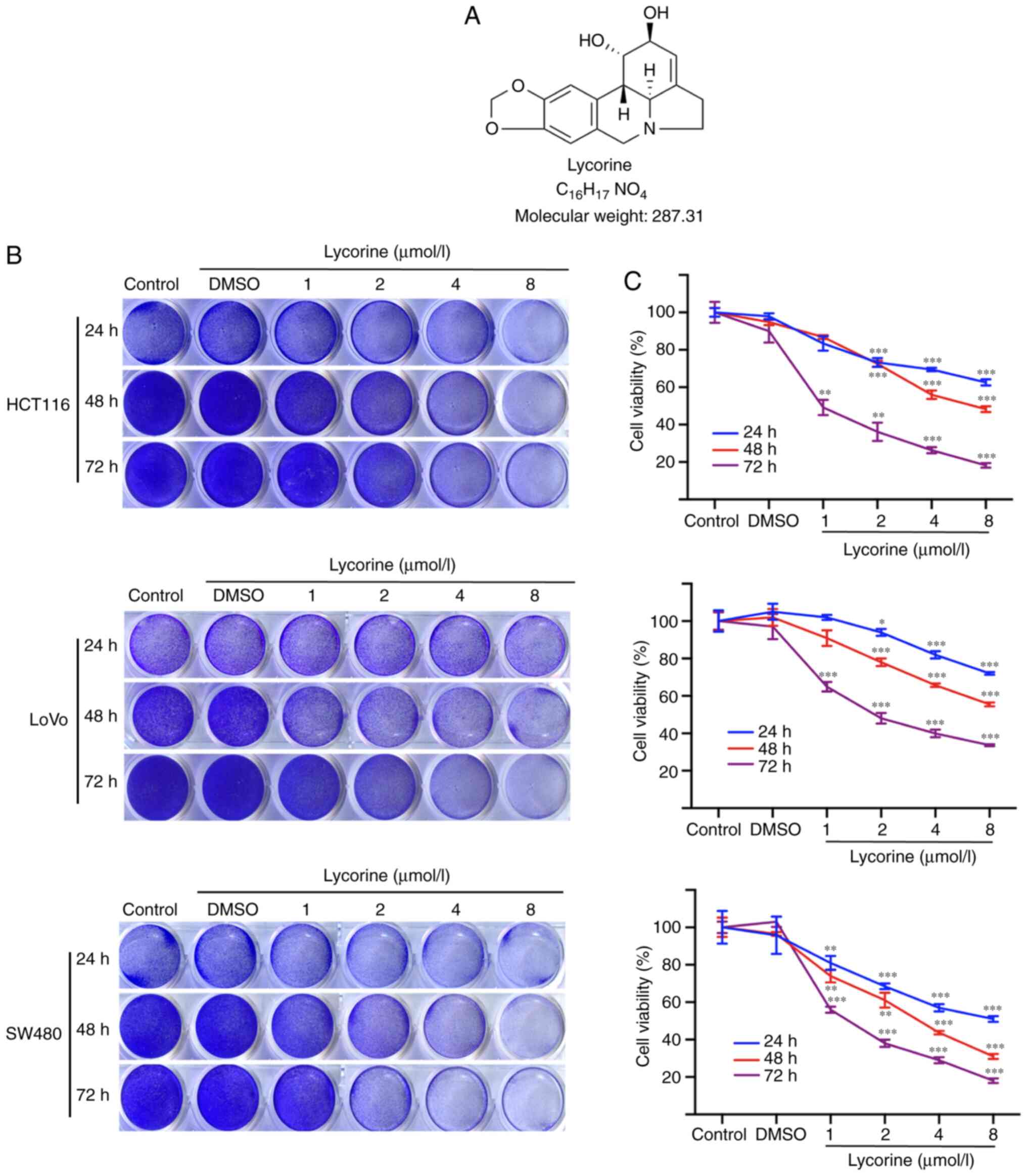
The chemical structure of lycorine is presented in
Fig. 1A. Firstly, the effects of
lycorine on human CRC cell proliferation were investigated. The
crystal violet staining assay results demonstrated that lycorine
suppressed cell proliferation in the three CRC cell lines in a
dose- and time-dependent manner (Fig. 1B
and C). Following treatment for 72 h, the IC50
values of lycorine in HCT116, LoVo and SW480 cells were 1.4, 3.8
and 1.3 µmol/l, respectively. The results indicated that HCT116 and
LoVo cells were less sensitive to lycorine compared with the SW480
cell line; therefore, the HCT116 and LoVo cell lines were selected
for subsequent experiments. The results suggested that lycorine
inhibited CRC cell proliferation in vitro.
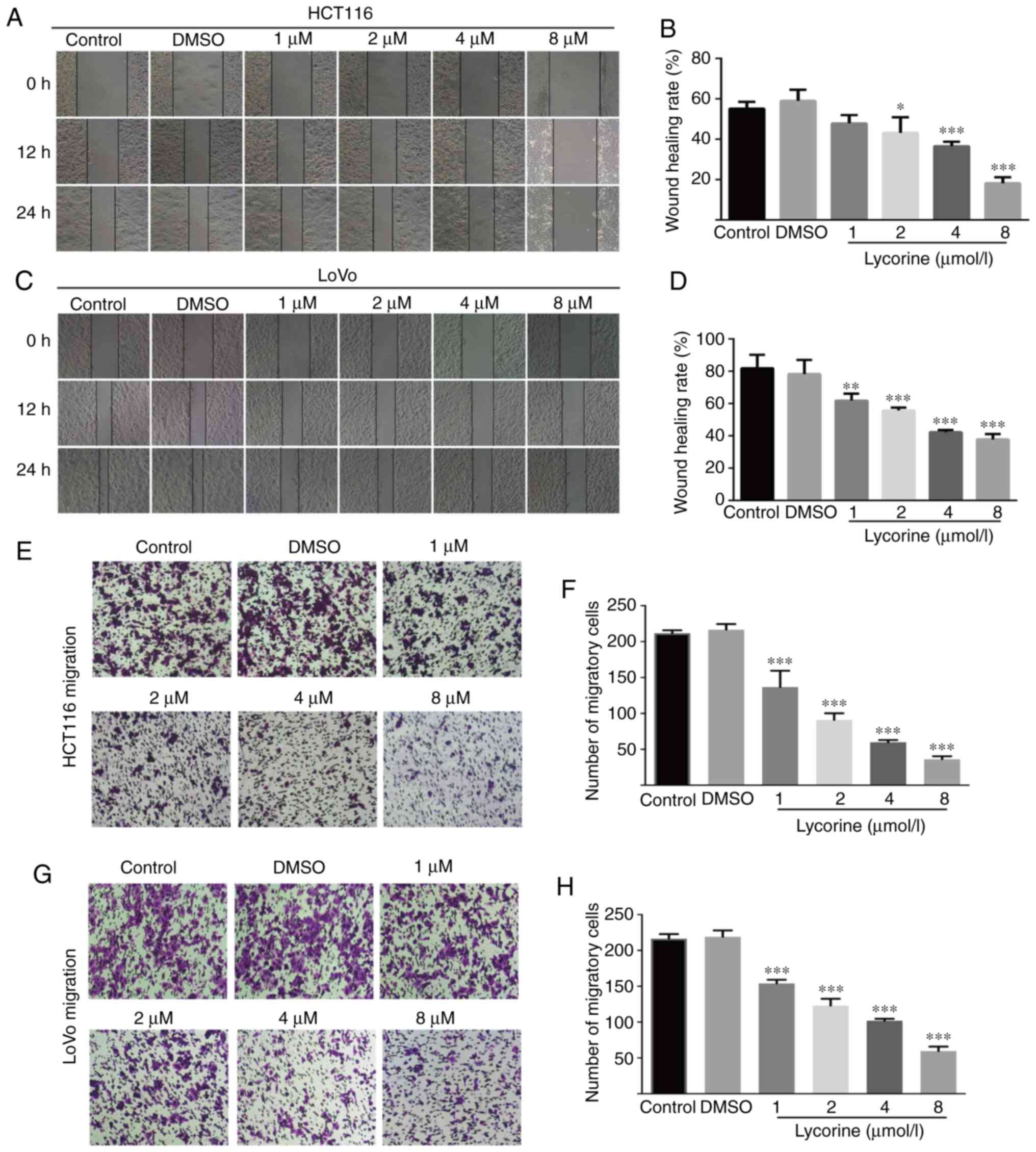
Lycorine decreases CRC cell migration
and invasion
Wound healing assays are affected by cell migration
and proliferation, and serum starvation is a common method used to
minimize cell proliferation (10);
however, the degree of cell tolerance in low-serum conditions
should be taken into consideration. Therefore, wound healing and
Transwell assays were performed to investigate lycorine
treatment-mediated alterations to cell migration and invasion,
respectively. Compared with the control group, lycorine (≥2 µmol/l)
significantly reduced the wound healing rate of CRC cells (Fig. 2A-D). Also, lycorine (≥4 µmol/l)
notably inhibited the wound healing rate of CRC cells to a similar
extent at both 12 and 24 h (Fig. 2A and
C). The aforementioned results suggested that lycorine
inhibited CRC cell migration (Fig.
2A-D). The inhibitory effect of lycorine on CRC cell migration
was further assessed by performing Transwell assays (Fig. 2E-H). Moreover, the Transwell invasion
assay results indicated that lycorine significantly decreased the
number of invading CRC cells compared with the control group
(Fig. 3A-D). The results suggested
that lycorine attenuated CRC cell migration and invasion.
Subsequently, western blotting was performed to evaluate the effect
of lycorine on epithelial-mesenchymal transition (EMT) and
extracellular matrix (ECM) degradation, which have been reported to
serve essential roles in tumor cell migration and invasion
(11,12). Compared with HCT116 cells, the
undifferentiated LoVo cell line displays a high EMT-signature,
which might be a more representative mesenchymal phenotype
(13). Therefore, in LoVo cells,
compared with the control group, lycorine significantly decreased
the protein expression levels of snail family transcriptional
repressor 1 (Snail), Vimentin and N-cadherin, which are EMT
inducers (11,14), but significantly increased the protein
expression levels of E-cadherin, which is an anti-EMT molecule
(14) (Fig.
3E). Moreover, the protein expression levels of ECM degradation
markers, including MMP-2, MMP-7 and MMP-9, were significantly
downregulated by lycorine (≥4 µmol/l) compared with the control
group. Collectively, the results suggested that lycorine inhibited
CRC cell migration and invasion.
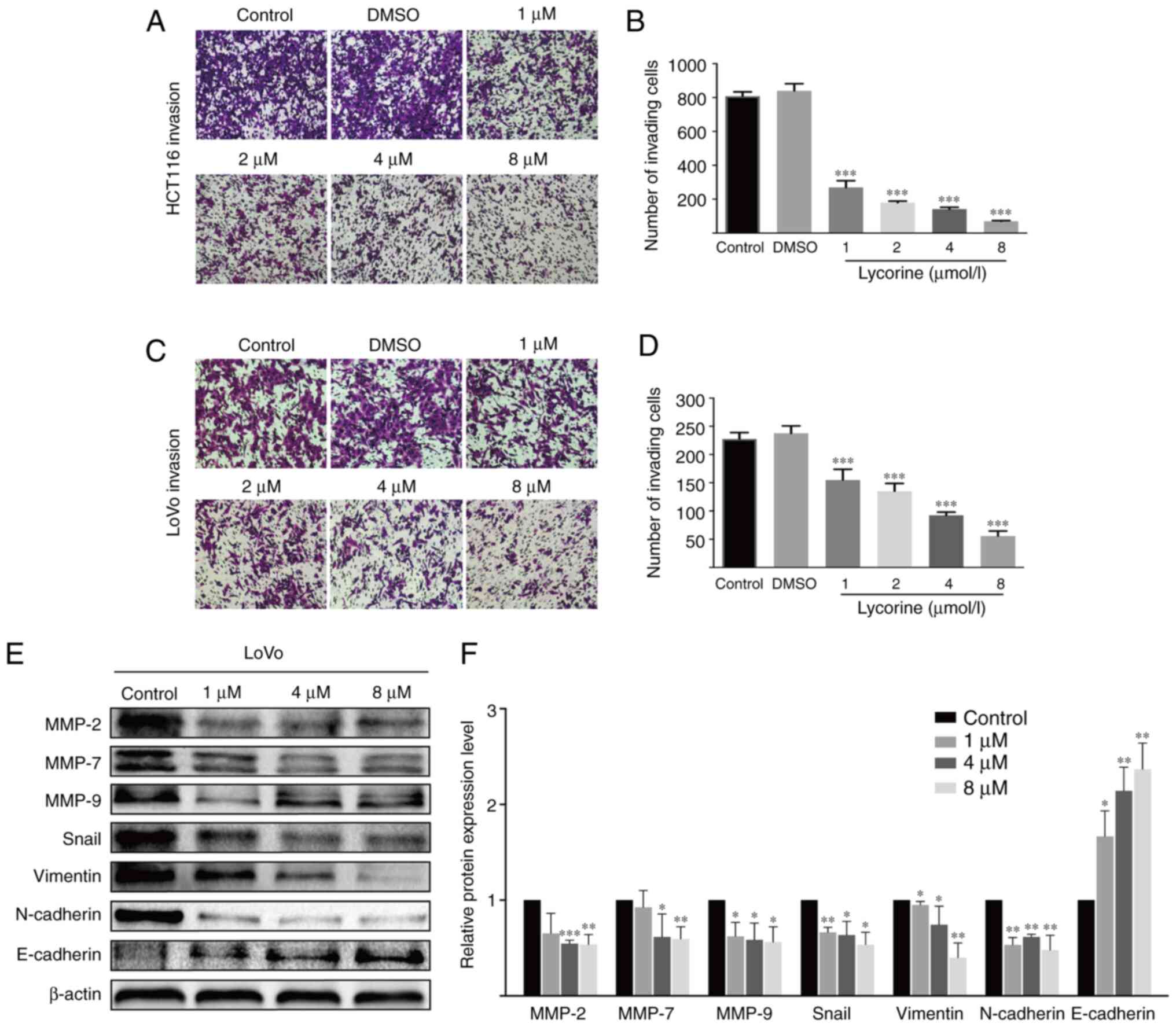 | Figure 3.Effect of lycorine on CRC cell
invasion in vitro. Effect of lycorine on HCT116 cell
invasion was (A) determined by performing Transwell invasion assays
(magnification, ×100) and (B) quantified. Effect of lycorine on
LoVo cell invasion was (C) determined by performing Transwell
invasion assays (magnification, ×100) and (D) quantified. Effect of
lycorine on the protein expression levels of MMP-2, MMP-7, MMP-9,
Snail, Vimentin, N-cadherin and E-cadherin in LoVo cells were (E)
determined via western blotting and (F) semi-quantified. Data are
presented as the mean ± SD (n=3). *P<0.05, **P<0.01 and
***P<0.001 vs. Control. CRC, colorectal cancer; MMP, matrix
metallopeptidase; Snail, snail family transcriptional repressor
1. |
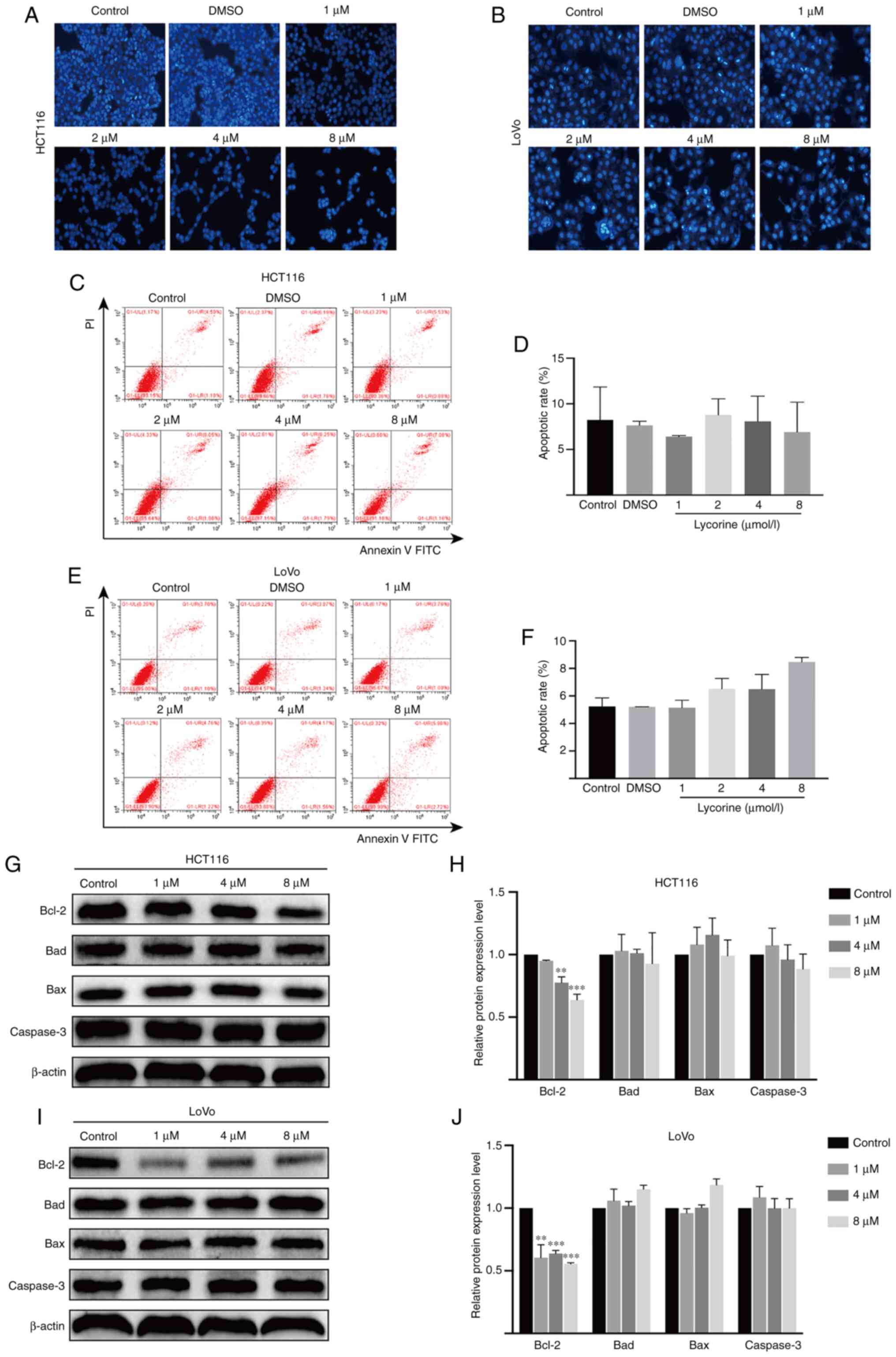
Lycorine displays no obvious effect on
CRC cell apoptosis
Hoechst 33258 staining was conducted to investigate
whether lycorine altered HCT116 and LoVo cell apoptosis. Compared
with the control group, karyopyknosis, karyorrhexis and apoptotic
bodies were not increased by lycorine treatment (Fig. 4A and B). The flow cytometry results
further demonstrated that lycorine did not significantly alter
HCT116 and LoVo cell apoptosis compared with the control group.
Moreover, the western blotting results indicated that the protein
expression levels of antiapoptotic Bcl-2 were significantly
decreased by lycorine (≥4 µmol/l) treatment compared with the
control group (Fig. 4G-J). Moreover,
the protein expression levels of Caspase-3, an important downstream
executor in the apoptosis cascade (15), and pro-apoptotic Bad and Bax were not
significantly altered by lycorine treatment compared with the
control group. The results indicated that lycorine-induced
inhibition of proliferation was not associated with apoptosis in
CRC cells.
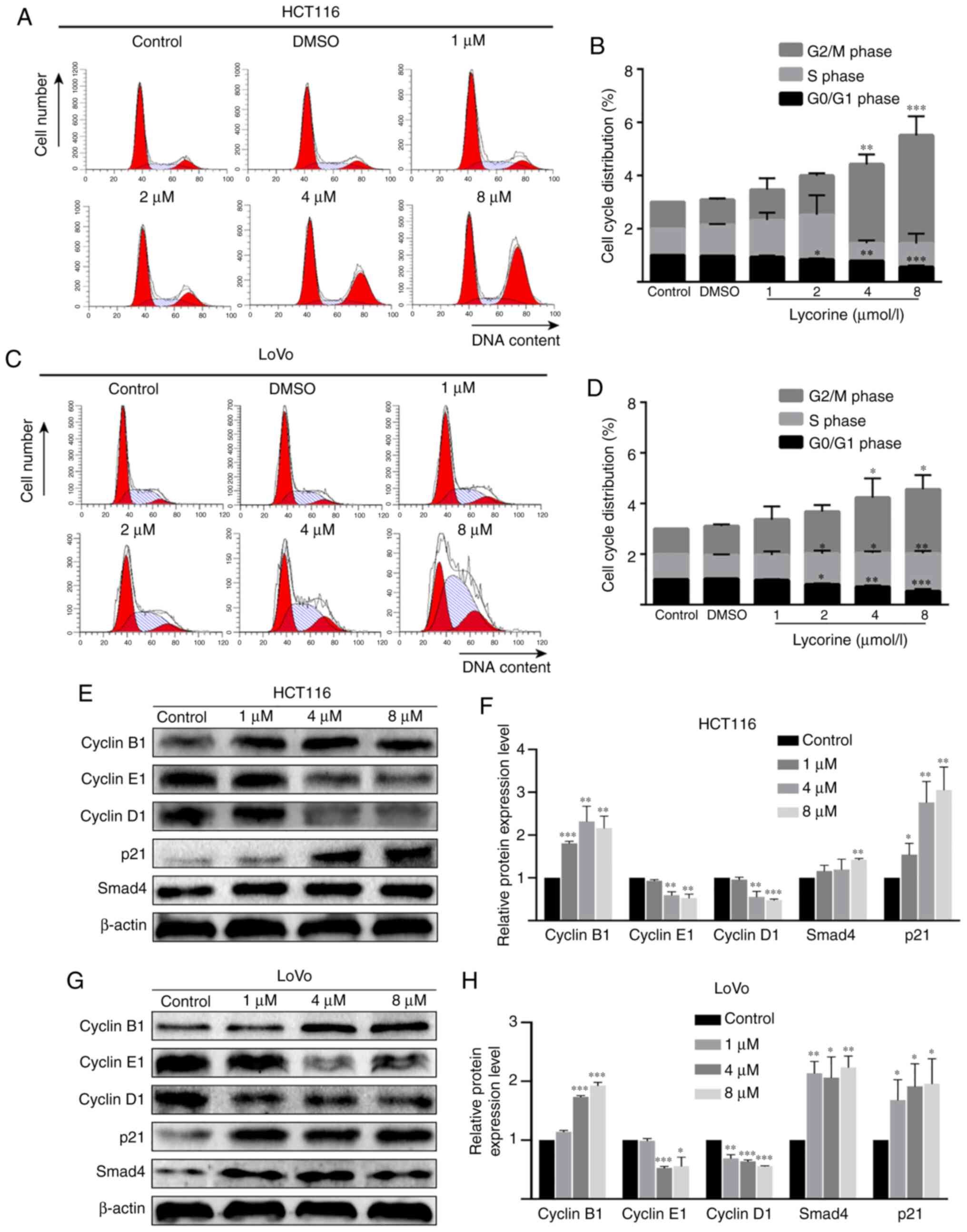
Lycorine induces cell cycle arrest in
CRC cell lines
As lycorine displayed no obvious effect on CRC cell
apoptosis, it was hypothesized that lycorine might exert cytostasis
to inhibit CRC cell proliferation. To investigate the hypothesis,
flow cytometry was performed to determine the effect of lycorine on
the cell cycle of CRC cells. Compared with the control group,
lycorine treatment (≥4 µmol/l) resulted in a significantly higher
proportion of cells in the G2/M phase in HCT116 cells,
but resulted in a significantly higher ratio of cells in the S and
G2/M phases in LoVo cells (Fig. 5A-D). In addition, the western blotting
results indicated that compared with the control group, cyclin B1
expression levels were significantly increased, but cyclin D1 and
cyclin E1 expression levels were significantly decreased by
lycorine (≥4 µmol/l) in HCT116 and LoVo cells (Fig. 5E-H). Notably, the protein expression
levels of cell cycle regulator p21 (≥1 µmol/l) and Smad4 (8 µmol/l
in HCT116 cells and ≥1 µmol/l in LoVo cells) were significantly
upregulated by lycorine treatment compared with the control group.
The results demonstrated that lycorine inhibited cell proliferation
potentially via disrupting CRC cell progression.
Lycorine promotes the activation of
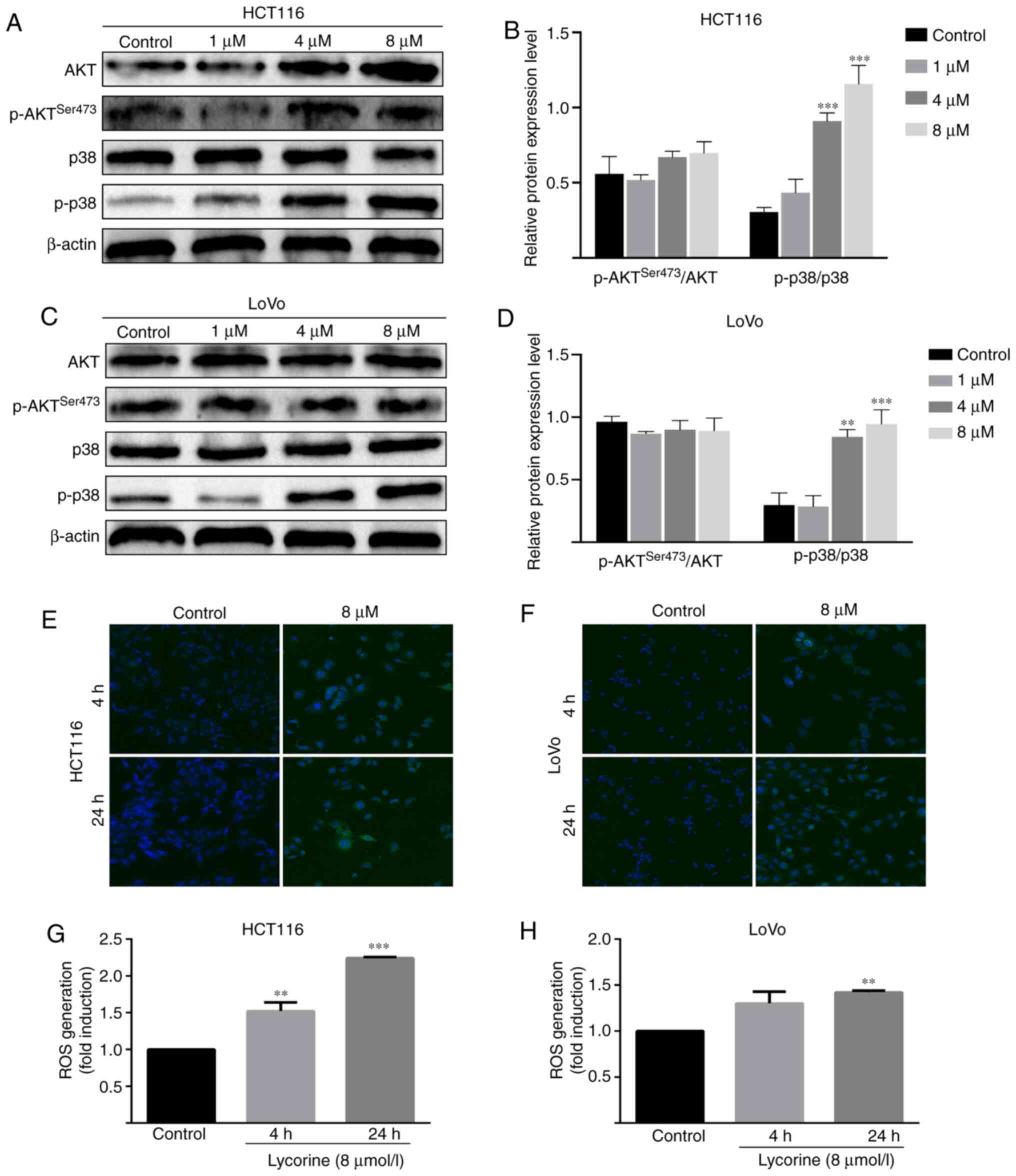
AKT and ROS/p38 signaling pathways in CRC cells
Subsequently, the possible mechanism underlying
lycorine-induced inhibitory effects against CRC cells were
investigated. Compared with the control group, the protein
expression level of phosphorylated-p38 was significantly increased
by lycorine (≥4 µmol/l) in HCT116 and LoVo cells (Fig. 6A-D). Furthermore, compared with the
control group, lycorine treatment significantly increased the
levels of intracellular ROS in HCT116 (at 4 and 24 h) and LoVo
cells (at 4 h) (Fig. 6E-H). The
results suggested that lycorine might activate the ROS/p38
signaling pathway in CRC cells. Moreover, compared with the control
group, the protein expression levels of phosphorylated AKT and
total AKT significantly increased by lycorine treatment (≥4 µmol/l)
in HCT116 cells (Fig. 6C).
Collectively, the results suggested that lycorine might enhance the
activation of AKT and ROS/p38 signaling pathways in CRC cells.
Discussion
Colorectal cancer is a life-threatening tumor
(1). Several drugs have been approved
for the chemotherapy of CRC, and although these drugs display
certain effects, patients typically develop drug resistance and the
drugs display strong cytotoxicity against healthy cells and organs
(16). For example, gastrointestinal
(GI) toxicity as an adverse event exists universally in systemic
therapy for CRC. Single 5-FU bolus or 5-FU-based combination
chemotherapy can result in GI side effects (17). Moreover, 5-FU causes myelosuppression,
hand-foot syndrome and cardiotoxicity (18,19).
Therefore, preoperative and postoperative adjuvant therapies for
CRC treatment require the introduction of more effective and safer
drugs. Lycorine is an alkaloid that is located in the bulbs of
plant lycoris, and has been reported to effectively inhibit several
types of cancer (6–9). In the present study, the anticancer
effect of lycorine on CRC was investigated. Lycorine treatment
inhibited cell proliferation in a dose- and time-dependent manner
in the three CRC cell lines. Moreover, the IC50 value of
lycorine following 72 h treatment was 1.4 µmol/l in HCT116, which
was similar to a previous study (20). Moreover, the results indicated that
compared with the control group, lycorine significantly inhibited
migration and invasion, but displayed no significant effect on CRC
cell apoptosis.
MMPs serve vital roles in tumor metastasis via
mediating ECM degradation (21).
MMP-9, a widely studied MMP, is a potential cancer biomarker due to
its involvement in numerous cancer-related processes, including
cell migration, invasion, angiogenesis and inflammation (22). Additionally, epithelial tumor cells
can obtain migratory and invading characteristics via the process
of EMT (23). The LoVo cell line is a
CRC cell line with a high degree of malignancy (13). The present study demonstrated that
lycorine treatment significantly downregulated the protein
expression levels of MMP-2, MMP-7 and MMP-9 in LoVo cells compared
with the control group. Furthermore, compared with the control
group, lycorine significantly increased the protein expression
level of E-cadherin, an anti-EMT regulator, and significantly
decreased the protein expression levels of Snail, Vimentin and
N-cadherin, which are pivotal EMT inducers. Collectively, the
results suggested that lycorine inhibited CRC cell migration and
invasion potentially via downregulating MMP protein expression
levels and reversing the EMT process in CRC cells.
Apoptosis is a highly regulated process of cellular
death. There are two classic apoptosis signaling pathways:
Extrinsic and intrinsic. The extrinsic apoptosis signaling pathway
is initialized by the interaction between death receptors and TNF,
Fas ligand or TNF superfamily member 10 ligand. By contrast, the
intrinsic apoptosis signaling pathway refers to
non-receptor-mediated initiation and mitochondrial regulation
(24). The mechanism underlying
lycorine-mediated apoptosis in tumors has been investigated.
Previous studies revealed that lycorine induced apoptosis via a
mitochondria-dependent apoptotic cascade or the death-receptor
signaling pathway in tumor cells, including bladder cancer T24
cells (7), leukemia Mcl-1 cells
(25) and HL-60 cells (26). In the present study, the Hoechst 33258
staining and flow cytometry results demonstrated that lycorine
displayed no significant effect on HCT116 and LoVo cell apoptosis
compared with the control group. Moreover, compared with the
control group, lycorine significantly decreased the expression
levels of Bcl-2, which displays an antiapoptotic effect by
inhibiting Bax and blocking the process of mitochondrial outer
membrane permeabilisation (27).
However, the protein expression levels of Bad, Bax or the apoptosis
effector caspase-3 were not significantly altered by lycorine
treatment compared with the control group, which was consistent
with the morphology and flow cytometry results. The results
indicated that lycorine might primarily exert cytostatic effects
rather than apoptosis inducing effects.
The AKT signaling pathway serves an important role
in response to cell survival (28).
Activation of the AKT signaling pathway may be related to the
cytostatic effects of several anticancer drugs. For example,
Richards et al (29) reported
that Crassin, a Diterpenoid natural compound against cancer cells,
induced cytostatic effects via activating the AKT signaling pathway
and then promoting ROS activation in triple-negative breast cancer
cells. Goyeneche et al (30)
and Wempe et al (31) reported
that AKT signaling was activated to exert cytostatic effects on
ovarian cancer cells in mifepristone-induced antitumor activity.
The western blotting results indicated that AKT activation might
serve a role in the cytostatic effect of lycorine against CRC
cells. Moreover, whether lycorine in combination with AKT
inhibitors could inhibit the cytostatic effect of lycorine on CRC
cells requires further investigation.
Previous studies have demonstrated that 5 µM
lycorine reduced mitosis in glioblastoma U373 cells and
non-small-cell lung cancer A549 cells by >90% without inducing
cell apoptosis (9,32), which suggested that lycorine may
display strong inhibitory effects on cancer, especially in those
naturally resistant to proapoptotic stimuli. In the present study,
based on the finding that lycorine did not alter apoptosis, it was
hypothesized that lycorine may primarily exert cytostatic effects
by regulating the cell cycle in CRC cells. The flow cytometry
results supported this hypothesis, demonstrating that lycorine
significantly induced cell cycle arrest at the G2/M
phase in HCT116 cells, and at the S and G2/M phase in
LoVo cells compared with the control group. Cancer cells display
cell cycle dysregulation, resulting in unscheduled and rapid cell
division, which is a hallmark of tumors (33). The cell cycle is controlled by a
subfamily of CDKs complexed with cyclin proteins (34). Aberrations of CDKs drive re-entry into
the cell cycle in cancer cells (35);
therefore, inducing cell cycle arrest via activation of checkpoints
to modulate CDK activity may serve as an effective therapeutic
strategy. For example, targeting the cyclin D1-Cdk4/6 complex as a
therapeutic invention has been focused on due to the invariable
deregulation of the complex in human tumors (36). CDK4/6 inhibitors are already available
for clinical use in the USA (37).
Meanwhile, in the present study, compared with the control group,
lycorine significantly downregulated the protein expression levels
of cyclin D1 and cyclin E1 in HCT116 and LoVo cells. p21 is a
negative regulator of the cell cycle that functions by dampening
the activity of cyclin-CDK complexes (38). The results of the present study
indicated that the protein expression levels of p21 were
significantly increased by lycorine treatment compared with the
control group. In CRC, Smad4 serves a tumor-suppressing role by
controlling a cohort of targeting genes, including p21, to trigger
cell cycle arrest at the G1/S phase (39). It has been reported that Smad4
expression was absent in 20–40% of cases, which promotes tumor
progression and is associated with poor prognosis (40). The present study demonstrated that
lycorine significantly upregulated the protein expression levels of
Smad4 compared with the control group. Collectively, the results
indicated that lycorine exerted anti-CRC effects potentially via
triggering cell cycle arrest in CRC cells.
p38 belongs to the MAPK family and is involved in
cell proliferation, migration, differentiation and apoptosis
(41,42). The roles of p38 in occurrence and
development of cancer have been previously investigated (43); however, the obtained results are
controversial, with certain studies suggesting a stimulatory role
of p38 in cancer and others proposing an inhibitory role of p38. A
recent study demonstrated that p38 interfered with initial steps of
colorectal adenomagenesis, and nuclear p38 was correlated with
low-grade dysplasia and decreased adenoma size (44). The results of the present study
demonstrated that lycorine promoted activation of the p38 signaling
pathway, leading to increased p38 phosphorylation in CRC cells
compared with the control group. Therefore, the mechanism
underlying p38 activation was investigated. p38 can be activated by
several stimuli, such as inflammatory factors, DNA damage and ROS
(42). ROS serves pivotal roles in
cell physiology and participates in several pathological processes.
When ROS generation exceeds the antioxidant capacity of the cell,
cells enter an oxidative stress state, causing cell dysfunction and
behavioral alterations, including cell cycle arrest, senescence and
cell death (45). Therefore, it was
hypothesized that p38 activation may be a consequence of ROS
generation. To verify whether lycorine affected ROS accumulation, a
DCFH-DA staining assay was performed. The results demonstrated that
ROS intracellular levels in HCT116 and LoVo cells were increased by
lycorine treatment compared with the control group. Therefore,
lycorine-induced antitumor effects might be closely related to
ROS/p38 signaling.
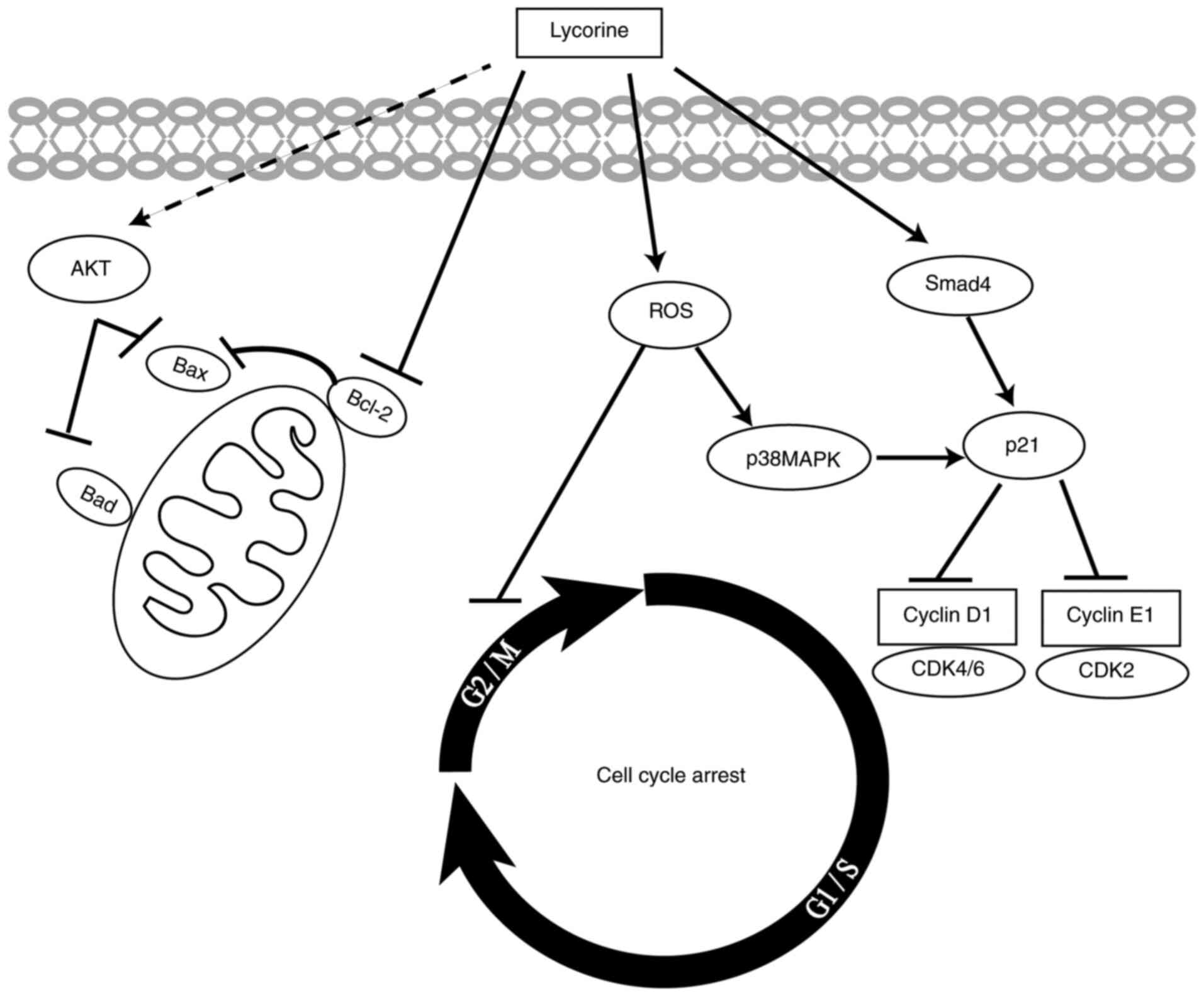
In conlusion, the present study demonstrated that
lycorine suppressed CRC cell proliferation, migration and invasion
and cell cycle progression in vitro. Moreover, to the best
of our knowledge, the present study was the first to suggest that
lycorine exerted cytostatic effects on CRC cells via ROS/p38 and
AKT signaling pathways (Fig. 7).
Therefore, the results of the present study suggested that lycorine
might serve as a therapeutic for CRC. Future studies should
investigate the correlation between ROS accumulation and p38
activation, as well as the potential of lycorine in combination
with other drugs in CRC treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81874001) and the
Scientific Research and Technology Development Program of Chongqing
(grant no. cstc2017jcyjAX0196).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL, PZ, XY, TY, HH, CY, LZ, SY and XL made
substantial contributions to the study design. JL and XL critically
revised the manuscript for important intellectual content. PZ
drafted the manuscript, and agreed to be accountable for the work
in ensuring that questions related to the integrity of any part of
the work are appropriately investigated and resolved. PZ, CY, LZ
and SY acquired the data. XY, TY and HH analyzed and interpreted
the data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
DCFH-DA
|
2′,7′-dichlorofluorescein-diacetate
|
|
ECM
|
extracellular matrix
|
|
EMT
|
epithelial-mesenchymal transition
|
|
MMP
|
matrix metallopeptidase
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wild CP, Weiderpass E and Stewart BW:
2020, World Cancer Report. Cancer Research for Cancer Prevention
Lyon, France: International Agency for Research on Cancer;
http://publications.iarc.fr/586
|
|
3
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mohammed AA, El-Tanni H, El-Khatib HM,
Mirza AA and El-Kashif AT: WITHDRAWN: Molecular classification of
colorectal cancer: Current perspectives and controversies. J Egypt
Natl Canc Inst. Jan 2–2016.(Epub ahead of print). doi:
10.1016/j.jnci.2015.11.004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kaya GI, Cicek D, Sarikaya B, Onur MA and
Somer NU: HPLC - DAD analysis of lycorine in Amaryllidaceae
species. Nat Prod Commun. 5:873–876. 2010.PubMed/NCBI
|
|
6
|
Luo Y, Roy M, Xiao X, Sun S, Liang L, Chen
H, Fu Y, Sun Y, Zhu M, Ye M and Liu J: Lycorine induces programmed
necrosis in the multiple myeloma cell line ARH-77. Tumour Biol.
36:2937–2945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang C, Wang Q, Li X, Jin Z, Xu P, Xu N,
Xu A, Xu Y, Zheng S, Zheng J, et al: Lycorine induces apoptosis of
bladder cancer T24 cells by inhibiting phospho-AKT and activating
the intrinsic apoptotic cascade. Biochem Biophys Res Commun.
483:197–202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ying X, Huang A, Xing Y, Lan L, Yi Z and
He P: Lycorine inhibits breast cancer growth and metastasis via
inducing apoptosis and blocking Src/FAK-involved pathway. Sci China
Life Sci. 60:417–428. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lamoral-Theys D, Andolfi A, Van
Goietsenoven G, Cimmino A, Le Calvé B, Wauthoz N, Mégalizzi V, Gras
T, Bruyère C, Dubois J, et al: Lycorine, the main phenanthridine
amaryllidaceae alkaloid, exhibits significant antitumor activity in
cancer cells that display resistance to proapoptotic stimuli: An
investigation of structure-activity relationship and mechanistic
insight. J Med Chem. 52:6244–6256. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Davis PK, Ho A and Dowdy SF: Biological
methods for cell-cycle synchronization of mammalian cells.
Biotechniques. 30:1322–1331. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuzhalin AE, Lim SY, Kutikhin AG and
Gordon-Weeks AN: Dynamic matrisome: ECM remodeling factors
licensing cancer progression and metastasis. Biochim Biophys Acta
Rev Cancer. 1870:207–228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Berg KCG, Eide PW, Eilertsen IA,
Johannessen B, Bruun J, Danielsen SA, Bjørnslett M, Meza-Zepeda LA,
Eknæs M, Lind GE, et al: Multi-omics of 34 colorectal cancer cell
lines-a resource for biomedical studies. Mol Cancer. 16:1162017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Serrano-Gomez SJ, Maziveyi M and Alahari
SK: Regulation of epithelial-mesenchymal transition through
epigenetic and post-translational modifications. Mol Cancer.
15:182016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meyers BM, Cosby R, Quereshy F and Jonker
D: Adjuvant chemotherapy for stage II and III colon cancer
following complete resection: A cancer care ontario systematic
review. Clin Oncol (R Coll Radiol). 29:459–465. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee CS, Ryan EJ and Doherty GA:
Gastro-intestinal toxicity of chemotherapeutics in colorectal
cancer: The role of inflammation. World J Gastroenterol.
20:3751–3761. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Xing X, Meng F, Wang Y and Zhong
D: Oral fluoropyrimidine versus intravenous 5-fluorouracil for the
treatment of advanced gastric and colorectal cancer: Meta-analysis.
J Gastroenterol Hepatol. 33:209–225. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdel-Rahman O: 5-Fluorouracil-related
cardiotoxicity; findings from five randomized studies of
5-fluorouracil-based regimens in metastatic colorectal cancer. Clin
Colorectal Cancer. 18:58–63. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang P, Yuan HH, Zhang X, Li YP, Shang LQ
and Yin Z: Novel lycorine derivatives as anticancer agents:
Synthesis and in vitro biological evaluation. Molecules.
19:2469–2480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Winer A, Adams S and Mignatti P: Matrix
metalloproteinase inhibitors in cancer therapy: Turning past
failures into future successes. Mol Cancer Ther. 17:1147–1155.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang H: Matrix metalloproteinase-9
(MMP-9) as a cancer biomarker and MMP-9 biosensors: Recent
advances. Sensors (Basel). 18:32492018. View Article : Google Scholar
|
|
23
|
Bates RC and Mercurio AM: The
epithelial-mesenchymal transition (EMT) and colorectal cancer
progression. Cancer Biol Ther. 4:365–370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu XS, Jiang J, Jiao XY, Wu YE, Lin JH
and Cai YM: Lycorine induces apoptosis and down-regulation of Mcl-1
in human leukemia cells. Cancer Lett. 274:16–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu J, Hu JL, Shi BW, He Y and Hu WX:
Up-regulation of p21 and TNF-alpha is mediated in lycorine-induced
death of HL-60 cells. Cancer Cell Int. 10:252010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song G, Ouyang G and Bao S: The activation
of Akt/PKB signaling pathway and cell survival. J Cell Mol Med.
9:59–71. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Richards CE, Vellanki SH, Smith YE and
Hopkins AM: Diterpenoid natural compound C4 (Crassin) exerts
cytostatic effects on triple-negative breast cancer cells via a
pathway involving reactive oxygen species. Cell Oncol (Dordr).
41:35–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goyeneche AA, Carón RW and Telleria CM:
Mifepristone inhibits ovarian cancer cell growth in vitro and in
vivo. Clin Cancer Res. 13:3370–3379. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wempe SL, Gamarra-Luques CD and Telleria
CM: Synergistic lethality of mifepristone and LY294002 in ovarian
cancer cells. Cancer Growth Metastasis. 6:1–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Van Goietsenoven G, Andolfi A, Lallemand
B, Cimmino A, Lamoral-Theys D, Gras T, Abou-Donia A, Dubois J,
Lefranc F, Mathieu V, et al: Amaryllidaceae alkaloids belonging to
different structural subgroups display activity against
apoptosis-resistant cancer cells. J Nat Prod. 73:1223–1227. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Malumbres M and Barbacid M: To cycle or
not to cycle: A critical decision in cancer. Nat Rev Cancer.
1:222–231. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schafer KA: The cell cycle: A review. Vet
Pathol. 35:461–478. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
O'Leary B, Finn RS and Turner NC: Treating
cancer with selective CDK4/6 inhibitors. Nat Rev Clin Oncol.
13:417–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goel S, DeCristo MJ, McAllister SS and
Zhao JJ: CDK4/6 inhibition in cancer: Beyond cell cycle arrest.
Trends Cell Biol. 28:911–925. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abbas T and Dutta A: p21 in cancer:
Intricate networks and multiple activities. Nat Rev Cancer.
9:400–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao M, Mishra L and Deng CX: The role of
TGF-β/SMAD4 signaling in cancer. Int J Biol Sci. 14:111–123. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Salovaara R, Roth S, Loukola A, Launonen
V, Sistonen P, Avizienyte E, Kristo P, Järvinen H, Souchelnytskyi
S, Sarlomo-Rikala M and Aaltonen LA: Frequent loss of SMAD4/DPC4
protein in colorectal cancers. Gut. 51:56–59. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cuenda A and Sanz-Ezquerro JJ: p38γ and
p38δ: From spectators to key physiological players. Trends Biochem
Sci. 42:431–442. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bonney EA: Mapping out p38MAPK. Am J
Reprod Immunol. 77:e126522017. View Article : Google Scholar
|
|
43
|
Martínez-Limón A, Joaquin M, Caballero M,
Posas F and de Nadal E: The p38 pathway: From biology to cancer
therapy. Int J Mol Sci. 21:19132020. View Article : Google Scholar
|
|
44
|
Handra-Luca A, Bendib M and Magkou C: P38
expression in colorectal adenomas: Relationships to cell
proliferation, stem phenotype and akt pathway proteins. Minerva
Gastroenterol Dietol. 66:208–210. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Moloney JN and Cotter TG: ROS signalling
in the biology of cancer. Semin Cell Dev Biol. 80:50–64. 2018.
View Article : Google Scholar : PubMed/NCBI
|