Introduction
Lung cancer, one of the most common globally
diagnosed cancers, has a high mortality rate. The percentage of
non-small cell lung cancer (NSCLC) is estimated at 85% among all
lung cancers (1). Although clinical
diagnosis and therapeutic strategies of NSCLC have advanced over
the past three decades, the 5-year overall survival rate remains
<16% (2). Among the reasons, tumor
metastasis is one of the main causes of clinical treatment failure
and recurrence (3,4). Therefore, it is necessary to elucidate
the underlying mechanisms of NSCLC cell growth and metastasis,
which may shed light on potential therapies for NSCLC.
MicroRNAs (miRNAs) belong to a class of small
non-coding RNAs which regulating gene expression and protein-coding
genes (5). Increasing studies have
revealed that miRNAs are closely associated with tumor cell
proliferation, migration, invasion, drug resistance and apoptosis
(6,7).
For instance, miR-410 was revealed to promote
epithelial-mesenchymal transition (EMT) and radio-resistance by
targeting the PI3K/mTOR axis in NSCLC (8). miR-219 functioned as a tumor suppressor,
which inhibited NSCLC and ovarian cancer cell growth and metastasis
(9,10).
miR-454-3p is a member of the
miR-130-3p/301-3p/454-3p miRNA cluster, which was revealed to play
a critical role in the progression and malignancies of various
cancers (11). At present, some
studies have revealed that miR-454-3p functioned as a suppressive
miRNA in several cancers, including glioblastoma (12), lung cancer (13), bladder cancer (14) and pancreatic ductal adenocarcinoma
(15). However, the detailed
functions of miR-454-3p in NSCLC proliferation and metastasis
remain to be characterized.
Transforming growth factor-β2 (TGFB2) is a protein
coding gene of TGF-β2, and it has been revealed to regulate tumor
cell growth, proliferation and metastasis (16,17).
Emerging evidence has revealed that TGF-β2 was upregulated in
numerous types of cancer (18). Bruna
et al revealed that TGF-β2 was abundant in glioma and
correlated with poor prognosis (19).
Yang et al have reported that TGF-β2 contributed to EMT and
tumor mutation burden in gastric cancer (17). EMT is a key biological process that
induces malignant tumor cell migration and invasion (20,21). TGF-β
is regarded as the most crucial factor for EMT (22). Consequently, targeting TGF-β2 could be
a promising treatment strategy for cancer. Whether the expression
of TGF-β2 is related with the expression level of miR-454-3p in
NSCLC remains to be elucidated.
In the present study, the expression of miR-454-3p
and TGFB2 was investigated in NSCLC tissues and cell lines.
Furthermore, it was explored how miR-454-3p and TGFB2 contribute to
the progression of NSCLC and the underlying mechanisms were also
investigated.
Materials and methods
Clinical specimens
The present study was carried out with a total
number of 56 human NSCLC tissue and 56 adjacent non-tumor tissue
samples. These patients were between 25–75 years old, and the ratio
of males to females was 1.15:1. Patients were confirmed to have no
other serious diseases except NSCLC. The 56 NSCLC patients were
clinically diagnosed and undergoing surgery (had never received any
neo-adjuvant treatment) at the Yuebei People's Hospital of Shaoguan
from March 2013 to October 2018. Patients provided written informed
consent for their participation in the present study. These
clinical samples were stored in liquid nitrogen until their use in
experiments. The research was approved and carried out according to
the ethical standards of the Ethics Committee of Yuebei People's
Hospital (approval no. DD-KY-2018310).
Cells and cell culture
The normal lung cell line BEAS-2B and NSCLC cell
lines A549, NCI-H1299, NCI-H1650, NCI-H460, NCI-H1975 were
purchased from American Type Culture Collection (ATCC). All the
cells were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
with 10% fetal bovine serum (FBS; Shanghai ExCell Biotech Co.,
Ltd.) and 1% penicillin/streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) in 5% CO2 at 37°C. By separately using
an STR Multi-Amplification Kit (Microreader 21 ID System; Suzhou
Microread Genetics) and Mycoplasma Detection Set (M&C Gene
Technology), all the cell lines were identified with no cross
contamination with other cell lines and negative for
mycoplasma.
Reverse transcription-quantitative
(RT-q)PCR
E.Z.N.A.® Total RNA Kit I (Omega Bio-Tek)
was used to extract total RNA from tissues and cultured cells in
accordance with the manufacturer's protocol. Then, cDNAs were
synthesized by All-in-One cDNA Synthesis SuperMix (Bimake) on a PCR
instrument (C1000; Bio-Rad Laboratories, Inc.). The thermocycling
conditions were as follows: 25°C/10 min; 42°C/30 min; and 85°C/5
min. Next, real-time qPCR was performed using 2X SYBR Green qPCR
Master Mix (Bimake) on a LightCycler 480 System (Roche
Diagnostics). The thermocycling conditions were as follows:
Pre-incubation at 95°C for 5 min; then 40 cycles of amplification
at 95°C for 10 sec, 56°C for 20 sec, 72°C for 20 sec; melting
curve, 1 cycle at 95°C for 5 sec, 65°C for 1 min and continuously
at 97°C; finally cooling at 95°C for 10 sec. miRNA and mRNA
expression were defined based on the quantification cycle
(Cq), and respectively normalized to U6 and GAPDH
levels. The relative expression levels were analyzed by using
2−ΔΔCq method (23). The
sequence of all primers applied in the present study are listed as
follows: miR-454-3p forward, 5′-ACCCTATCAATATTGTCTCTGC-3′ and
reverse, 5′-GCGAGCACAGAATTAATACGAC-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; TGFB2 forward,
5′-GTTCGATTTGACGTCTCAGCAAT-3′ and reverse,
5′-CAATCCGTTGTTCAGGCACTCT-3′; GAPDH forward,
5′-TGCACCACCAACTGCTTAGC-3′ and reverse,
5′-GGCATGGACTGTGGTCATGAG-3′.
Cell transient transfection
A549 and NCI-H1650 cells were seeded in 6-well
plates (2.1×104 cells/well) and incubated overnight.
After cell fusion reached 80%, the cells were transfected with i)
negative control (NC) or miR-454-3p mimics. The sequence of the
miR-454-3p mimic was 5′-UAGUGCAAUAUUGCUUAUAGGGU-3′ and that of the
negative control 5′-UCACAACCUCCUAGAAAGAGUAGA-3′; ii) NC with Vector
(pCMV3), pCMV3-TGFB2 plasmid, or pCMV3-TGFB2 plasmid with
miR-454-3p mimics using Lipofectamine™ 3000 transfection reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The concentration of NC and miR-454-3p
mimics was 100 pmol and the mass of pCMV3 and pCMV3-TGFB2 was 2 µg.
miR-454-3p mimics and negative control (NC) oligonucleotides were
synthesized by Shanghai GenePharma Co., Ltd. Vector (pCMV3) and
pCMV3-TGFB2 were purchased from Origene Technologies, Inc.. After
incubation with transfection reagent for 6 h, the cells were
cultured using complete medium for another 48 h in 5%
CO2 at 37°C. Then, the transfected cells were used to
perform subsequent experiments.
Bioinformatics analysis
Bioinformatics analysis was conducted to predict the
assumed targets of miR-454-3p using TargetScan (version 6.0;
www.targetscan.org).
Luciferase reporter assay
Luciferase reporter plasmids, pmirGLO-TGFB2-3′-UTR
wild-type (WT), and pmirGLO-TGFB2-3′-UTR mutant (Mut) were
synthesized by Shanghai GenePharma Co., Ltd. A549 and NCI-H1650
cells were seeded in 96-well plates (7.0×103 cells/well)
and cultured for 24 h. Then, the pmirGLO-TGFB2-3′-UTR-WT or
pmirGLO-TGFB2-3′-UTR-Mut plasmid was co-transfected with NC or
miR-454-3p mimics into the cells using Lipofectamine™ 3000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
After transfection for 48 h, a Dual Luciferase Reporter Assay kit
(Promega Corporation) was used to measure the luciferase signals
and the Renilla luciferase activity was used for
normalization.
Cell proliferation assay
A549 and NCI-H1650 cells were plated into 96-well
plates (5.0×103 cells/well) and incubated overnight.
After transfection, the cells were cultured for another 24 h. Cell
proliferation was analyzed by Cell Counting Kit-8 (CCK-8) assay
(Beyotime Institute of Biotechnology) at 0, 24, 48 and 72 h,
whereupon 10 µl CCK-8 solution was added to each well. Following
incubation at 37°C for another 1 h, the optical density (OD) at 450
nm was measured using a microplate reader (Bio-Rad Laboratories,
Inc.).
5-Ethynyl-2-deoxyuridine (EdU)
proliferation assay
Transfected cells were cultured in 96-well plates
(6.0×103 cells/well). After a 24-h incubation, the cells
were subjected to the iClick™ EdU Andy Fluor 555 Imaging Kit
(GeneCopoeia, Inc.) in accordance with the indicated protocol. The
images were acquired with a fluorescence microscope (magnification,
×10) (Zeiss AG) and the EdU-positive cells were calculated by
Image-Pro Plus 6.0 software (Media Cybernetics, Inc.).
Wound healing assay
A wound-healing assay can be used to evaluate cell
migration capacity (24). A549 and
NCI-H1650 cells were transfected with miRNA mimics and/or plasmid
in 12-well plates (1.5×104 cells/well). After incubation
for another 24 h, the cells were starved with serum-free DMEM for 6
h and then cell monolayers were scratched with a 10-µl pipette tip.
Next, the cells were washed gently with phosphate-buffered saline
(PBS) to remove the cell debris and then cultured with total
medium. Images were captured at 0 and 12 h after the scratch by an
optical microscope (magnification, ×5) (Zeiss AG). The wound widths
were analyzed with ImageJ 1.38 software (National Institutes of
Health).
Invasion assays
The invasion assays were conducted with 24-well
Boyden chambers (8 µm pore size; Corning, Inc.). First, the upper
chambers were pre-coated with diluted Matrigel (Corning, Inc.) and
allowed to settle for 30 min at 37°C. The transfected cells
(1.0×105 cells/ml) were resuspended in serum-free DMEM
medium and seeded into the upper chamber (100 µl/well). The bottom
chamber was filled with DMEM supplemented with 10% FBS (600
µl/well). After incubation for 24 h, the invasive cells were fixed
with 4% paraformaldehyde for 30 min and then stained with 0.1%
crystal violet for 15 min at room temperature. A cotton swab was
used to rub away the non-invading cells on the inner side of upper
chamber. The invasive cells in the lower surface were photographed
with an inverted microscope (magnification, ×10) (Olympus
Corporation) and counted by Image-Pro Plus 6.0 software.
Western blotting
A549 and NCI-H1650 cells after transfection were
collected and the total protein of cells was extracted using RIPA
buffer (Thermo Fisher Scientific, Inc.) containing proteinase
inhibitors (Roche Diagnostics). A Pierce™ Bicinchoninic Acid assay
kit (Thermo Fisher Scientific, Inc.) was used to detect the protein
concentrations. Equal amounts of protein (30 µg) were separated by
10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and then transferred onto 4.5 µm polyvinylidene fluoride
(PVDF) membranes (EMD Millipore). Subsequently, 5% skim milk in TBS
containing 0.1% Tween-20 (TBST) was used to block the membranes for
1 h at room temperature. The membranes were incubated with primary
antibodies against E-cadherin (cat. no. 3195), N-cadherin (cat. no.
4061) and GAPDH (cat. no. 5174S), which were purchased from Cell
Signaling Technology, Inc.. The dilution of the primary antibodies
was 1:1,000. After washing with TBST for three times, the
horseradish peroxidase (HRP)-conjugated secondary antibody (goat
anti-rabbit IgG; cat. no. sc-2004; 1:1,000; Santa Cruz
Biotechnology, Inc.) was probed with the blots for 1 h at room
temperature. After washing with TBST, the protein bands were
visualized by an enhanced chemiluminescence (ECL; Pierce; Thermo
Fisher Scientific, Inc.) solutions.
Statistical analysis
The experimental data were statistically analyzed
with GraphPad Prism 8.0 software (GraphPad Software, Inc.). Each
experiment was repeated at least three times and the results are
expressed as the mean ± SD. The unpaired Student's t-test was
conducted for comparisons between two groups and one-way ANOVA
followed by the Tukey's post hoc test was used to analyze the
significance among more than two groups. Survival analysis was
investigated using Kaplan-Meier method followed by the
Gehan-Breslow-Wilcoxon test which was used to analyze the
significance. The correlation between miR-454-3p and TGFB2
expression were evaluated using Spearman's analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-454-3p is downregulated in NSCLC
tissues and cell lines
To understand the role of miR-454-3p in NSCLC
progression, the expression level of miR-454-3p in NSCLC tissues
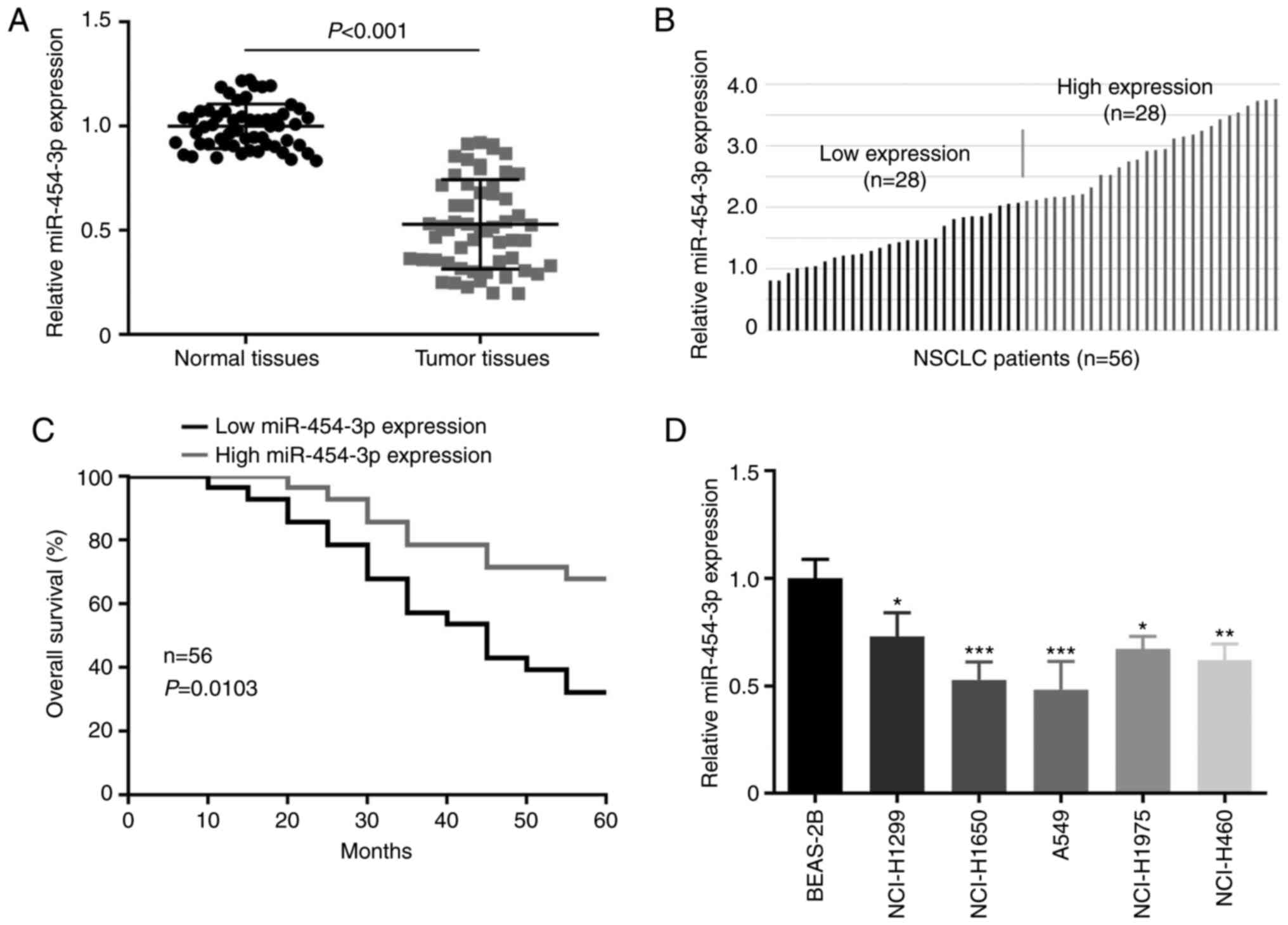
and cells was examined by RT-qPCR assay. As presented in Fig. 1A, miR-454-3p was downregulated in
NSCLC tissues by comparison with adjacent normal tissues. In the
clinicopathological analysis of NSCLC patients, miR-454-3p was
significantly associated with tumor size, pathological stage and
tumor metastasis, but not with age and sex (Table I). Then, the association between
overall survival and miR-454-3p expression in NSCLC patients was
analyzed. The 56 NSCLC tissues were divided into a low-miR-454-3p
expression group and a high-miR-454-3p expression group (Fig. 1B). Kaplan-Meier analysis revealed that
the low miR-454-3p expression was associated with worse overall
survival in patients with NSCLC (Fig.
1C). The expression of miR-454-3p in NSCLC cells was also
detected. The RT-qPCR assay revealed that miR-454-3p expression was
significantly lower in 5 NSCLC cell lines (NCI-H1299, NCI-H1650,
A549, NCI-H1975 and NCI-H460) than in normal lung cell line BEAS-2B
(Fig. 1D).
 | Table I.Association between miR-454-3p and
clinicopathological characteristics of patients with NSCLC. |
Table I.
Association between miR-454-3p and
clinicopathological characteristics of patients with NSCLC.
|
|
| Expression level of
miR-454-3p |
|---|
|
|
|
|
|---|
| Pathological
parameters | n | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.7825 |
|
<50 | 21 | 10 | 11 |
|
|
≥50 | 35 | 18 | 17 |
|
| Sex |
|
|
|
|
|
Male | 30 | 14 | 16 | 0.5920 |
|
Female | 26 | 14 | 12 |
|
| Tumor size |
|
|
| 0.0331a |
| <3
cm | 27 | 17 | 10 |
|
| ≥3
cm | 29 | 10 | 19 |
|
| Pathological
stage |
|
|
| 0.0044b |
|
Well | 20 | 14 | 6 |
|
|
Poor | 36 | 11 | 25 |
|
| Metastasis |
|
|
| 0.0014b |
|
Negative | 31 | 22 | 9 |
|
|
Positive | 25 | 7 | 18 |
|
miR-454-3p inhibits the proliferation
of NSCLC cells
A549 and NCI-H1650 cells were transfected with NC or
miR-454-3p mimics. The transfection efficiency was determined by
RT-qPCR and it was revealed that the expression of miR-454-3p was
significantly increased in both A549 and NCI-H1650 cells after
transfection with miR-454-3p mimics (Fig.
2A). Then, CCK-8 and EdU cell proliferation assays were
performed to investigate the effect of miR-454-3p overexpression on
NSCLC cell proliferation. The CCK-8 assay revealed that miR-454-3p
overexpression significantly suppressed the proliferation rate of
A549 and NCI-H1650 cells (Fig. 2B and
C). EdU assay also revealed that the overexpression of
miR-454-3p markedly reduced the number of EdU-positive cells
(Fig. 2D and E). These results
provided evidence that miR-454-3p functions as a tumor suppressor
and inhibits the proliferation of NSCLC cells.
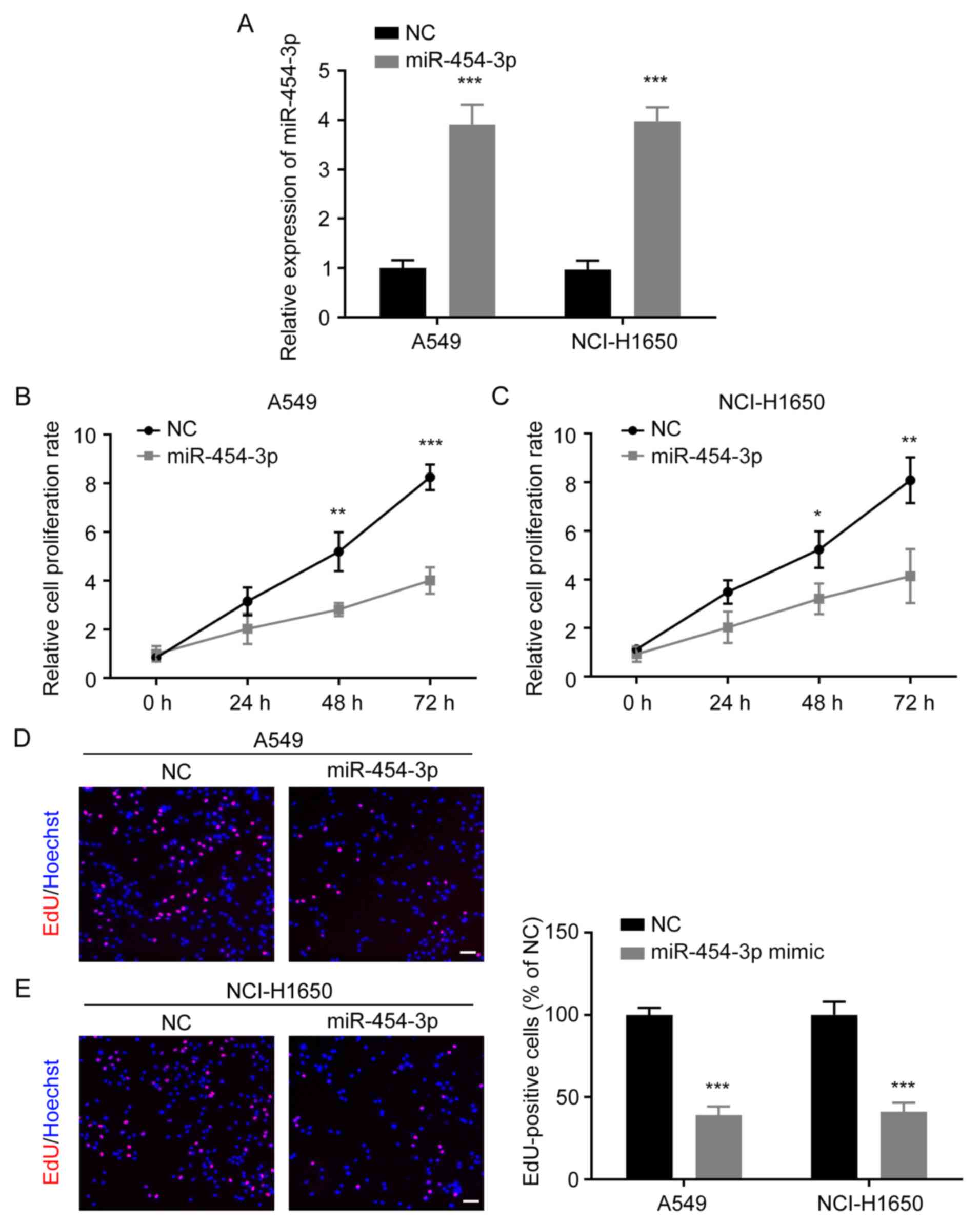 | Figure 2.Ectopic expression of miR-454-3p
suppresses NSCLC cell proliferation. (A) The expression of
miR-454-3p in NSCLC cells transfected with miR-454-3p mimics was
evaluated by qPCR assay. (B and C) A549 and NCI-H1650 cells were
transfected with miR-454-3p mimics for 24 h, and the cell
proliferation rate was determined by CCK-8 assay. (D and E) The
effect of miR-454-3p on A549 and NCI-H1650 cell proliferation was
detected by EdU cell proliferation kit. EdU-positive cells were
photographed under an inverted fluorescence microscope. Scale bar,
100 µm. Data are presented as the mean ± SD, n=3. *P<0.05,
**P<0.01 and ***P<0.001 vs. cells transfected with the
NC-mimic group. miR, microRNA; NSCLC, non-small cell lung cancer;
qPCR, quantitative PCR; CCK-8, Cell Counting Kit-8; EdU,
5-ethynyl-2-deoxyuridine; NC, negative control mimics; miR-454-3p,
miR-454-3p mimics. |
miR-454-3p attenuates metastases and
suppresses EMT in NSCLC cells
To further determine the biological effect of
miR-454-3p on NSCLC cell metastasis, wound-healing and Transwell
invasion assays were conducted. The wound-healing assay revealed
that the migratory capability of A549 and NCI-H1650 cells was
attenuated when miR-454-3p was overexpressed (Fig. 3A and B). Furthermore, miR-454-3p
significantly decreased the number of invasive A549 and NCI-H1650
cells (Fig. 3C). Next, the role of
miR-454-3p in EMT processes was evaluated. The present results
revealed that the overexpression of miR-454-3p led to a decrease in
mesenchymal marker N-cadherin and increased the expression of
epithelial marker E-cadherin (Fig. 3D and
E). Collectively, these data indicated that miR-454-3p may
block EMT to inhibit NSCLC cell metastasis.
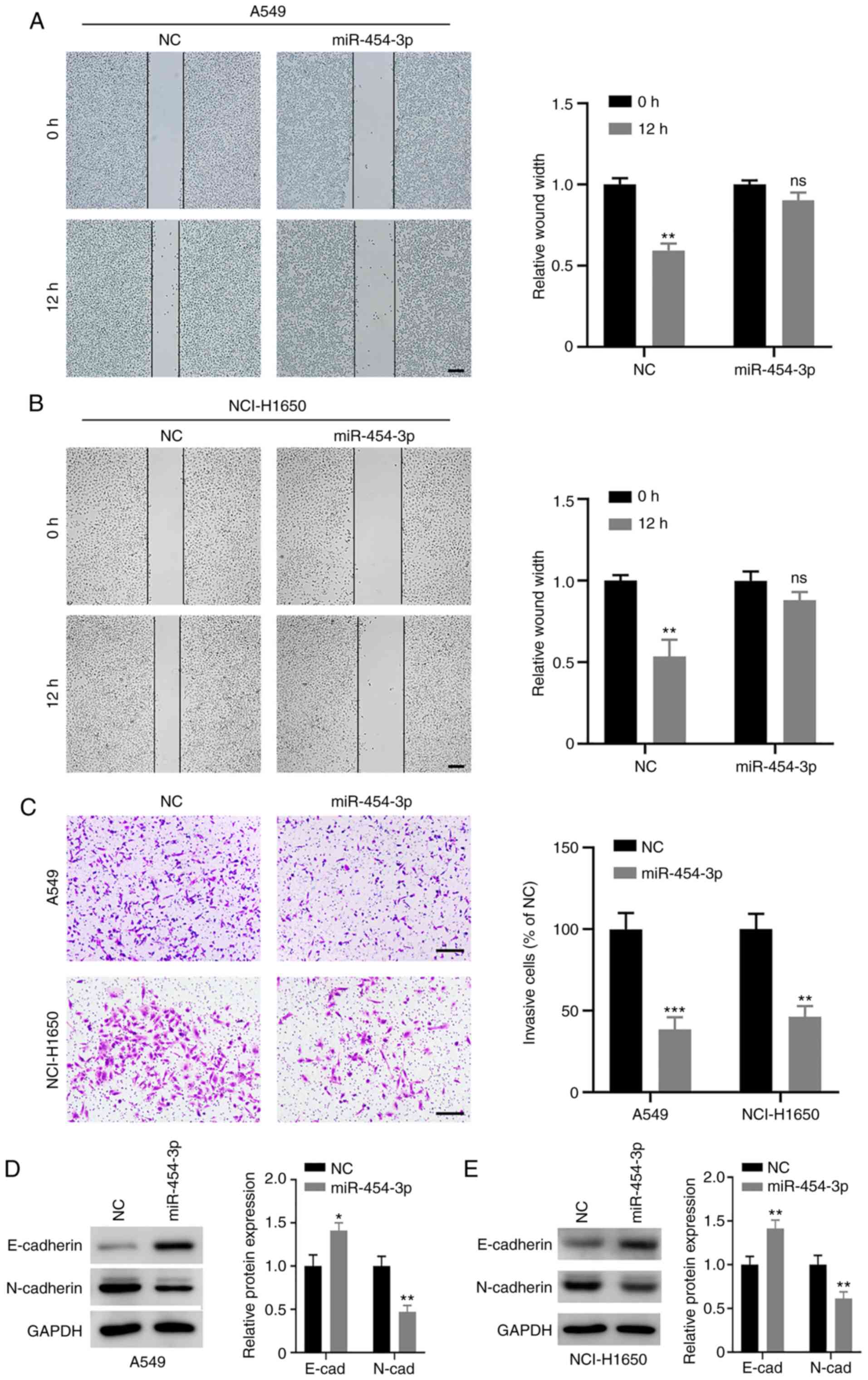 | Figure 3.Effect of miR-454-3p on migration,
invasion, and EMT in NSCLC cells. (A and B) A549 and NCI-H1650
cells were transfected with NC or miR-454-3p mimics, and the
migration capacity of cells was assessed with a wound healing
assay. Scale bar, 200 µm. (C) The effect of miR-454-3p
overexpression on NSCLC cell invasion was examined using a
Transwell invasion assay. Scale bar, 200 µm. (D and E) The protein
levels of EMT markers in A549 and NCI-H1650 cells after
transfection with NC or miR-454-3p mimics were determined by
western blotting. Data are presented as the mean ± SD, n=3.
*P<0.05, **P<0.01 and ***P<0.001 compared with the NC
group. miR, microRNA; EMT, epithelial-mesenchymal transition;
NSCLC, non-small cell lung cancer; NC, negative control mimics; ns,
not significant; miR-454-3p, miR-454-3p mimics. |
TGFB2 is a direct target of miR-454-3p
in NSCLC
TargetScan was selected for bioinformatics analysis
to identify a potential target gene of miR-454-3p. The results
suggested that TGFB2 is a putative target of miR-454-3p (Fig. 4A). To confirm whether the 3′-UTR of
TGFB2 could be directly targeted by miR-454-3p, a luciferase
reporter assay was performed. The results revealed that miR-454-3p
overexpression resulted in significantly decreased the luciferase
activity in the WT 3′-UTR of TGFB2, but not the Mut reporter
(Fig. 4B and C). In addition, RT-qPCR
assays were performed to investigate the mRNA levels of TGFB2 in
A549 and NCI-H1650 cells transfected with NC or miR-454-3p mimics.
As revealed in Fig. 4D and E,
overexpression of miR-454-3p significantly attenuated TGFB2 mRNA
expression. Consequently, these results demonstrated that TGFB2 is
a direct target of miR-454-3p in NSCLC cells.
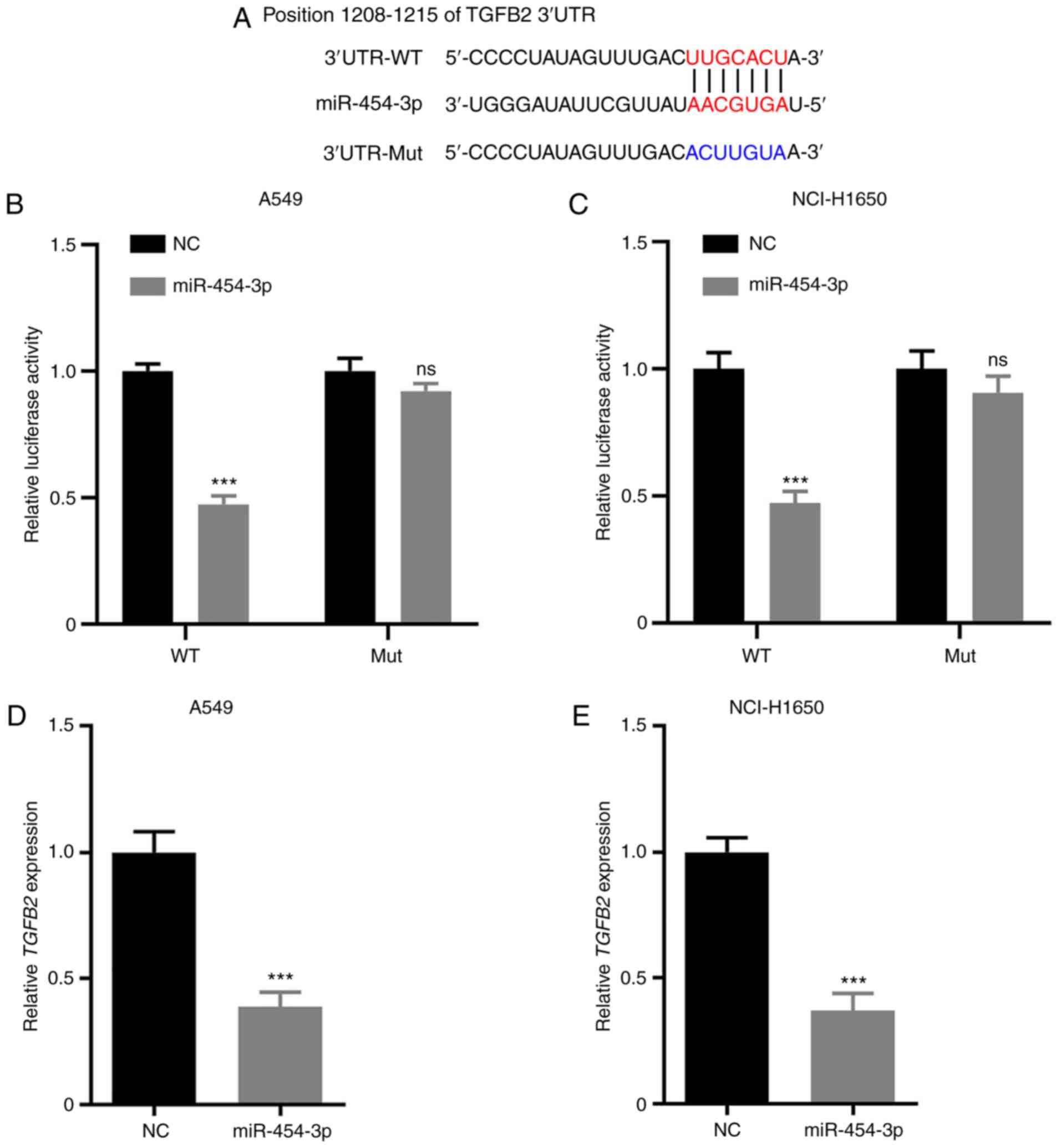 | Figure 4.TGFB2 is a direct target of
miR-454-3p in NSCLC cells. (A) The predicted seed region of
miR-454-3p at the 3′UTR of TGFB2 was analyzed with TargetScan
software and the mutated version is presented. (B and C) A549 and
NCI-H1650 cells were co-transfected with miR-454-3p mimics and
TGFB2-WT or TGFB2-Mut for 48 h. Then, the luciferase activity was
detected using the Dual Luciferase Reporter Assay kit. (D and E)
The TGFB2 mRNA expression in A549 and NCI-H1650 cells transfected
with NC or miR-454-3p mimics was assessed by RT-qPCR. Data are
expressed as the mean ± SD, n=3. ***P<0.001 compared with NC
group. TGFB2, transforming growth factor-β2; miR, microRNA; NSCLC,
non-small cell lung cancer; WT, wild-type; Mut, mutant; NC,
negative control mimics; RT-qPCR, reverse
transcription-quantitative PCR; ns, not significant; miR-454-3p,
miR-454-3p mimics. |
miR-454-3p is negatively correlated
with TGFB2 expression in NSCLC tissues
To determine the association between miR-454-3p and
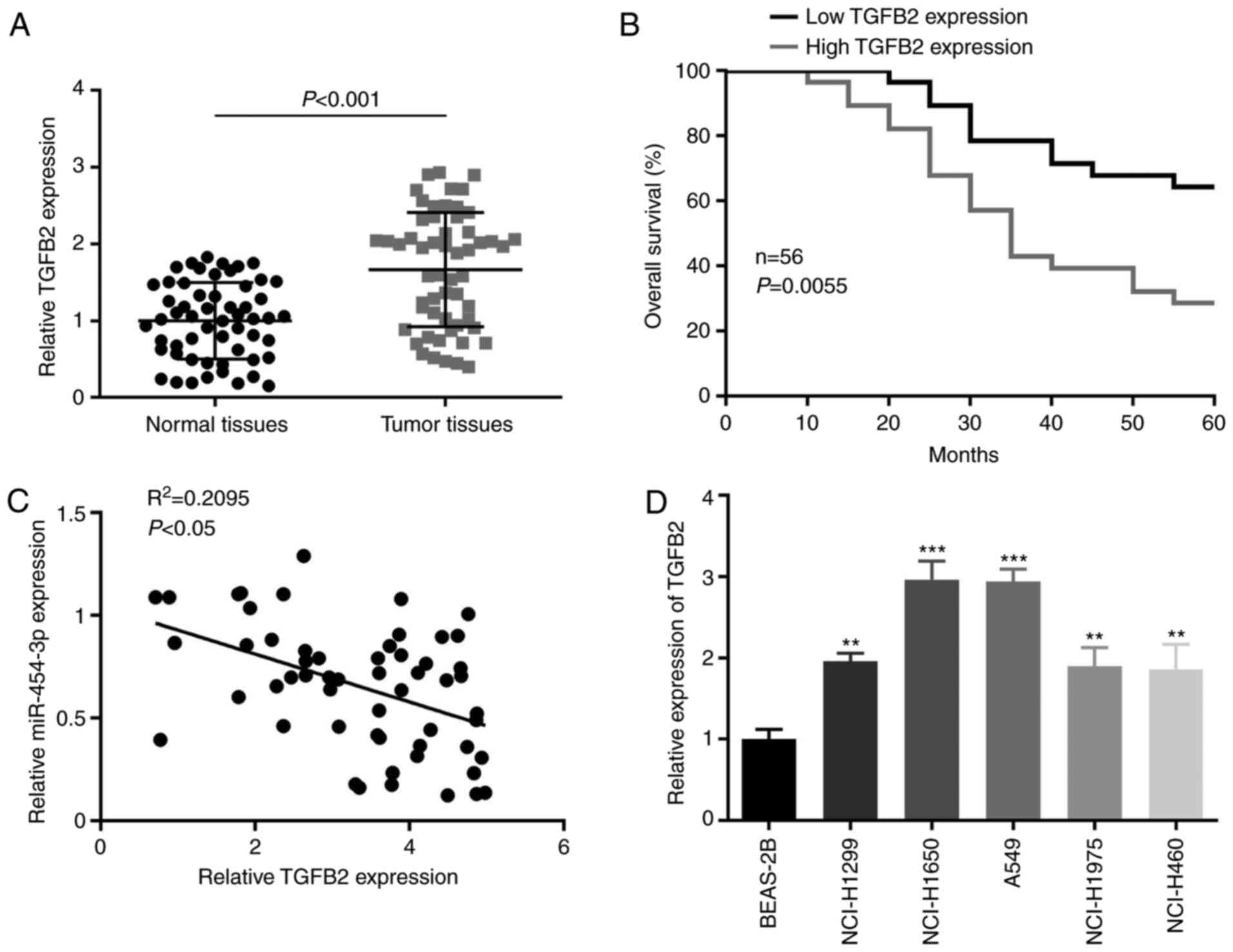
TGFB2 in NSCLC tissues, the expression of TGFB2 in NSCLC tissues
and adjacent non-cancerous tissues was examined. It was revealed
that the expression level of TGFB2 was significantly higher in
NSCLC tissues than in non-tumor tissues (Fig. 5A) and high TGFB2 expression indicated
poor prognosis in patients with NSCLC (Fig. 5B). In addition, it was observed that
the expression of miR-454-3p had a negative correlation with TGFB2
expression in NSCLC tissues (Fig.
5C). Furthermore, when compared with normal lung cell line
BEAS-2B, TGFB2 expression was significantly higher in all five
NSCLC cell lines (NCI-H1299, NCI-H1650, A549, NCI-H1975 and
NCI-H460) (Fig. 5D). Collectively,
the present results revealed that TGFB2 expression was negatively
correlated with miR-454-3p in NSCLC tissues and cells, which may
confirm that TGFB2 could be a target of miR-454-3p.
TGFB2 overexpression abrogates
miR-454-3p-mediated inhibition of NSCLC cell proliferation
Next, the effect of TGFB2 and miR-454-3p on NSCLC
cell proliferation was investigated. First, A549 and NCI-H1650
cells were co-transfected with TGFB2-overexpressing plasmid and
miR-454-3p mimics. RT-qPCR assays revealed that TGFB2 was
significantly upregulated in A549 and NCI-H1650 cells after
transfection with TGFB2 overexpression plasmid, which could be
significantly inhibited by miR-454-3p mimics (Fig. 6A and B). Then, CCK-8 and EdU cell
proliferation assays were performed to examine the proliferation of
NSCLC cells. The cell proliferation rate was significantly
increased in NSCLC cells co-transfected with TGFB2 plasmid and
miR-454-3p mimics compared with cells transfected with miR-454-3p
mimics alone (Fig. 6C and D).
Furthermore, the number of EdU-positive A549 and NCI-H1650 cells in
the co-transfection groups was significantly higher than cells
transfected with miR-454-3p mimics alone (Fig. 6E and F). The results indicated that
the overexpression of TGFB2 abrogated the miR-454-3p-mediated
inhibitory effect on NSCLC cell proliferation.
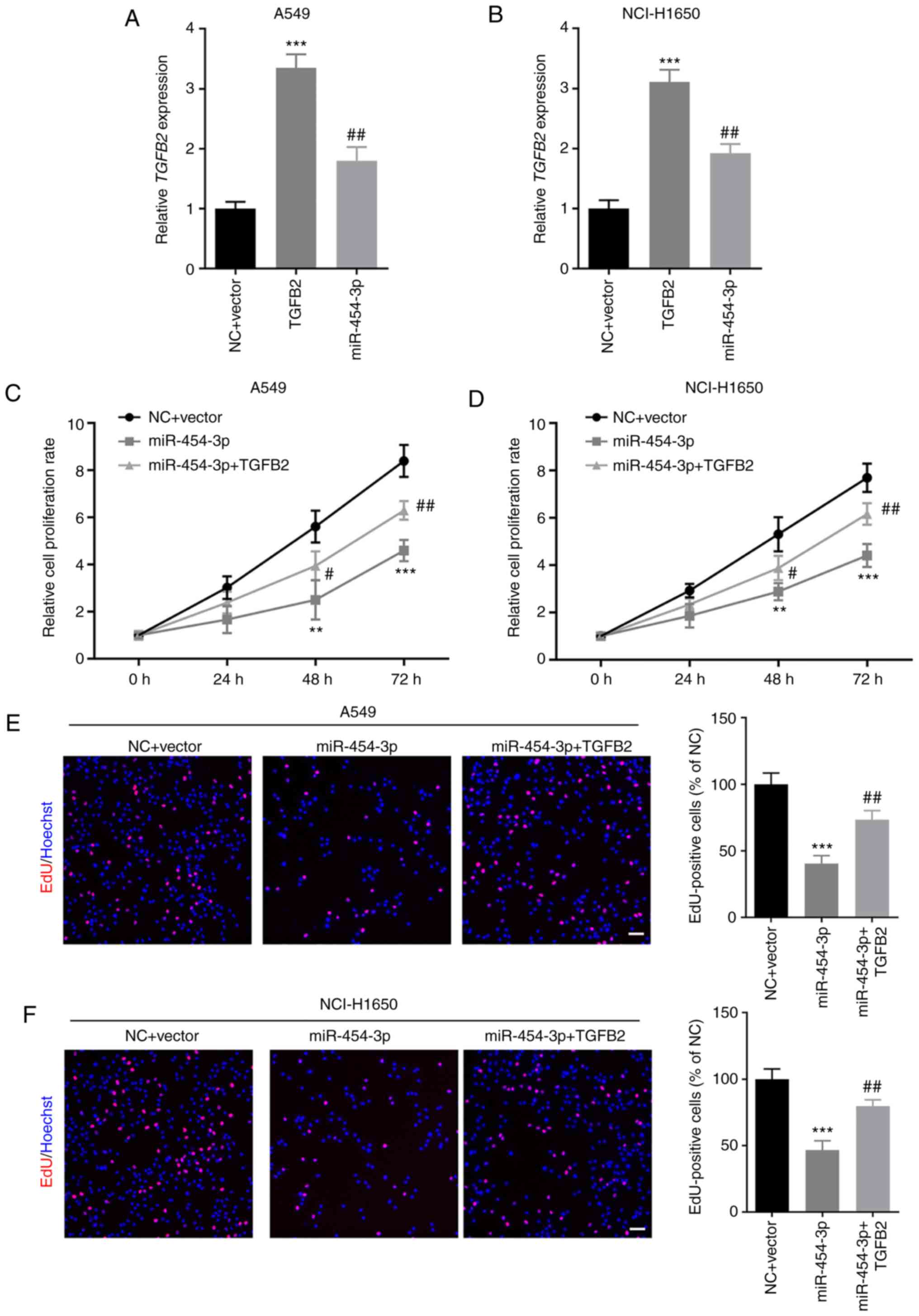 | Figure 6.TGFB2 overexpression counteracts the
miR-454-3p-mediated suppressive effects on NSCLC cell
proliferation. (A and B) A549 and NCI-H1650 cells were
co-transfected with TGFB2-overexpressing plasmid and miR-454-3p
mimics and then the expression of TGFB2 was determined by RT-qPCR
assay. (C-F) The effect of miR-454-3p and TGFB2 on the
proliferation of A549 and NCI-H1650 cells was examined by (C and D)
CCK-8 assay and (E and F) EdU cell proliferation assay. Scale bar,
100 µm. Data are presented as the mean ± SD, n=3. **P<0.01 and
***P<0.001 compared with cells transfected with the NC + Vector
group. #P<0.05 and ##P<0.01 compared
with cells transfected with miR-454-3p mimics. TGFB2, transforming
growth factor-β2; miR, microRNA; NSCLC, non-small cell lung cancer;
RT-qPCR, reverse transcription-quantitative PCR; CCK-8, Cell
Counting Kit-8; EdU, 5-ethynyl-2-deoxyuridine; NC, negative control
mimics; miR-454-3p, miR-454-3p mimics. |
TGFB2 reverses miR-454-3p-mediated
inhibition of metastasis and EMT in NSCLC cells
Subsequently, the role of TGFB2 in
miR-454-3p-mediated NSCLC cell metastasis and EMT were evaluated.
As revealed in Fig. 7A-D, the number
of migrated and invasive cells was markedly decreased in A549 and
NCI-H1650 cells transfected miR-454-3p mimics, which could be
reversed by overexpression of TGFB2. In addition, TGFB2
overexpression decreased the expression E-cadherin and increased
the expression of N-cadherin in A549 and NCI-H1650 cells (Fig. 7E and F). Collectively, these data
demonstrated that miR-454-3p inhibited the metastasis and EMT of
NSCLC cells via antagonizing TGFB2 expression.
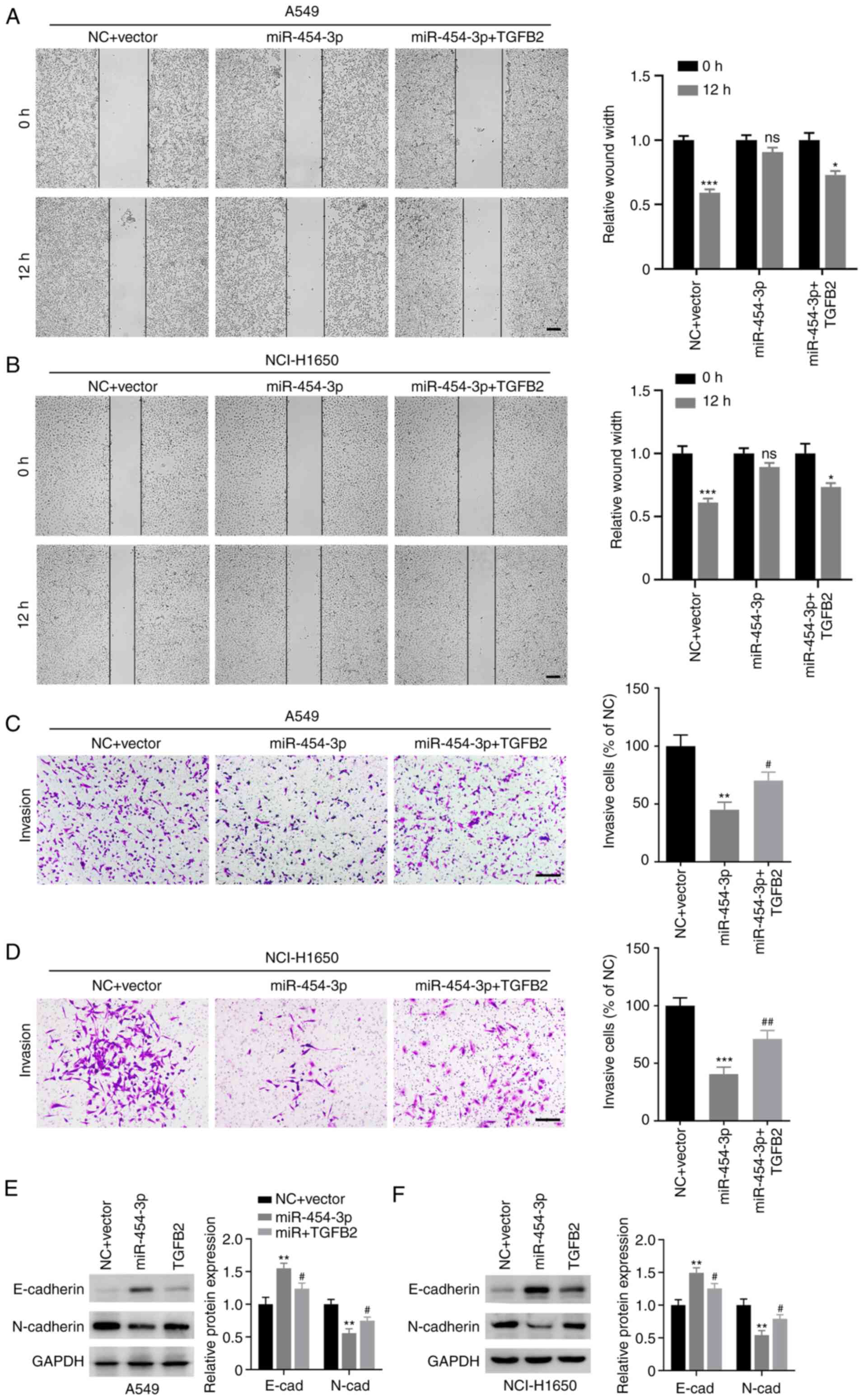 | Figure 7.TGFB2 reverses miR-454-3p-mediated
inhibition of migration, invasion and EMT in NSCLC cells. A549 and
NCI-H1650 cells were transfected with TGFB2-overexpressing plasmid
and miR-454-3p mimics. (A and B) A wound healing assay was
performed to evaluate cell migration capacity. Scale bar, 200 µm.
(C and D) Transwell invasion assay was conducted to investigate
cell invasion capacity. Scale bar, 200 µm. (E and F) Western blot
analysis was used to reveal the expression of E-cadherin and
N-cadherin in cells after the indicated transfections. Data are
presented as the mean ± SD, n=3. *P<0.05, **P<0.01 and
***P<0.001 vs. cells transfected with the NC + Vector group.
#P<0.05 and ##P<0.01 vs. cells
transfected with miR-454-3p mimics. TGFB2, transforming growth
factor-β2; miR, microRNA; EMT, epithelial-mesenchymal transition;
NSCLC, non-small cell lung cancer; NC, negative control mimics;
miR-454-3p, miR-454-3p mimics; ns, no significance. |
Discussion
NSCLC is closely associated with high cancer-related
incidence and fatality rate (25).
Challenges remain due to failure of early diagnosis of the cancer,
metastasis or drug resistance (26–28). The
last two decades have witnessed the rapid developments of tumor
epigenetic alternations (29,30). miRNAs play an important role in
tumorigenesis and development (31,32). It is
urgent to determine the expression, functions and potential targets
of dysregulated miRNAs in NSCLC, which could provide effective
strategies to improve the treatment and prognosis of patients.
It has been reported that miR-454-3p was
downregulated in various cancers and its ectopic expression
impaired tumor malignant behaviors (12,33). For
example, miR-454-3p suppressed cell proliferation and induced
apoptosis by directly downregulating nuclear factor of activated
T-cells c2 (NFATc2) in glioblastoma (GBM) (12). It also targeted cytoplasmic
polyadenylation element-binding protein 1 (CPEB1) to inhibit GBM
cell proliferation and mobility (34). For these reasons, it may be regarded
as a tumor suppressor and a novel biomarker for GBM therapy. In
addition, low miR-454-3p expression in bladder cancer tissues,
predicted poor prognosis of bladder cancer patients (14). Moreover, miR-454-3p functioned as a
tumor inhibitor of esophageal cancer by inhibiting the ERK/AKT
signaling pathways (35). With regard
to NSCLC, the expression of miR-454-3p was downregulated in NSCLC
tissues and cell lines (13). Further
study suggested that aberrant expression of miR-454-3p
significantly restrained NSCLC cell proliferation and accelerated
cell apoptosis by downregulating calbindin 1 (CALB1) (13). These studies indicated that miR-454-3p
could be a tumor-suppressive miRNA as revealed in a great number of
cancer types. Conversely, it was observed to be upregulated in
breast cancer and further enhanced cell metastasis, leading to a
shorter relapse-free survival of patients with breast cancer
(36). Song et al expounded
that miR-454-3p depletion reduced cervical cancer cell growth,
facilitating cancer cell apoptosis (37). Thus, miR-454-3p can function as either
a tumorigenic or tumor suppressive miRNA, which may depend on the
type of cancer. In present study, the expression of miR-454-3p was
revealed to be decreased in NSCLC tissues and cells. Further
mechanistic studies suggested that its aberrant expression
attenuated migration and invasion and had an impact on E-cadherin
and N-cadherin expression in NSCLC cells.
To gain insight into the mechanism of miR-454-3p in
NSCLC, TargetScan database was applied to predict the potential
target genes of miR-454-3p. Among the candidate genes, a
complementary miR-454-3p site was present in the 3′-UTR of TGFB2
mRNA. Previous evidence has revealed the involvement of TGFB2 in
numerous malignancies (38,39). As a result, TGFB2 was selected for
subsequent investigations. Dual luciferase reporter assays revealed
that TGFB2 could directly bind to miR-454-3p in NSCLC cells. TGFB2
is known as an isoform of the TGFB family. TGFBs could function as
oncogenic genes and bind with a range of TGFB receptors and
activate transcription factors such as the SMAD superfamily members
which regulate cell homeostasis (39–41). TGFBs
(TGFB1, TGFB2, and TGFB3) were upregulated in gastric carcinoma,
however, only high expression of TGFB2 was associated with poor
prognosis of gastric carcinoma patients (42). Schlingensiepen et al also
reported that the high expression of TGFB2 was revealed in
high-grade gliomas and indicated an unfavorable prognosis of glioma
patients (43). Moreover, TGFB2 could
induce EMT and promote tumor metastasis in nasopharyngeal carcinoma
(44) and gallbladder carcinoma
(45). The major biological function
of miRNAs is to regulate the expression of genes by
post-transcriptional mRNA regulation (31). Some miRNAs have been reported to
regulate TGFB2 in malignant tumors. miR-200a suppressed renal cell
carcinoma development by directly targeting TGFB2 (46). A recent study revealed that TGFB2
downregulation mediated by miR-324-5p mimics could inhibit
gallbladder carcinoma cell migration, invasion and EMT, which
contributed to suppression of gallbladder carcinoma metastatic
behaviors (45). In the present
study, the results revealed that TGFB2 was markedly upregulated in
NSCLC tissues and cell lines. Moreover, it was negatively
correlated with miR-454-3p in NSCLC tissues. Further research
corroborated that TGFB2 reversed miR-454-3p-mediated migration,
invasion and E-cadherin and N-cadherin expression in NSCLC
cells.
TGFB2 is a hallmark in various malignant tumors, and
treatment of cancer based on the specific inhibition of TGFB2 is
currently being developed (17). The
mode of action, effectuality and tolerability in vitro and
in vivo of Trabedersen (AP-12009), an antisense
oligodeoxynucleotide inhibitor targeting TGFB2, has been confirmed
(47). Furthermore, AP-12009 is being
evaluated in a phase I/II dose escalation study with high-grade
glioma, metastatic colorectal carcinoma, metastatic melanoma, and
advanced pancreatic carcinoma (47,48). In
present study, the regulatory effect of miR-454-3p/TGFB2
concatenation on NSCLC malignant behaviors in vitro was
confirmed. However, the study of miR-454-3p and TGFB2 in
vivo remains to be further investigated. In addition, studies
using TGFB2 inhibitors can be carried out in vitro and in
vivo, which may provide a more rationale and potential
treatment target for NSCLC patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets generated and analyzed during the study
are available from the corresponding author upon reasonable
request.
Authors' contributions
HL and RW conceived and designed this study. YL and
LK collected the samples and performed the experiments. YX and TY
analyzed and verified the data. HL wrote the manuscript. RW revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The human tissue used in the present study were
obtained from Yuebei People's Hospital of Shaoguan (Shaoguan,
China). The data were published after internal ethical approval and
patient consent were obtained. The proposed experiments and
procedures were reviewed and approved by the Ethics Committee of
Yuebei People's Hospital. The present study was performed according
to the Declaration of Helsinki of 1975 and its succeeding
amendments.
Patient consent for publication
Not applicable.
Competing interests
The author declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Didkowska J, Wojciechowska U, Mańczuk M
and Łobaszewski J: Lung cancer epidemiology: Contemporary and
future challenges worldwide. Ann Transl Med. 4:1502016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jonna S and Subramaniam DS: Molecular
diagnostics and targeted therapies in non-small cell lung cancer
(NSCLC): An update. Discov Med. 27:167–170. 2019.PubMed/NCBI
|
|
4
|
Foster CC, Pitroda SP and Weichselbaum RR:
Beyond palliation: The rationale for metastasis-directed therapy
for metastatic non-small cell lung cancer. J Thorac Oncol.
14:1510–1512. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galagali H and Kim JK: The multifaceted
roles of microRNAs in differentiation. Curr Opin Cell Biol.
67:118–140. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee TJ, Yuan X, Kerr K, Yoo JY, Kim DH,
Kaur B and Eltzschig HK: Strategies to modulate MicroRNA functions
for the treatment of cancer or organ injury. Pharmacol Rev.
72:639–667. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu M, Wang G, Tian W, Deng Y and Xu Y:
miRNA-based therapeutics for lung cancer. Curr Pharm Des.
23:5989–5996. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan Y, Liao H, Pu Q, Ke X, Hu X, Ma Y,
Luo X, Jiang Q, Gong Y, Wu M, et al: miR-410 induces both
epithelial-mesenchymal transition and radioresistance through
activation of the PI3K/mTOR pathway in non-small cell lung cancer.
Signal Transduct Target Ther. 5:852020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun X, Xu M, Liu H and Ming K:
MicroRNA-219 is downregulated in non-small cell lung cancer and
inhibits cell growth and metastasis by targeting HMGA2. Mol Med
Rep. 16:3557–3564. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xing F, Song Z and He Y: miR-219-5p
inhibits growth and metastasis of ovarian cancer cells by targeting
HMGA2. Biol Res. 51:502018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shao Y, Liang B, Long F and Jiang SJ:
Diagnostic MicroRNA biomarker discovery for non-small-cell lung
cancer adenocarcinoma by integrative bioinformatics analysis.
Biomed Res Int. 2017:25630852017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zuo J, Yu H, Xie P, Liu W, Wang K and Ni
H: miR-454-3p exerts tumor-suppressive functions by down-regulation
of NFATc2 in glioblastoma. Gene. 710:233–239. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jin C, Lin T and Shan L: Downregulation of
calbindin 1 by miR-454-3p suppresses cell proliferation in nonsmall
cell lung cancer in vitro. Cancer Biother Radiopharm. 34:119–127.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang S, Zhang G, Zheng W, Xue Q, Wei D,
Zheng Y and Yuan J: miR-454-3p and miR-374b-5p suppress migration
and invasion of bladder cancer cells through targetting ZEB2.
Biosci Rep. 38:BSR201814362018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fan Y, Shi C, Li T and Kuang T:
microRNA-454 shows anti-angiogenic and anti-metastatic activity in
pancreatic ductal adenocarcinoma by targeting LRP6. Am J Cancer
Res. 7:139–147. 2017.PubMed/NCBI
|
|
16
|
Abraham CG, Ludwig MP, Andrysik Z, Pandey
A, Joshi M, Galbraith MD, Sullivan KD and Espinosa JM: ΔNp63α
suppresses TGFB2 expression and RHOA activity to drive cell
proliferation in squamous cell carcinomas. Cell Rep. 24:3224–3236.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang B, Bai J, Shi R, Shao X, Yang Y, Jin
Y, Che X, Zhang Y, Qu X, Liu Y and Li Z: TGFB2 serves as a link
between epithelial-mesenchymal transition and tumor mutation burden
in gastric cancer. Int Immunopharmacol. 84:1065322020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dave H, Trivedi S, Shah M and Shukla S:
Transforming growth factor beta 2: A predictive marker for breast
cancer. Indian J Exp Biol. 49:879–887. 2011.PubMed/NCBI
|
|
19
|
Bruna A, Darken RS, Rojo F, Ocaña A,
Peñuelas S, Arias A, Paris R, Tortosa A, Mora J, Baselga J and
Seoane J: High TGFbeta-Smad activity confers poor prognosis in
glioma patients and promotes cell proliferation depending on the
methylation of the PDGF-B gene. Cancer Cell. 11:147–160. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mittal V: Epithelial mesenchymal
transition in aggressive lung cancers. Adv Exp Med Biol. 890:37–56.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Singh M, Yelle N, Venugopal C and Singh
SK: EMT: Mechanisms and therapeutic implications. Pharmacol Ther.
182:80–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang H, Brown CW and Matzuk MM: Genetic
analysis of the mammalian transforming growth factor-beta
superfamily. Endocr Rev. 23:787–823. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodriguez LG, Wu X and Guan JL:
Wound-healing assay. Methods Mol Biol. 294:23–29. 2005.PubMed/NCBI
|
|
25
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hung JJ, Jeng WJ, Hsu WH, Chou TY, Huang
BS and Wu YC: Predictors of death, local recurrence, and distant
metastasis in completely resected pathological stage-I
non-small-cell lung cancer. J Thorac Oncol. 7:1115–1123. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee WC, Reuben A, Hu X, McGranahan N, Chen
R, Jalali A, Negrao MV, Hubert SM, Tang C, Wu CC, et al: Multiomics
profiling of primary lung cancers and distant metastases reveals
immunosuppression as a common characteristic of tumor cells with
metastatic plasticity. Genome Biol. 21:2712020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lim ZF and Ma PC: Emerging insights of
tumor heterogeneity and drug resistance mechanisms in lung cancer
targeted therapy. J Hematol Oncol. 12:1342019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Diaz-Lagares A, Mendez-Gonzalez J, Hervas
D, Saigi M, Pajares MJ, Garcia D, Crujerias AB, Pio R, Montuenga
LM, Zulueta J, et al: A novel epigenetic signature for early
diagnosis in lung cancer. Clin Cancer Res. 22:3361–3371. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Duruisseaux M and Esteller M: Lung cancer
epigenetics: From knowledge to applications. Semin Cancer Biol.
51:116–128. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iqbal MA, Arora S, Prakasam G, Calin GA
and Syed MA: MicroRNA in lung cancer: Role, mechanisms, pathways
and therapeutic relevance. Mol Aspects Med. 70:3–20. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu KL, Tsai YM, Lien CT, Kuo PL and Hung
AJ: The roles of MicroRNA in lung cancer. Int J Mol Sci.
20:16112019. View Article : Google Scholar
|
|
33
|
Jiang D, Li H, Xiang H, Gao M, Yin C, Wang
H, Sun Y and Xiong M: Long chain non-coding RNA (lncRNA) HOTAIR
knockdown increases miR-454-3p to suppress gastric cancer growth by
targeting STAT3/cyclin D1. Med Sci Monit. 25:1537–1548. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hui X, Zhang S and Wang Y: miR-454-3p
suppresses cell migration and invasion by targeting CPEB1 in human
glioblastoma. Mol Med Rep. 18:3965–3972. 2018.PubMed/NCBI
|
|
35
|
Yan A, Wang C, Zheng L, Zhou J and Zhang
Y: MicroRNA-454-3p inhibits cell proliferation and invasion in
esophageal cancer by targeting insulin-like growth factor 2
mRNA-binding protein 1. Oncol Lett. 20:3592020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ren L, Chen H, Song J, Chen X, Lin C,
Zhang X, Hou N, Pan J, Zhou Z, Wang L, et al: miR-454-3p-mediated
Wnt/β-catenin signaling antagonists suppression promotes breast
cancer metastasis. Theranostics. 9:449–465. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song Y, Guo Q, Gao S and Hua K: miR-454-3p
promotes proliferation and induces apoptosis in human cervical
cancer cells by targeting TRIM3. Biochem Biophys Res Commun.
516:872–879. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Strauss J, Heery CR, Schlom J, Madan RA,
Cao L, Kang Z, Lamping E, Marté JL, Donahue RN, Grenga I, et al:
Phase I trial of M7824 (MSB0011359C), a bifunctional fusion protein
targeting PD-L1 and TGFβ, in advanced solid tumors. Clin Cancer
Res. 24:1287–1295. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Derynck R, Akhurst RJ and Balmain A:
TGF-beta signaling in tumor suppression and cancer progression. Nat
Genet. 29:117–129. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moustakas A and Heldin CH: Mechanisms of
TGFβ-induced epithelial-mesenchymal transition. J Clin Med.
5:632016. View Article : Google Scholar
|
|
42
|
Vagenas K, Spyropoulos C, Gavala V and
Tsamandas AC: TGFbeta1, TGFbeta2, and TGFbeta3 protein expression
in gastric carcinomas: Correlation with prognostics factors and
patient survival. J Surg Res. 139:182–188. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Schlingensiepen KH, Schlingensiepen R,
Steinbrecher A, Hau P, Bogdahn U, Fischer-Blass B and Jachimczak P:
Targeted tumor therapy with the TGF-beta 2 antisense compound AP
12009. Cytokine Growth Factor Rev. 17:129–139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao L, Lin L, Pan C, Shi M, Liao Y, Bin J
and Liao W: Flotillin-2 promotes nasopharyngeal carcinoma
metastasis and is necessary for the epithelial-mesenchymal
transition induced by transforming growth factor-β. Oncotarget.
6:9781–9793. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang X, Zhang L, Chen M and Liu D:
miR-324-5p inhibits gallbladder carcinoma cell metastatic
behaviours by downregulation of transforming growth factor beta 2
expression. Artif Cells Nanomed Biotechnol. 48:315–324. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lu R, Ji Z, Li X, Qin J, Cui G, Chen J,
Zhai Q, Zhao C, Zhang W and Yu Z: Tumor suppressive microRNA-200a
inhibits renal cell carcinoma development by directly targeting
TGFB2. Tumour Biol. 36:6691–6700. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jaschinski F, Rothhammer T, Jachimczak P,
Seitz C, Schneider A and Schlingensiepen KH: The antisense
oligonucleotide trabedersen (AP 12009) for the targeted inhibition
of TGF-β2. Curr Pharm Biotechnol. 12:2203–2213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bogdahn U, Hau P, Stockhammer G,
Venkataramana NK, Mahapatra AK, Suri A, Balasubramaniam A, Nair S,
Oliushine V, Parfenov V, et al: Targeted therapy for high-grade
glioma with the TGF-β2 inhibitor trabedersen: Results of a
randomized and controlled phase IIb study. Neuro Oncol. 13:132–142.
2011. View Article : Google Scholar : PubMed/NCBI
|