Introduction
Toxicarioside G (TCG) is a cardenolide isolated from
Calotropis gigantean. Apart from their traditional use in
the treatment of congestive heart failure and arrhythmia,
cardenolides have recently been proven to exert potent anticancer
activities (1–5). Previous studies have demonstrated that
other cardenolides isolated from Calotropis gigantean
exhibited significant cytotoxicity against numerous types of human
cancer cells, such as hepatoma carcinoma, gastric cancer and lung
cancer (6–8). Consistent with these observations, TCG
has been found to exert significant anticancer effects in various
cancer cell types, such as hepatoma carcinoma, gastric cancer and
cervical cancer cell lines (6).
However, to the best of our knowledge, the underlying mechanisms by
which TCG inhibits tumor growth remains poorly understood.
Autophagy is a conserved catabolic process whereby
aggregated proteins or damaged organelles are sequestered by
double-membrane autophagosomes and degraded in autolysosomes, which
maintains cellular homeostasis under physiological conditions
(9,10). Induction of autophagy has been
observed in various types of cancer cells challenged with intra-
and extracellular stresses; however, the role of autophagy in
regulating the fate of cancer cells remains controversial (11–13). On
the one hand, inhibition of autophagy was discovered to promote
cell death or chemosensitivity, suggesting that autophagy is
required for cancer progression (14,15). On
the other hand, activation of autophagy has been found to inhibit
cell proliferation or cell death, indicating that autophagy may
play a tumor-suppressive role (16,17).
Therefore, understanding the role of autophagy and the related
mechanisms in cancer cells is important for enhancing the
effectiveness of cancer treatment.
The Hippo signaling pathway is an evolutionarily
conserved pathway that controls organ size and tissue homeostasis
(18,19). In recent years, the Hippo signaling
pathway has emerged as a key regulator in cancer development and
progression (20). The kinase cascade
of Mst1/2 and large tumor suppressor kinase 1/2 (LATS1/2)
represents a core component of the mammalian Hippo signaling
pathway (21). When the Hippo pathway
is activated by upstream signals, Mst1/2 phosphorylates and
activates LATS1/2, which subsequently phosphorylates Yes1
associated transcriptional regulator (YAP). This phosphorylation
event retains YAP in the cytoplasm, which prevents YAP from being
translocated into the nucleus to interact with the TEA domain
transcription factor 1 family of transcription factors and activate
its downstream genes, such as cellular communication network factor
2 (CTGF) and cellular communication network factor 1 (CCN1)
(22,23). As a key downstream effector of the
Hippo signaling pathway, YAP has been discovered to play important
roles in cell proliferation and the survival of cancer cells. YAP
amplification has been frequently observed in a large number of
human cancer types, including in lung, liver, colon and gastric
cancers (19,24). In addition, the abnormal activation of
YAP was found to be associated with the progression of numerous
types of cancer, such as lun, colorectal and lover cancer
suggesting that YAP may play an important role in tumorigenesis
(24–26).
The aim of the present study was to investigate the
effect of TCG on the SW480 colorectal cancer cell and determine
whether autophagy and Yes1 associated transcriptional regulator
(YAP) were associated with TCG-mediated inhibition of cell
proliferation and viability.
Materials and methods
Cell culture and reagents
The human colorectal cancer cell lines, SW480 and
HT-29, were purchased from the American Type Culture Collection.
The cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Sigma-Aldrich;
Merck KGaA), and maintained at 37°C in a humidified incubator with
5% CO2. The cells were treated with TCG (purity, ≥95%),
which was kindly gifted by Professor HaoFu Dai (Chinese Academy of
Tropical Agricultural Sciences, Haikou, China).
The PI3K inhibitor, 3-methyladenine (3-MA; cat. no.
189490) and chloroquine (CQ; cat. no. C6628) were purchased from
Sigma-Aldrich (Merck KGaA). Verteporfin (VP; cat. no. HY-B0146) was
purchased from MedChemExpress. The cells were treated with 10 mM
3-MA, 5 µM CQ, or 10 µM VP at 37°C for 24 h.
Cytotoxicity assay
Cell cytotoxicity was measured using a MTT assay.
Briefly, SW480 or HT-29 cells were seeded (1×106/well)
into 96-well plates and treated with indicated concentrations of
TCG or DMSO (or 10 mM 3-MA, 5 µM CQ and 10 µM VP) for 24 h at 37°C.
Following incubation, 10 µl MTT was added to each well and the
samples were incubated for a further 4 h at 37°C. Then, 200 µl DMSO
was added to each well to dissolve purple formazan. The absorbance
was measured at 570 nm using an ELISA reader (Bio-Rad Laboratories,
Inc.).
Bromodeoxyuridine (BrdU) incorporation
assay
The SW480 cells were seeded (1×106/well)
into 96-well plates and treated with indicated concentrations of
TCG or DMSO (or 10 mM 3-MA and 10 µM VP) for 24 h. Following
incubation, cell proliferation was determined using a BrdU Cell
Proliferation ELISA kit (cat. no. ab126556; Abcam) according to the
manufacturer's protocol.
Colony formation assay
The SW480 cells were seeded (1×103/well)
into 24-well plates and treated with indicated concentrations of
TCG or DMSO (or 3-MA according to each experiment requires).
Following 7 days of incubation, the colonies were stained with
Giemsa for 15 min at 37°C, then washed three times with PBS. The
visible colonies were visualized using a Molecular Imager Gel Do
XR+ system (Bio-Rad Laboratories, Inc.) and counted using ImageJ
1.47 software (National Institutes of Health).
Lactate dehydrogenase (LDH) release
assay
The SW480 cells were seeded (1×106/well)
into 96-well plates and treated with indicated concentrations of
TCG or DMSO (or 10 mM 3-MA and 10 µM VP) for 24 h. Following
incubation, the cytotoxicity was measured using a LDH release kit
(cat. no. C0016; Beyotime Institute of Biotechnology) according to
the manufacturer's protocol.
Flow cytometry
Briefly, the SW480 cells were seeded
(3×105/well) into 6-well plates and treated with TCG (0,
0.2 and 0.4 µM) for 24 h. Following incubation, the cells were
harvested and washed with PBS, then resuspended in PI/AnnexinV
solution (Nanjing KeyGen Biotech Co., Ltd.). Apoptosis was
subsequently analyzed using a FACSCalibur flow cytometer (BD
Biosciences) and FlowJo v7.6.1 software (FlowJo LLC).
Western blot analysis
Total protein was extracted from the SW480 cells
treated with 0, 0.2 and 0.4 µM TCG (or control, 0.4 µM TCG, 10 mM
3-MA, 0.4 µM TCG+10 mM 3-MA; or control, 0.4 µM TCG, 5 µM CQ, 0.4
µM TCG+5 µM CQ) using RIPA lysis buffer (Thermo Fisher Scientific,
Inc.) and quantified using a BCA protein assay kit (Thermo Fisher
Scientific, Inc.). The proteins were separated using 10% SDS-PAGE
and transferred onto PVDF membranes. After blocking with 5% BSA
(Sigma-Aldrich; Merck KGaA) for 30 min at 37°C, the membranes were
incubated at 4°C overnight with the following primary antibodies:
Anti-poly (ADP-ribose) polymerase 1 (PARP; cat. no. ab191217;
1:1,000; Abcam), anti-caspase-3 (cat. no. ab32351; 1:1,000; Abcam),
anti-LC3B (cat. no. L7543; 1:2,000; Sigma-Aldrich; Merck KGaA),
anti-Beclin1 (cat. no. ab210498; 1:1,000; Abcam), anti-autophagy
related 5 (ATG5; cat. no. ab108327; 1:1,000; Abcam), anti-P62 (cat.
no. ab109012; 1:1,000; Abcam), anti-phosphorylated (p)-YAP (cat.
no. ab76252; 1:1,000; Abcam), anti-YAP (cat. no. ab52771; 1:1,000;
Abcam), anti-LATS1 (cat. no. 3477; 1:1,000; Cell Signaling
Technology, Inc.) and anti-β-actin (cat. no. ab8226; 1:2,000;
Abcam). Following incubation with the primary antibodies, the
membranes were incubated with secondary antibodies (anti-rabbit
IgG; cat. no. ab6721; 1:2,000; Abcam) at room temperature for 2 h.
The protein bands were visualized using an enhanced
chemiluminescence reagent (cat. no. WBKLS0100; MilliporeSigma) and
the protein ratios were calculated following densitometric analysis
using ImageJ v1.47 software (National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the SW480 cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.).
Total RNA was reverse transcribed into cDNA using reverse
transcriptase and random hexamers from a RevertAid First Strand
cDNA Synthesis kit (Thermo Fisher Scientific, Inc.). The following
temperature protocol was used: Priming at 25°C for 5 min, RT at
42°C for 60 min, then inactivation at 70°C for 5 min. qPCR was
subsequently performed by mixing cDNA, gene-specific primers and IQ
SYRB Green Supermix (Agilent Technologies, Inc.) and detected using
a Mx3005P Real-Time PCR system (Agilent Technologies, Inc.)
according to the manufacturer's protocol. The following primers
were used for the qPCR: CTGF forward, 5′-AAAAGTGCATCCGTACTCCCA-3′
and reverse, 5′-CCGTCGGTACATACTCCACAG-3′; CCN1 forward,
5′-AGCCTCGCATCCTATACAACC-3′ and reverse,
5′-TTCTTTCACAAGGCGGCACTC-3′; and GAPDH forward,
5′-GAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. The following thermocycling
conditions were used for qPCR: Initial denaturation at 95°C for 2
min, followed by 40 cycles of denaturation at 94°C for 15 sec, and
annealing/extension at 60°C for 1 min. Expression levels were
quantified using the 2−∆∆Cq method and fold changes were
obtained by normalization to GAPDH expression (27).
Immunofluorescence assay
The SW480 cells treated with 0.4 µM TCG or control
(or 5 µM CQ) were fixed with 4% paraformaldehyde at room
temperature for 30 min, washed with PBS, then incubated with 0.1%
Triton X-100 for permeabilization. Non-specific binding was
performed by blocking with 5% BSA (Sigma-Aldrich; Merck KGaA) for
30 min at 37°C and the cells were then incubated with anti-LC3B
(1:200; cat. no. L7543; Sigma-Aldrich; Merck KGaA), anti-lysosomal
associated membrane protein 2 (LAMP2; 1:200; cat. no. sc-20004;
Santa Cruz Biotechnology, Inc.) and anti-YAP (1:200; cat. no.
ab52771; Abcam) primary antibodies overnight at 4°C. Following
incubation with the primary antibody, the cells were incubated with
an Alexa Fluor 488-conjugated goat anti-rabbit IgG (1:1,000; cat.
no. ab150077; Abcam) or an Alexa Fluor 594-conjugated donkey
anti-mouse IgG (1:1,000; cat. no. ab150108; Abcam) secondary
antibody at room temperature for 1 h. Nuclei were stained with 10
ug/ml DAPI for 5 min at 37°C. The stained cells were
visualized using a confocal laser scanning microscope (FV1000;
Olympus Corporation).
Cell transfection
Small interfering (si) RNA targeting Atg5 and YAP,
and scramble siRNA were synthesized by Shanghai GenePharma Co.,
Ltd. The sequences of the siRNAs are as follows: Atg5 siRNA,
5′-GCAACUCUGGAUGGGAUUGTT-3′; YAP siRNA,
5′-CGAGAUGAGAGCACAGACAdTdT-3′; negative control (NC) of Atg5
(siScramble#1; 5′-GGAAAGAGCUGCAUAUUAATT-3′); and NC of YAP,
(siScramble#2; 5′-UAAGGCUAUGAAGAGAUAC-3′). The SW480 cells were
transfected with the siRNAs (50 nmol/l) at 37°C for 48 h using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. At 72 h
post-transfection, the cells were harvested and used for subsequent
experiments.
The pEGFP-LC3 plasmid was kindly gifted by Dr Lu
Zhang (Sichuan University, Chengdu, China). The SW480 cells were
transfected with the pEGFP-LC3 plasmid (2.5 µg) at 37°C using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. After
36 h at 37°C, the cells were treated with DMSO or TCG (0.4 µM) for
another 36 h at 37°C. Formation of EGFP-LC3 puncta was visualized
using fluorescence microscopy (FV1000; Olympus Corporation).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism v6.0 software (GraphPad Software, Inc.). Statistical
differences between groups were determined using a one-way ANOVA
followed by Tukey's post hoc test or an unpaired Student's t-test.
The data are presented as the mean ± SD. P<0.05 was considered
to indicate a statistically significant difference.
Results
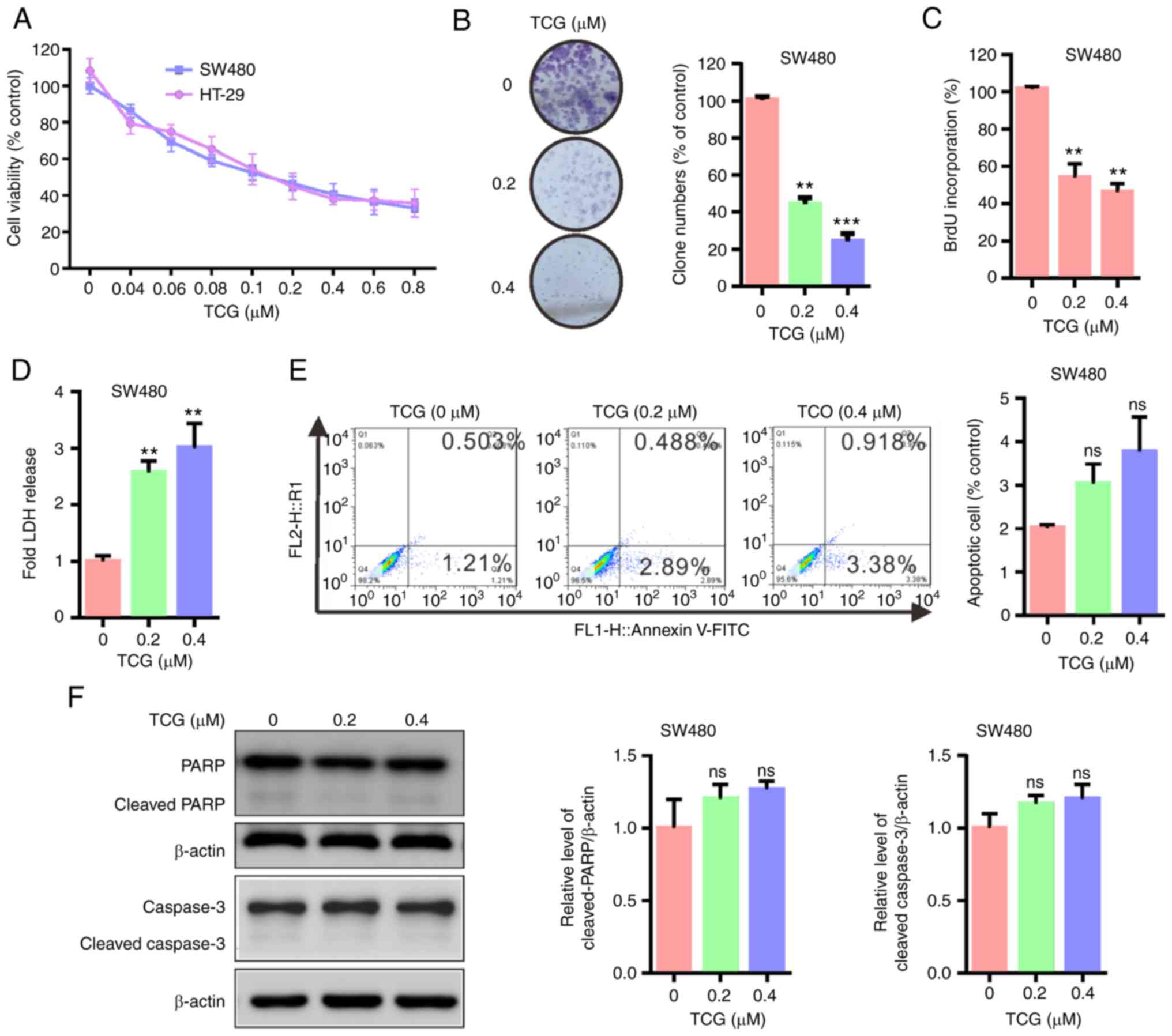
TCG inhibits SW480 cell cytotoxicity
and proliferation, independent of apoptosis
To determine whether TCG exhibits anticancer effects
against colorectal cancer cells, cell viability was determined
using a MTT assay. As shown in Fig.
1A, TCG treatment decreased the cytotoxicity of the colorectal
cancer cells in a dose-dependent manner compared with that in the
control group. Consistent with this finding, the results of the
colony formation and BrdU incorporation assays revealed that TCG
treatment inhibited SW480 cell proliferation compared with that in
the control group (Fig. 1B and C).
Furthermore, the LDH release assay showed that TCG treatment
increased SW480 cell death compared with that in the control group
(Fig. 1D). Taken together, these
results indicated that TCG may inhibit SW480 cell proliferation and
viability in vitro.
To further evaluate whether apoptosis was associated
with the anticancer effect of TCG, the apoptotic ratio was analyzed
using flow cytometry. As shown in Fig.
1E, TCG treatment for 24 h did not significantly increase the
levels of apoptosis in SW480 cells compared with that in the
control group. Consistent with this observation, the expression
levels of cleaved PARP and cleaved caspase-3 were not altered
between TCG-treated and control cells (Fig. 1F). These findings indicated that TCG
may not induce apoptosis in the SW480 cell line. Taken together,
these data suggested that TCG may inhibit SW480 cell viability and
proliferation, independent of apoptosis.
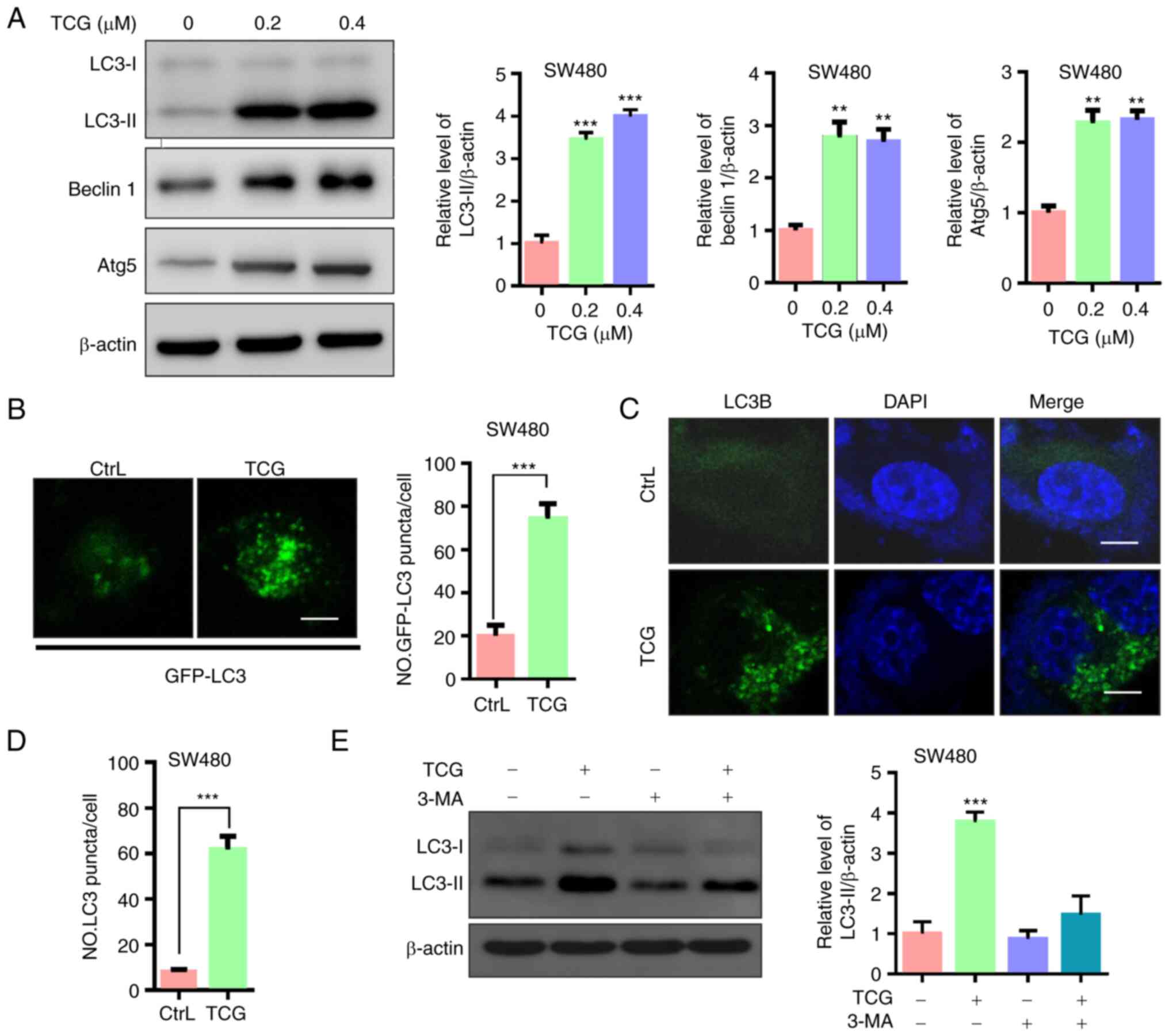
TCG induces autophagy in the SW480
cell line
Accumulating evidence has indicated that autophagy
is involved in drug-mediated anticancer therapy (10,28,29);
therefore, the present study investigated whether TCG induced
autophagy in the SW480 cell line. First, LC3-II accumulation, a
hallmark of autophagy, and the levels of Beclin 1 and Atg5, two
autophagy-related proteins, were analyzed. As shown in Fig. 2A, TCG treatment markedly increased
LC3-II expression in the SW480 cell lines. In addition, the
expression levels of Beclin1 and Atg5 were found to be increased in
the TCG-treated cells. To confirm this observation, a GFP-tagged
LC3B plasmid was used, and a marked increase in GFP-tagged LC3B
puncta was observed in TCG-treated cells compared with that in the
control cells (Fig. 2B). Next, the
distribution of endogenous LC3 puncta, another classical marker of
autophagy, was examined. The results revealed that TCG treatment
significantly increased the number of endogenous LC3 puncta in the
SW480 cell lines (Fig. 2C and D).
Furthermore, treatment with 3-MA (an autophagy inhibitor) markedly
decreased LC3-II expression in the TCG-treated cells (Fig. 2E). Taken together, these findings
indicated that TCG may induce autophagy in SW480 cells.
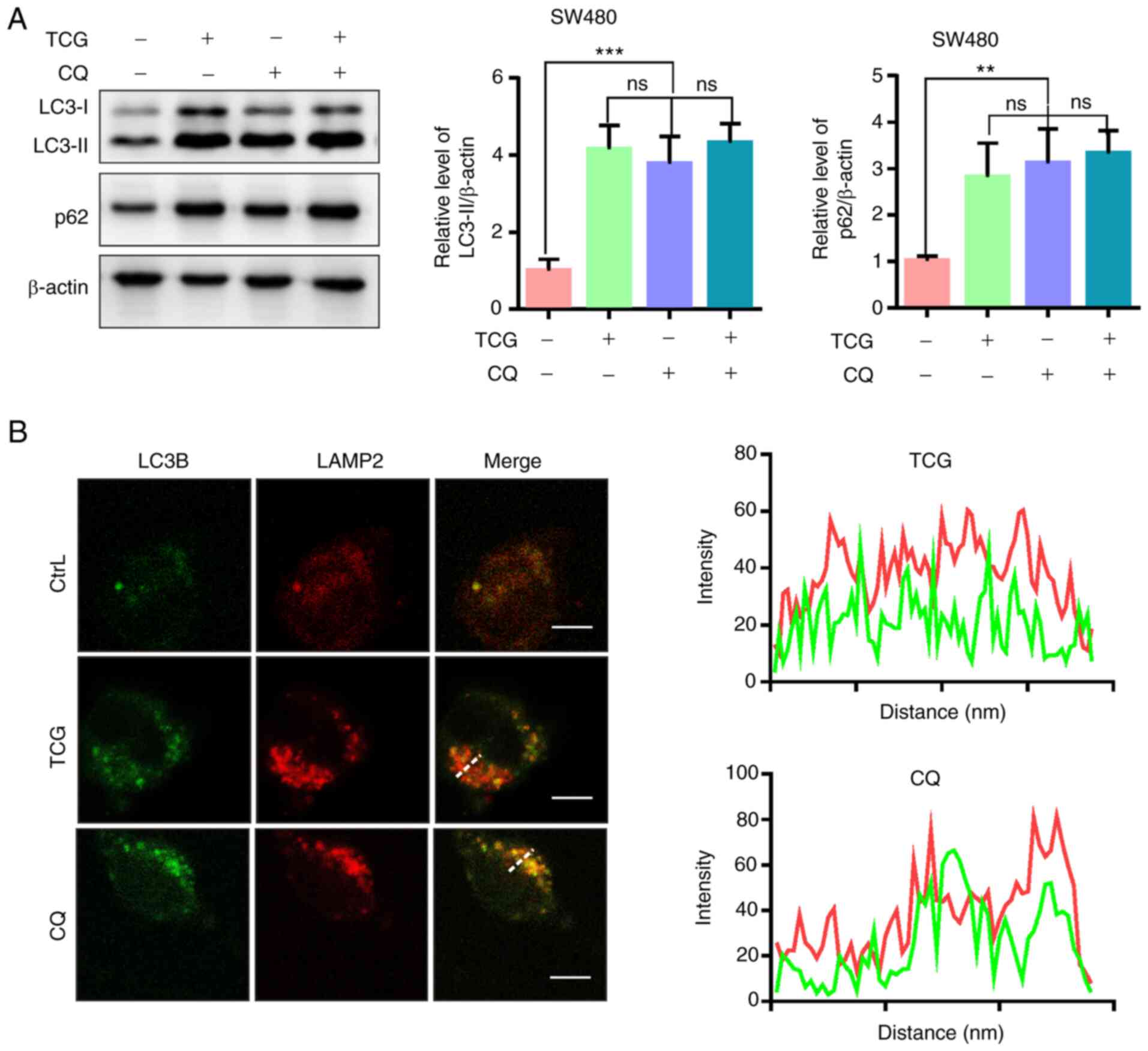
TCG induces lethal autophagy arrest in
the SW480 cell lines
In addition to autophagy initiation, LC3-II
accumulation may result from impaired autophagic flux (30). Therefore, the present study
investigated whether TCG induced complete autophagic flux in the
SW480 cell line. The protein expression levels of LC3-II and P62
(an autophagy-specific substrate) were investigated by adding CQ (a
lysosomal inhibitor) to the TCG-treated cells to CQ. The results
revealed that both LC3-II and P62 were significantly increased in
TCG-treated cells, but expression was not further increased upon
combined treatment with CQ (Fig. 3A).
Furthermore, the colocalization of LC3B with LAMP2 (a lysosomal
marker) was not observed in the TCG-treated cells (Fig. 3B), suggesting that TCG hinders the
fusion of autophagosomes with lysosomes. These results indicated
that TCG may induce autophagosome accumulation by blocking
autophagosome-lysosome fusion.
To determine whether autophagy was involved in the
TCG-mediated inhibition of cell viability and proliferation, the
SW480 cell line was treated with TCG in combination with 3-MA or
CQ. As shown in Fig. 4A and B, 3-MA
treatment markedly restored the TCG-mediated inhibition of cell
viability, whereas CQ treatment did not affect cell viability. The
results of the BrdU incorporation and colony formation assays
showed that 3-MA treatment significantly restored the TCG-mediated
suppression of cell proliferation (Fig.
4C and D). In addition, 3-MA treatment markedly decreased
TCG-induced cytotoxicity, as evidenced by the LDH release assay
(Fig. 4E). Atg5 knockdown
significantly decreased Atg5 protein expression levels in the SW480
cell line compared with that in the control group (Fig. 4F and G). Knockdown of Atg5 also
prevented the TCG-mediated suppression of cell viability, as
evidenced by MTT and LDH release assays (Fig. 4H and I). Taken together, these results
suggested that TCG may inhibit SW480 cell viability and
proliferation by promoting lethal autophagosome accumulation.
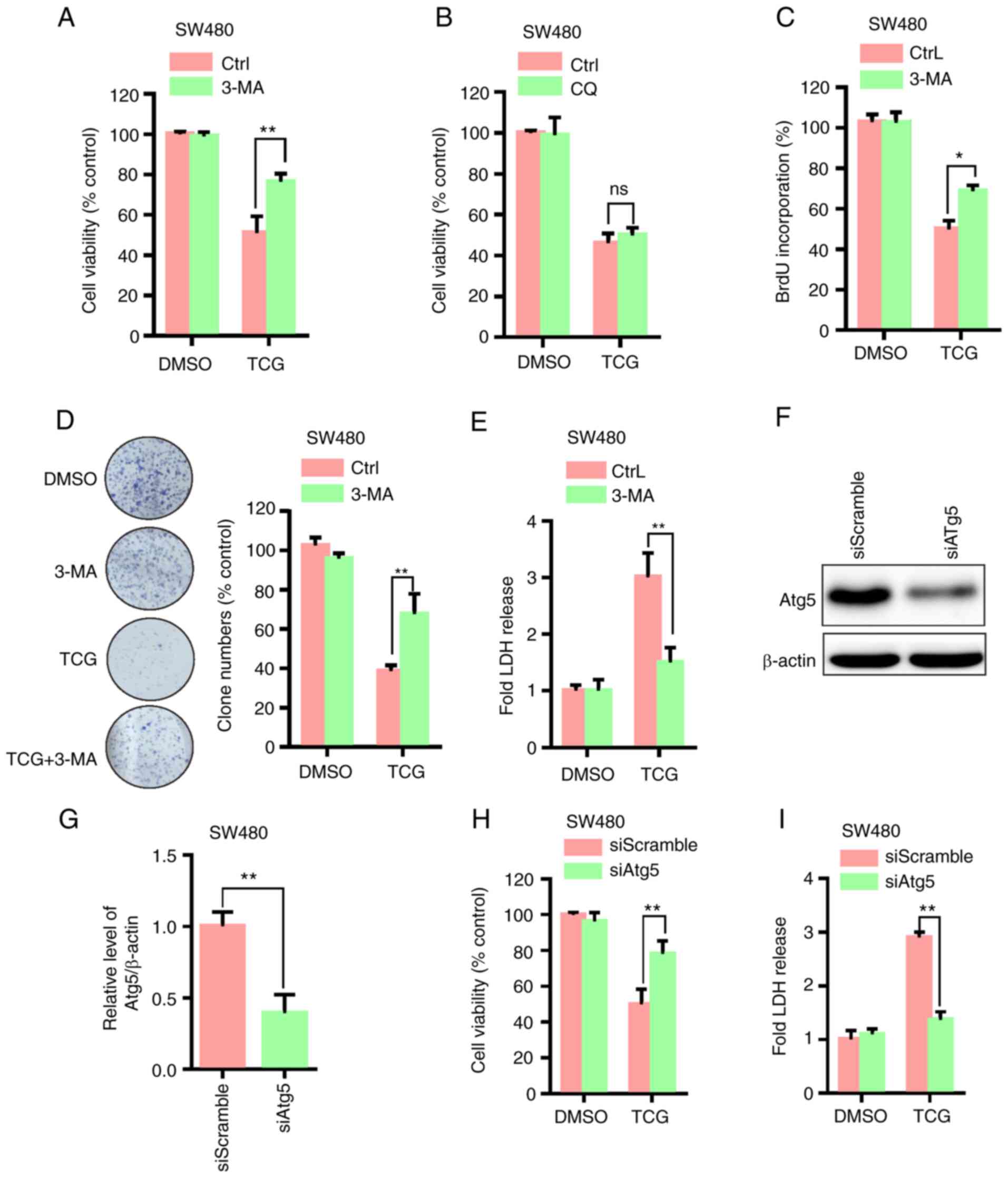 | Figure 4.TCG inhibits SW480 cell viability and
proliferation by promoting autophagosome accumulation. MTT assay
was used to determine the viability of cells treated with DMSO or
TCG for 24 h in the presence or absence of (A) 3-MA or (B) CQ. (C)
BrdU incorporation assay was used to determine the proliferation of
cells treated as described in part (A). (D) Colony formation assay
was used to determine the proliferation of cells treated with DMSO
or TCG for 7 days in the presence or absence of 3-MA. (E) LDH
release assay was performed using SW480 cells treated as described
in part (A). (F and G) Effect of Atg5 knockdown on Atg5 protein
expression. (H) MTT assay was used to determine the viability of
cells transfected with Atg5 siRNA or siScramble followed by
treatment with or without TCG for 24 h. (I) LDH release assay was
performed using SW480 cells transfected as described in part. (H)
The experiments were repeated in triplicate. *P<0.05,
**P<0.01. TCG, toxicarioside G; CQ, chloroquine; 3-MA,
3-methyladenine; BrdU, bromodeoxyuridine; LDH, lactate
dehydrogenase; si, small interfering; Atg5, autophagy related 5;
DMSO, dimethyl sulfoxide; Ctrl, control; ns, non-significant. |
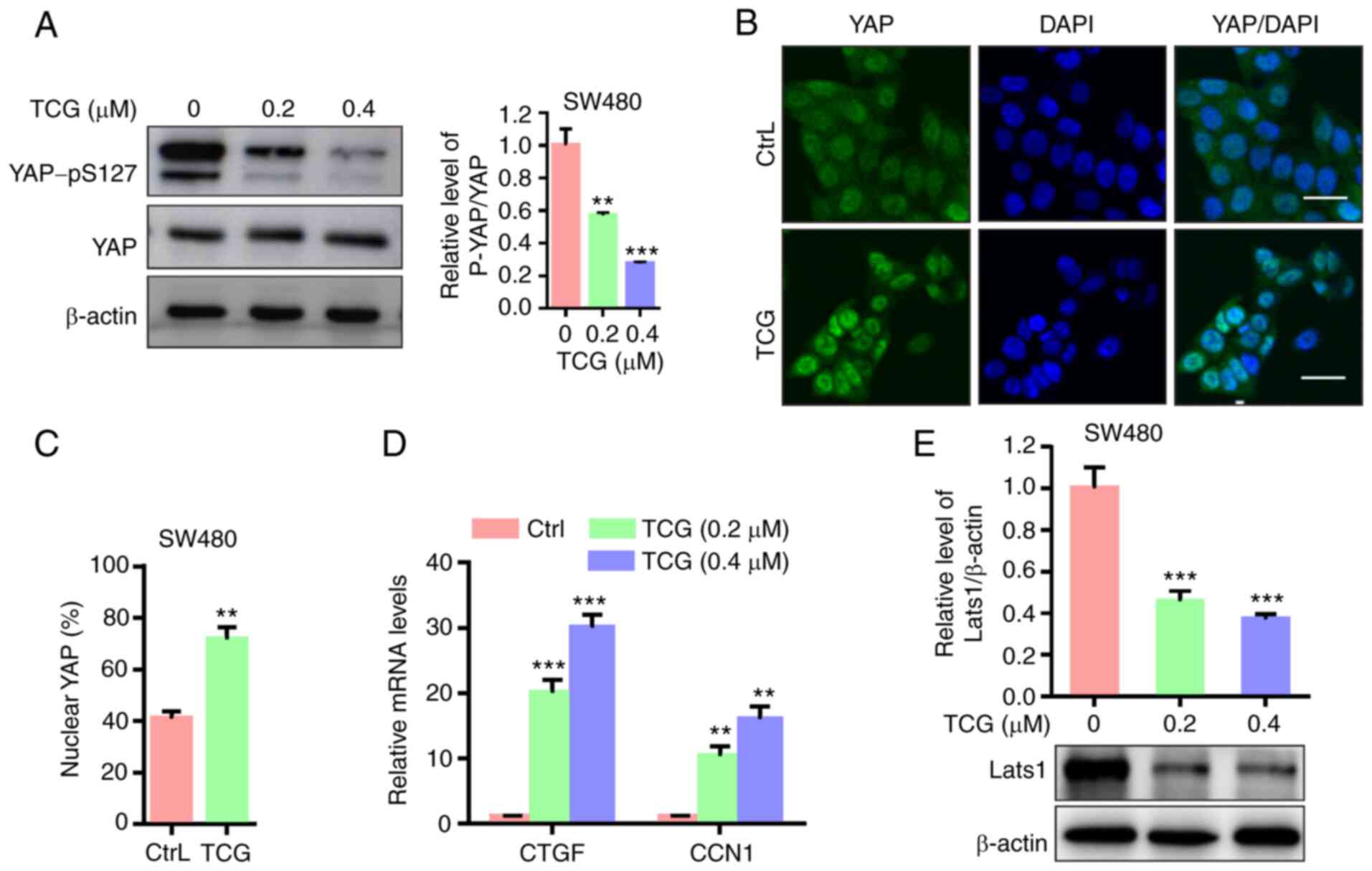
YAP is activated in TCG-treated SW480
cells
As the Hippo signaling pathway has been found to
play a key role in the development of numerous types of human
cancer (31), the present study aimed
to determine whether the Hippo signaling pathway was associated
with the TCG-mediated inhibition of SW480 cell viability and
proliferation. The results demonstrated that TCG treatment promoted
YAP dephosphorylation at serine 127 in the SW480 cell line
(Fig. 5A). In addition, YAP nuclear
localization was found to be elevated in the TCG-treated cells
(Fig. 5B and C). The mRNA expression
levels of downstream target genes of YAP were also investigated. As
shown in Fig. 5D, TCG treatment
markedly upregulated the mRNA expression levels of CTGF and CCN1.
Furthermore, TCG treatment significantly downregulated LATS1
protein expression level in the SW480 cell line (Fig. 5E). Collectively, these results
indicated that TCG may enhance YAP dephosphorylation and nuclear
localization, and downstream target gene expression, suggesting
that YAP may be activated in the TCG-treated SW480 cell line.
Inhibition of YAP activity enhances
the TCG-induced antiproliferative effect in the SW480 cell
line
To determine the role of YAP in the TCG-mediated
inhibition of cell viability and proliferation, cell viability and
proliferation were measured following treatment with the YAP
inhibitor, VP. The results of the MTT assay showed that VP
treatment significantly decreased cell viability compared with that
in cells treated with TCG alone (Fig.
6A). In addition, VP treatment further suppressed cell
proliferation in TCG-treated cells, as demonstrated by the BrdU
incorporation assay (Fig. 6B).
Furthermore, VP treatment markedly increased TCG-induced
cytotoxicity, which was evidenced using a LDH release assay
(Fig. 6C). YAP knockdown
significantly decreased YAP protein expression levels in the SW480
cell line compared with that in the control group (Fig. 6D and E). Knockdown of YAP further
enhanced the TCG-mediated inhibition of cell viability and
proliferation, which was evidenced by MTT, BrdU incorporation and
LDH release assays (Fig. 6F-H). Taken
together, these findings indicated that TCG-induced YAP activation
may play a protective role against the TCG-mediated inhibition of
cell viability and proliferation.
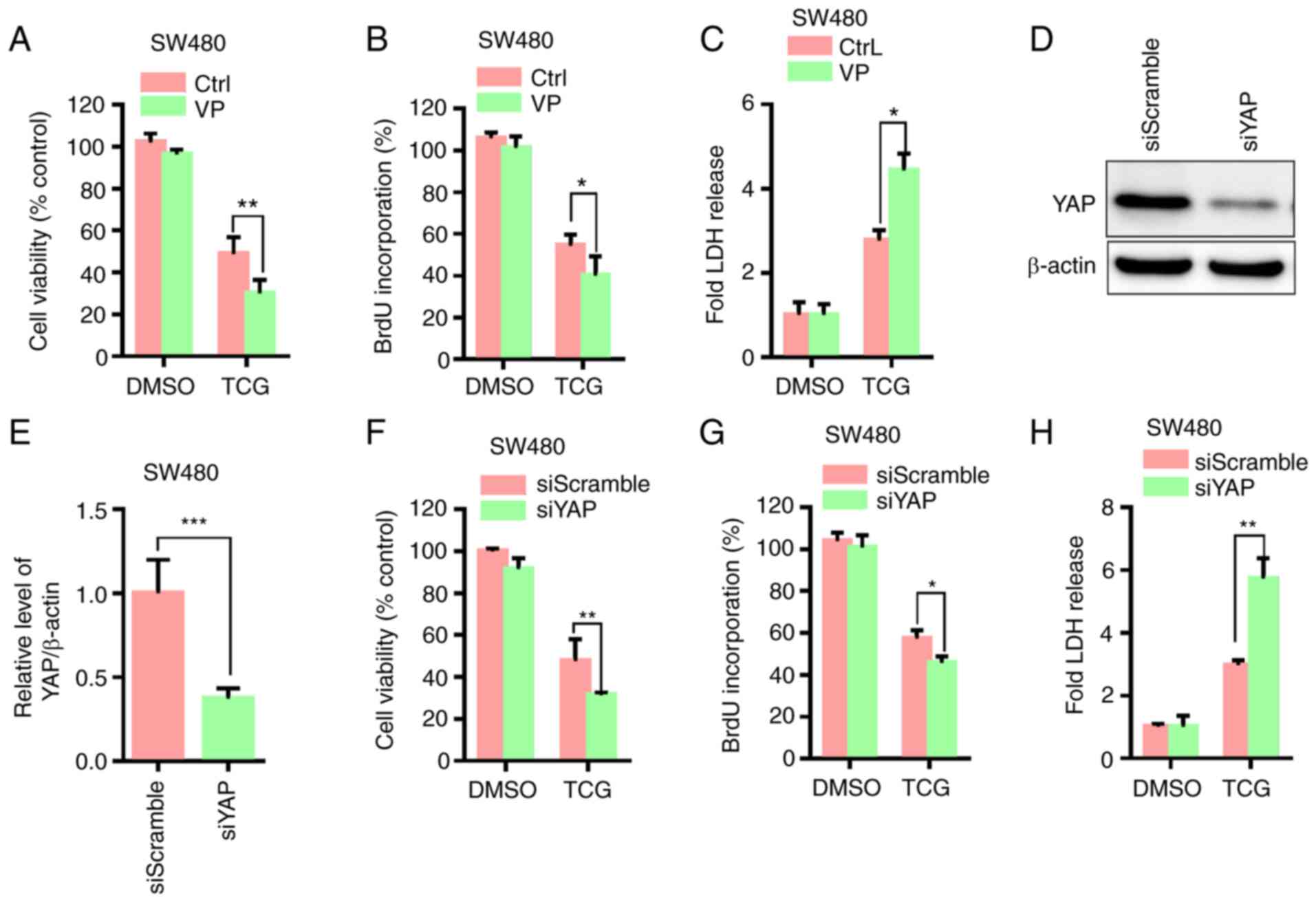 | Figure 6.Inhibition of YAP activity enhances
TCG-induced lethal autophagosome accumulation in the SW480 cell
line. (A) MTT, (B) BrdU incorporation and (C) LDH release assays
were used to determine the viability, proliferation and LDH levels
in cells treated with DMSO or TCG for 24 h in the presence or
absence of VP. (D) Effect of YAP knockdown on YAP protein
expression and the results were (E) statistically analyzed. (F) MTT
assay was used to determine the viability of cells transfected with
YAP siRNA or siScramble, followed by treatment with or without TCG
for 24 h. (G) BrdU incorporation assay was used to determine the
proliferation of cells transfected as described in part (F). (H)
LDH release assay was performed using SW480 cells transfected as
described in part. (F) The experiments were repeated in triplicate.
*P<0.05, **P<0.01, ***P<0.001. YAP, Yes1 associated
transcriptional regulator; TCG, toxicarioside G; VP, verteporfin;
BrdU, bromodeoxyuridine; LDH, lactate dehydrogenase; si, small
interfering; CtrL, control; DMSO, dimethyl sulfoxide. |
Discussion
In recent years, natural products have been of
significant interest due to the use of their compounds for
medicinal purposes. Numerous natural products derived from plants
have exhibited potent anticancer activities and have been
successfully used in cancer treatment, such as vincristine,
etoposide, irinotecan and paclitaxel (32–34).
Cardenolides, a class of natural products, including digitoxin,
oleandrin and ouabain, have received considerable attention due to
their reported anticancer activities (1,35,36). TCG is a cardenolide isolated from
Calotropis gigantea, which has been shown to exert potential
anticancer activities in several types of cancer cell line
(6). However, to the best of our
knowledge, the molecular mechanisms of TCG remain largely unknown.
The results of the present study demonstrated that TCG inhibited
the viability and proliferation of the SW480 cell line. In
addition, TCG induced autophagosome accumulation and the inhibition
of autophagy restored the TCG-mediated inhibition of cell viability
and proliferation, suggesting that TCG may induce lethal autophagy
arrest. The data further showed that TCG induced YAP activation in
the SW480 cell line, while the inhibition of YAP activity enhanced
the TCG-induced effects on cell viability and proliferation,
indicating that YAP may play a protective role in TCG-treated
cells.
Accumulating evidence has indicated that autophagy
is induced following treatment with numerous anticancer agents
(15,37); however, the reported role of autophagy
is paradoxical. Some anticancer agents have been shown to induce
cytoprotective autophagy and inhibition of autophagy rendered tumor
cells vulnerable to these drug treatments (28,38). On
the other hand, other anticancer agents were found to induce
cytotoxic autophagy and inhibition of autophagy promoted cancer
cell proliferation (17,39). The results of the present study
revealed that TCG induced autophagy in the SW480 cell line and
inhibition of autophagy favored cancer cell proliferation,
indicating that TCG may induce cytotoxic autophagy in the SW480
cell line. Autophagy arrest, which is involved in cytotoxic
autophagy, has been frequently observed in response to
chemotherapy. For example, elaiophylin, a natural product, was
reported to block the autophagic flux and promote the accumulation
of autophagosomes by attenuating lysosomal cathepsin activity,
resulting in lethal autophagy arrest (40). Regorafenib, an oral multi-kinase
inhibitor, induced lethal autophagy arrest in glioblastoma by
inhibiting autophagosome-lysosome fusion (39). Consistent with these observations, the
present study demonstrated that TCG induced lethal autophagy arrest
in the SW480 cell line by blocking autophagosome-lysosome
fusion.
The Hippo signaling pathway plays an important role
in regulating numerous aspects of tumor biology, and the
dysregulation of the Hippo signaling pathway components has been
associated with aberrant cell proliferation and tumor formation
(31,41). YAP, a key downstream effector of the
Hippo signaling pathway, regulates several context-specific
transcriptional programs, and was discovered to promote
proliferation and tumor growth (19,25).
Amplification of YAP, and high expression levels and nuclear
localization, have been frequently observed in numerous types of
cancer, including lung, colon and breast cancer, and the abnormal
activation of YAP has been associated with tumorigenesis and tumor
progression through its ability to act as a powerful tumor promoter
(18,24,25).
Conversely, in some circumstances, YAP has been discovered to
inhibit cell expansion, and also control organ size and growth,
which indicated that it may function as a possible tumor suppressor
(42). The results of the current
study revealed that TCG promoted YAP dephosphorylation, nuclear
localization and downstream target gene expression, suggesting that
YAP may be activated in the TCG-treated SW480 cell line. The
inhibition of YAP activity enhanced the TCG-mediated inhibition of
cell viability and proliferation, indicating that YAP activation
may play a protective role in TCG-treated cells.
Recently, the emerging link between the Hippo
signaling pathway and autophagy has attracted significant attention
due to their complex and reciprocal interactions, which have been
found to be involved in a wide range of human diseases, including
cancer (43,44). Accumulating data have suggested that
the Hippo signaling pathway may control autophagy through various
mechanisms and the inhibition of YAP function was found to reduce
basal autophagy levels (45,46). On the other hand, autophagy has been
suggested to regulate the Hippo signaling pathway via numerous
different mechanisms. Since YAP acts as an autophagic substrate,
the expression levels of the YAP protein and YAP target genes are
regulated by autophagy (47,48). The results of the present study
demonstrated that TCG induced autophagy, while activating YAP. The
findings further revealed that YAP activation restored the
inhibited cell viability and proliferation mediated by TCG-induced
lethal autophagy arrest in the SW480 cell line.
Admittedly, the present study on TCG-induced
autophagy arrest is preliminary and also focuses on the SW480 cell
line. Further investigation is required to verify the effect of TCG
on autophagy in an additional cell line, to clarify the association
between autophagy and the cell cycle, and to perform in vivo
experiments.
In conclusion, the findings of the present study
suggested that TCG may inhibit SW480 cell proliferation and
viability, which may be independent of apoptosis, but associated
with autophagy and the Hippo signaling pathway. TCG induced high
levels of autophagosome accumulation by blocking
autophagosome-lysosome fusion, thereby resulting in lethal
autophagy arrest. Furthermore, YAP was found to be activated in the
TCG-treated cells and YAP activation attenuated TCG-induced lethal
autophagy arrest in the SW480 cell line. These results suggested
that TCG may represent a potential anticancer agent for colorectal
cancer, and the combined use of YAP inhibitors and TCG may
represent a promising strategy for anticancer therapy.
Acknowledgements
Not applicable.
Funding
This study was funded by the National Natural
Science Foundation of China (grant no. 81860429).
Availability of data and materials
All datasets generated and/or analyzed during this
study are available from the corresponding author upon reasonable
request.
Authors' contributions
LZ and JW designed and performed the experiments,
analyzed the data, and drafted the manuscript. JLiu, JLia and YW
conducted the experiments and provided the materials for the
biological assays. QC and YH conceived the study, supervised all
the research and revised the manuscript. All authors contributed to
the article and approved the final version. LZ, JW and YH confirm
the authenticity of all the raw data.
Ethics approval and consent to
participate
The animal study was reviewed and approved by the
Ethics Committee of Hainan Medical University (approval no.
HY-2018-1005).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Newman RA, Yang P, Pawlus AD and Block KI:
Cardiac glycosides as novel cancer therapeutic agents. Mol Interv.
8:36–49. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cerella C, Dicato M and Diederich M:
Assembling the puzzle of anti-cancer mechanisms triggered by
cardiac glycosides. Mitochondrion. 13:225–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McConkey DJ, Lin Y, Nutt LK, Ozel HZ and
Newman RA: Cardiac glycosides stimulate Ca2+ increases
and apoptosis in androgen-independent, metastatic human prostate
adenocarcinoma cells. Cancer Res. 60:3807–3812. 2000.PubMed/NCBI
|
|
4
|
Hou Y, Shang C, Meng T and Lou W:
Anticancer potential of cardiac glycosides and steroid-azole
hybrids. Steroids. 171:1088522021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Skubnik J, Pavlickova V and Rimpelova S:
Cardiac glycosides as immune system modulators. Biomolecules.
11:6592021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dai HF, Gan YJ, Que DM, Wu J, Wen ZC and
Mei WL: Two new cytotoxic cardenolides from the latex of antiaris
toxicaria. J Asian Nat Prod Res. 11:832–837. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dai HF, Gan YJ, Que DM, Wu J, Wen ZC and
Mei WL: A new cytotoxic 19-nor-cardenolide from the latex of
antiaris toxicaria. Molecules. 14:3694–3699. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang YH, Sun Y, Huang FY, Li YN, Wang CC,
Mei WL, Dai HF, Tan GH and Huang C: Toxicarioside O induces
protective autophagy in a sirtuin-1-dependent manner in colorectal
cancer cells. Oncotarget. 8:52783–52791. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Janku F, McConkey DJ, Hong DS and Kurzrock
R: Autophagy as a target for anticancer therapy. Nat Rev Clin
Oncol. 8:528–539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xia H, Green DR and Zou W: Autophagy in
tumour immunity and therapy. Nat Rev Cancer. 21:281–297. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kroemer G, Marino G and Levine B:
Autophagy and the integrated stress response. Mol Cell. 40:280–293.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rubinsztein DC, Codogno P and Levine B:
Autophagy modulation as a potential therapeutic target for diverse
diseases. Nat Rev Drug Discov. 11:709–730. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maes H, Rubio N, Garg AD and Agostinis P:
Autophagy: Shaping the tumor microenvironment and therapeutic
response. Trends Mol Med. 19:428–446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marino G, Niso-Santano M, Baehrecke EH and
Kroemer G: Self-consumption: The interplay of autophagy and
apoptosis. Nat Rev Mol Cell Biol. 15:81–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi YH, Ding ZB, Zhou J, Hui B, Shi GM, Ke
AW, Wang XY, Dai Z, Peng YF, Gu CY, et al: Targeting autophagy
enhances sorafenib lethality for hepatocellular carcinoma via ER
stress-related apoptosis. Autophagy. 7:1159–1172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Elgendy M, Sheridan C, Brumatti G and
Martin SJ: Oncogenic Ras-induced expression of Noxa and Beclin-1
promotes autophagic cell death and limits clonogenic survival. Mol
Cell. 42:23–35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dou Q, Chen HN, Wang K, Yuan K, Lei Y, Li
K, Lan J, Chen Y, Huang Z, Xie N, et al: Ivermectin induces
cytostatic autophagy by blocking the PAK1/akt axis in breast
cancer. Cancer Res. 76:4457–4469. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu FX, Zhao B and Guan KL: Hippo pathway
in organ size control, tissue homeostasis, and cancer. Cell.
163:811–828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zanconato F, Cordenonsi M and Piccolo S:
YAP/TAZ at the roots of cancer. Cancer Cell. 29:783–803. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Crunkhorn S: New route to targeting the
Hippo pathway. Nat Rev Drug Discov. 20:3442021. View Article : Google Scholar
|
|
21
|
Meng Z, Moroishi T and Guan KL: Mechanisms
of Hippo pathway regulation. Genes Dev. 30:1–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hao Y, Chun A, Cheung K, Rashidi B and
Yang X: Tumor suppressor LATS1 is a negative regulator of oncogene
YAP. J Biol Chem. 283:5496–5509. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao B, Ye X, Yu J, Li L, Li W, Li S, Yu
J, Lin JD, Wang CY, Chinnaiyan AM, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cottini F, Hideshima T, Xu C, Sattler M,
Dori M, Agnelli L, ten Hacken E, Bertilaccio MT, Antonini E, Neri
A, et al: Rescue of Hippo coactivator YAP1 triggers DNA
damage-induced apoptosis in hematological cancers. Nat Med.
20:599–606. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Avruch J, Zhou D and Bardeesy N: YAP
oncogene overexpression supercharges colon cancer proliferation.
Cell Cycle. 11:1090–1096. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao B, Li L, Lei Q and Guan KL: The
Hippo-YAP pathway in organ size control and tumorigenesis: An
updated version. Genes Dev. 24:862–874. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang PM, Liu YL, Lin YC, Shun CT, Wu MS
and Chen CC: Inhibition of autophagy enhances anticancer effects of
atorvastatin in digestive malignancies. Cancer Res. 70:7699–7709.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Limpert AS, Lambert LJ, Bakas NA, Bata N,
Brun SN, Shaw RJ and Cosford NDP: Autophagy in cancer: Regulation
by small molecules. Trends Pharmacol Sci. 39:1021–1032. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klionsky DJ, Abdalla FC, Abeliovich H,
Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M,
Agostinis P, Aguirre-Ghiso JA, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy. Autophagy.
8:445–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Harvey KF, Zhang X and Thomas DM: The
Hippo pathway and human cancer. Nat Rev Cancer. 13:246–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mishra BB and Tiwari VK: Natural products:
An evolving role in future drug discovery. Eur J Med Chem.
46:4769–4807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Prassas I and Diamandis EP: Novel
therapeutic applications of cardiac glycosides. Nat Rev Drug
Discov. 7:926–935. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Maoyuan YW, Wenli M, Yuanyuan D, Shenglan
L, Zhunian W and Haofu D: Cytotoxic cardenolides from the roots of
Calotropis gigantea. Modern Pharm Res. 1:4–9. 2008.
|
|
37
|
Wang K, Liu R, Li J, Mao J, Lei Y, Wu J,
Zeng J, Zhang T, Wu H, Chen L, et al: Quercetin induces protective
autophagy in gastric cancer cells: Involvement of Akt-mTOR- and
hypoxia-induced factor 1α-mediated signaling. Autophagy. 7:966–978.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu R, Li J, Zhang T, Zou L, Chen Y, Wang
K, Lei Y, Yuan K, Li Y, Lan J, et al: Itraconazole suppresses the
growth of glioblastoma through induction of autophagy: Involvement
of abnormal cholesterol trafficking. Autophagy. 10:1241–1255. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang J, Zhang L, Chen H, Lei Y, Zhang T,
Wang Y, Jin P, Lan J, Zhou L, Huang Z, et al: Regorafenib induces
lethal autophagy arrest by stabilizing PSAT1 in glioblastoma.
Autophagy. 16:106–122. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao X, Fang Y, Yang Y, Qin Y, Wu P, Wang
T, Lai H, Meng L, Wang D, Zheng Z, et al: Elaiophylin, a novel
autophagy inhibitor, exerts antitumor activity as a single agent in
ovarian cancer cells. Autophagy. 11:1849–1863. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu FX, Luo J, Mo JS, Liu G, Kim YC, Meng
Z, Zhao L, Peyman G, Ouyang H, Jiang W, et al: Mutant Gq/11 promote
uveal melanoma tumorigenesis by activating YAP. Cancer Cell.
25:822–830. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Barry ER, Morikawa T, Butler BL, Shrestha
K, de la Rosa R, Yan KS, Fuchs CS, Magness ST, Smits R, Ogino S, et
al: Restriction of intestinal stem cell expansion and the
regenerative response by YAP. Nature. 493:106–110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang D, He J, Huang B, Liu S, Zhu H and Xu
T: Emerging role of the Hippo pathway in autophagy. Cell Death Dis.
11:8802020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tang F and Christofori G: The cross-talk
between the Hippo signaling pathway and autophagy:Implications on
physiology and cancer. Cell Cycle. 19:2563–2572. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Totaro A, Zhuang Q, Panciera T, Battilana
G, Azzolin L, Brumana G, Gandin A, Brusatin G, Cordenonsi M and
Piccolo S: Cell phenotypic plasticity requires autophagic flux
driven by YAP/TAZ mechanotransduction. Proc Natl Acad Sci USA.
116:17848–17857. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tang F and Christofori G: LATS1-Beclin1
mediates a non-canonical connection between the Hippo pathway and
autophagy. Mol Cell Oncol. 7:17573782020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee YA, Noon LA, Akat KM, Ybanez MD, Lee
TF, Berres ML, Fujiwara N, Goossens N, Chou HI, Parvin-Nejad FP, et
al: Autophagy is a gatekeeper of hepatic differentiation and
carcinogenesis by controlling the degradation of Yap. Nat Commun.
9:49622018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhou Y, Wang Y, Zhou W, Chen T, Wu Q,
Chutturghoon VK, Lin B, Geng L, Yang Z, Zhou L and Zheng S: YAP
promotes multi-drug resistance and inhibits autophagy-related cell
death in hepatocellular carcinoma via the RAC1-ROS-mTOR pathway.
Cancer Cell Int. 19:1792019. View Article : Google Scholar : PubMed/NCBI
|