Introduction
Preeclampsia is a heterogeneous pregnancy disorder
that may develop in late pregnancy, and is characterized by
hypertension and proteinuria which may occur after 20 weeks of
gestation. It affects 2–8% of pregnancies worldwide (1,2).
Preeclampsia has preclinical (symptomless) and clinical
(symptomatic) stages. Until recently, only the symptomatic final
stage preeclampsia could be detected by clinical screening.
Although extensive research on the pathophysiology of the disease
has been conducted, the etiology of preeclampsia remains poorly
understood. A two-stage disorder theory is accepted for
preeclampsia: Stage 1, poor placentation at the early stage of
pregnancy; and stage 2, placental oxidative stress at the late
stage of pregnancy. Stage 1 is preclinical and characterized by
faulty trophoblast invasion and spiral artery (SA) remodeling,
resulting in failure to remodel the SAs supplying the
uteroplacental circulation, and placental hypoxia (3).
Trophoblast differentiation, invasion and SA
remodeling are regulated by several trophoblast- and/or
decidua-derived factors, including cytokines, growth factors,
hormones and oxygen, in the first trimester (4–6). Recent
studies have reported that impaired invasion of trophoblasts, and
the subsequent incomplete SA remodeling, may result in preeclampsia
(6,7). Consequently, elucidating the
trophoblast invasion mechanisms is crucial in understanding the
pathophysiological mechanisms of preeclampsia. The decidual
leukocyte population, in particular uterine natural killer (uNK)
cells, is proposed to have a major role in the regulation of
trophoblast invasion (8,9). Reduced numbers of uNK cells are
observed in patients with preeclampsia and intrauterine growth
restriction (10), which are
conditions associated with poor SA remodeling and reduced
trophoblast invasion in the decidua (11).
Unique long 16 binding proteins (ULBPs) which are
also termed as retinoic acid early transcripts (RAET), encoded by
RAET1 genes, are a family of ligands for natural-killer group 2D
(NKG2D) receptors in humans that are frequently expressed by tumor
cells and mediate biological functions of NK cells (12). The majority of studies of NKG2D
ligands have traditionally demonstrated their expression only on
infected or transformed cells (13,14);
whereas other studies have demonstrated that several normal cells
and tissues also express NKG2D ligands (15,16).
Constitutive intracellular presence of ULBP1-4 has been recently
described in normal human bronchial epithelium (17). A study by Hedlund et al
(18) demonstrated that ULBP1-5 are
constitutively transcribed and expressed as proteins in human early
placenta (8–16 weeks), and have localized expression on the
membrane of exosomes of the multivesicular late endosomes in the
syncytiotrophoblast (STB). A previous study using DNA microarray
analysis and validation by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR), demonstrated that ULBP1 was
upregulated in preeclamptic placentas (19).
Considering that inadequate invasion of trophoblasts
in the first trimester may lead to preeclampsia and the role of uNK
cells in the regulation of trophoblast invasion, it was
hypothesized that ULBP1 may inhibit the invasion of extravillous
trophoblasts (EVTs) by altering cytokines secreted by uNK cells via
binding to NKG2D. Although the differential expression of ULBP1 in
preeclampsia in the first trimester is difficult to determine, the
differential expression of genes or proteins detected in full-term
placenta may provide an indication to investigate the mechanism.
The present study was performed to determine the expression levels
of ULBP1 in placentas collected following cesarean section from
women with preeclampsia and normal pregnant women. The functions of
ULBP1 in trophoblast invasion were also investigated.
Materials and methods
Ethics statement
Ethical approval was granted by the Ethics Committee
of The First Affiliated Hospital of China Medical University
(Shenyang, China) and methods were carried out in accordance with
the committee guidelines. Informed consent was obtained from all
participating patients.
Tissue collection
The present study included 30 pregnant women with
preeclampsia and 30 normal pregnant women. Human placental tissues
were collected at the time of cesarean section from the Department
of Obstetrics between September 2014 and August 2015, The First
Affiliated Hospital of China Medical University (Shenyang, China).
The clinical characteristics of the patients included in the
present study are summarized in Table
I. Preeclampsia was diagnosed according to the reported
criteria (20). Patients enrolled in
the preeclampsia group had no history of pre-existing or chronic
hypertension, although they exhibited ≥140 mmHg systolic or ≥90
mmHg diastolic pressure on two occasions at least 4 h apart after
20 weeks of gestation and ≥300 mg per 24-h urine collection after
20 weeks of gestation. Chorionic tissues were obtained from four
different parts of the placenta, from which the amniotic membrane
and maternal decidual tissues were removed. Tissues were frozen and
stored at −80°C until use. Decidual samples were obtained from
women undergoing elective surgical termination of pregnancy at
12–14 weeks of gestation (as determined by ultrasound measurement
of crown rump length or biparietal diameter). Following collection,
decidual tissue was immediately suspended in sterile saline,
transported to the laboratory and washed two to three times in
sterile phosphate-buffered saline (PBS) to remove excess blood.
 | Table I.Clinical characteristics of pregnant
women enrolled on the present study. |
Table I.
Clinical characteristics of pregnant
women enrolled on the present study.
| Clinical
characteristic | Normal pregnancy
(n=30) | Preeclampsia
(n=30) | P-value |
|---|
| Maternal age
(years) | 29.50±3.21 | 29.8±3.05 | 0.71 |
| Body mass index
(kg/m2) | 24.82±1.89 | 25.67±1.45 | 0.06 |
| Systolic blood
pressure (mmHg) | 119.67±9.24 | 155.05±10.53 |
<0.05a |
| Diastolic blood
pressure (mmHg) | 68.5±6.33 | 105.13±9.18 |
<0.05a |
| 24 h urine protein
(g) | 0.047±0.025 | 2.54±1.40 |
<0.05a |
| Gestational age at
delivery (weeks) | 38.84±0.54 | 37.26±1.91 |
<0.05a |
| Fetal birth weight
(g) | 3433.33±320.78 | 2580.00±600.87 |
<0.05a |
Immunocytochemistry
Formalin-fixed and paraffin-embedded tissue sections
of 4-µm thickness were prepared. Immunostaining was performed using
a streptavidin-peroxidase method. Sections were incubated with a
ULBP1 primary antibody (1:100; sc-33456; Santa Cruz Biotechnology
Inc., Dallas, TX, USA) at 4°C overnight, followed by a biotinylated
goat serum anti-rabbit immunoglobulin (Ig)G secondary antibody
(1:100; 0017; Ultrasensitive; Fuzhou Maixin Biotech Co., Fuzhou,
China) at room temperature for 10 min. Subsequent to washing with
PBS three times, the sections were incubated with horseradish
peroxidase-conjugated streptavidin-biotin (1:1:1; 0017;
Ultrasensitive; Fuzhou Maixin Biotech Co.) at room temperature for
10 min, developed using 3,3-diaminobenzidine tetrahydrochloride,
lightly counterstained with hematoxylin, dehydrated in alcohol and
mounted. Sections were observed under an Olympus BX51 microscope
(Olympus Corp., Tokyo, Japan). As a negative control,
isotype-specific IgG was used instead of primary antibody to
exclude nonspecific binding of the secondary antibody.
RT-qPCR
Total RNA was extracted from human placental tissues
using TRIzol reagent (Ambion; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and were treated with DNase I (Promega Corp.,
Madison, WI, USA). cDNA was generated from 1 µg total RNA using a
PrimeScript Reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China), according to the manufacturers' instructions. The PCR
reaction (20 µl final volume) contained 10 µl SYBR Premix Ex
Taq, 2 µl cDNA, 0.8 µl of each primer and 6.4 µl double
distilled H2O. qPCR analysis for ULBP1 and GAPDH was
performed as follows: 95°C for 30 sec; 45 cycles of 95°C for 5 sec;
and 60°C for 20 sec using a Roche Lightcycler 480 (Roche
Diagnostics GmbH, Mannheim, Germany). Gene-specific amplifications
were confirmed with melting curve analysis following RT-qPCR.
Relative gene expression levels were determined using the threshold
cycle method (2−ΔΔCq method) (21) with reference to the endogenous GAPDH
control. The following primers were used: ULBP1 forward,
5′-CAGCAGACGATGAGGACATT-3′ and reverse,
5′-GACAGAAAGTGGCAGAAGGTG-3′; and GAPDH forward,
5′-CATGAGAAGTATGACAACAGCCT-3′ and reverse,
5′-AGTCCTTCCACGATACCAAAGT-3′.
Western blotting
Human placental tissues were lysed in a
radioimmunoprecipitation assay lysis buffer (Nanjing KeyGen Biotech
Co., Ltd., Nanjing, China), and protein concentrations were
determined using an Enhanced BCA Protein Assay kit (Beyotime
Institute of Biotechnology, Beijing, China). A total of 20 µg of
proteins were separated by 10% SDS-PAGE and transferred onto a
polyvinylidene difluoride membrane (Merck Millipore, Darmstadt,
Germany). Membranes were blocked for 1 h at room temperature with
TBST (50 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20, pH 7.0)
containing 5% nonfat dry milk and incubated with primary antibodies
against ULBP1 (1:200; sc-33456; Santa Cruz Biotechnology) and
β-actin (1:8,000; 66009-1; Proteintech Group, Inc., Chicago, IL,
USA) overnight at 4°C. The membranes were washed with TBST three
times for 10 min, followed by incubation with
horseradish-peroxidase-labeled secondary antibodies (1:12,000;
SB-0071; Dingguo, Beijing, China) for 1 h at room temperature.
Protein expression was visualized using an enhanced
chemiluminescence system (Tanon5200; Tanon Science and Technology
Co., Ltd., Shanghai, China). Bands were analyzed using densitometry
with ImageJ software (version 1.46r; National Institutes of Health,
Bethesda, MA, USA).
uNK cell isolation
Total decidual cell isolates and purified
CD56+CD3− uNK cell isolates were prepared by
enzymatic disaggregation and immunomagnetic selection (MACS) as
previously described (22,23). The tissue was trimmed into 1-mm
pieces and enzymatically digested for 20 min, using vigorous
shaking, with type I DNase and IV collagenase in RPMI 1640 medium
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA).
Supernatants were collected and incubated overnight in a tissue
culture dish in a standard 37°C environment with 5% CO2.
Nonadherent cells were collected and loaded onto Ficoll-Paque
density gradient media (GE Healthcare Bio-Sciences, Pittsburgh, PA,
USA) to purify the lymphocyte population. Cell suspensions were
subjected to MACS (MidiMACS; Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany) with an NK Cell Isolation kit (Miltenyi Biotec
GmbH) to obtain uNK cell suspensions. uNK cells were plated on a
24-well plate at 5×104 cells/well in 600 µl RPMI-1640
medium supplemented with 1,000 U/ml penicillin, 1 mg/ml
streptomycin, 2 mM L-glutamine and 10% fetal bovine serum (Hyclone;
GE Healthcare Life Sciences), with or without 5 µg/ml recombinant
ULBP1-Fc chimera protein (SinoBiological Inc., Beijing, China), and
incubated for 72 h in a standard 37°C environment (5%
CO2). Cell viability was routinely tested by Trypan blue
exclusion. Viability, which was calculated as viability = (total
cells-dead cells)/total cells, was 80–90% after 72 h of cell
culture. Cell-free conditioned medium was removed and stored at
−20°C until required for invasion assay and ELISA.
Cell culture
EVT cell line HTR-8/SVneo cells were kindly provided
by Dr Charles H. Graham (Queen's University, Kingston, ON, Canada)
and cultured in RPMI-1640 medium supplemented with 10% FBS in a
standard 37°C environment (5% CO2) in an air
incubator.
Invasion assay
Cell invasion assays were performed to investigate
the effect of ULBP1 on the invasion of HTR-8/SVneo cells.
HTR-8/SVneo cells were placed in the upper chamber of an 8-µm
Transwell plate (Costar, New York, NY, USA) in 80 µl
growth-factor-reduced Matrigel (1:9; BD Biosciences, San Jose, CA,
USA). Inserts were pre-coated with 80 µl Matrigel matrix (1:9; BD
Biosciences). The different uNK cell conditioned media (cultured
with or without the ULBP1 protein, 33% v/v) was added to the lower
chambers. To investigate whether trophoblast invasion was mediated
by cytokines secreted by uNK cells, two representative neutralizing
antibodies for cytokines that inhibit or stimulate trophoblast cell
invasion were added to the 72-h cell supernatants cultured with
ULBP1. These antibodies included specific anti-interferon (IFN)-γ
(mouse anti-human IFN-γ, 10 µg/ml; D044-3) for stimulating
trophoblast cell invasion and a specific anti-interleukin (IL)-8
neutralizing antibody (goat anti-human IL-8, 0.5 µg/ml; MAB208;
both R&D Systems Europe, Ltd.) for inhibition of trophoblast
cell invasion. A total of 1×105 HTR-8/SVneo cells in 200
µl of serum-free medium were plated in the upper chamber, and a
total of 600 µl media was added in the lower chamber. Following 24
h, the membranes of the Transwell chamber were fixed in methanol
and stained with hematoxylin and eosin (OriGene Technologies,
Beijing, China). Finally, the number of cells was counted by
capturing images of the membrane with an Olympus CKX41 microscope
(Olympus Corp.). Each experiment was performed in duplicate in 10
cases. The invasion index was the average number of invaded cells
in test filters divided by the average number of invaded cells in
control filters for any given experiment.
Flow cytometry
Following 72 h of incubation of uNK cells with or
without recombinant ULBP1-Fc (5 µg/ml), surface antigen
fluorescence-activated cell sorting analysis was performed to
detect the modulation of NKG2D by recombinant ULBP1-Fc. Cells were
washed twice with ice-cold PBS. The washes were followed by
incubation of the cells with mouse anti-NKG2D antibody (320805;
BioLegend, Inc., San Diego, CA, USA) for 30 min on ice. Subsequent
to two washes with PBS, a FACScan flow cytometer (BD Pharmingen,
San Diego, CA, USA) was used for analysis. Background staining was
estimated after incubation with phycoerythrin-labeled isotype
antibodies (400113, 400107 and 400117; 5 µl per 106
cells in 100 µl staining volume, all BioLegend, Inc., San Diego,
CA, USA). ELISA kits (R&D Systems Europe, Ltd.) were used to
measure tumor necrosis factor (TNF)-α (DTA00C), IFN-γ (DIF50),
transforming growth factor (TGF)-β1 (DB100B), IL-6 (D6050) and IL-8
(D8000C). Cell-free supernatants were analyzed using ELISA
following 72 h incubation of uNK cells with or without recombinant
ULBP1-Fc (5 µg/ml). Each experiment was performed in triplicate in
10 cases.
Statistical analysis
Statistical analyses were performed using SPSS
version 17.0 (SPSS, Inc., Chicago, IL, USA). Results are presents
as the mean + standard error. Differences between two groups were
analyzed using Student's t-test. P<0.05 was considered to
indicate a statistically significant result.
Results
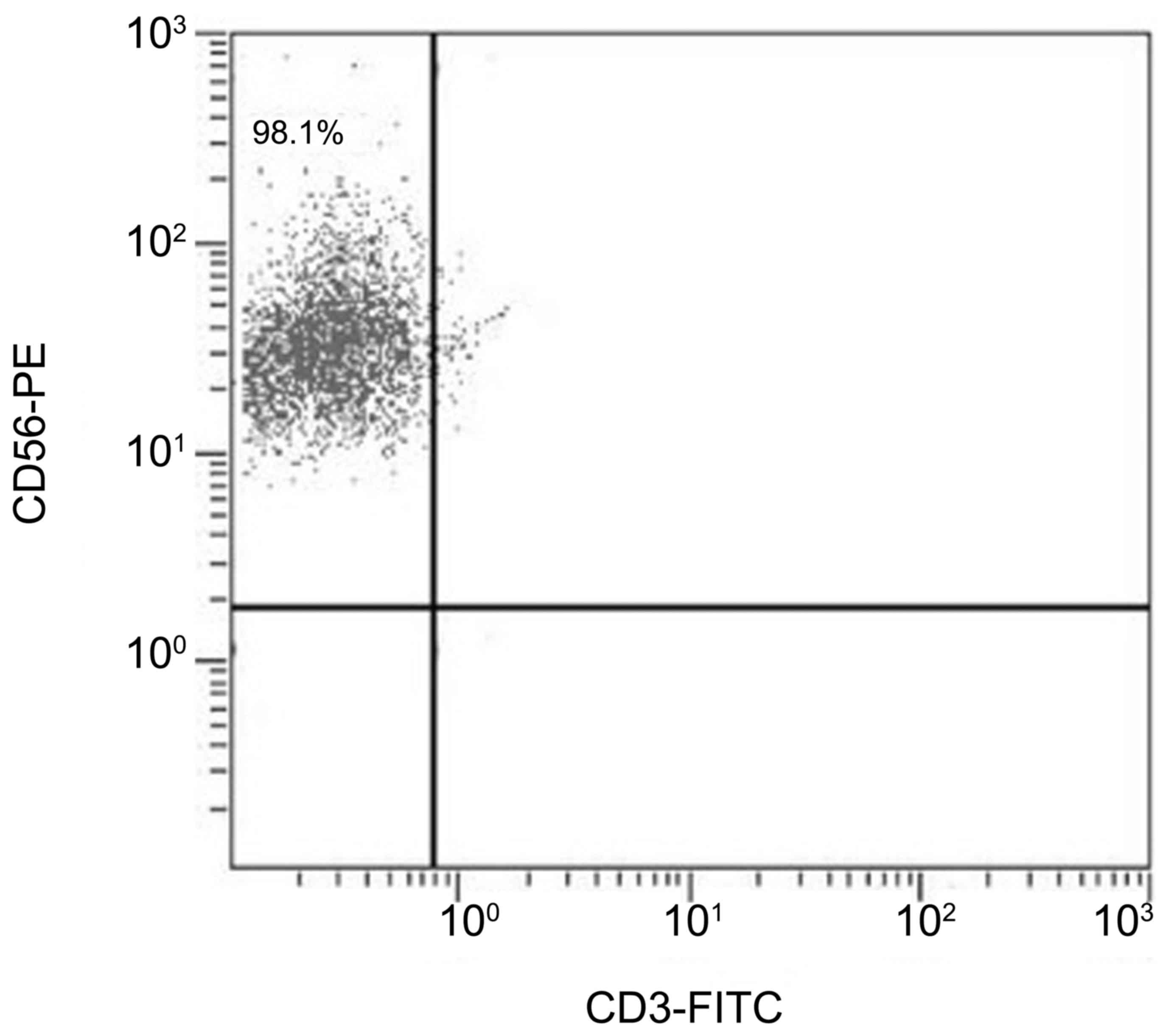
uNK cell isolation
CD56+CD3− cell-enriched
isolates were demonstrated to be consistently >95% pure by flow
cytometry (Fig. 1).
mRNA and protein expression levels of
ULBP1 are upregulated in preeclamptic placentas
RT-qPCR was used to investigate the mRNA expression
levels of ULBP1 in preeclamptic and normal placentas. As shown in
Fig. 2A, the mRNA expression levels
of ULBP1 in preeclampstic placentas were significantly upregulated
compared with that in normal placentas (P<0.05). In agreement
with the results of RT-qPCR, western blotting demonstrated that
ULBP1 protein expression levels were significantly higher in
preeclamptic placentas compared with that of normal placentas
(P<0.05; Fig. 2B).
Immunocytochemistry demonstrated that ULBP1 was predominantly
located in the cytoplasm of STB and that normal and preeclamptic
placentas expressed ULBP1; however, ULBP1 expression levels were
greater in preeclamptic placentas, as compared with normal
placentas (Fig. 2C).
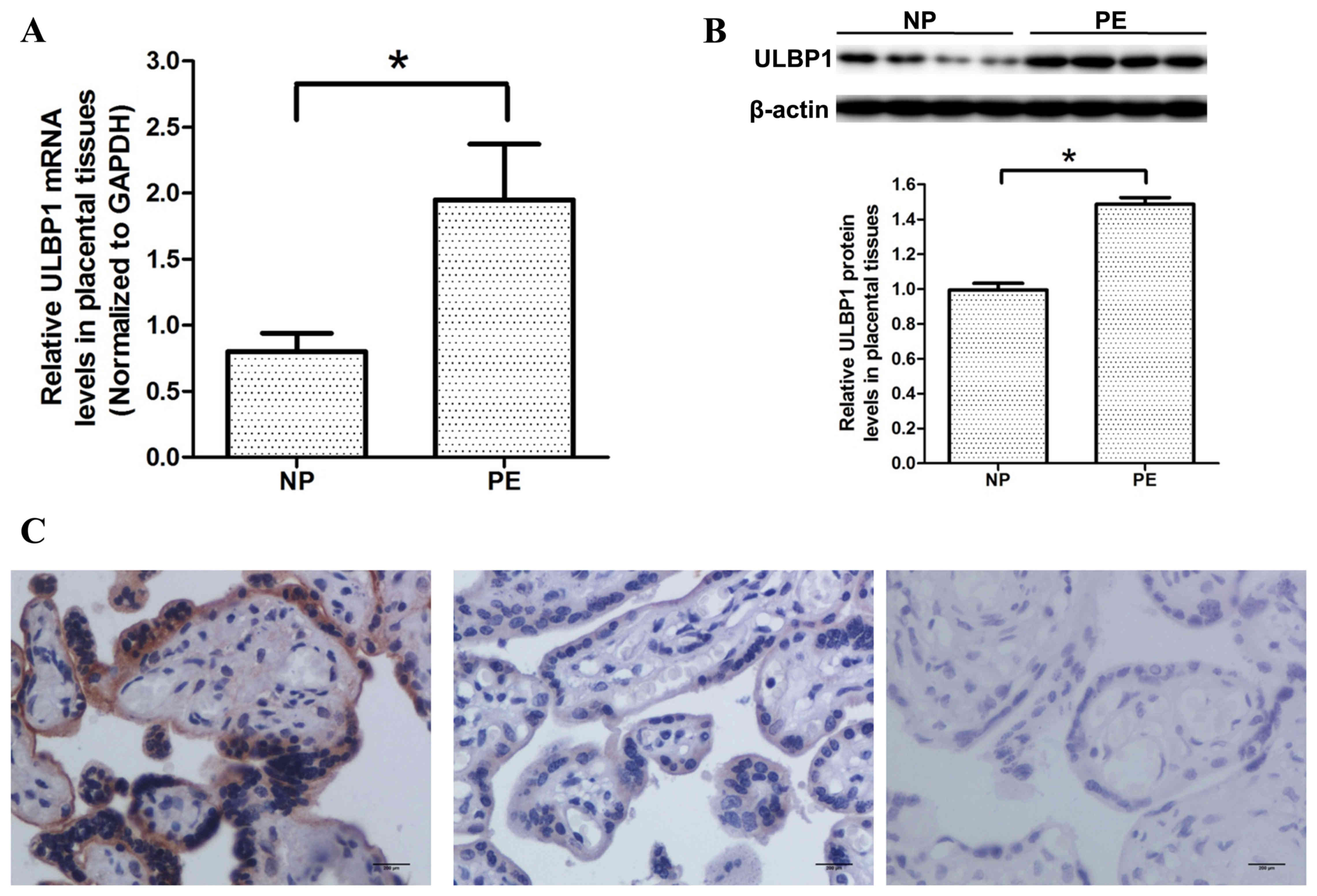 | Figure 2.Expression levels of ULBP1 in
placentas from pregnant women with PE and women with NP were
determined by RT-qPCR, western blotting, and immunohistochemistry
analysis. (A) RT-qPCR analysis of ULBP1 mRNA expression levels in
placentas from PE and NP women (n=30 for each group). (B) Western
blot analysis of ULBP1 protein expression in placentas from PE and
NP women. Upper panel, a typical result of western blotting; lower
panel, bar chart according to the statistical analysis based on the
result of three independently repeated experiments (n=30 for each
group). (C) Immunostaining of ULBP1 in placentas from PE and NP
women. Data are presented as the mean + standard error of the mean.
Left panel, PE; middle panel, NP; right panel, negative control;
scale bars, 200 µm. *P<0.05, PE vs. NP. ULBP1, unique long 16
binding protein 1; PE, preeclampsia; NP, normal pregnancy; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction. |
uNK cell culture supernatants with
ULBP1 inhibit the invasion of HTR-8/SVneo cells
Following incubation of uNK cells with or without
ULBP1 for 72 h, it was possible to determine the effect of ULBP1 on
the invasion of EVT. When EVT was cultured in the presence of uNK
cell culture supernatants with ULBP1, there was a significant
decrease in EVT invasion compared to the cell culture supernatants
without ULBP1 (P<0.05; Fig. 3A and
B). Following the addition of specific anti-IFN-γ or anti-IL-8
neutralizing antibody to uNK cell culture supernatants with ULBP1,
anti-IFN-γ neutralizing antibody significantly decreased the
inhibitory effect (P<0.05; Fig.
3C) and anti-IL-8 neutralizing antibody enhanced the inhibitory
effect of ULBP1 (P<0.05; Fig.
3D).
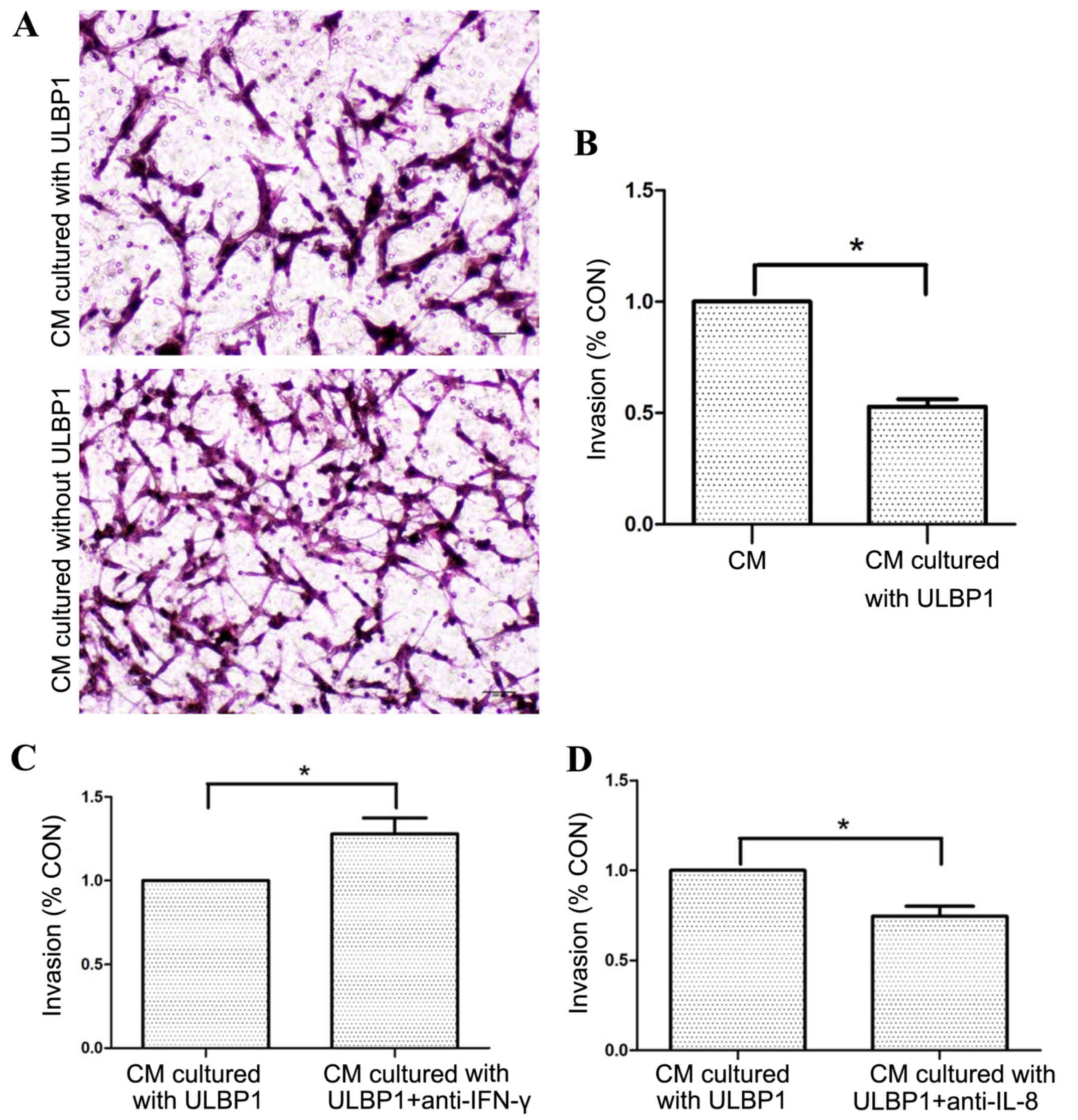 | Figure 3.uNK cell supernatants cultured with
ULBP1 inhibited HTR-8/SVneo cell invasion. (A) Representative
images of invaded cells cultured with different CM in the Transwell
invasion assay. Upper panel, the CM is uNK cell supernatants
cultured with ULBP1; lower panel, the CM is uNK cell supernatants
cultured without ULBP1; scale bars, 200 µm. Cells were stained with
hematoxylin and eosin (magnification, ×100). (B) Statistical bar
graphs exhibiting the effect of uNK cell supernatants cultured with
or without ULBP1 on the invasion of HTR-8/SVneo cells in a
Transwell invasion assay. Data are expressed as invasion index
(n=10 in duplicate). (C) Statistical bar graphs exhibiting the
effect of the addition of IFN-γ neutralizing Ab to the uNK cell
supernatants cultured with ULBP1 on invasion of HTR-8/SVneo cells
in a Transwell invasion assay (n=10 in duplicate). (D) Statistical
bar graphs exhibiting the effect of the addition of IL-8
neutralizing Ab to the uNK cell supernatants cultured with ULBP1 on
invasion of HTR-8/SVneo cells in a Transwell invasion assay (n=10
in duplicate). Data are presented as the mean + standard error of
the mean. *P<0.05. uNK, uterine natural killer; ULBP1, unique
long 16 binding protein 1; HTR-8/SVneo, extravillous trophoblast
cell line; IFN, interferon; Ab, antibody; IL, interleukin; CM,
condition medium. |
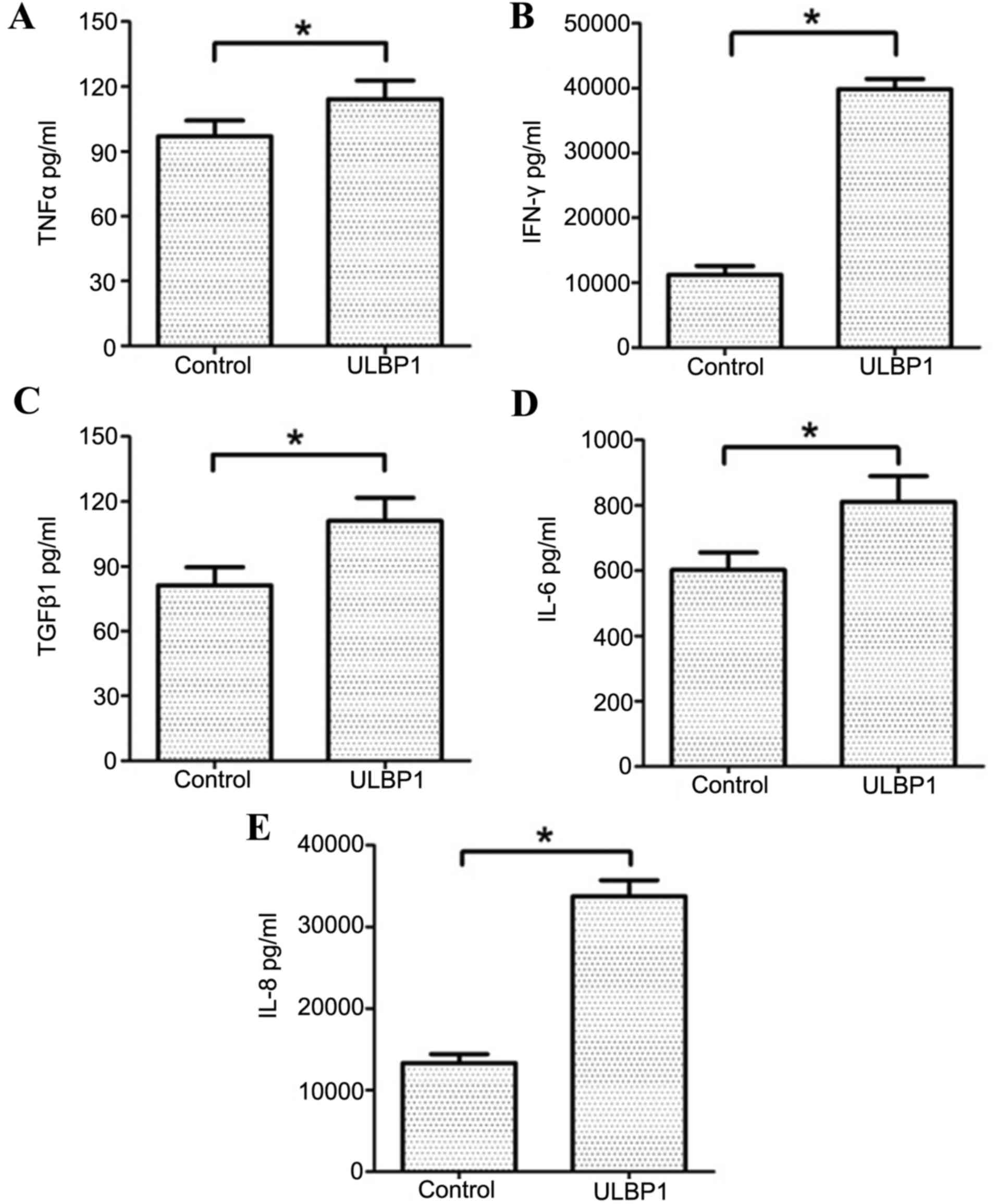
ULBP1 stimulates the production of
TNF-α, IFN-γ, TGF-β1, IL6 and IL-8 by downregulating NKG2D
expression
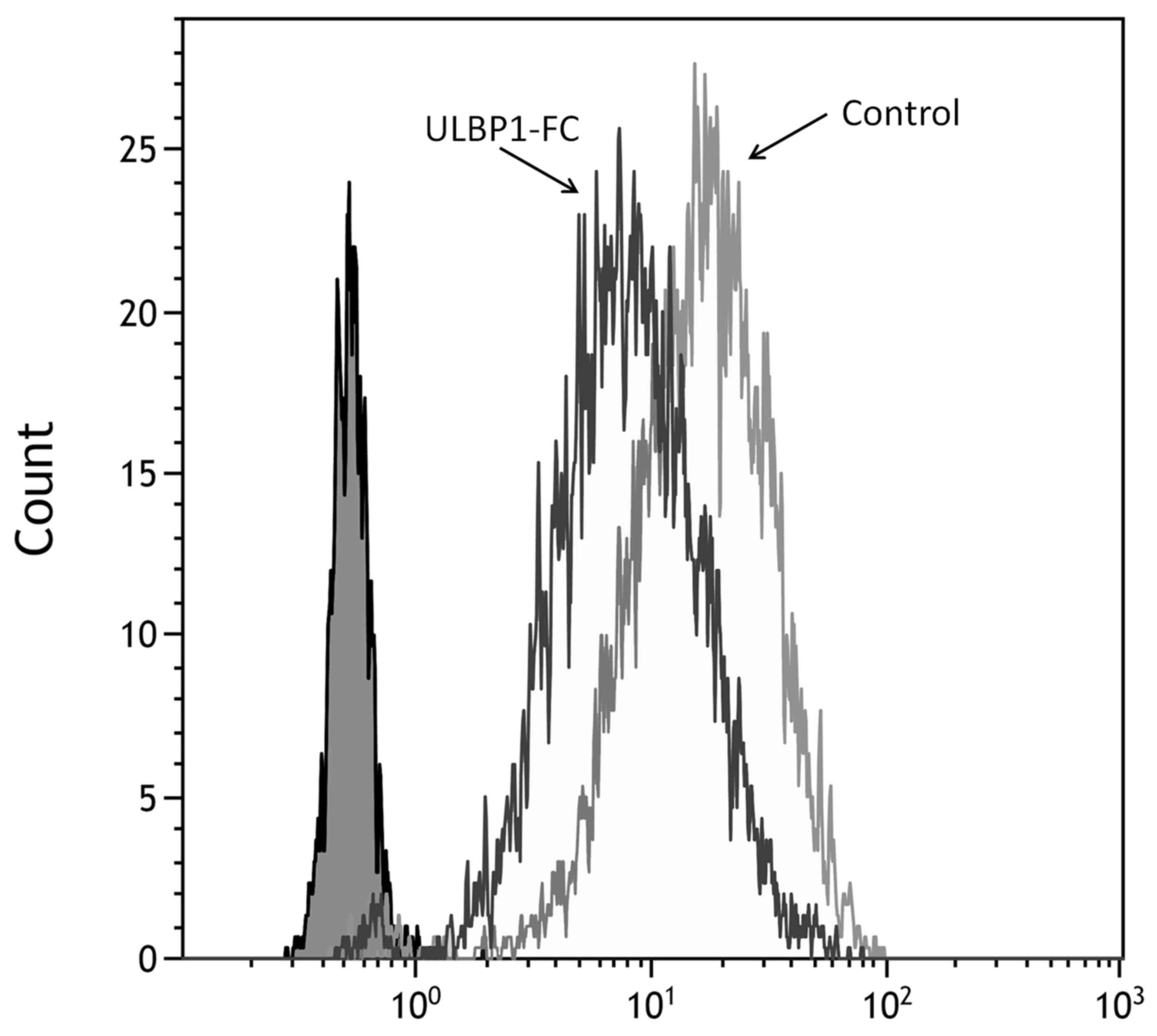
ULBP1 decreased surface NKG2D expression levels, as
monitored by staining with anti-NKG2D antibody, after incubation of
uNK cells for 72 h (Fig. 4).
Cell-free supernatants were analyzed, using ELISA, following 72 h
incubation for the presence of TNF-α, IFN-γ, TGF-β1, IL-6 and IL-8.
TNF-α, IFN-γ, TGF-β1, IL-6 and IL-8 were all significantly
increased in cell-free supernatants incubated with ULBP1
(P<0.05; Fig. 5) compared with
the controls.
Discussion
uNK cells, CD56+CD16−, are
distinct from peripheral blood (pb)NK cells and constitute up to
70% of the decidual leukocyte population in the first half of
pregnancy (24). uNK cells are
considered to have a cytokine-secreting role rather than a
cytotoxic defensive role, which is the predominant role of pbNK
cells (25). As previously
demonstrated, certain cytokines have the potential to inhibit
trophoblast invasion in in vitro invasion assays. These
cytokines include TNF-α (26),
TGF-β1 (9) and IFN-γ (27). Certain cytokines stimulate EVT
invasion, incuding IL-8 (8,28) and IL-6 (29). A study by Hanna et al
(8) demonstrated that uNK cells
induced EVT invasion; however, pbNK cells were unable to do this.
It is evident that uNK cells are important for the achievement and
maintenance of pregnancy. Although uNK cells have reduced
cytotoxic-defensive ability compared with pbNK cells, they do
retain low cytotoxic activity (30,31). uNK
cells express cytotoxic proteins, including perforin, granzymes A
and B and granulysin; therefore, uNK cells have cytolytic capacity.
This cytotoxic machinery does not result in apoptosis of the
invading trophoblast, although apoptosis may occur when responding
to infection (32); however, the
ability of uNK cells to destroy EVT remains unclear, with some
evidence demonstrating that expression of human leukocyte antigen-G
by EVT protects EVT from uNK cell attack (33).
The specific gestational period span of uNK cells
selected for the present study was based on the research of Lash
et al (34), which
demonstrated that uNK cell supernatants were able to stimulate EVT
invasion at 12–14 weeks gestation; however, they were unable to
stimulate EVT invasion at 8–10 weeks gestation. It was suggested
that, prior to 10 weeks gestational age, uNK cells had a role in
the initial stages of trophoblast-independent SA remodeling and
that, after 12 weeks gestational age, uNK cells had a role in
regulating EVT invasion. The focus of the present study was on EVT
invasion regulated by ULBP1 via uNK cells, therefore, uNK cells at
12–14 weeks were used.
NK cell function is regulated by a delicate balance
of signals initiated from various activating and inhibitory
receptors. NKG2D was originally identified, in 1991, as a key
activating receptor on all NK cells (35), including uNK cells (8). In humans, NKG2D was first identified as
a receptor for major histocompatibility complex class I
chain-related A and B and, subsequently, for ULBP 1–5 molecules
(36). Binding of ULBPs to NK cells
has several biological consequences, including the downregulation
of NKG2D expression (37) and the
increased production of cytokines and chemokines (38). Flow cytometry analysis demonstrated
that ULBP1 downregulated the expression of NKG2D on uNK cells in
the present study and, subsequently, all measured cytokines and
chemokines secreted by uNK cells increased. The effect of ULBP1 on
uNK cells was the same as pbNK cells (37).
In the present study, ULBP1 expression in the
placenta was predominately located in STB, which is consistent with
a previous study (18).
Additionally, it was demonstrated that ULBP1 expression was
upregulated in preeclamptic placenta. A study by Hedlund et
al (18) demonstrated that
ULBP1-5 were all expressed in the placenta; however, a different
study demonstrated differential expression of ULBP1 only. This may
have been due to the different binding capacity to NKG2D, which may
have induced varying secretory ability (37). Further investigation is required to
investigate whether other ULBPs are involved in preeclampsia.
As a ligand of NKG2D, ULBP1 is associated with tumor
cells (39,40). The present study demonstrated that
ULBP1 is associated with preeclampsia. NKG2D ligands may be
upregulated in response to the stresses of pregnancy, such as
hypoxia (41). Persistent hypoxia
may result in the failure of trophoblasts to differentiate from the
proliferative to invasive phenotype, resulting in shallow
trophoblast invasion and inadequate transformation of the spiral
arteries (42). Considerable
evidence supports a role for hypoxia in creating an environment
that predisposes women to implantation disorders and preeclampsia
(3,42). This may explain the upregulation of
ULBP1 in preeclampsia.
ULBP1 is produced and retained in endosomal
multivesicular bodies of the STB on exosomes in the first trimester
(18). ULBP1 may bind to uNK cells
on decidua via exosomes that are able to carry proteins to target
cells. Therefore, there may be an association between ULBP1 and EVT
invasion mediated by uNK cells. The present study focused on the
secretion from uNK cells following binding of ULBP1. The secretion
of cytokines related to the invasion of EVT was detected. The human
EVT cell line HTR-8/SVneo, which is widely used to study
trophoblast biology (43,44), was utilized in the present study. The
expression of NKG2D was downregulated when uNK cells were incubated
with ULBP1. Cytokine secretion of TNF-α, TGF-β1, IFN-γ, IL-8 and
IL6 by uNK cells significantly increased in the present study
following ULBP1 binding. Human uNK cells produce a variety of
cytokines and growth factors, including TNF-α, IL-10,
granulocyte-macrophage colony-stimulating factor, IL-1, TGF-β1,
colony-stimulating factor-1, leukemia inhibitory factor, IFN-γ
(45–47), IL-6 (9), IL-8 and interferon-inducible protein-10
(8), and the roles that these
cytokines and growth factors have in EVT invasion has been
investigated in previous studies. A study by Bauer et al
(26) demonstrated that TNF-α
inhibits trophoblast invasion through the elevation of plasminogen
activator inhibitor-1. A study by Lash et al (27) demonstrated that IFN-γ inhibits EVT
cell invasion by a mechanism that involves alterations to apoptosis
and protease levels. It was also demonstrated that invasion of EVT
cells was inhibited in the presence of TGF-β1, 2 and 3, which was
associated with a decrease in secreted matrix metalloproteinase
(MMP)-9 and urokinase plasminogen activator levels (7). A study by De Oliveira et al
(28) concluded that IL-8 stimulates
trophoblast invasion by a mechanism that may involve increased
secretion of MMP-2 by EVT. IL-6 stimulates trophoblast cell
migration and invasion, which may be partly attributable to
stimulation of expression of the integrin subunits (29); however, different results on the
effect of IL-6 on EVT invasion have been demonstrated, with the
study by Champion et al (48)
demonstrating that IL-6 has no effect on EVT invasion. Such
discrepancies between results may be attributable to differences in
cell type and quantification methods used. The present study
measured IL-6 as the cell line used was the same as in the study
conducted by Jovanović and Vićovac (29). In addition, the present study aimed
to indicate the dysfunction of uNK cells in preeclampsia by means
of measuring cytokine levels. The levels of cytokines secreted by
uNK cells, regardless of their effect on EVT invasion, became
unbalanced after ULBP1 binding to uNK.
In conclusion, trophoblast invasion is regulated by
crosstalk between trophoblasts and decidual cells in a paracrine
and autocrine manner, which involves growth factors, cytokines,
chemokines and adhesion molecules. uNK cells produce a wide range
of cytokines, chemokines and growth factors that may have diverse
effects on trophoblast invasion. Upregulation of ULBP1 disturbs the
reproductive balance on the maternal-fetal surface. This balance is
complicated and is based on various factors, rather than on one
particular factor. The results of the present study demonstrate
that ULBP1 is significantly higher in preeclampsia placentas
compared with normal placentas. As an active ligand of NK cells,
ULBP1 results in the secretion of cytokines that increase EVT
invasion. As a result of imbalance, invasion of EVT was inhibited,
which is a characteristic of preeclampsia. This finding suggests
that inappropriate crosstalk at the fetal-maternal interface
disturbs the physiological balance and induces adverse pregnancy.
Further investigation is required to determine whether it is
possible for ULBP1 to be used as a predictor for other pregnancy
complications.
References
|
1
|
Altinbas S, Toğrul C, Orhan A, Yücel M and
Danisman N: Increased MPV is not a significant predictor for
preeclampsia during pregnancy. J Clin Lab Ana. 26:403–406. 2012.
View Article : Google Scholar
|
|
2
|
Kaleli I, Kaleli B, Demir M, Yildirim B,
Cevahir N and Demir S: Serum levels of neopterin and interleukin-2
receptor in women with severe preeclampsia. J Clin Lab Anal.
19:36–39. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Amash A, Huleihel M, Sheiner E, Sapir O
and Holcberg G: Preeclampsia as a maternal vascular disease.
Harefush. 146:707–712, 733. 2007.
|
|
4
|
Kharfi A, Giguère Y, Sapin V, Massé J,
Dastugue B and Forest JC: Trophoblastic remodeling in normal and
preeclamptic pregnancies: Implication of cytokines. Clin Biochem.
36:323–331. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lala PK and Chakraborty C: Factors
regulating trophoblast migration and invasiveness: Possible
derangements contributing to pre-eclampsia and fetal injury.
Placenta. 24:575–587. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ji L, Brkić J, Liu M, Fu G, Peng C and
Wang YL: Placental trophoblast cell differentiation: Physiological
regulation and pathological relevance to preeclampsia. Mol Aspects
Med. 34:981–1023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Linzke N, Schumacher A, Woidacki K, Croy
BA and Zenclussen AC: Carbon monoxide promotes proliferation of
uterine natural killer cells and remodeling of spiral arteries in
pregnant hypertensive heme oxygenase-1 mutant mice. Hypertension.
63:580–588. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hanna J, Goldman-Wohl D, Hamani Y, Avraham
I, Greenfield C, Natanson-Yaron S, Prus D, Cohen-Daniel L, Arnon
TI, Manaster I, et al: Decidual NK cells regulate key developmental
processes at human fetal-maternal interface. Nat Med. 12:1065–1074.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lash GE, Otun HA, Innes BA, Bulmer JN,
Searle RF and Robson SC: Inhibition of trophoblast cell invasion by
TGFβ1, 2, and 3 is associated with a decrease in active proteases.
Biol Reprod. 73:374–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Williams PJ, Bulmer JN, Searle RF, Innes
BA and Robson SC: Altered decidual leucocyte populations in the
placental bed in pre-eclampsia and foetal growth restriction: A
comparison with late normal pregnancy. Reproduction. 138:177–184.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stallmach T, Hebisch G, Orban P and Lu X:
Aberrant positioning of trophoblast and lymphocytes in the
feto-maternal interface with pre-eclampsia. Virchows Arch.
434:207–211. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao W, Xi X, Hao Z, Li W, Kong Y, Cui L,
Ma C, Ba D and He W: RAET1E2, a soluble isoform of the UL16-binding
protein RAET1E produced by tumor cells, inhibits NKG2D-mediated NK
cytotoxicity. J Biol Chem. 282:18922–18928. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salih HR, Antropius H, Gieseke F, Lutz SZ,
Kanz L, Rammensee HG and Steinle A: Functional expression and
release of ligands for the activating immunoreceptor NKG2D in
leukemia. Blood. 102:1389–1396. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pende D, Rivera P, Marcenaro S, Chang CC,
Biassoni R, Conte R, Kubin M, Cosman D, Ferrone S, Moretta L and
Moretta A: Major histocompatibility complex class I-related chain A
and UL16-binding protein expression on tumor cell lines of
different histotypes: Analysis of tumor susceptibility to
NKG2D-dependent natural killer cell cytotoxicity. Cancer Res.
62:6178–6186. 2002.PubMed/NCBI
|
|
15
|
Mincheva-Nilsson L, Nagaeva O, Chen T,
Stendahl U, Antsiferova J, Mogren I, Hernestål J and Baranov V:
Placenta-derived soluble MHC class I chain-related molecules
down-regulate NKG2D receptor on peripheral blood mononuclear cells
during human pregnancy: A possible novel immune escape mechanism
for fetal survival. J Immunol. 176:3585–3592. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Upshaw JL and Leibson PJ: NKG2D-mediated
activation of cytotoxic lymphocytes: Unique signalling pathways and
distinct functional outcomes. Semin Immunol. 18:167–175. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Borchers MT, Harris NL, Wesselkamper SC,
Vitucci M and Cosman D: NKG2D ligands are expressed on stressed
human airway epithelial cells. Am J Physiol. 291:L222–L231.
2006.
|
|
18
|
Hedlund M, Stenqvist AC, Nagaeva O,
Kjellberg L, Wulff M, Baranov V and Mincheva-Nilsson L: Human
placenta expresses and secretes NKG2D ligands via exosomes that
down-modulate the cognate receptor expression: Evidence for
immunosuppressive function. J Immunol. 183:340–351. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meng T, Chen H, Sun M, Wang H, Zhao G and
Wang X: Identification of differential gene expression profiles in
placentas from preeclamptic pregnancies versus normal pregnancies
by DNA microarrays. OMICS. 16:301–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Steegers EA, von Dadelszen P, Duvekot JJ
and Pijnenborg R: Pre-eclampsia. Lancet. 376:631–644. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lash GE, Schiessl B, Kirkley M, Innes BA,
Cooper A, Searle RF, Robson SC and Bulmer JN: Expression of
angiogenic growth factors by uterine natural killer cells during
early pregnancy. J Leukoc Biol. 80:572–580. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vassiliadou N and Bulmer JN: Functional
studies of human decidua in spontaneous early pregnancy loss:
Effect of soluble factors and purified CD56+ lymphocytes on killing
of natural killer- and lymphokine-activated killer-sensitive
targets. Biol Reprod. 58:982–987. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moffett-King A: Natural killer cells and
pregnancy. Nat Rev Immunol. 2:656–663. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tabiasco J, Rabot M, Aguerre-Girr M, El
Costa H, Berrebi A, Parant O, Laskarin G, Juretic K, Bensussan A,
Rukavina D and Le Bouteiller P: Human decidual NK cells: Unique
phenotype and functional properties-a review. Placenta. 27 Suppl
A:S34–S39. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bauer S, Pollheimer J, Hartmann J,
Husslein P, Aplin JD and Knöfler M: Tumor necrosis factor-alpha
inhibits trophoblast migration through elevation of plasminogen
activator inhibitor-1 in first-trimester villous explant cultures.
J Clin Endocrinol Metab. 89:812–822. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lash GE, Otun HA, Innes BA, Kirkley M, De
Oliveira L, Searle RF, Robson SC and Bulmer JN: Interferon-gamma
inhibits extravillous trophoblast cell invasion by a mechanism that
involves both changes in apoptosis and protease levels. FASEB J.
20:2512–2518. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De Oliveira LG, Lash GE, Murray-Dunning C,
Bulmer JN, Innes BA, Searle RF, Sass N and Robson SC: Role of
interleukin 8 in uterine natural killer cell regulation of
extravillous trophoblast cell invasion. Placenta. 31:595–601. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jovanović M and Vićovac L: Interleukin-6
stimulates cell migration, invasion and integrin expression in
HTR-8/SVneo cell line. Placenta. 30:320–328. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
King A, Birkby C and Loke YW: Early human
decidual cells exhibit NK activity against the K562 cell line but
not against first trimester trophoblast. Cell Immunol. 118:337–344.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ritson A and Bulmer JN: Isolation and
functional studies of granulated lymphocytes in first trimester
human decidua. Clin Exp Immunol. 77:263–268. 1989.PubMed/NCBI
|
|
32
|
Le Bouteiller P, Siewiera J, Casart Y,
Aguerre-Girr M, El Costa H, Berrebi A, Tabiasco J and
Jabrane-Ferrat N: The human decidual NK-cell response to virus
infection: What can we learn from circulating NK lymphocytes? J
Reprod Immunol. 88:170–175. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rouas-Freiss N, Gonçalves RM, Menier C,
Dausset J and Carosella ED: Direct evidence to support the role of
HLA-G in protecting the fetus from maternal uterine natural killer
cytolysis. Proc Natl Acad Sci USA. 94:11520–11525. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lash GE, Otun HA, Innes BA, Percival K,
Searle RF, Robson SC and Bulmer JN: Regulation of extravillous
trophoblast invasion by uterine natural killer cells is dependent
on gestational age. Hum Reprod. 25:1137–1145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Houchins JP, Yabe T, McSherry C and Bach
FH: DNA sequence analysis of NKG2, a family of related cDNA clones
encoding type II integral membrane proteins on human natural killer
cells. J Exp Med. 173:1017–1020. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
González S, López-Soto A, Suarez-Alvarez
B, López-Vázquez A and López-Larrea C: NKG2D ligands: Key targets
of the immune response. Trends Immunol. 29:397–403. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song H, Kim J, Cosman D and Choi I:
Soluble ULBP suppresses natural killer cell activity via
down-regulating NKG2D expression. Cell Immunol. 239:22–30. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kubin M, Cassiano L, Chalupny J, Chin W,
Cosman D, Fanslow W, Müllberg J, Rousseau AM, Ulrich D and Armitage
R: ULBP1, 2, 3 Novel MHC class I related molecules that bind to
human cytomegalovirus glycoprotein UL16, activate NK cells. Eur J
Immunol. 31:1428–1437. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim SJ, Ha GH, Bae JH, Kim GR, Son CH,
Park YS, Yang K, Oh SO, Kim SH and Kang CD: COX-2- and endoplasmic
reticulum stress-independent induction of ULBP-1 and enhancement of
sensitivity to NK cell-mediated cytotoxicity by celecoxib in colon
cancer cells. Exp Cell Res. 330:451–459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cho H, Chung JY, Kim S, Braunschweig T,
Kang TH, Kim J, Chung EJ, Hewitt SM and Kim JH: MICA/B and ULBP1
NKG2D ligands are independent predictors of good prognosis in
cervical cancer. BMC Cancer. 14:9572014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Weier JF, Weier HU, Jung CJ, Gormley M,
Zhou Y, Chu LW, Genbacev O, Wright AA and Fisher SJ: Human
cytotrophoblasts acquire aneuploidiesas they differentiate to an
invasive phenotype. Dev Biol. 279:420–432. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Caniggia I, Mostachfi H, Winter J,
Gassmann M, Lye SJ, Kuliszewski M and Post M: Hypoxia-inducible
factor-1 mediates the biological effects of oxygen on human
trophoblast differentiation through TGFbeta(3). J Clin Invest.
105:577–587. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Suman P, Godbole G, Thakur R,
Morales-Prieto DM, Modi DN, Markert UR and Gupta SK: AP-1
transcription factors, mucin-type molecules and MMPs regulate the
IL-11 mediated invasiveness of JEG-3 and HTR-8/SVneo trophoblastic
cells. PLoS One. 7:e297452012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Graham CH, Hawley TS, Hawley RG,
MacDougall JR, Kerbel RS, Khoo N and Lala PK: Establishment and
characterization of first trimester human trophoblast cells with
extended lifespan. Exp Cell Res. 206:204–211. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jokhi PP, King A, Sharkey AM, Smith SK and
Loke YW: Screening for cytokine messenger ribonucleic acids in
purified human decidual lymphocyte populations by the
reverse-transcriptase polymerase chain reaction. J Immunol.
153:4427–4435. 1994.PubMed/NCBI
|
|
46
|
Jokhi PP, King A and Loke YW: Cytokine
production and cytokine receptor expression by cells of the human
first trimester placental-uterine interface. Cytokine. 9:126–137.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rieger L, Kammerer U, Hofmann J, Sütterlin
M and Dietl J: Choriocarcinoma cells modulate the cytokine
production of decidual large granular lymphocytes in coculture. Am
J Reprod Immunol. 46:137–143. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Champion H, Innes BA, Robson SC, Lash GE
and Bulmer JN: Effects of interleukin-6 on extravillous trophoblast
invasion in early human pregnancy. Mol Hum Reprod. 18:391–400.
2012. View Article : Google Scholar : PubMed/NCBI
|