Introduction
In recent years, due to improvements in living
standards and an ageing population, the incidence and morbidity of
hypertension, coronary heart disease (CHD), diabetes, pulmonary
hypertension and other cardiovascular diseases has increased(6 in
many countries around the world (1–3).
Cardiovascular disease has become one of the major diseases
threatening human health. According to the latest statistics from
the World Health Organization, each year 17.3 million people
succumb to diversified cardiovascular and cerebrovascular diseases,
accounting for ~30% of the global mortality rate, which is
considerably higher than the mortality rate of other diseases,
including cancer and HIV/AIDS (4).
Thus cardiovascular disease has been referred to as the number one
enemy of the second health revolution (5).
It has been demonstrated that the transforming
growth factor (TGF)β1/SMAD signaling pathway, which participates in
the formation and progression of coronary atherosclerosis,
remodeling following myocardial infarction, cell apoptosis and
inflammatory reactions, is a crucial regulator during the genesis
and development of CHD and has become a topic of considerable
interest within the field of CHD (6). Different inflammatory cytokines may
induce various types of cardiovascular and cerebrovascular diseases
in patients with CHD (7). TGF-β1 has
attracted widespread attention since it serves multiple important
roles in certain important disease processes (8). TGFβ binds a heterodimer of TGFβ type I
and type II receptors (9) and the
downstream signaling molecules TGFβ1, SMAD2 and SMAD3 are
phosphorylated and consequently activated (10). Polymers are formed from the activated
SMAD2 and SMAD3, together with SMAD4, and are subsequently
translocated to the cell nucleus to regulate corresponding gene
expression (11).
Rutin, also referred to as rutoside and purple
quercetin, is a flavonoid compound that may be obtained from a wide
variety of sources (12).
Flavonoids, of which rutin is a typical representative, serve a
positive role in the treatment of cardiovascular and
cerebrovascular diseases (13).
Rutin is an active component of medicinally active plants, and a
topic of recent research (14). Its
effects include maintaining blood vessel elasticity, reducing blood
vessel brittleness and lowering capillary permeability, as well as
preventing cerebral hemorrhage, retinal hemorrhage and other
complications induced by hypertension (12,15).
Therefore, the aims of the current study were to investigate
whether rutin is able to inhibit CHD and to assess its underlying
mechanisms of action on CHD.
Materials and methods
Animals and reagents
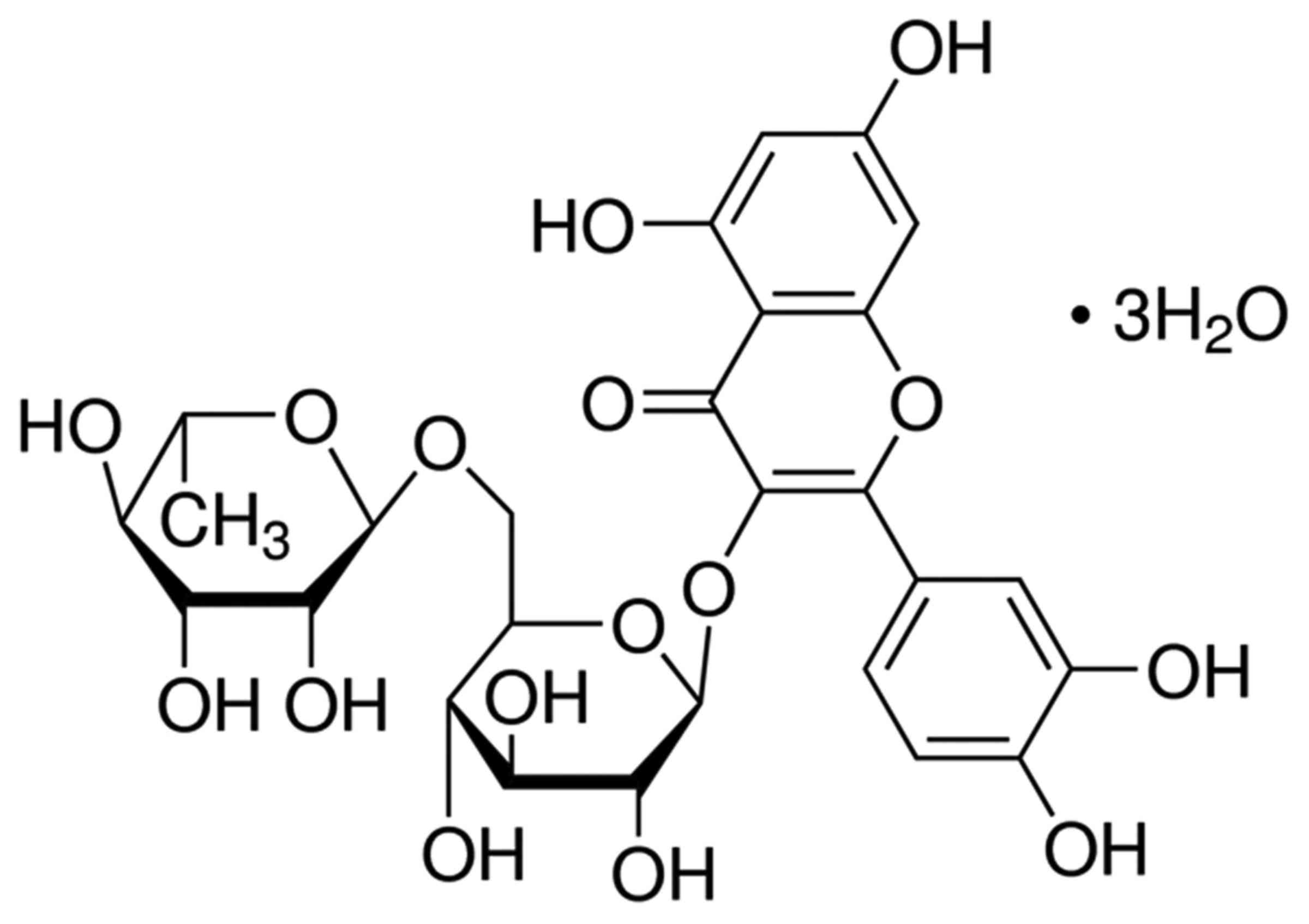
Rutin was acquired from Sigma-Aldrich (Merck
Millipore; Darmstadt, Germany). The chemical structure of rutin is
presented in Fig. 1. Male Chinese
miniature pigs (n=24; weight range, 20–30 kg) were acquired from
the Institute of Laboratory Animal Science, Jining Medical
University (Jining, China). Pigs had ad libitum access to a
standard laboratory diet (3% fat) and water and were housed in a
climate-controlled farmhouse at 22–24°C, 60–70% humidity and a 12-h
light-dark cycle. The animals were randomly assigned to one of four
groups: CHD model (n=6), CHD model + 15 mg/kg rutin treatment
(n=6), CHD model + 45 mg/kg rutin treatment (n=6), and
sham-operated control groups (n=6). Pigs in the CHD model group and
the two rutin treatment + CHD groups were fed with a high-fat diet
(20%) for 2 weeks. The pigs were anesthetized with 30 mg/kg sodium
barbital (Sigma-Aldrich; Merck Millipore) via the ear vein prior to
modeling. The common carotid artery was isolated and, following the
embedding of a 6F arterial sheath and injection of 200 U/kg
heparin, the distal heart was ligated. A guidewire was advanced
into the left anterior descending artery and anangioplasty balloon
(VistaBrite; Cordis Corporation, Hialeah, FL, USA) was introduced
into the left anterior descending branch and pressured with air
(10–12 atm) for 30 sec three times, following maintenance of the
balloon pressure air gap (1–1.5 atm). Following removal of the
catheter and balloon, the common carotid artery was ligated. Pigs
in the CHD group were fed a high-fat diet for the following 8
weeks, and those in the CHD model + rutin groups, which were also
fed a high-fat diet, were injected with rutin (15 or 45 mg/kg) for
eight weeks (16). Meanwhile, pigs
in the sham group were administered with normal saline. Ethical
approval was granted for the present study by the Ethics Committee
of the First School of Clinical Medicine at Jining Medical
University.
Echocardiography
At the end of the treatment period, anS5-1 linear
probe with an iE33 xMatrix Ultrasound instrument (Philips
Healthcare, Andover, MA, USA) was used to perform an
echocardiographic analysis. The left ventricular ejection fraction
(LVEF) and systolic internal diameter (LVID) were calculated.
Blood and urine chemistry
At the end of the treatment period, pigs were
anesthetized (20–30 mg/kg pentobarbital sodium; Sigma-Aldrich;
Merck Millipore) and blood was sampled through the eyes. Urine
protein concentration, blood urea nitrogen (BUN) and plasma
creatinine (Scr) levels were measured using ELISA kits (BUN,
C013-1; Scr, C011-2; Nanjing Jiancheng, Ltd., Nanjing, China).
Assessment of the infarcted
myocardium
All pigs were sacrificed via exsanguination
following administration of 30 mg/kg pentobarbital sodium at 8
weeks after treatment. The left ventricle was washed with
physiological saline and refrigerated at −80°C for 5–10 min. Heart
tissue samples were cut into 2 mm-thick sections. The tissue
sections were stained with 1% 2,3,5-triphenyltetrazolium chloride
(Sigma-Aldrich; Merck Millipore) for 30 min at room temperature.
The infarct size was measured by volume and weight as a percentage
of the left ventricle.
Assessment of antioxidant profile
The left ventricle from each pig was homogenized
with radioimmunoprecipitation assay (RIPA) lysis buffer (Nanjing
Jiancheng, Ltd.) and centrifuged at 12,000 × g for 10 min at
4°C. Activities of superoxide dismutase (SOD), catalase (CAT),
glutathione-S-transferase (GST) and glutathione peroxidase (GSH-Px)
were determined using ELISA kits (Wuhan Elabscience Biotechnology
Co., Ltd., Wuhan, China) according to the manufacturer's
instructions.
Western blot analysis
Heart tissue lysate was prepared using an RIPA lysis
buffer (Nanjing Jiancheng, Ltd.) and quantified using the Bradford
protein assay (Nanjing Jiancheng Ltd.). Equal amounts of protein
(50 µg) were separated using 10–12% dodecyl sulfate-polyacrylamide
gel electrophoresis (SDS-PAGE) and electroblotted onto
nitrocellulose membranes. Following blocking with 5% bovine serum
albumin (Nanjing Jiancheng, Ltd.) at 37°C for 1 h, the membranes
were incubated with antibodies against TGFβ1 (sc-130348; 1:500;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), SMAD2 (sc-101153;
1:500; Santa Cruz Biotechnology, Inc.),
phosphorylated-extracellular signal-regulated kinases (p-ERK) 1/2
(sc-136521; 1:500; Santa Cruz Biotechnology, Inc.), anti-p-Akt
(sc-7985-R; 1:500; Santa Cruz Biotechnology, Inc.) and β-actin
(sc-47778; 1:2,000; Santa Cruz Biotechnology, Inc.) overnight at
4°C. Membranes were then incubated with a horseradish
peroxidase-conjugated antibody against rabbit or mouse IgG at a
1:2,000 dilution (sc-2005 and sc-358914; Santa Cruz Biotechnology,
Inc.) for 1 h at room temperature and viewed using enhanced
chemiluminescence (ECL) reagent (Nanjing Beyotime Ltd.) in
accordance with the manufacturer's instructions. Protein bands were
analyzed and quantified using Image J version 3.0 (National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All values are presented as the mean ± standard
error and were analyzed by analysis of variance and post hoc
Bonferroni test. P<0.01 was considered to indicate a
statistically significant difference.
Results
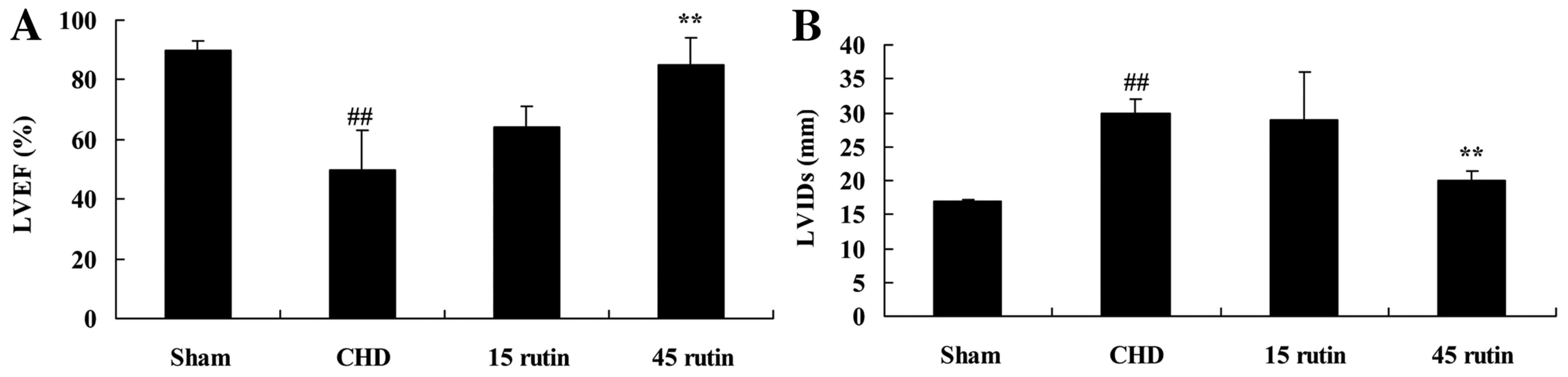
Rutin changes LVEF and LVID values in
CHD model pigs
Following treatment with rutin for eight weeks, the
mean LVEF of the CHD model pigs was lower than that of the sham
group (P<0.01), and the mean LVID of the CHD model pigs was
higher than that of the sham group (P<0.01). However, treatment
of CHD model pigs with 45 mg/kg rutin attenuated LVEF and the
increase in LVID that occurred following CHD modeling in the
absence of rutin treatment (P<0.01; Fig. 2).
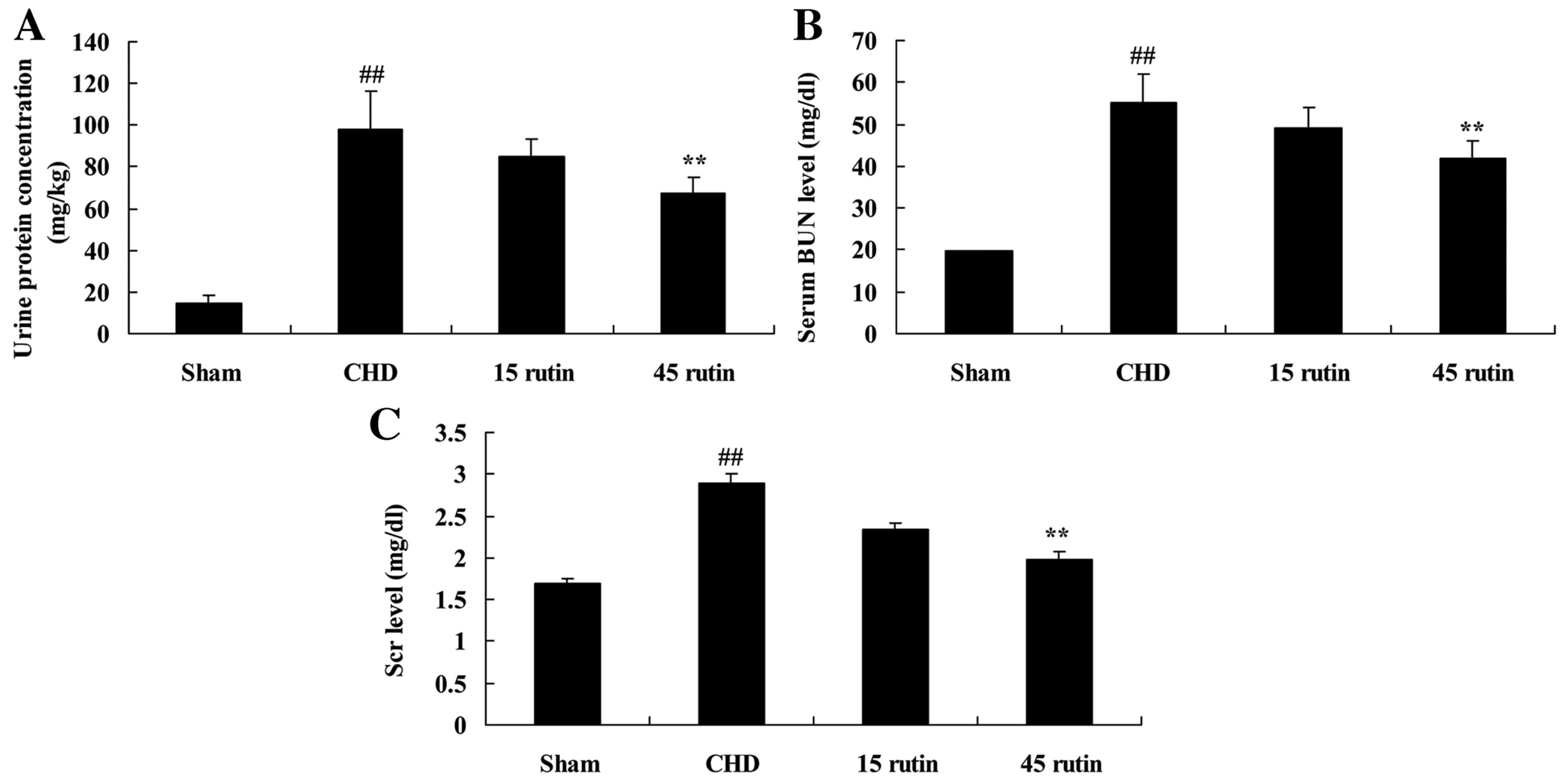
Rutin changes biochemical parameters
in CHD model pigs
Compared with the sham-operated control group, the
CHD group exhibited a significant elevation of urine protein
concentration, serum BUN and Scr expression levels (P<0.01;
Fig. 3). The administration of 45
mg/kg rutin significantly attenuated the CHD-induced increases in
urine protein concentration, BUN and Scr expression levels
(P<0.01; Fig. 3).
Rutin reduces infarction of the
myocardium in CHD model pigs
Rutin had a protective effect against myocardial
infarction in CHD model pigs. CHD induced infarction of the
myocardium, which did not occur in the sham-operated control group
(P<0.01). However, the infarct size was significantly reduced
following treatment with 45 mg/kg rutin compared with the infarct
size in CHD model pigs (P<0.01; Fig.
4).
Rutin increases antioxidant activities
in CHD model pigs
The CHD model pigs exhibited an inhibited
antioxidant profile compared with the sham-operated control group.
The activities of the antioxidant enzymes SOD, CAT, GST and GSH-Px
were significantly reduced in the CHD group (P<0.01 vs. the sham
control; Fig. 5). This reduction in
SOD, CAT, GST and GSH-Px activity was significantly attenuated by
treatment with 45 mg/kg rutin (P<0.01; Fig. 5).
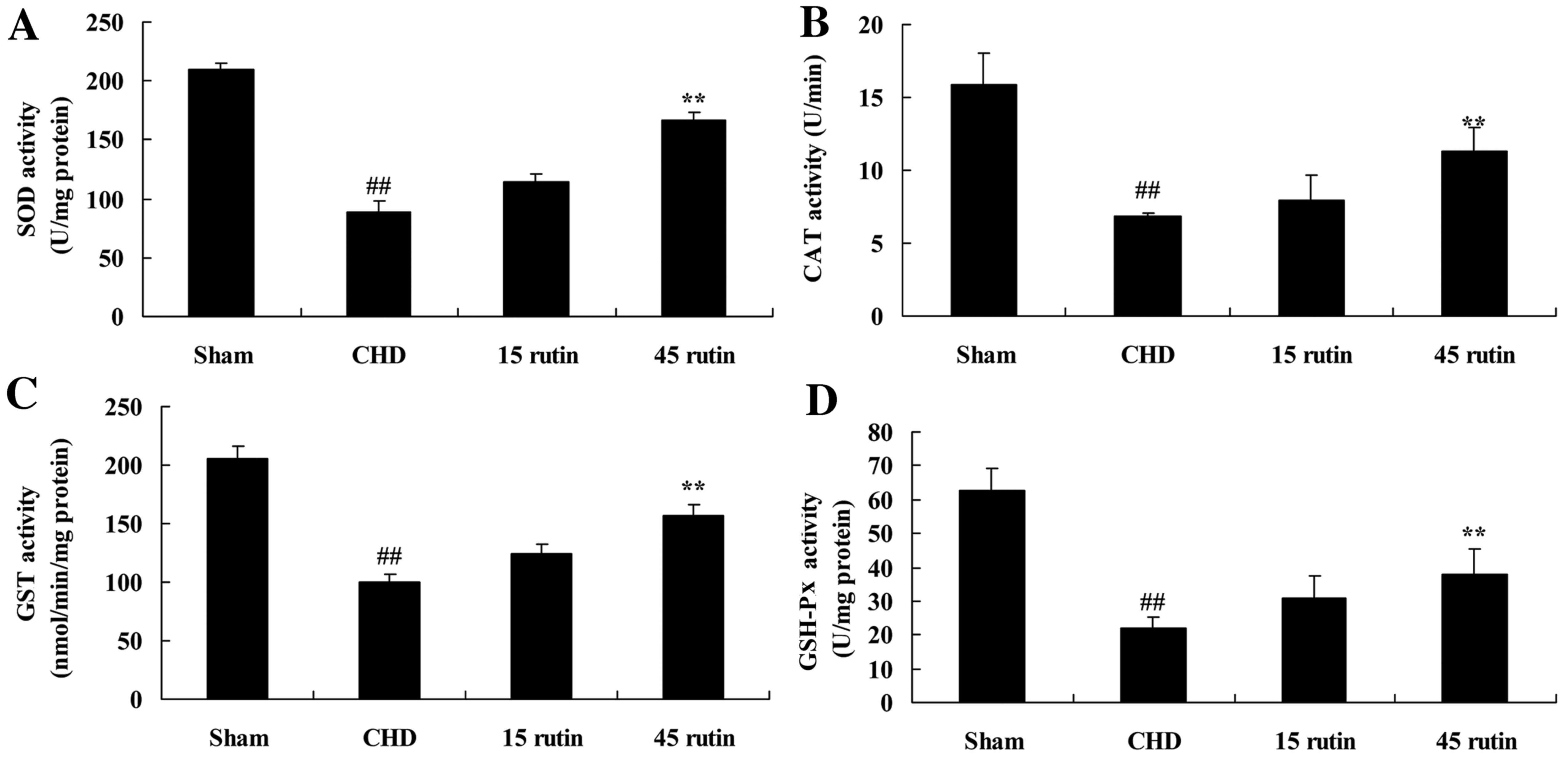 | Figure 5.Rutin changes the antioxidant profile
in CHD model pigs. (A) SOD, (B) CAT, (C) GST and (D) GSH-Px
activity in the four groups. n=3 per group. ##P<0.01
vs. the sham group; **P<0.01 vs. the CHD model group. SOD,
superoxide dismutase; CAT, catalase; GST,
glutathione-S-transferase; GSH-Px, glutathione peroxidase; sham,
sham-operated control group; CHD, coronary heart disease model
group; 15 rutin, 15 mg/kg rutin treated CHD group; 45 rutin, 45
mg/kg rutin treated CHD group. |
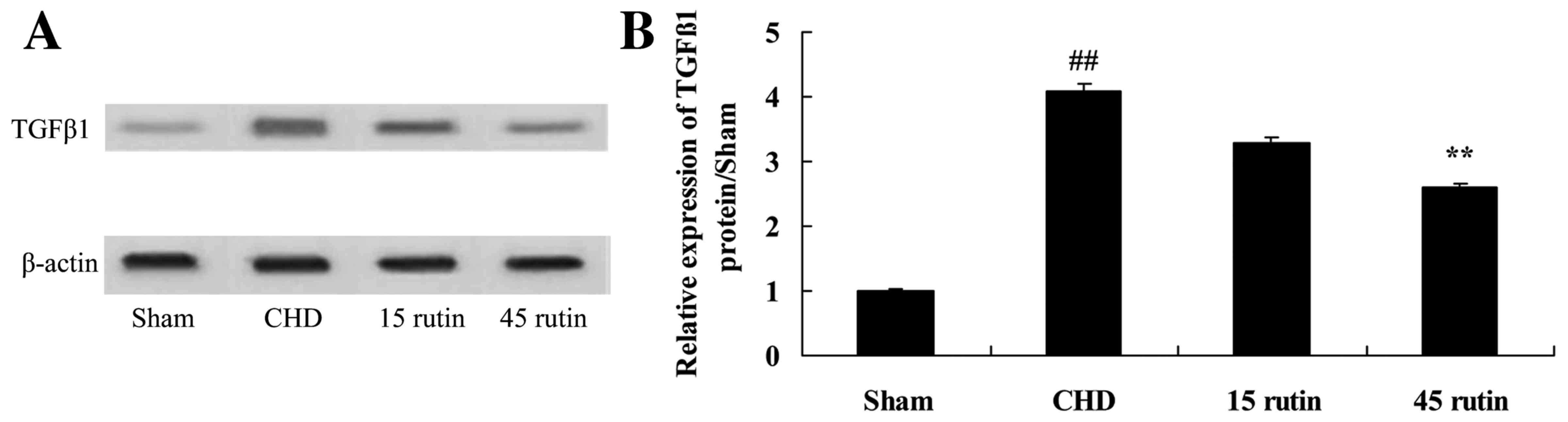
Rutin reduces TGFβ1 expression in CHD
model pigs
TGFβ1 protein expression levels were increased in
the CHD model pigs compared with the sham-operated control group
(P<0.01; Fig. 6). However, the
CHD-induced increase in TGFβ1 protein expression was significantly
attenuated following treatment with 45 mg/kg rutin (P<0.01;
Fig. 6).
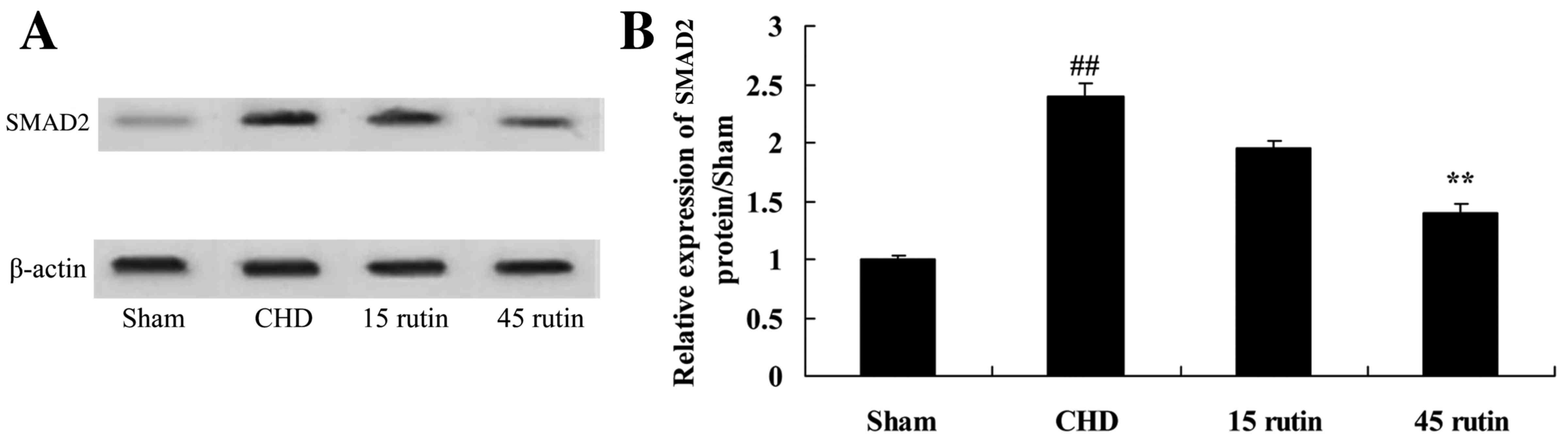
Rutin changes SMAD2 expression in CHD
model pigs
Western blotting analyses indicated that expression
of SMAD2 protein was increased in the CHD pigs compared with the
sham-operated control group (P<0.01; Fig. 7). However, the CHD model pigs that
received treatment with 45 mg/kg rutin exhibited decreased
expression levels of SMAD2 protein compared with the CHD group
(P<0.01).
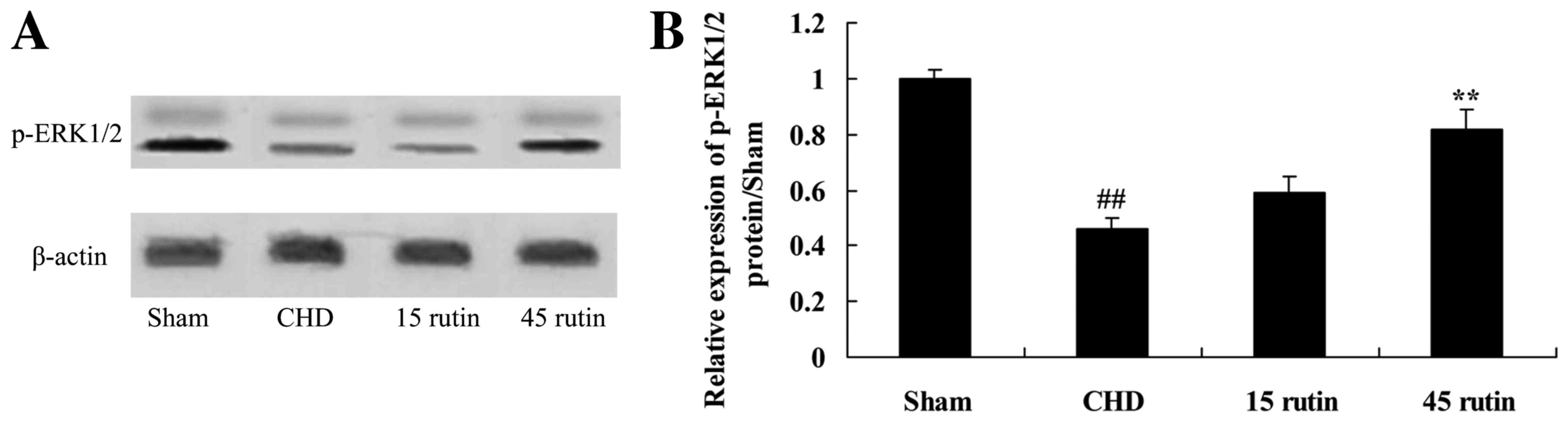
Rutin increases p-ERK1/2 expression in
CHD model pigs
There was a reduction in p-ERK1/2 protein expression
in the CHD model pigs compared with the sham-operated control group
(P<0.01; Fig. 8). However,
p-ERK1/2 protein expression was restored in CHD model pigs treated
with 45 mg/kg rutin compared with the untreated CHD group
(P<0.01; Fig. 8).
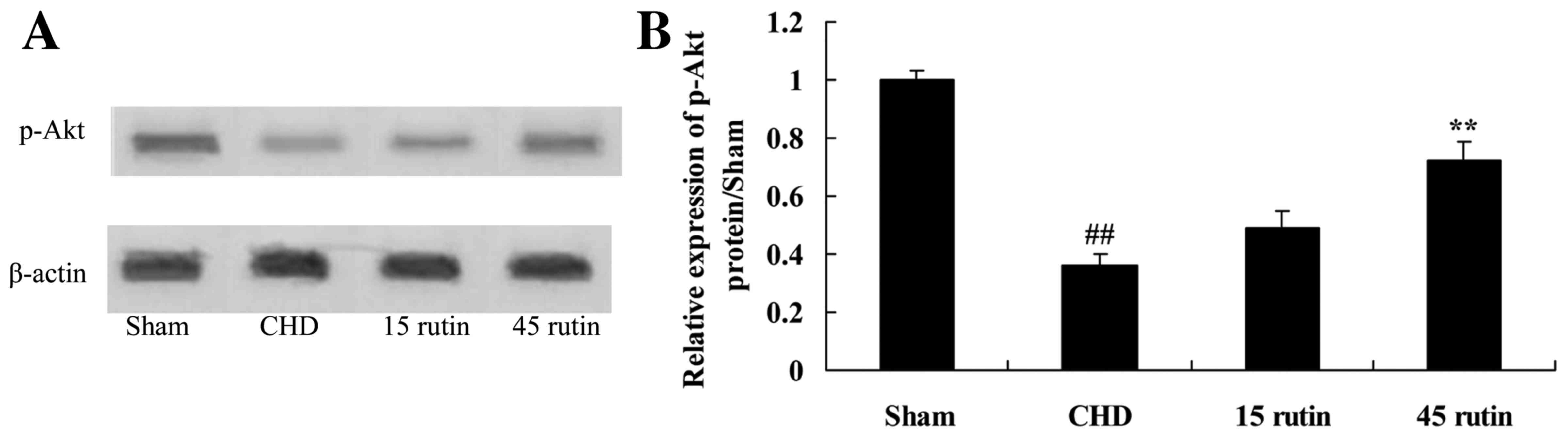
Rutin increases p-Akt expression in
CHD model pigs
Compared with the sham-operated control group, p-Akt
protein expression of the CHD model pigs was suppressed (P<0.01;
Fig. 9). Treatment with rutin
increased the p-Akt protein expression in the CHD model pigs
(P<0.01; Fig. 9).
Discussion
CHD is a set of clinical syndromes presenting as
different degrees of myocardial ischemia caused by coronary
atherosclerotic plaque rupture, which is accompanied by platelet
aggregation, thrombosis and different degrees of stenosis that may
involve the complete occlusion of coronary vessels (17). It poses a great threat to human
health and life due to its characteristics of emergent onset,
further complications and a high mortality rate (18,19). The
morbidity and mortality of coronary heart disease in China has been
gradually rising, and the rate of increase appears to be escalating
(20,21). In the present study of a porcine
model of CHD, rutin treatment significantly increased the LVEF,
reduced the LVID, attenuated the increase in urine protein
concentration, BUN and Scr expression levels induced by CHD, and
limited the area of infarcted myocardium.
Oxidative stress occurs when the body is exposed to
various types of noxious stimuli, and excessive quantities of
highly reactive molecules, such as reactive oxygen species and
reactive nitrogen species are generated (20). When the generation of oxidative
species exceeds their removal, an imbalance in the oxidation and
antioxidant system occurs, leading to vascular endothelial tissue
damage (21). The oxidized
low-density lipoprotein cholesterol (OX-LDL) that is generated by
oxidative stress has an important effect on atherosclerosis
(22). It directly and indirectly
contributes to atherosclerosis via a variety of mechanisms,
including endothelial cell injury and endothelium dysfunction; in
addition, LDL in the circulation is promoted to enter subcutaneous
clearance (23). LDL has a
chemotactic effect on monocytes and T lymphocytes (24). Furthermore, it may inhibit the
activity of macrophages, causing them to remain within the arterial
wall to increase inflammatory reactions of the local blood vessel
wall (25). The present study
demonstrated that treatment with rutin significantly reversed the
CHD-induced reductions of SOD, CAT, GST and GSH-Px activities in
CHD model pigs. This builds on the results from Yu et al
(14) who previously found that
rutin inhibits amylin-induced neurocytotoxicity and oxidative
stress.
The signaling pathways associated with TGFβ1/SMAD2
serve an important role in the biological functions of TGFβ1,
including induction of cell proliferation, immune suppression and
inflammation (26). A significant
enhancement was previously observed between TGFβ1 expression in
patients with hypertrophic cardiomyopathy and dilated
cardiomyopathy in the animal model (27). In addition, a number of studies have
indicated that the TGFβ signaling pathway may inhibit the
inflammatory gene expression caused by myocardial infarction with
inflammatory infiltration (28,29).
TGFβ1 is of great significance in various pathological processes,
including infarction, myocardial remodeling and interstitial
fibrosis (30). An analysis of TGFβ1
and SMAD2 levels and their correlation with CHD risk factors
indicated that TGFβ1 is associated with the expression of
lipoprotein A and uric acid (31,32).
Increased levels of lipoprotein A and uric acid are considered as
markers of atherosclerosis, which indicates that TGFβ1 may regulate
atherosclerosis, thereby accelerating the establishment of CHD. The
level of SMAD2 is closely associated with blood glucose content,
which indicates that SMAD2 may affect the onset and development of
CHD and diabetes mellitus (11). The
current study observed that treatment with rutin significantly
suppressed the CHD-induced TGFβ1/SMAD2 signaling pathway in CHD
model pigs. This is in accordance with a previous study by Han
et al (16), which suggested
that rutin ameliorates renal fibrosis and proteinuria by activating
TGFβ1-SMAD2 signaling in 5/6-nephrectomized rats.
The ERK1/2 signaling pathway is a typical
mitogen-activated protein kinase (MAPK) signal transduction pathway
(33). It may be activated by
insulin, growth factors and serum (33). Furthermore, it is activated through
the phosphorylation of numerous types of growth factors, ion beam
radiation and hydrogen peroxide (34). ERK1/2 enters the cell nucleus and
acts as a transcription factor, accelerating the transcription and
expression of certain genes and is closely associated with cell
proliferation and differentiation (35). Studies have indicated that ERK1/2
mediates the stress and inflammatory response (36,37).
ERK1/2 is an essential signal transduction pathway in the MAPK
family and it has been demonstrated that ERK1/2 is closely
associated with atherosclerosis (38). Jeong et al (39) suggested that rutin inhibits
myocardial ischemia/reperfusion-induced apoptosis via ERK1/2 and
PI3 K/Akt signaling in vitro, and the current study
indicated that treatment with rutin recovered p-ERK1/2 protein
expression in CHD model pigs.
Akt activates NF-κB and other nuclear transcription
factors, produces a large number of pro-inflammatory cytokines and
induces or maintains inflammation (40). It also stimulates sustained cell
growth, inhibits cell apoptosis and promotes carcinogenesis
(40). In the present study, it was
demonstrated that treatment with rutin increased p-Akt protein
expression levels in CHD model pigs. This is in agreement with a
study by Moghbelinejad et al (41), which indicated that rutin may be
protective against the neurotoxic effects of amyloid β on memory in
rats through actions on ERK and PI3K/Akt.
In conclusion, rutin inhibits CHD in a porcine
model; this may be due to an anti-oxidative effect, suppression of
TGFβ1 and SMAD2 and the modulation of ERK1/2 and Akt signaling. The
results of the current study support the notion that rutin may be
developed as a novel therapeutic strategy to treat CHD in the
future.
Acknowledgements
The present study was supported by a grant from the
Natural Science Foundation of Shandong Province (grant no.
Q2007C10).
References
|
1
|
Ueno T, Chow LW and Toi M: Increases in
circulating VEGF levels during COX-2 inhibitor treatment in breast
cancer patients. Biomed Pharmacother. 60:277–279. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chuah BY, Putti T, Salto-Tellez M,
Charlton A, Iau P, Buhari SA, Wong CI, Tan SH, Wong AL, Chan CW, et
al: Serial changes in the expression of breast cancer-related
proteins in response to neoadjuvant chemotherapy. Ann Oncol.
22:1748–1754. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kullo IJ, Jouni H, Olson JE, Montori VM
and Bailey KR: Design of a randomized controlled trial of
disclosing genomic risk of coronary heart disease: The myocardial
infarction genes (MI-GENES) study. BMC Med Genomics. 8:512015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pradeepa R, Surendar J, Indulekha K,
Chella S, Anjana RM and Mohan V: Prevalence of metabolic syndrome
and its association with coronary artery disease among an urban
elderly south indian population (CURES- 145). J Assoc Physicians
India. 64:20–25. 2016.PubMed/NCBI
|
|
5
|
Nissen SE, Yeomans ND, Solomon DH, Lüscher
TF, Libby P, Husni ME, Graham DY, Borer JS, Wisniewski LM, Wolski
KE, et al: Cardiovascular safety of celecoxib, naproxen, or
ibuprofen for arthritis. N Engl J Med. 375:2519–2529. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shirai T, Nazarewicz RR, Wallis BB, Yanes
RE, Watanabe R, Hilhorst M, Tian L, Harrison DG, Giacomini JC,
Assimes TL, et al: The glycolytic enzyme PKM2 bridges metabolic and
inflammatory dysfunction in coronary artery disease. J Exp Med.
213:337–354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li H, Sun K, Zhao R, Hu J, Hao Z, Wang F,
Lu Y, Liu F and Zhang Y: Inflammatory biomarkers of coronary heart
disease. Front Biosci (Landmark Ed). 22:504–515. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li S, Fan Q, He S, Tang T, Liao Y and Xie
J: MicroRNA-21 negatively regulates treg cells through a
TGF-β1/Smad-independent pathway in patients with coronary heart
disease. Cell Physiol Biochem. 37:866–878. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thakur A, Schalk D, Tomaszewski E,
Kondadasula SV, Yano H, Sarkar FH and Lum LG: Microenvironment
generated during EGFR targeted killing of pancreatic tumor cells by
ATC inhibits myeloid-derived suppressor cells through COX2 and PGE2
dependent pathway. J Transl Med. 11:352013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schaan BD, Quadros AS, Sarmento-Leite R,
De Lucca G Jr, Bender A and Bertoluci M: ‘Correction:’ Serum
transforming growth factor beta-1 (TGF-beta-1) levels in diabetic
patients are not associated with pre-existent coronary artery
disease. Cardiovasc Diabetol. 6:192007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang J, Zeini M, Lin CY, Lin CJ, Xiong Y,
Shang C, Han P, Li W, Quertermous T, Zhou B and Chang CP:
Epicardial calcineurin-NFAT signals through Smad2 to direct
coronary smooth muscle cell and arterial wall development.
Cardiovasc Res. 101:120–129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hosseinzadeh H and Nassiri-Asl M: Review
of the protective effects of rutin on the metabolic function as an
important dietary flavonoid. J Endocrinol Invest. 37:783–788. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chuffa LG, Fioruci-Fontanelli BA, Bordon
JG, Pires RB, Braga CP, Seiva FR and Fernandes AA: Rutin
ameliorates glycemic index, lipid profile and enzymatic activities
in serum, heart and liver tissues of rats fed with a combination of
hypercaloric diet and chronic ethanol consumption. Indian J Biochem
Biophys. 51:215–222. 2014.PubMed/NCBI
|
|
14
|
Yu XL, Li YN, Zhang H, Su YJ, Zhou WW,
Zhang ZP, Wang SW, Xu PX, Wang YJ and Liu RT: Rutin inhibits
amylin-induced neurocytotoxicity and oxidative stress. Food Funct.
6:3296–3306. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Benavente-Garcia O and Castillo J: Update
on uses and properties of citrus flavonoids: New findings in
anticancer, cardiovascular and anti-inflammatory activity. J Agric
Food Chem. 56:6185–6205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han Y, Lu JS, Xu Y, Zhang L and Hong BF:
Rutin ameliorates renal fibrosis and proteinuria in
5/6-nephrectomized rats by anti-oxidation and inhibiting activation
of TGFβ1-smad signaling. Int J Clin Exp Pathol. 8:4725–4734.
2015.PubMed/NCBI
|
|
17
|
Ammenwerth E, Woess S, Baumgartner C, Fetz
B, van der Heidt A, Kastner P, Modre-Osprian R, Welte S and Poelzl
G: Evaluation of an integrated telemonitoring surveillance system
in patients with coronary heart disease. Methods Inf Med.
54:388–397. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Salfati E, Nandkeolyar S, Fortmann SP,
Sidney S, Hlatky MA, Quertermous T, Go AS, Iribarren C, Herrington
DM, Goldstein BA and Assimes TL: Susceptibility loci for clinical
coronary artery disease and subclinical coronary atherosclerosis
throughout the life-course. Circ Cardiovasc Genet. 8:803–811. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Jiang Y, Nguyen HD, Poo DC and
Wang W: The effect of a smartphone-based coronary heart disease
prevention (SBCHDP) programme on awareness and knowledge of CHD,
stress and cardiac-related lifestyle behaviours among the working
population in Singapore: A pilot randomised controlled trial.
Health Qual Life Outcomes. 15:492017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Riegersperger M, Covic A and Goldsmith D:
Allopurinol, uric acid and oxidative stress in cardiorenal disease.
Int Urol Nephrol. 43:441–449. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Prasad K and Dhar I: Oxidative stress as a
mechanism of added sugar-induced cardiovascular disease. Int J
Angiol. 23:217–226. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Turan T, Menteşe Ü, Ağaç MT, Akyüz AR, Kul
S, Aykan AÇ, Bektaş H, Korkmaz L, Öztaş Menteşe S, Dursun İ and
Çelik Ş: The relation between intensity and complexity of coronary
artery lesion and oxidative stress in patients with acute coronary
syndrome. Anatol J Cardiol. 15:795–800. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Wang M, Li Z, Bi X, Song J, Weng
S and Fu G: NADPH oxidase activation played a critical role in the
oxidative stress process in stable coronary artery disease. Am J
Transl Res. 8:5199–5210. 2016.PubMed/NCBI
|
|
24
|
Zurgil N, Solodeev I, Gilburd B, Shafran
Y, Afrimzon E, Avtalion R, Shoenfeld Y and Deutsch M: Monitoring
the apoptotic process induced by oxidized low-density lipoprotein
in Jurkat T-lymphoblast and U937 monocytic human cell lines. Cell
Biochem Biophys. 40:97–113. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pintér Ö, Hardi P, Nagy T, Gasz B, Kovács
V, Arató E, Sínay L, Lénárd L and Jancsó G: The role of GST
polymorphism in reperfusion induced oxidative stress, inflammatory
responses and clinical complications after surgical and
percutaneous coronary intervention. Clin Hemorheol Microcirc.
66:261–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Crobu F, Palumbo L, Franco E, Bergerone S,
Carturan S, Guarrera S, Frea S, Trevi G, Piazza A and Matullo G:
Role of TGF-beta1 haplotypes in the occurrence of myocardial
infarction in young Italian patients. BMC Med Genet. 9:132008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Purnomo Y, Piccart Y, Coenen T, Prihadi JS
and Lijnen PJ: Oxidative stress and transforming growth
factor-beta1-induced cardiac fibrosis. Cardiovasc Hematol Disord
Drug Targets. 13:165–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo L, Zhang Y, Zhang L, Huang F, Li J and
Wang S: MicroRNAs, TGF-β signaling and the inflammatory
microenvironment in cancer. Tumour Biol. 37:115–125. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shen Y, Zhang C and Chen Y: TGF-β in
inflammatory bowel diseases: A tale of the janus-like cytokine.
Crit Rev Eukaryot Gene Expr. 25:335–347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Valls N, Gormaz JG, Aguayo R, González J,
Brito R, Hasson D, Libuy M, Ramos C, Carrasco R, Prieto JC, et al:
Amelioration of persistent left ventricular function impairment
through increased plasma ascorbate levels following myocardial
infarction. Redox Rep. 21:75–83. 2016.PubMed/NCBI
|
|
31
|
Forbes K, Souquet B, Garside R, Aplin JD
and Westwood M: Transforming growth factor-{beta} (TGF{beta})
receptors I/II differentially regulate TGF{beta}1 and IGF-binding
protein-3 mitogenic effects in the human placenta. Endocrinology.
151:1723–1731. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Thu VT, Kim HK, le T Long, Thuy TT, Huy
NQ, Kim SH, Kim N, Ko KS, Rhee BD and Han J: NecroX-5 exerts
anti-inflammatory and anti-fibrotic effects via modulation of the
TNFα/Dcn/TGFβ1/Smad2 pathway in hypoxia/reoxygenation-treated rat
hearts. Korean J Physiol Pharmacol. 20:305–314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhong Y, Cheng CF, Luo YZ, Tian CW, Yang
H, Liu BR, Chen MS, Chen YF and Liu SM: C-reactive protein
stimulates RAGE expression in human coronary artery endothelial
cells in vitro via ROS generation and ERK/NF-κB activation. Acta
Pharmacol Sin. 36:440–447. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tarone G, Sbroggiò M and Brancaccio M: Key
role of ERK1/2 molecular scaffolds in heart pathology. Cell Mol
Life Sci. 70:4047–4054. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu J, Wang L, Akinyi M, Li Y, Duan Z, Zhu
Y and Fan G: Danshensu protects isolated heart against ischemia
reperfusion injury through activation of Akt/ERK1/2/Nrf2 signaling.
Int J Clin Exp Med. 8:14793–14804. 2015.PubMed/NCBI
|
|
36
|
Lee YH, Lee SJ, Jung JE, Kim JS, Lee NH
and Yi HK: Terrein reduces age-related inflammation induced by
oxidative stress through Nrf2/ERK1/2/HO-1 signalling in aged HDF
cells. Cell Biochem Funct. 33:479–486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hao Q, Chen X, Wang X, Dong B and Yang C:
Curcumin attenuates angiotensin ii-induced abdominal aortic
aneurysm by inhibition of inflammatory response and ERK signaling
pathways. Evid Based Complement Alternat Med. 2014:2709302014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pang X, Liu J, Li Y, Zhao J and Zhang X:
Emodin inhibits homocysteine-induced c-reactive protein generation
in vascular smooth muscle cells by regulating ppargamma expression
and ROS-ERK1/2/p38 signal pathway. PLoS One. 10:e01312952015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeong JJ, Ha YM, Jin YC, Lee EJ, Kim JS,
Kim HJ, Seo HG, Lee JH, Kang SS, Kim YS and Chang KC: Rutin from
Lonicera japonica inhibits myocardial ischemia/reperfusion-induced
apoptosis in vivo and protects H9c2 cells against hydrogen
peroxide-mediated injury via ERK1/2 and PI3K/Akt signals in vitro.
Food Chem Toxicol. 47:1569–1576. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hamamdzic D, Fenning RS, Patel D, Mohler
ER III, Orlova KA, Wright AC, Llano R, Keane MG, Shannon RP,
Birnbaum MJ and Wilensky RL: Akt pathway is hypoactivated by
synergistic actions of diabetes mellitus and hypercholesterolemia
resulting in advanced coronary artery disease. Am J Physiol Heart
Circ Physiol. 299:H699–H706. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moghbelinejad S, Nassiri-Asl M, Farivar
TN, Abbasi E, Sheikhi M, Taghiloo M, Farsad F, Samimi A and Hajiali
F: Rutin activates the MAPK pathway and BDNF gene expression on
beta-amyloid induced neurotoxicity in rats. Toxicol Lett.
224:108–113. 2014. View Article : Google Scholar : PubMed/NCBI
|