Introduction
Paraquat (PQ; 1,1′-dimethyl-4,4′-bipyridinium) is a
herbicide that is widely used in developing countries worldwide,
which may cause severe toxicity in animals and humans (1). PQ poisoning was reported to account for
up to a third of all suicides around the world (2). PQ poisoning may cause patients to
develop acute multi-organ failure, pulmonary fibrosis and finally
mortality due to respiratory failure (3). Pulmonary fibrosis is a typical feature
of PQ poisoning, the onset of which may be several days or weeks
following ingestion of PQ (4).
Although PQ-induced pulmonary fibrosis has a high mortality rate,
the molecular mechanisms underlying its toxicity are largely
unknown, which makes it particularly hard to treat.
Epithelial-mesenchymal transition (EMT) has been
reported to be associated with pulmonary fibrosis following PQ
exposure (3). It has also been
reported that factors, including epithelial growth factor,
transforming growth factor-β1 (TGF-β1), insulin-like growth factor
and interleukin-17, may induce EMT (5). Myofibroblasts are key mediators in
fibrosis. In murine models of hepatic and renal fibrosis, ~40% of
α-smooth muscle actin (SMA)-positive myofibroblasts were derived
from epithelial cells via EMT (6).
However, to the best of our knowledge, the roles and mechanisms of
rapamycin in the EMT process of A549 and MRC-5 cells remain
unknown.
In the present study, the roles and mechanisms of
rapamycin on PQ-induced pulmonary fibrosis were investigated. It
was revealed that rapamycin alleviated PQ-induced EMT in A549 and
MRC-5 cells, and PQ activated the Wnt signaling pathway. Rapamycin
inhibited the effects of PQ, and the activation of the Wnt
signaling pathway attenuated the inhibitory effects of rapamycin on
PQ-induced EMT.
Materials and methods
Reagents
PQ, rapamycin and lithium chloride (LiCl) were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Antibodies against E-cadherin (ab1416), β-actin (ab8227) and α-SMA
(ab7817) were purchased from Abcam (Cambridge, UK). Antibodies
against Wnt1, low-density lipoprotein receptor-related protein
(LRP)5 and LRP6 were provided by Santa Cruz Biotechnology, Inc.,
(Dallas, TX, USA). Antibodies against β-catenin were purchased from
Cell Signaling Technology, Inc., (Danvers, MA, USA).
Cell culture and treatment
A549 (human lung adenocarcinoma epithelial cells)
and MRC-5 (human fetal lung fibroblast cells) cells were purchased
from American Type Culture Collection (Manassas, VA, USA) and
cultured in Dulbecco's modified Eagle medium (DMEM) (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin and 100 U/ml streptomycin at 37°C in a humidified
5% CO2 atmosphere.
A previous study demonstrated that 300 µmol/l PQ
treatment for 6 days induced the EMT of A549 cells; therefore, this
concentration was selected for use within the present study
(3). Prior to treatment, A549 cells
and MRC-5 cells were incubated in serum-free DMEM for 12 h at 37°C.
To evaluate the effects of Wnt signaling on EMT, 300 µmol/l PQ, 100
nM rapamycin and 10 µmol/l LiCl were added to the cells for 48 h.
All the reagents were added again when the medium was changed every
day.
In the present study, four treatment groups were
established to study the effects of rapamycin on the PQ-induced EMT
and the Wnt signaling pathway in A549 and MRC-5 cells: The negative
control group, the PQ-treated group, the rapamycin-treated group
and the PQ + rapamycin-treated group. Subsequently, five groups
were established to confirm the roles of Wnt signaling pathway in
the inhibitory effects of rapamycin on PQ-induced EMT in A549 and
MRC-5 cells: The negative control group, the PQ-treated group, the
LiCl-treated group, the PQ + rapamycin-treated group and the LiCl +
PQ + rapamycin-treated group.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from A549 and MRC-5cells
using a total RNA rapid extraction kit (BioTeke Corporation,
Beijing, China) according to manufacturer's protocol. A total of 1
µg RNA was reverse transcribed into cDNA using M-MLV reverse
transcriptase (BioTeke Corporation) in the presence of oligo (dT)
and random 50 primers (Invitrogen; Thermo Fisher Scientific, Inc.).
The reaction program was 42°C for 50 min, and then 72°C for 10 min.
The instruments in this section were pre-treated with surface RNase
Erase (Tiandz, Inc., Beijing, China) and the reagents were
RNase-free. The cDNA (1 µl for each reaction) was used for qPCR to
detect the gene expression levels using 2XPower Taq PCR Master mix
(BioTeke Corporation) and SYBR Green (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China) with GAPDH as the
internal control. The PCR procedure was set as follows: 95°C for 10
min, followed by 38 cycles of 95°C for 12 secs 60°C for 18 sec and
72°C for 30 sec, and finally 4°C for 5 min. Calculations were
performed using the 2−ΔΔCq method described previously
(7). The primers used were are as
follows: GAPDH, forward 5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse
5′-GGCTGTTGTCATACTTCTCATGG-3′; E-cadherin, forward
5′-AAGGCACGCCTGTCGAAGCA-3′ and reverse
5′-ACGTTGTCCCGGGTGTCATCCT-3′; α-SMA, forward
5′-TACTACTGCTGAGCGTGAGA-3′ and reverse 5′-CATCAGGCAACTCGTAACTC-3′;
Wnt1, forward 5′-CCGATGGTGGGGTATTGTGAA-3′ and reverse
5′-TCCCCGGATTTTGGCGTATC-3′; β-catenin, forward
5′-GCCAGTGGATTCCGTACTGT-3′ and reverse 5′-GAGCTTGCTTTCCTGATTGC-3′;
LRP5, forward 5′-GGGAGACGCCAAGACAGACAAGATCG-3′ and reverse
5′-GGTGAAGACCAAGAAGGCCTCAGG-3′; and LRP6, forward
5′-ATTGTAGTTGGAGGCTTGGAGGATGC-3′ and reverse
5′-CCATCCATTCCAGCACGTTCTATC-3′.
Western blot analysis
Protein was extracted from A549 and MRC-5cells using
a whole-cell lysis kit (CWBio, Beijing, China, http://www.cwbiotech.com) and the concentration of
protein was measured using a bichinchoninic acid protein
quantitative kit (Beyotime Institute of Biotechnology, Haimen,
China). Following denaturation by boiling for 5 min, the protein
samples (40 µg for each lane) were separated by 10% SDS-PAGE and
transferred to polyvinylidene fluoride membranes (Merck KGaA).
Following blocking with 5% skimmed milk at room temperature for 1
h, the membranes were incubated with the primary antibodies
anti-E-cadherin (1:1,000), anti-α-SMA (1:1,000), anti-Wnt1
(1:2,000), anti-LRP5 (1:2,000), anti-LRP6 (1:1,000), anti-β-catenin
(1:1,000) and anti-β-actin (1:5,000)at 4°C overnight. Following
rinsing with Tris-buffered saline with Tween-20, the membranes were
incubated, with goat anti-rabbit immunoglobulin (Ig) G labeled with
horseradish peroxidase (HRP;sc-2007; dilution 1:5,000) or goat
anti-mouse IgG-HRP (sc-2005; dilution 1:5,000; both Santa Cruz
Biotechnology, Inc.) at 37°C for 45 min. The membranes were
visualized using an enhanced chemiluminescent reagent (Thermo
Fisher Scientific, Inc.). The optical density values of bands were
analyzed using Image Lab software v.3.0 (Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Fluorescent immunostaining
A549 cells were grown on cover slips at 37°C in DMEM
cell culture medium, and then fixed with 4% paraformaldehyde for 10
min at room temperature, and blocked with 1% bovine serum albumin
(Thermo Fisher Scientific, Inc.) in PBS for 1 h at room
temperature, and then stained with primary antibodies against
E-cadherin (dilution 1:500) overnight at 4°C. The cells were
subsequently incubated with goat anti-mouse secondary antibody
(dilution 1:2,000, ab150115, Abcam) for 1 h at room temperature.
The nuclei were stained with 4′,6-diamidino-2-phenylindole for 1 h
at room temperature. The fluorescence images were observed under a
fluorescence microscope at a magnification of ×400.
Statistical analysis
The data in the present study were presented as the
mean ± standard deviation of three or five individual experiments.
The results were analyzed by one-way analysis of variance with post
hoc comparisons with Tukey's honest significant difference test
using GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
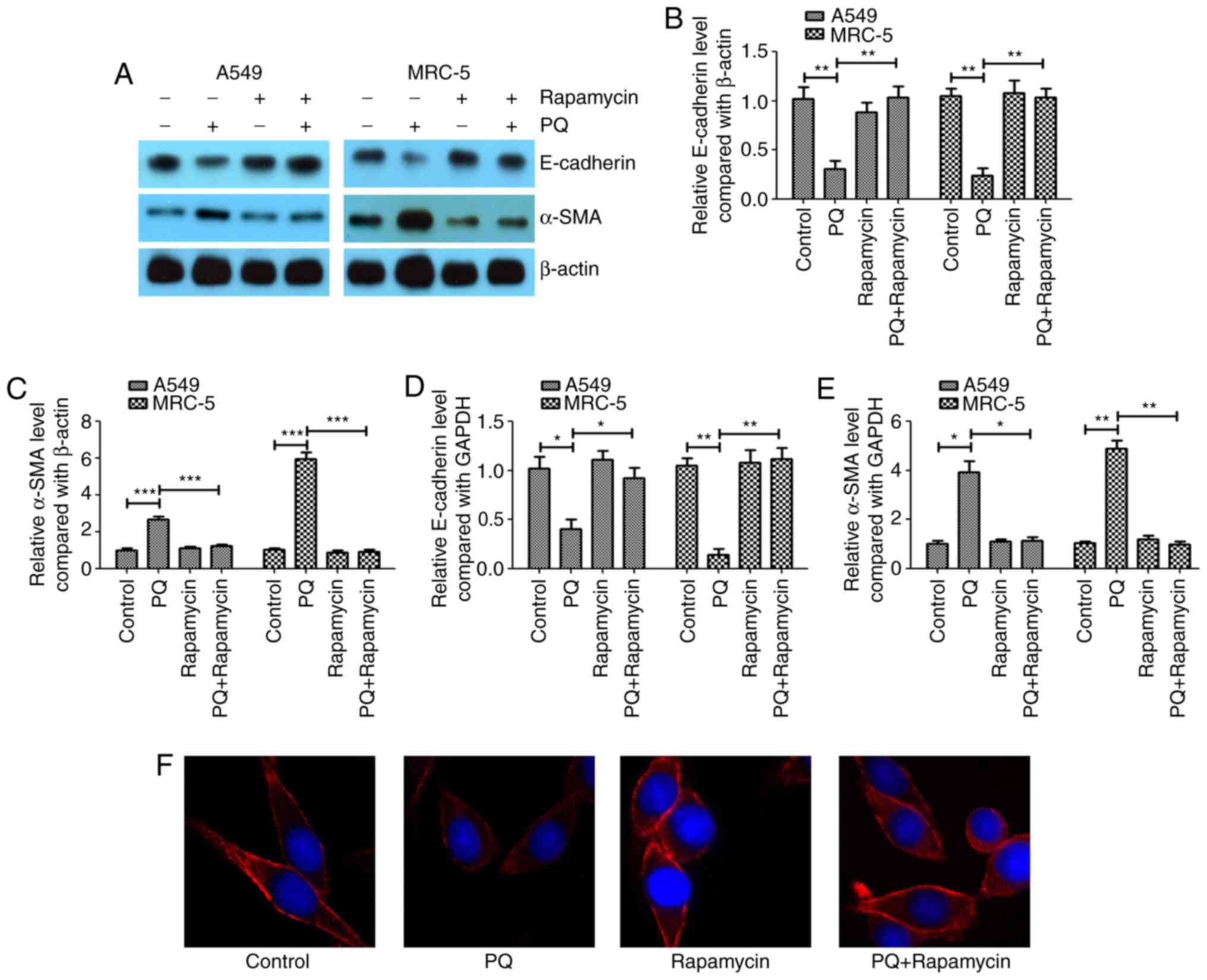
Rapamycin alleviates PQ-induced EMT in
A549 and MRC-5 cells
In the present study, A549 and MRC-5 cells were
exposed to 300 µmol/l PQ for 6 days as previously described
(3). A western blotting assay
revealed that PQ induced EMT in A549 and MRC-5 cells (Fig. 1A), as the epithelial cell marker
E-cadherin was significantly downregulated in the PQ group compared
with the level in the control (Fig.
1B). Furthermore, the mesenchymal cell marker α-SMA was
significantly upregulated in the PQ group compared with the level
in the control (Fig. 1C). It was
observed that rapamycin may inhibit the EMT process that was
induced by PQ as E-cadherin was significantly increased and α-SMA
was significantly decreased in the PQ + rapamycin group compared
with the level in the PQ group.
The mRNA expression levels of E-cadherin and α-SMA
were investigated using RT-qPCR, and it was observed that PQ
significantly decreased the mRNA expression level of E-cadherin
(Fig. 1D) and significantly
increased the mRNA level of α-SMA (Fig.
1E) compared with the levels observed in the control group.
Rapamycin significantly inhibited the effect of PQ on α-SMA and
E-cadherin in A549 and MRC-5 cells. Using immunofluorescent
staining, it was further demonstrated that rapamycin notably
upregulated the expression of E-cadherin compared with the level in
the PQ group in A549 cells (Fig.
1F).
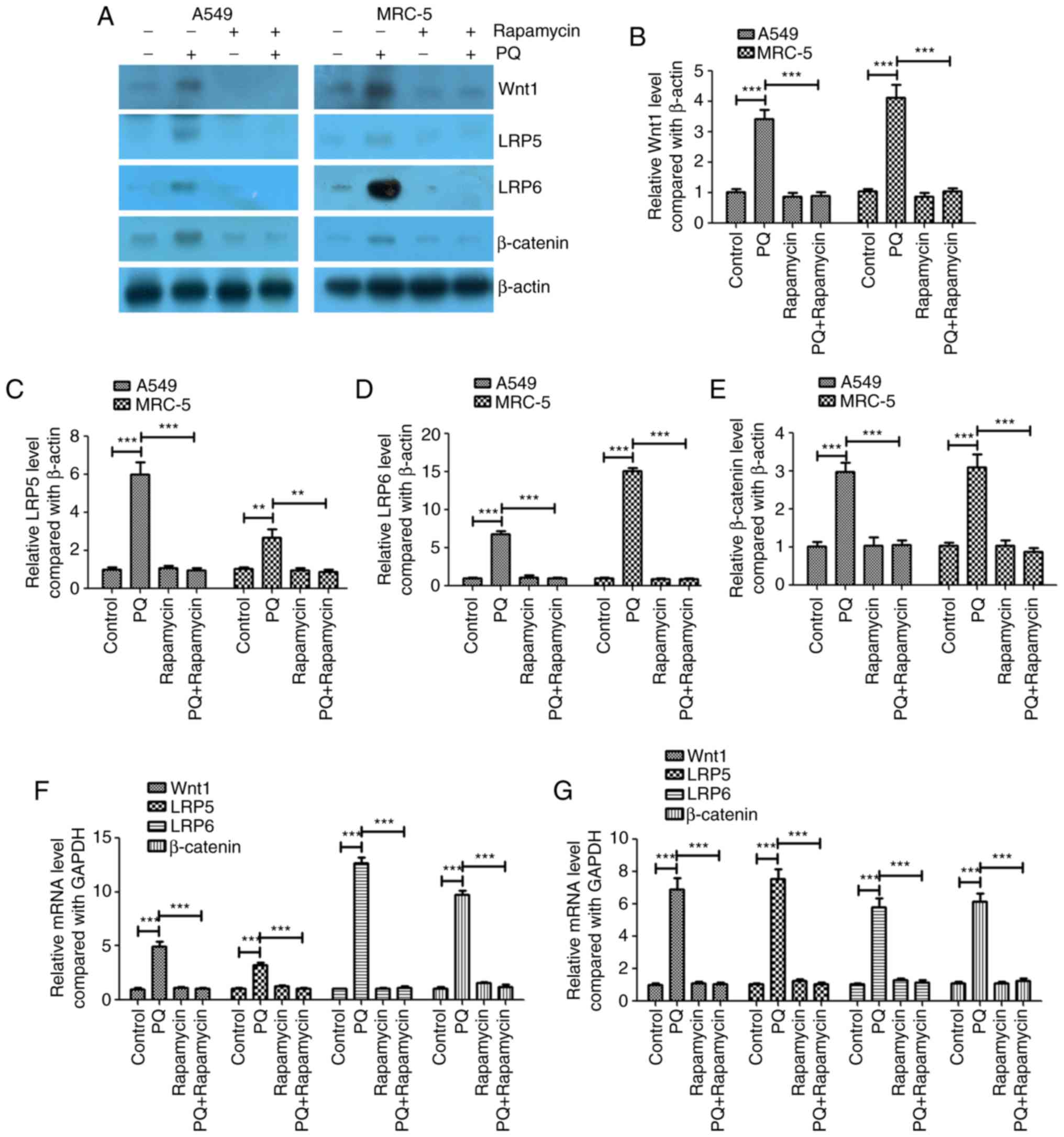
PQ activates the Wnt signaling pathway
in A549 and MRC-5 cells, and rapamycin inhibits these effects of
PQ
The present study investigated whether the Wnt
signaling pathway served an important role in the PQ-induced EMT
process. The protein and mRNA expression levels of Wnt1, LRP5 and
LRP6 were investigated and it was revealed that PQ significantly
increased the protein and mRNA expression levels of Wnt1, LRP5 and
LRP6 compared with the levels in the control group (Fig. 2). Rapamycin significantly decreased
the protein and mRNA expression levels of Wnt1, LRP5 and LRP6
compared with the levels in the PQ group in A549 and MRC-5 cells
(Fig. 2). PQ also significantly
upregulated the key regulator of the Wnt signaling pathway,
β-catenin, at the mRNA and protein levels, and rapamycin
significantly decreased β-catenin expression compared with the
level observed in the PQ group in A549 and MRC-5 cells (Fig. 2). These results indicate that PQ
activated the Wnt signaling pathway in A549 and MRC-5 cells and
rapamycin inhibited the effects of PQ.
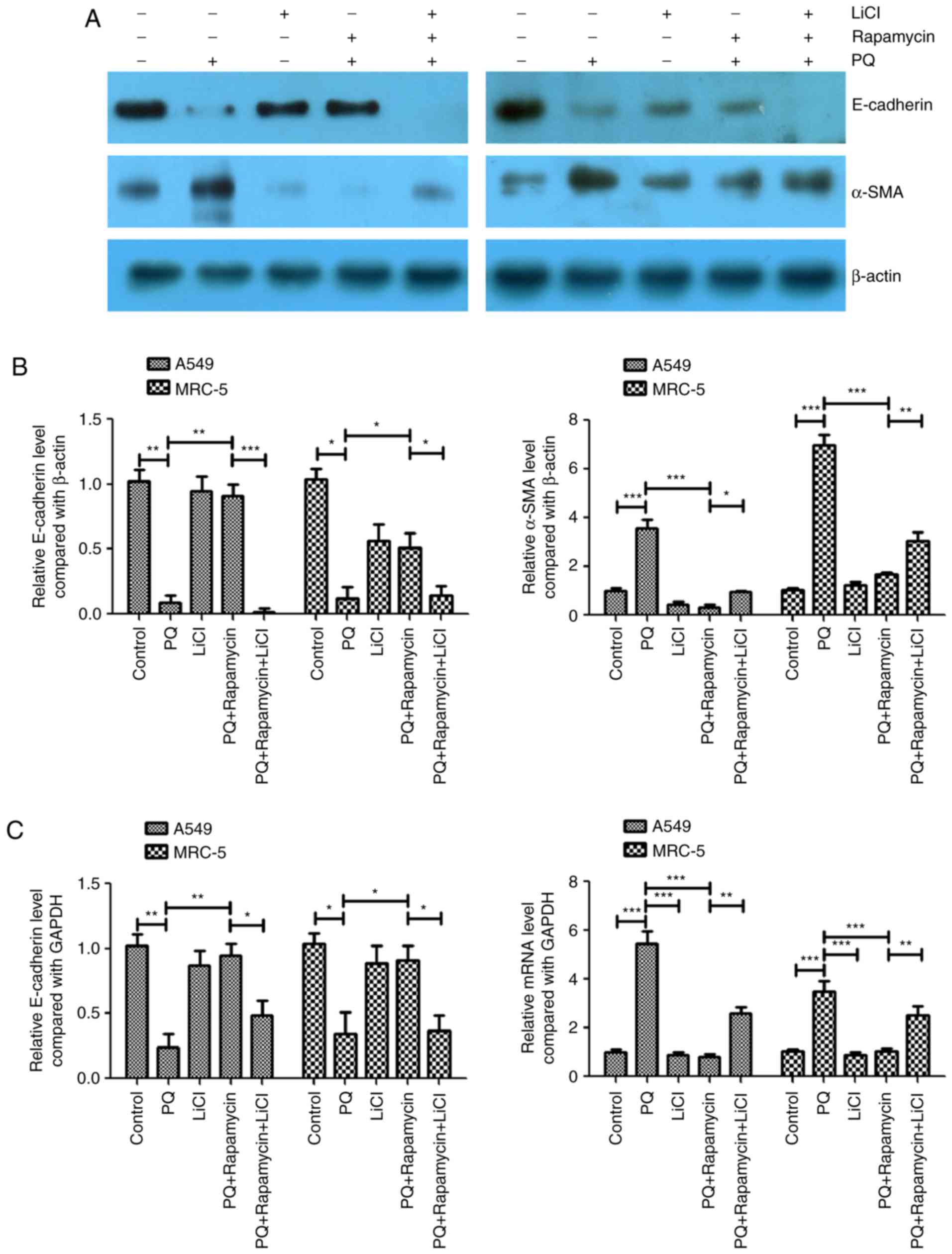
Activation of the Wnt signaling
pathway attenuates the inhibitory effects of rapamycin on
PQ-induced EMT
LiCl is an activator of the Wnt/catenin signaling
pathway (6–9) and this activator was used to further
study the association between the Wnt signaling pathway and
PQ-induced EMT in the present study. In the PQ-treated A549 and
MRC-5 cells, rapamycin inhibited PQ-induced EMT by significantly
upregulating E-cadherin and significantly downregulating α-SMA
protein levels compared with the levels in the PQ only group
(Fig. 3A-C). However, LiCl
significantly inhibited the effects of rapamycin as it caused a
significant decrease in the protein expression level of E-cadherin
and a significant increase in the protein expression of α-SMA
compared with the levels observed in the PQ+ rapamycin group
(Fig. 3A and B). The mRNA expression
levels of E-cadherin and α-SMA were measured using RT-qPCR and
similar results were observed. Rapamycin significantly increased
the mRNA expression level of E-cadherin and significantly decreased
the level of α-SMA compared with the levels in the PQ only group,
while LiCl significantly inhibited the effects of rapamycin
compared with the PQ + rapamycin group (Fig. 3B and C).
Discussion
Pulmonary fibrosis is a chronic lung disease, which
causes fibrosis of the lung parenchyma and loss of lung function
(10). Aging, smoking and infection
have been reported as risk factors for pulmonary fibrosis (11). The pathological characteristics of
pulmonary fibrosis are repetitive microscopic alveolar epithelial
cell injury, dysregulated repair, fibroblast proliferation and
accumulation of extracellular matrix, which ultimately result in
respiratory failure (12). EMT is an
important mechanism of lung fibrogenesis through the generation of
mesenchymal-type myofibroblasts from lung epithelial cells
(13). EMT is also considered to be
the key process that leads to end-stage lung fibrosis, which occurs
in a number of different types of inflammatory interstitial lung
diseases and chronic obstructive pulmonary disease (5).
PQ may induce EMT-like cellular responses resulting
in fibrogenesis and the prevention of apoptosis in human pulmonary
epithelial cells (14). Previous
studies reported that PQ promoted the EMT process by activating the
TGF-β/Smad signaling pathway (15,16).
Lysyl oxidase may promote EMT during PQ-induced pulmonary fibrosis
(17). In PQ poisoning-induced early
pulmonary fibrosis, hypoxia-inducible factor-1α enhanced Snail and
β-catenin and promoted EMT (18).
The results of the present study demonstrated that PQ significantly
upregulated the expression levels of Wnt1, LRP5, LRP6 and
β-catenin. Rapamycin was revealed to inhibit these effects of PQ on
the Wnt signaling pathway genes. When the Wnt signaling pathway was
activated using LiCl, EMT was promoted. These findings suggest that
rapamycin protects against PQ-induced pulmonary EMT by suppressing
the Wnt/β-catenin signaling pathway.
Yang et al (19) revealed that silencing the mechanistic
target of rapamycin (mTOR) using small interfering RNA effectively
inhibited the expression level of mTOR in the lung tissues of
PQ-poisoned rats and further decreased the fibrosis of lung tissues
caused by PQ. Rapamycin is an inhibitor of mTOR and may activate
autophagy (20,21). The present study revealed that
rapamycin significantly inhibited PQ-induced EMT and the Wnt
signaling pathway. However, whether mTOR is associated with the
regulation of PQ-induced EMT requires further study to confirm.
Using a murine model, Chung et al (22) demonstrated that rapamycin treatment
reduced inflammatory cytokine expression, extracellular matrix
production and senescence in type II pneumocytes, and that
rapamycin protected against radiation-induced pulmonary fibrosis.
Phosphoinositide 3-kinase (PI3K) was reported as a promising
therapeutic target for idiopathic pulmonary fibrosis (23). Rapamycin may increase connective
tissue growth factor expression in lung fibroblasts by regulating
the PI3K signaling pathway (24).
However, the association between the Wnt and PI3K/mitogen-activated
protein kinase signaling pathways requires further
investigation.
Snail and Twist are important transcription factors
of EMT associated with pulmonary fibrosis (25,26). In
further studies, which transcription factor is involved in the
process of PQ-induced EMT should be studied.
In conclusion, the results of the present study
suggested that rapamycin alleviated PQ-induced EMT in A549 and
MRC-5 cells. It was also observed that PQ activated the Wnt
signaling pathway in A549 and MRC-5 cells and that rapamycin
inhibited these effects of PQ. Activation of the Wnt signaling
pathway attenuated the inhibitory effects of rapamycin on
PQ-induced EMT. Further investigation into the effects of PQ and
rapamycin on EMT are required, particularly on the expression of
additional EMT-associated proteins, including zonula occludens-1,
cytokeratin, vimentin and N-cadherin. The role of autophagy in
PQ-induced pulmonary fibrosis should also be investigated.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81560015 and
81360015), Yunnan Applied Basic Research Projects (grant no.
2013FB049) and Yunnan Applied Basic Research Projects-Joint Special
Project (established by Yunnan Provincial Science and Technology
Department and Kunming Medical University) [grant nos. 2014FB046
and 2017FE468 (−005)].
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Weng CH, Chen HH, Hu CC, Huang WH, Hsu CW,
Fu JF, Lin WR, Wang IK and Yen TH: Predictors of acute kidney
injury after paraquat intoxication. Oncotarget. 8:51345–51354.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sabzghabaee AM, Eizadi-Mood N, Montazeri
K, Yaraghi A and Golabi M: Fatality in paraquat poisoning.
Singapore Med J. 51:496–500. 2010.PubMed/NCBI
|
|
3
|
Li T, Yang X, Xin S, Cao Y and Wang N:
Paraquat poisoning induced pulmonary epithelial mesenchymal
transition through Notch1 pathway. Sci Rep. 7:9242017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suntres ZE: Role of antioxidants in
paraquat toxicity. Toxicology. 180:65–77. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Willis BC and Borok Z: TGF-beta-induced
EMT: Mechanisms and implications for fibrotic lung disease. Am J
Physiol Lung Cell Mol Physiol. 293:L525–L534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li H, Hao Y, Zhang H, Ying W, Li D, Ge Y,
Ying B, Cheng B, Lian Q and Jin S: Posttreatment with Protectin DX
ameliorates bleomycin-induced pulmonary fibrosis and lung
dysfunction in mice. Sci Rep. 7:467542017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xia MY, Zhao XY, Huang QL, Sun HY, Sun C,
Yuan J, He C, Sun Y, Huang X, Kong W and Kong WJ: Activation of
Wnt/β-catenin signaling by lithium chloride attenuates
d-galactose-induced neurodegeneration in the auditory cortex of a
rat model of aging. FEBS Open Bio. 7:759–776. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang X, Tang S, Dai C, Li D, Zhang S, Deng
S, Zhou Y and Xiao X: Quinocetone induces mitochondrial apoptosis
in HepG2 cells through ROS-dependent promotion of VDAC1
oligomerization and suppression of Wnt1/β-catenin signaling
pathway. Food Chem Toxicol. 105:161–176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lucarini L, Durante M, Lanzi C, Pini A,
Boccalini G, Calosi L, Moroni F, Masini E and Mannaioni G:
HYDAMTIQ, a selective PARP-1 inhibitor, improves bleomycin-induced
lung fibrosis by dampening the TGF-β/SMAD signalling pathway. J
Cell Mol Med. 21:324–335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wuyts WA, Agostini C, Antoniou KM, Bouros
D, Chambers RC, Cottin V, Egan JJ, Lambrecht BN, Lories R, Parfrey
H, et al: The pathogenesis of pulmonary fibrosis: A moving target.
Eur Respir J. 41:1207–1218. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Borensztajn K, Crestani B and Kolb M:
Idiopathic pulmonary fibrosis: From epithelial injury to
biomarkers-insights from the bench side. Respiration. 86:441–452.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim KK, Kugler MC, Wolters PJ, Robillard
L, Galvez MG, Brumwell AN, Sheppard D and Chapman HA: Alveolar
epithelial cell mesenchymal transition develops in vivo during
pulmonary fibrosis and is regulated by the extracellular matrix.
Proc Natl Acad Sci USA. 103:pp. 13180–13185. 2006; View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamada A, Aki T, Unuma K, Funakoshi T and
Uemura K: Paraquat induces epithelial-mesenchymal transition-like
cellular response resulting in fibrogenesis and the prevention of
apoptosis in human pulmonary epithelial cells. PLoS One.
10:e01201922015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han YY, Shen P and Chang WX: Involvement
of epithelial-to-mesenchymal transition and associated transforming
growth factor-β/Smad signaling in paraquat-induced pulmonary
fibrosis. Mol Med Rep. 12:7979–7984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xie L, Zhou D, Xiong J, You J, Zeng Y and
Peng L: Paraquat induce pulmonary epithelial-mesenchymal transition
through transforming growth factor-β1-dependent mechanism. Exp
Toxicol Pathol. 68:69–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Zhu Y, Tan J, Meng X, Xie H and
Wang R: Lysyl oxidase promotes epithelial-to-mesenchymal transition
during paraquat-induced pulmonary fibrosis. Mol Biosyst.
12:499–507. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu Y, Tan J, Xie H, Wang J, Meng X and
Wang R: HIF-1α regulates EMT via the Snail and β-catenin pathways
in paraquat poisoning-induced early pulmonary fibrosis. J Cell Mol
Med. 20:688–697. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang W, Zhao X, Liang R and Chen D:
Effects of small RNA interference targeting mammalian target of
rapamycin on paraquat-induced pulmonary fibrosis in rats. Zhonghua
Wei Zhong Bing Ji Jiu Yi Xue. 29:830–835. 2017.(In Chinese).
PubMed/NCBI
|
|
20
|
Chen LM, Song TJ, Xiao JH, Huang ZH, Li Y
and Lin TY: Tripchlorolide induces autophagy in lung cancer cells
by inhibiting the PI3K/AKT/mTOR pathway and improves cisplatin
sensitivity in A549/DDP cells. Oncotarget. 8:63911–63922.
2017.PubMed/NCBI
|
|
21
|
Song Y, Zhang P, Sun Y, Li X, Chen L, Xiao
Y and Xing Y: AMPK activation-dependent autophagy compromises
oleanolic acid-induced cytotoxicity in human bladder cancer cells.
Oncotarget. 8:67942–67954. 2017.PubMed/NCBI
|
|
22
|
Chung EJ, Sowers A, Thetford A,
McKay-Corkum G, Chung SI, Mitchell JB and Citrin DE: Mammalian
target of rapamycin inhibition with rapamycin mitigates
radiation-induced pulmonary fibrosis in a murine model. Int J
Radiat Oncol Biol Phys. 96:857–866. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mercer PF, Woodcock HV, Eley JD, Platé M,
Sulikowski MG, Durrenberger PF, Franklin L, Nanthakumar CB, Man Y,
Genovese F, et al: Exploration of a potent PI3 kinase/mTOR
inhibitor as a novel anti-fibrotic agent in IPF. Thorax.
71:701–711. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu X, Dai H, Geng J, Wan X, Huang X, Li F,
Jiang D and Wang C: Rapamycin increases CCN2 expression of lung
fibroblasts via phosphoinositide 3-kinase. Lab Invest. 95:846–859.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu Y, Tai W, Qu X, Wu W, Li Z, Deng S,
Vongphouttha C and Dong Z: Rapamycin protects against
paraquat-induced pulmonary fibrosis: Activation of Nrf2 signaling
pathway. Biochem Biophys Res Commun. 490:535–540. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pozharskaya V, Torres-Gonzalez E, Rojas M,
Gal A, Amin M, Dollard S, Roman J, Stecenko AA and Mora AL: Twist:
A regulator of epithelial-mesenchymal transition in lung fibrosis.
PLoS One. 4:e75592009. View Article : Google Scholar : PubMed/NCBI
|