Introduction
Antigen-specific cancer immunotherapy using the
induction of tumor-specific reactions without autoimmunity is a
potentially attractive option for the treatment of cancer. However,
immunotherapy for hepatocellular carcinoma (HCC) is still in the
preclinical or early clinical trial phases (I and II) of
development (1,2). Glypican-3 (GPC3), a carcinoembryonic
antigen, is overexpressed in 72–81% of HCC cases, and is correlated
with poor prognosis; therefore, it is an ideal target for HCC
(3–7). Recently, a phase I clinical study of
a GPC3-derived peptide vaccine reported its safety and efficacy for
the treatment of advanced HCC (8).
Although vaccine-induced GPC3-peptide-specific cytotoxic T
lymphocytes (CTLs) are often tumor reactive in vitro
(9) and correlate with overall
survival, no complete response was observed when GPC3 peptide
vaccination was used as monotherapy in patients with advanced HCC
(8).
Programmed death-1 (PD-1) is expressed on activated
T and B cells, and elicits inhibitory signals (10). Its ligand PD-L1 is member of the B7
family, and interacts with PD-1 (11). Several studies have shown that the
PD-1/PD-L1 pathway plays a critical role in compromised tumor
immunity (12,13). PD-1 antibody blockade exerts
antitumor effects in clinical trials (14,15).
High expression levels of PD-1 on T cells, both in
tumor-infiltrating lymphocytes (TILs) and peripheral blood
mononuclear cells (PBMCs), were correlated with poor prognosis in
HCC patients after surgical resection (16). In addition, PD-L1 expression in HCC
was correlated with tumor aggressiveness and postoperative
recurrence (17).
In animal models, PD-1 blockade exerts synergistic
effects with various tumor vaccines to enhance tumor
antigen-specific T cell responses and suppress tumors in
vivo (18–20). It was reported that melanoma
vaccine-induced CTLs become exhausted, which could be reversed by
blocking the inhibitory pathways (21). However, a study evaluating the
combination of a cancer vaccine and an anti-PD-1 blocking antibody
(αPD-1 Ab) for HCC has not been conducted. Therefore, the aim of
this study was to investigate whether αPD-1 Ab would enhance the
antitumor effects of a peptide vaccine by analyzing CTLs isolated
from the PBMCs of vaccinated patients, as well as from a mouse
model.
Materials and methods
Patient samples
Three clinical trials were conducted using
GPC3-derived peptide vaccines. A phase I trial (n=33) was performed
in patients with advanced or metastatic HCC (8) (University Hospital Medical
Information Network Clinical Trials Registry; UMIN-CTR no.
000001395). Subsequently, a phase II trial was performed using a
GPC3-derived peptide vaccine as an adjuvant therapy in patients
with HCC (UMIN-CTR: 000002614, on-going). Finally, a pilot study of
liver biopsies taken before and after GPC3 peptide vaccination is
being performed for advanced HCC (UMIN-CTR: 000005093, on-going).
These trials were approved by the Ethics Committee of the National
Cancer Center, Japan, and conformed to the ethical guidelines of
the 1975 Declaration of Helsinki. All patients were enrolled after
providing written informed consent. Patients were injected
intradermally with HLA-A24-restricted GPC3298–306
(EYILSLEEL) or HLA-A2-restricted GPC3144–152 (FVGEFFTDV)
peptide vaccines emulsified with incomplete Freund’s adjuvant (IFA,
Montanide ISA-51VG; SEPPIC).
Peripheral blood (30 ml) was obtained at the
National Cancer Center Hospital East. PBMCs were isolated using
standard Ficoll density gradient centrifugation from buffy coats.
The remaining PBMCs were used after immunological monitoring in
clinical trials. The immunological analyses were approved by the
Ethics Committee of the National Cancer Center, Japan.
Cell lines
The human liver cancer cell lines SK-Hep-1
(GPC3−, HLA-A*02:01/A*24:02),
SK-Hep-1/GPC3 (GPC3+,
HLA-A*02:01/A*24:02), and HepG2
(GPC3+, HLA-A*02:01/A*24:02) were
available in our laboratory and were used as the target cells
(6,9). SK-Hep-1/GPC3 is an established stable
GPC3-expressing cell line that was transfected with the human GPC3
gene, whereas SK-Hep-1/vec is an established counterpart cell line
that was transfected with an empty vector. The mouse lymphoma cell
line RMA (OVA-, H-2Kb) was provided by Dr Yasuharu
Nishimura (Kumamoto University, Japan). Cells were cultured at 37°C
in RPMI-1640 or DMEM (Sigma-Aldrich) supplemented with 10% fetal
bovine serum (FBS), 100 U/ml penicillin and 100 μg/ml streptomycin
in a humidified atmosphere containing 5% CO2.
Synthetic peptides and cytokines
The peptides used in this study were as follows:
HLA-A*02:01-restricted GPC3144–152
(FVGEFFTDV) peptide (American Peptide Co.), HLA-A*24:
02-restricted GPC3298–306 (EYILSLEEL) peptide (American
Peptide Co.), HLA-A*02:01-restricted human
immunodeficiency virus (HIV)77–85 (SLYNTYATL) peptide
(ProImmune), and H-2Kb-restricted ovalbumin
(OVA)257–264 (SIINFEKL) peptide (AnaSpec). The peptides
were dissolved and diluted in 7% NaHCO3 or dimethyl
sulfoxide (DMSO). Where appropriate, liver cancer cell cultures
were treated with 100 U/ml recombinant interferon (IFN)-γ
(PeproTech).
Ex vivo Dextramer staining and flow
cytometry
PBMCs were stained using HLA-A*02:01
Dextramer-RPE [GPC3144–152 (FVGEFFTDV),
HIV19–27 (TLNAWVKVV) or negative control; Immudex] and
HLA-A*24:02 Dextramer-RPE [GPC3298–306
(EYILSLEEL), HIV583–591 (RYLKDQQLL); Immudex] for 15 min
at room temperature, followed by anti-CD8-FITC (clone T8, Beckman
Coulter), anti-PD-1-APC (clone EH12.2H7, BioLegend), or isotype
control-APC (clone MOPC-21, BioLegend) for 20 min at 4°C. Flow
cytometry was performed using a FACSCanto II (BD Biosciences).
Blocking antibody
GPC3 peptide-specific CTL clones were established
from PBMCs as described previously (9). The CTL clones were cultured in AIM-V
medium (Life Technologies) supplemented with 10% human AB serum in
the presence of 10 μg/ml anti-PD-1 (clone J116, eBioscience) or 10
μg/ml control (clone MOPC-21, BioXcell) monoclonal antibodies for 2
days.
CD107a assay
GPC3 peptide-specific CTL clones were incubated with
SK Hep-1/vec pulsed with GPC3144–152 or
HIV19–27 peptide and HepG2 at a 1:1 ratio for 3.5 h at
37°C. CTL clones were stained with anti-CD107a-APC (clone LAMP-1,
BD Bioscience) during the incubation period, followed by
anti-CD8-FITC (clone LT8, ProImmune) for 20 min at 4°C.
Mice
Female C57BL/6 mice (6–8 weeks old) were purchased
from Japan Charles River Laboratories (Yokohama, Japan), and were
maintained under specific pathogen-free conditions. The Animal
Research Committee of the National Cancer Center, Japan, approved
all studies. All animal procedures were performed according to the
guidelines for the Animal Research Committee of the National Cancer
Center, Japan. Ether was used for mouse euthanasia and
anesthesia.
In vivo tumor growth inhibition
assays
It was reported previously that intratumoral (i.t.)
injection of OVA257–264 peptide (SIINFEKL) effectively
inhibited OVA-negative tumor growth and survival in a peptide
vaccine model using C57BL/6 mice (22). RMA cells (1×105
cells/100 μl PBS) were implanted on the backs of C57BL/6 mouse on
day 0. They were then injected with 50-μg peptide mixed with an
equal volume of incomplete Freund’s adjuvant (IFA, Montanide
ISA-51VG; SEPPIC) on days 7 and 14. The total volume of injected
vaccine solution was 100 μl in all experiments. For in vivo
therapeutic experiments, anti-mouse PD-1 (clone 4H2) and control Ab
(clone MOPC-21, BioXcell) were provided by Ono Pharmaceutical Co.,
Ltd. The anti-mouse PD-1 Ab (clone 4H2) used in the present study
is a chimeric rat Ab containing the murine IgG1 Fc region (23). Anti-PD-1 or control Abs (200
μg/day) were injected intraperitoneally (i.p.) on days 7 and 14.
Tumor volume was monitored twice per week, and was calculated using
the following formula: tumor volume (mm3) = a × b × b ×
0.5, where a is the longest diameter, b is the shortest diameter,
and 0.5 is a constant to calculate the volume of an ellipsoid.
Mouse health, behavior and mortality were checked daily. All mice
were maintained until they showed signs of morbidity or the length
or width of the tumors exceeded 30 mm, at which point they were
sacrificed for reasons of animal welfare (22).
IFN-γ enzyme-linked immunospot (ELISPOT)
analysis
The BDTM ELISPOT set (BD Biosciences) was
used to assess the levels of IFN-γ, as described previously
(24). Briefly, CD8-positive
splenocytes (5×105) were added to the plate as effector
cells. Then, either bone marrow-derived dendritic cells (BM-DCs;
1×105) pulsed with OVA peptide (10 μg/ml; as target
cells) or non-pulsed BM-DCs (1×105; as control cells)
were added. The plate was then incubated for 37°C, for 20 h in the
presence of 5% CO2. Spots were counted automatically
using the Eliphoto system (Minerva Tech).
Isolation of mouse tumors and flow
cytometry
The mice were sacrificed and the dorsal tumors were
dissected, cut into small pieces, and digested with collagenase (1
mg/ml) for 20 min at 37°C. After the intratumoral injection of
OVA257–264 peptide, tumor cells were isolated and
stained with anti-mouse H-2Kb bound to
OVA257–264 peptide (SIINFEKL)-PE (clone 25-D1.16,
BioLegend) or isotype control-PE (MOPC-21, BioLegend). To analyze
the local accumulation of antigen-specific CTLs in mice, isolated
tumor cells including tumor-infiltrating lymphocytes were stained
with H-2Kb OVA Tetramer-PE [OVA257–264
(SIINFEKL); MBL] for 30 min at room temperature. They were then
incubated with anti-mouse CD8-FITC (clone KT15, MBL), anti-mouse
PD-1-PE-Cy7 (clone 29F.1A12, BioLegend), anti-mouse CTLA-4-APC
(clone UC10-4B, BioLegend), or anti-mouse LAG-3-PerCP-Cy5.5 (clone
RTK2071, BioLegend) for 20 min at 4°C.
Quantitative real-time PCR
The tumors implanted into mice were dissected. Total
RNA was isolated from homogenized tumors using RNeasy mini kit
(Qiagen) according to the manufacturer’s instructions. First-strand
complementary deoxyribonucleic acid (cDNA) was synthesized using a
PrimeScript® II first-strand cDNA Synthesis kit
(Takara). Quantitative real-time PCR was then performed on an
Applied Biosystems 7500 FAST Real-time PCR system using Power
SYBR® Green (Applied Biosystems). We assessed the
expression of the chemokines CXCL10, CXCL12, and CCL3, and compared
them to β-actin. Data ware analyzed using delta-delta CT
methods. Primer sequences of the chemokines were as described
(25), and were purchased from
Sigma Genosys.
Statistical analysis
All statistical analyses were performed using PASW
Statistics software, version 18.0 (SPSS Inc.). Statistical
significance was defined as a value of P<0.05 based on a
two-tailed test.
Results
PD-1 expression ex vivo in GPC3
peptide-specific CTLs after vaccination in patients
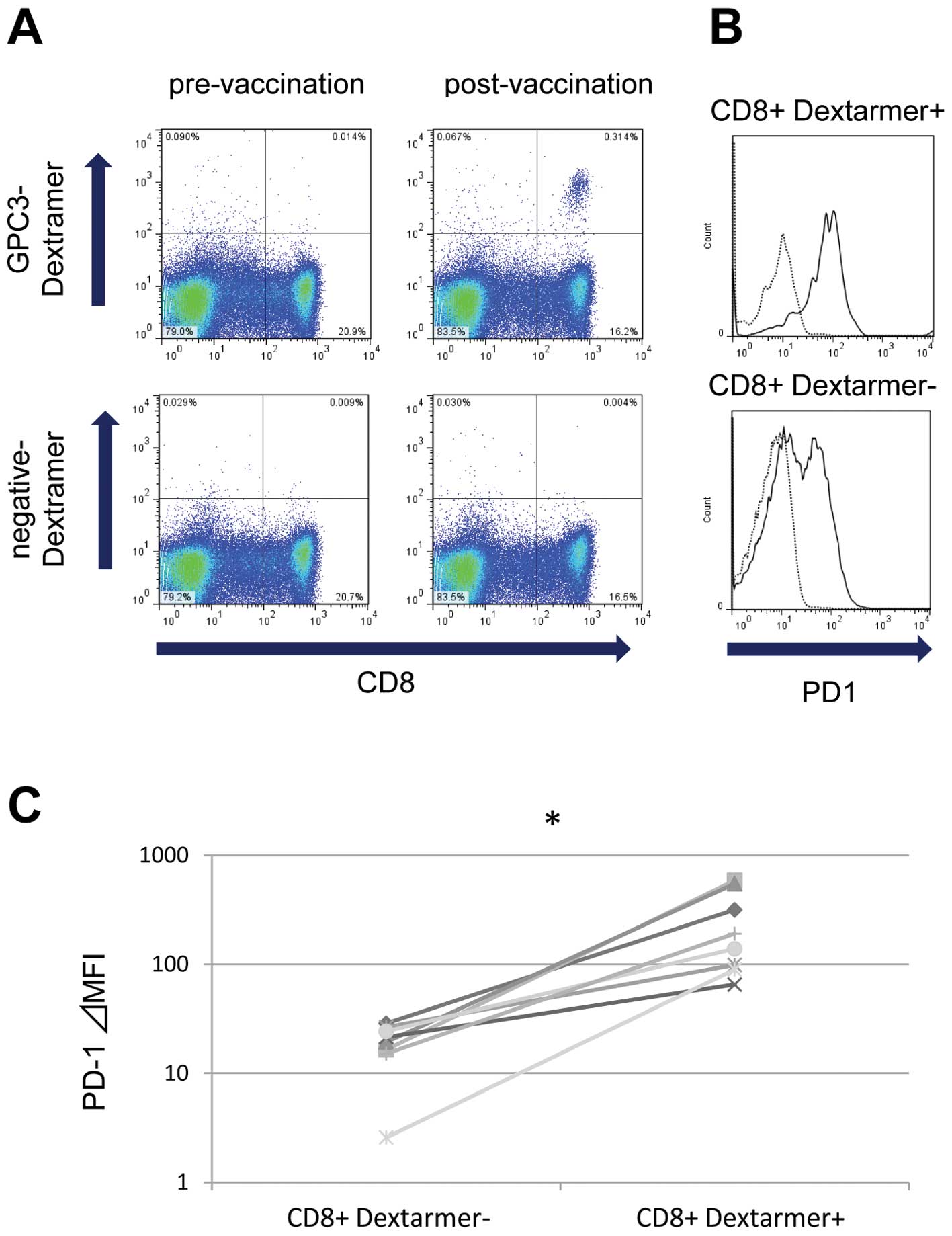
To investigate whether vaccine-induced CTLs were
affected by the PD-1/PD-L1 pathway, we measured the ex vivo
expression of PD-1 on vaccine-induced GPC3-specific CTLs using flow
cytometry with the GPC3-Dextramer. We used PBMCs obtained from
eight patients during clinical trials of the GPC3 peptide vaccine.
After vaccination, the frequency of GPC3-specific CTLs increased
and could be detected ex vivo, as shown in the
representative case 1 (Fig. 1A).
GPC3-Dextramer-positive CD8 lymphocytes had a higher expression of
PD-1 compared with GPC3-Dextramer-negative CD8 lymphocytes
(Fig. 1B; representative case 1).
In all eight patients with detectable GPC3-specific CTLs ex
vivo after vaccination, PD-1 expression levels were
significantly higher in GPC3-Dextramer-positive CD8 lymphocytes
compared with GPC3-Dextramer-negative CD8 lymphocytes (Fig. 1C). Before vaccination, no
GPC3-Dextramer-positive CD8 lymphocytes were detected ex
vivo; therefore, PD-1 expression was not analyzed.
PD-1 blockade augments the GPC3-specific
CTL clones that degranulate against liver cancer cell lines
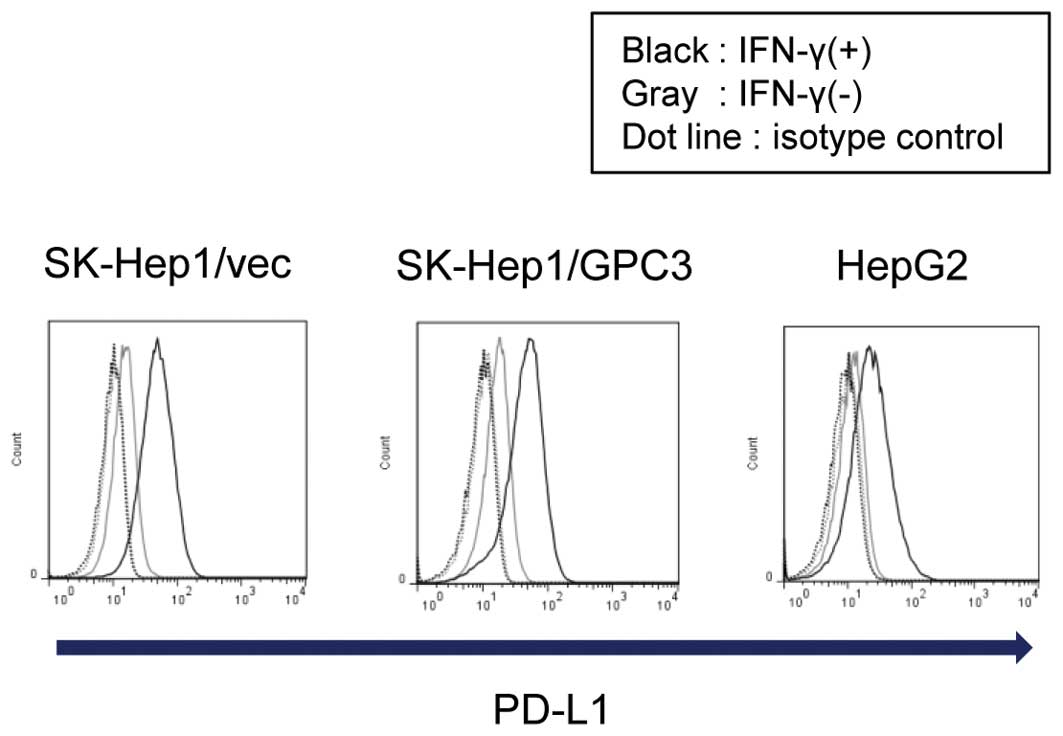
SK-Hep1/vec, SK-Hep1/GPC3, and HepG2 liver cancer
cell lines cultured with IFN-γ exhibited marked induction of PD-L1
on their surface (Fig. 2). This
suggests that liver cancer cells are invaded by IFN-γ-producing
CTLs via the PD-L1-mediated ligation of PD-1. Previously, several
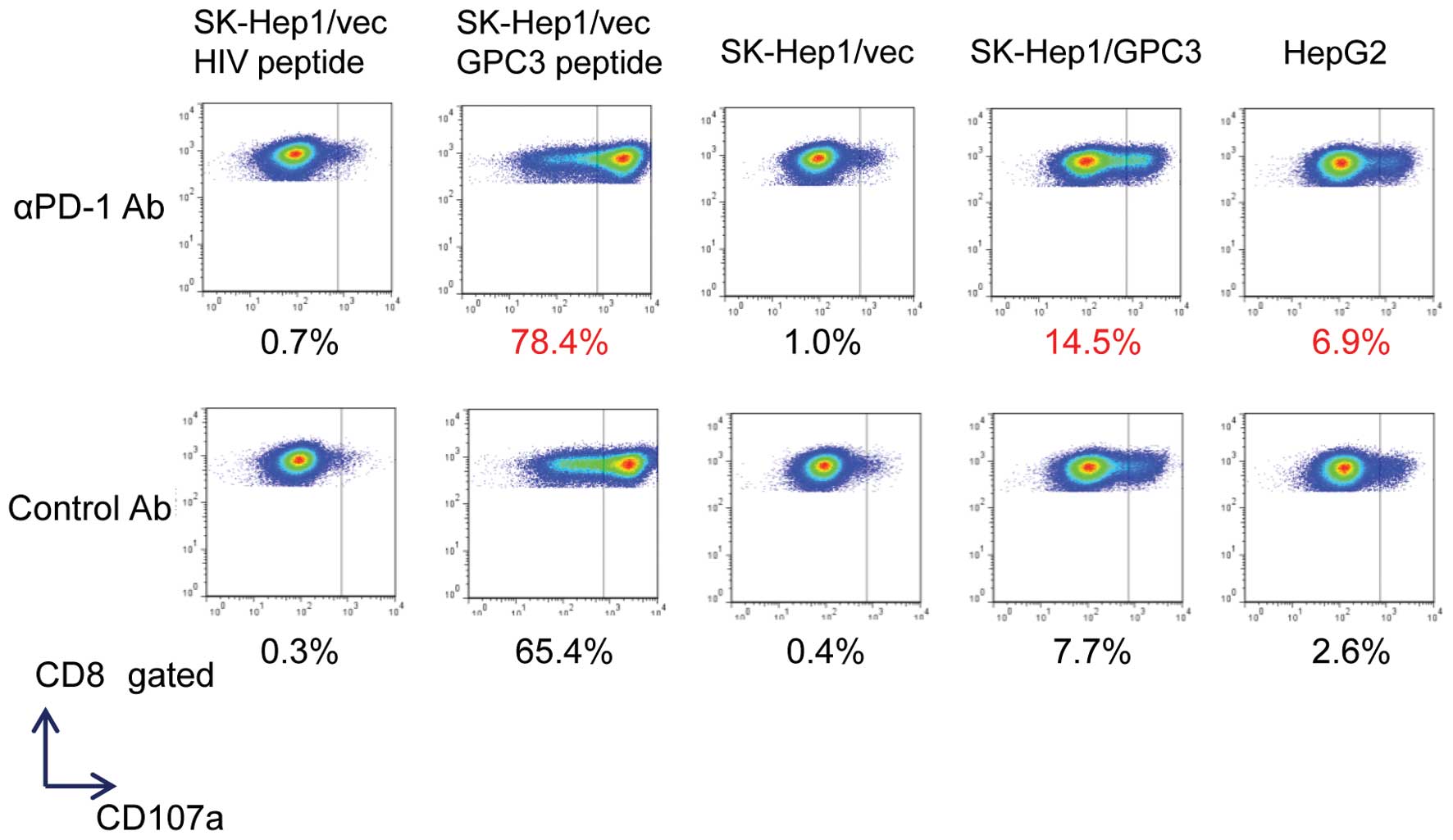
GPC3 peptide-specific CTL clones were established from PBMCs
isolated from vaccinated patients. These clones exhibited cytotoxic
activity against cancer cells expressing GPC3 endogenously
(9,26). Therefore, the CD107a
(lysosomal-associated membrane protein-1)-mediated externalization
of GPC3 peptide-specific CTL clones was examined upon exposure to
liver cancer cell lines. The externalization of CD107a could be a
surrogate marker to identify the antigen-specific CTLs that
degranulate against tumor cells (27). CTL clones mobilized CD107a in
response to SK-Hep1/vec pulsed with GPC3144–152 peptide,
SK-Hep-1/GPC3, and HepG2 (GPC3+,
HLA-A*02:01+), but not in response to pulsed
SK-Hep1/vec with HIV19–27 (Fig. 3). Furthermore, PD-1 blockade
enriched the population of GPC3-specific CTLs that degranulated
against only GPC3-positive liver cancer cell lines (SK-Hep1/vec
pulsed with GPC3144–152 peptide, SK-Hep1/GPC3 and
HepG2). These results suggest that blocking the interaction between
PD-1 and PD-L1 enhanced the antitumor effect of CTLs in liver tumor
cells that evade CTLs via PD-L1 expression.
Combination of a peptide vaccine and
αPD-1 Ab suppresses tumor growth in vivo synergistically
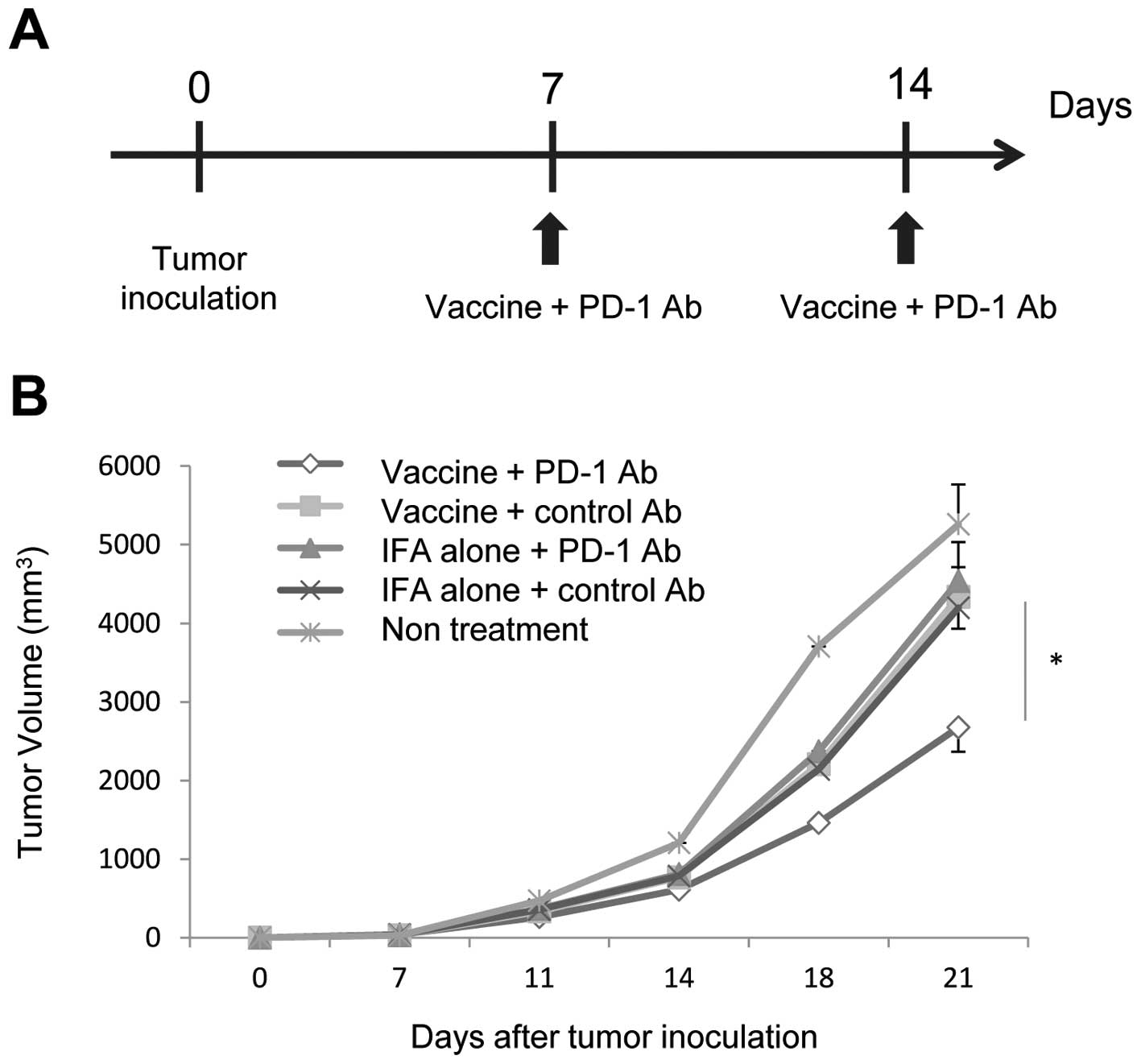
Intratumoral injection with OVA257–264
peptide (SIINFEKL) effectively inhibited the growth of OVA-negative
tumors in a mouse model treated with a peptide vaccine (22). Therefore, we performed in
vivo therapeutic experiments using intratumoral OVA peptide
vaccine and αPD-1 Ab in tumor implanted mice. Mice were implanted
with RMA tumor cells on day 0, and established tumors (3–6 mm in
diameter) were treated with OVA peptide emulsified with IFA
(vaccine) or vehicle emulsified with IFA (IFA alone) in combination
with αPD-1 Ab or control Ab on day 7. An additional dose of vaccine
and αPD-1 Ab was administered on day 14 after tumor inoculation
(Fig. 4A). On day 21, one mouse in
the untreated group was dead, and all other mice were alive. The
tumor volume of mice treated using the combination therapy of
vaccine and αPD-1 Ab was significantly less than those treated with
the appropriate control (Fig. 4B,
n=10). Treatment with vaccine/control Ab or IFA alone/αPD-1 Ab did
not inhibit tumor growth compared with IFA alone/control Ab
treatment. These data suggest that the combination of peptide
vaccine and αPD-1 Ab had a synergistic antitumor effect.
Vaccine and αPD-1 Ab treatment increases
the number of peptide-specific CTLs within mouse tumors
The loading of injected peptide onto major
histocompatibility complex (MHC) class I molecules in tumor cells
in vivo was reported previously using IFN-γ ELISPOT assays
(22). In the present study, RMA
(OVA-, H-2Kb) tumor cells were inoculated onto the backs
of C57/BL6 mice. When the tumor diameter reached 3–6 mm, 50 μg
H-2Kb-restricted OVA257–264 peptide was
injected into the tumor. After 96 h, the tumors were dissected, cut
into small pieces, and digested using collagenase. To investigate
whether the injected peptide was loaded onto the MHC class I
molecules in the tumor cells in a solid mass, flow cytometry using
anti-mouse H-2Kb bound to OVA257–264 peptide
was performed. The loading of H-2Kb-restricted
OVA257–264 peptide onto MHC class I of tumor cells was
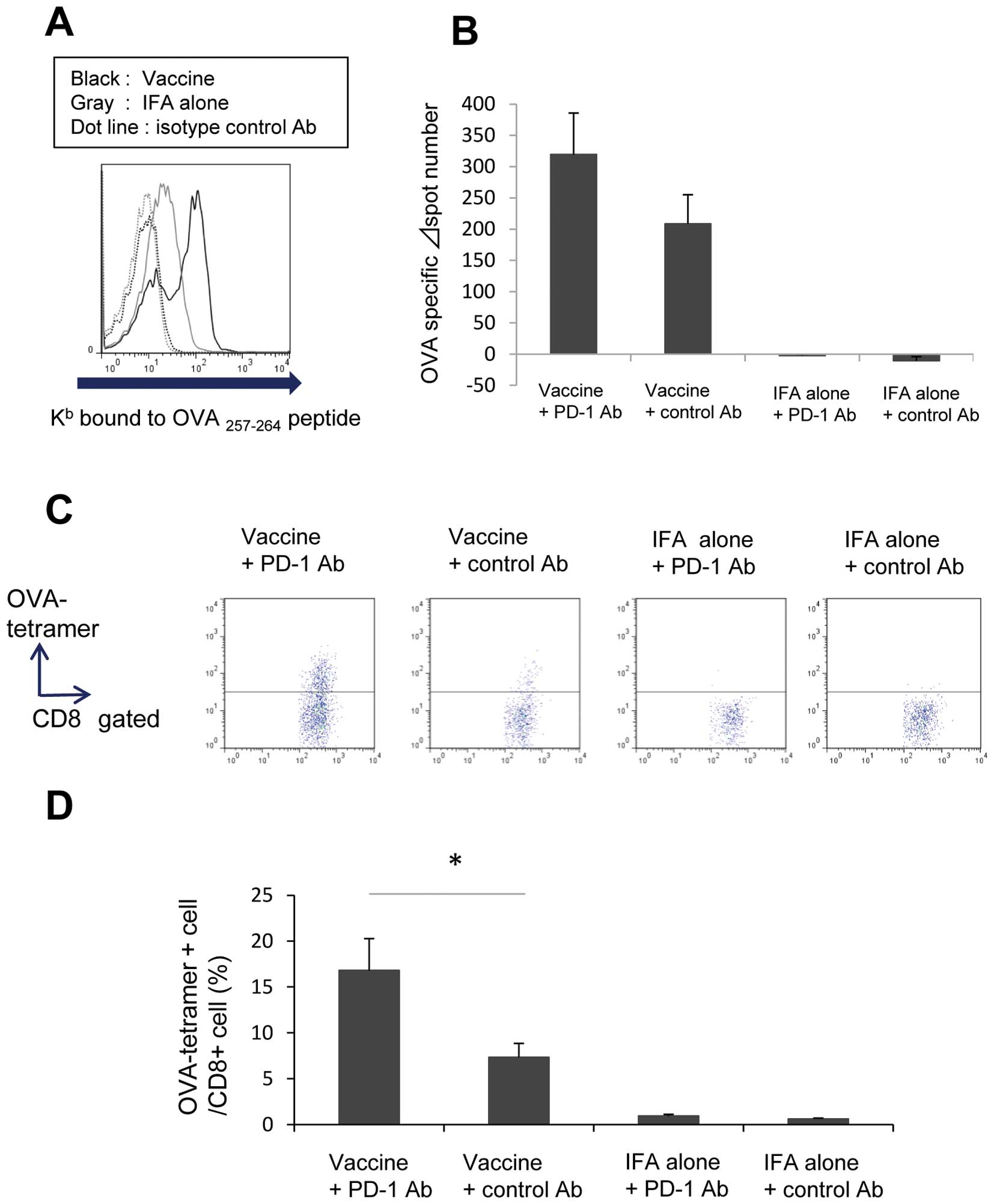
detected (Fig. 5A).
To evaluate the immunological response to
intratumoral OVA peptide vaccine and αPD-1 Ab, the spleens and
tumors of mice treated with the same schedule were analyzed as
described previously (Fig. 4A).
Peptide-specific immune responses were detected in the spleens of
mice treated with intratumoral OVA peptide injection using IFN-γ
ELISPOT assays (Fig. 5B). Mice
that received the combination of intratumoral OVA peptide injection
and αPD-1 Ab exhibited an increased number of OVA peptide-specific
CTLs compared with those treated with control Ab on day 14
(n=10).
To obtain direct evidence that the combination of
peptide vaccine and αPD-1 Ab led to the local accumulation of
antigen-specific CTLs, an OVA tetramer assay was performed in mice.
OVA-tetramer-positive CD8 lymphocytes could be detected within a
tumor using flow cytometry on day 21. Mice that received the
combination of OVA peptide vaccine and αPD-1 Ab had a significantly
increased number of OVA peptide-specific CTLs compared with those
treated with control Ab (Fig. 5C and
D; n=8).
Inhibitory receptors on
tumor-infiltrating T lymphocytes and the expression of
chemokines
The expression of inhibitory receptors on
peptide-specific CTLs at the tumor site was assessed to investigate
the mechanism of CTL accumulation in the tumors of mice treated
with the combination therapy of peptide vaccine and αPD-1 Ab.
RMA-bearing mice were treated with intratumoral OVA peptide
injection combined with αPD-1 Ab or control Ab, as described
previously (Fig. 4A). The
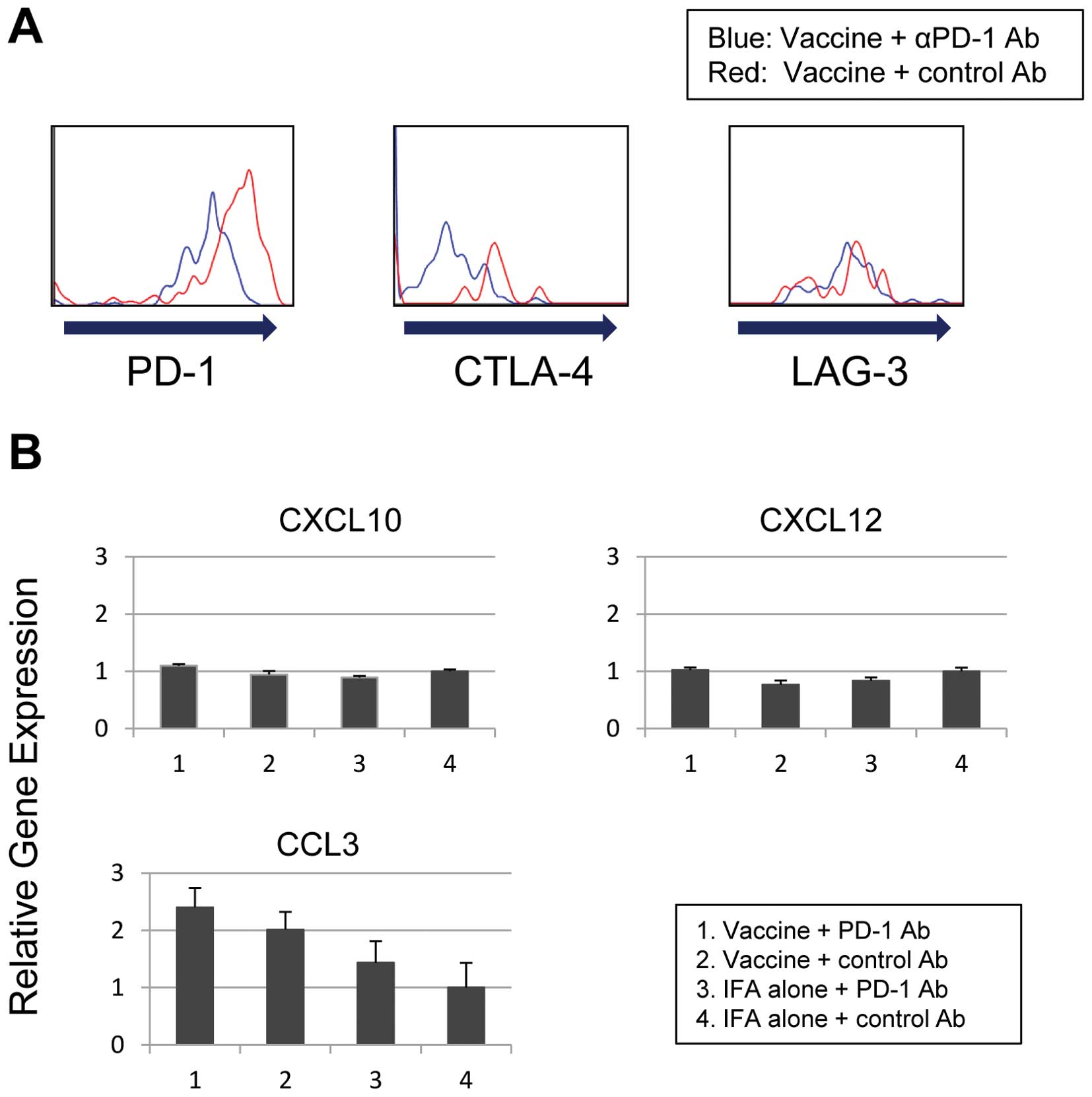
expression of PD-1, CTLA-4, and LAG-3 in OVA tetramer-positive CD8
lymphocytes within the tumor on day 21 was analyzed using flow
cytometry. The expression of the inhibitory receptors PD-1 and
CTLA-4 was decreased in OVA-tetramer positive CD8 lymphocytes in
the αPD-1 Ab group compared with the control Ab group (Fig. 6A). However, αPD-1 Ab treatment did
not decrease LAG-3 expression in OVA tetramer-positive CD8
lymphocytes.
The expression of chemokines within the tumor on day
21 was examined using quantitative real-time PCR. The expression of
the chemokine CCL3 was elevated in mice treated with the
combination of intratumoral OVA peptide injection and αPD-1 Ab
(Fig. 6B). The expression of the
chemokines CXCL10 and CXCL12 was unchanged.
Discussion
Many tumor antigens have been identified in HCC, and
their potential clinical utility for the development of
cancer-specific immunotherapy has been investigated (28–31).
GPC3 is a promising target of antigen-specific immunotherapy
because it is overexpressed specifically in human HCC (3,4). In
addition, it promotes tumor growth by stimulating canonical Wnt
signaling (32) or the Hippo
pathway (33). A phase I clinical
trial of a GPC3-derived peptide vaccine in patients with advanced
HCC showed that it had the potential to improve overall survival,
which was associated with vaccine-induced CTLs (8). However, the antitumor effects of the
peptide-based tumor vaccine alone were not satisfactory in patients
with advanced HCC (8,29–31).
Several studies identified molecules associated with the tumor
escape mechanism, such as PD-1/PD-L1, Fas/ FasL, and Decoy receptor
3, which might explain the poor immunogenicity and limitations of
the antitumor effects of cancer vaccines alone in patients with
advanced HCC (16,17,34,35).
Therefore, the present study examined whether blocking PD-1/PD-L
enhanced the antitumor effects of peptide vaccines in HCC.
The inhibitory receptor PD-1, was upregulated in
GPC3-specific CTLs of HCC patients vaccinated using GPC3 peptide,
consistent with previous reports of melanoma vaccine trials
(21,27). CTLs for some tumor antigens might
not be detected directly ex vivo. The ex vivo
analysis of antigen-specific CTLs from uncultured PBMCs could
provide strong and novel immunological evidence in HCC vaccine
trials. Fourcade et al reported that the upregulation of
PD-1 and Tim-3 on CTLs was correlated with the expansion of
melanoma-peptide vaccine-induced NY-ESO-1-specific CTLs (21). Further studies are necessary to
understand the potential clinical efficacy of vaccine-induced
CTLs.
In this experimental model, IFN-γ induced PD-L1
expression in liver cancer cell lines. It was also demonstrated
that blocking PD-1 increased the number of GPC3-specific CTL clones
that degranulate against these liver cancer cell lines in
vitro. These results suggest that blocking the interaction
between PD-1 and PD-L1 enhanced the antitumor effects of CTL in
liver cancer cells that evaded CTLs by expressing PD-L1. In
contrast, Xu et al reported that αPD-L1 or αCTLA-4 Abs did
not enhance cytokine secretion and the proliferation of peripheral
GPC3-specific CD8+ T-cell from HCC patients
significantly (36). Differences
in the effects of blocking PD-1 and PD-L1 might account for the
differences between spontaneous GPC3-specific CTLs and
vaccine-induced CTLs.
The combination of a peptide vaccine with αPD-1 Ab
enhanced tumor suppression and antigen-specific T cell infiltration
into the tumors of mouse models. The exact mechanisms by which CTLs
accumulate into tumors by blocking PD-1 are unclear. A previous
study in a mouse model of adoptive cell transfer demonstrated that
blocking PD-1 increased the production of CXCL10 by bone
marrow-derived myeloid cells, which enhanced the recruitment of
CTLs in the tumor (25). We
hypothesize that the αPD-1 Ab affected chemokine expression, which
resulted in recruitment of vaccine-induced CTLs to the tumor. In
the present study, the experimental model did not show a change in
the expression of CXCL10. However, the expression of CCL3 was
elevated by the combination treatment with vaccine and αPD-1 Ab.
Furthermore, blocking PD-1 decreased the expression of inhibitory
receptors in peptide-specific CTLs at the tumor site. Recently,
mouse models revealed that peptide/IFA vaccination increased the
antigen-driven expression of the inhibitory receptors PD-1, LAG-3,
CTLA-4, and Tim-3 in CTLs, suggesting partial exhaustion (37). PD-1 blockade might be a rational
strategy that could be used to rescue CTLs in a state of
exhaustion. Interestingly, αPD-1 Ab therapy did not decrease LAG-3
expression in TILs; however, CTLA-4 expression was decreased,
suggesting the partial rescue of CTL from exhaustion. A previous
study reported that dual treatment with αLAG-3 and αPD-1 Ab was
effective in mice with established tumors (38) as well as during the in vitro
expansion of human NY-ESO-1-specific CTLs (39). Furthermore, Sierro et al
reported that blocking both PD-1 and PD-L1 might further enhance
the antitumor effects of tumor vaccines in mouse models (40).
Based on the results of this clinical trial, the
GPC3 peptide vaccine has fewer side effects due to its antigen
specificity (8). Enhancing GPC3
peptide vaccine therapy is considered to be promising in terms of
sustained tumor control in HCC patients. These data suggest that
use of αPD-1 Ab could enhance the antitumor effects of a peptide
vaccine, and provide the foundation for the clinical development of
a combination therapy.
Acknowledgements
We thank Kayoko Shoda for technical assistance. We
also thank Dr Shigehisa Kitano (National Cancer Center), Masashi
Minami, Takao Yoshida and Hirotsugu Takano (Ono Pharmaceutical Co.)
for scientific advice. Y.S. would like to thank the Foundation for
Promotion of Cancer Research (Japan) for the Third-Term
Comprehensive Control Research for Cancer for the award of a
research resident fellowship. This study was supported in part by
the National Cancer Center Research and Development Fund (25-A-7),
as well as Research for Promotion of Cancer Control Programmes,
Research on Applying Health Technology, and Third Term
Comprehensive Control Research for Cancer from the Ministry of
Health, Labor and Welfare, Japan and a research funding from Ono
Pharmaceutical Co., Ltd. T.N. is a scientific advisor for Ono
Pharmaceutical Co., Ltd.
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
CTL
|
cytotoxic T lymphocyte
|
|
GPC3
|
glypican-3
|
|
PD-1
|
programmed death-1
|
|
PBMC
|
peripheral blood mononuclear cell
|
|
HLA
|
human leukocyte antigen
|
|
IFN-γ
|
interferon-γ
|
|
MHC
|
major histocompatibility complex
|
References
|
1
|
Breous E and Thimme R: Potential of
immunotherapy for hepatocellular carcinoma. J Hepatol. 54:830–834.
2011. View Article : Google Scholar
|
|
2
|
Greten TF, Manns MP and Korangy F:
Immunotherapy of hepatocellular carcinoma. J Hepatol. 45:868–878.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakatsura T, Yoshitake Y, Senju S, et al:
Glypican-3, overexpressed specifically in human hepatocellular
carcinoma, is a novel tumor marker. Biochem Biophys Res Commun.
306:16–25. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Capurro M, Wanless IR, Sherman M, et al:
Glypican-3: a novel serum and histochemical marker for
hepatocellular carcinoma. Gastroenterology. 125:89–97. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakatsura T, Komori H, Kubo T, et al:
Mouse homologue of a novel human oncofetal antigen, glypican-3,
evokes T-cell-mediated tumor rejection without autoimmune reactions
in mice. Clin Cancer Res. 10:8630–8640. 2004. View Article : Google Scholar
|
|
6
|
Komori H, Nakatsura T, Senju S, et al:
Identification of HLA-A2- or HLA-A24-restricted CTL epitopes
possibly useful for glypican-3-specific immunotherapy of
hepatocellular carcinoma. Clin Cancer Res. 12:2689–2697. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shirakawa H, Suzuki H, Shimomura M, et al:
Glypican-3 expression is correlated with poor prognosis in
hepatocellular carcinoma. Cancer Sci. 100:1403–1407. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sawada Y, Yoshikawa T, Nobuoka D, et al:
Phase I trial of a glypican-3-derived peptide vaccine for advanced
hepatocellular carcinoma: immunologic evidence and potential for
improving overall survival. Clin Cancer Res. 18:3686–3696. 2012.
View Article : Google Scholar
|
|
9
|
Yoshikawa T, Nakatsugawa M, Suzuki S, et
al: HLA-A2-restricted glypican-3 peptide-specific CTL clones
induced by peptide vaccine show high avidity and antigen-specific
killing activity against tumor cells. Cancer Sci. 102:918–925.
2011. View Article : Google Scholar
|
|
10
|
Agata Y, Kawasaki A, Nishimura H, et al:
Expression of the PD-1 antigen on the surface of stimulated mouse T
and B lymphocytes. Int Immunol. 8:765–772. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Freeman GJ, Long AJ, Iwai Y, et al:
Engagement of the PD-1 immunoinhibitory receptor by a novel B7
family member leads to negative regulation of lymphocyte
activation. J Exp Med. 192:1027–1034. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iwai Y, Ishida M, Tanaka Y, Okazaki T,
Honjo T and Minato N: Invovement of PD-L1 on tumor cells in the
escape from host immune system and tumor immunotherapy by PD-L1
blockade. Proc Natl Acad Sci USA. 99:12294–12297. 2002.PubMed/NCBI
|
|
13
|
Iwai Y, Terawaki S and Honjo T: PD-1
blockade inhibits hematogenous spread of poorly immunogenic tumor
cells by enhanced recruitment of effector T cells. Int Immunol.
17:133–144. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Topalian SL, Hodi FS, Brahmer JR, et al:
Safety, activity, and immune correlates of anti-PD-1 antibody in
cancer. N Engl J Med. 366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wolchok JD, Kluger H, Callahan MK, et al:
Nivolumab plus ipilimumab in advanced melanoma. N Engl J Med.
369:122–133. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi F, Shi M, Zeng Z, et al: PD-1 and
PD-L1 upregulation promotes CD8(+) T-cell apoptosis and
postoperative recurrence in hepatocellular carcinoma patients. Int
J Cancer. 128:887–896. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao Q, Wang XY, Qiu SJ, et al:
Overexpression of PD-L1 significantly associates with tumor
aggressiveness and postoperative recurrence in human hepatocellular
carcinoma. Clin Cancer Res. 15:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McGray AJ, Bernard D, Hallett R, et al:
Combined vaccination and immunostimulatory antibodies provides
durable cure of murine melanoma and induces transcriptional changes
associated with positive outcome in human melanoma patients.
Oncoimmunology. 1:419–431. 2012. View Article : Google Scholar
|
|
19
|
Mkrtichyan M, Najjar YG, Raulfs EC, et al:
Anti-PD-1 synergizes with cyclophosphamide to induce potent
antitumor vaccine effects through novel mechanisms. Eur J Immunol.
41:2977–2986. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duraiswamy J, Kaluza KM, Freeman GJ and
Coukos G: Dual blockade of PD-1 and CTLA-4 combined with tumor
vaccine efficacy restorres T cell rejection function in tumors.
Cancer Res. 73:3591–3603. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fourcade J, Sun Z, Pagliano O, et al: PD-1
and Tim-3 regulate the expansion of tumor antigen-specific
CD8+ T cells induced by melanoma vaccines. Cancer Res.
74:1045–1055. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nobuoka D, Yoshikawa T, Takahashi M, et
al: Intratumoral peptide injection enhances tumor cell antigenicity
recognized by cytotoxic T lymphocytes: a potential option for
improvement in antigen-specific cancer immunotherapy. Cancer
Immunol Immunother. 62:639–652. 2013. View Article : Google Scholar
|
|
23
|
Li B, VanRoey M, Wang C, Chen TH, Korman A
and Jooss K: Anti-programmed death-1 synergizes with granulocyte
macrophage colony-stimulating factor - secreting tumor cell
immunotherapy providing therapeutic benefit to mice with
established tumors. Clin Cancer Res. 15:1623–1634. 2009. View Article : Google Scholar
|
|
24
|
Iwama T, Horie K, Yoshikawa T, et al:
Identification of an H2-Kb or H2-Db
restricted and glypican-3-derived cytotoxic T-lymphocyte epitope
peptide. Int J Oncol. 42:831–838. 2013.
|
|
25
|
Peng W, Liu C, Xu C, et al: PD-1 blockade
enhances T-cell migration to tumors by elevating IFN-γ inducible
chemokines. Cancer Res. 72:5209–5218. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tada Y, Yoshikawa T, Shimomura M, et al:
Analysis of cytotoxic T lymphocytes from a patient with
hepatocellular carcinoma who showed a clinical response to
vaccination with a glypican-3-derived peptide. Int J Oncol.
43:1019–1026. 2013.
|
|
27
|
Wong RM, Scotland RR, Lau RL, et al:
Programmed death-1 blockade enhances expansion and functional
capacity of human melanoma antigen-specific CTLs. Int Immunol.
19:1223–1234. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mizukoshi E, Nakamoto Y, Arai K, et al:
Comparative analysis of various tumor-associated antigen-specific
T-cell responses in patients with hepatocellular carcinoma.
Hepatology. 53:1206–1216. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Butterfield LH, Ribas A, Meng WS, et al:
T-cell responses to HLA-A*0201 immunodominant peptides
derived from alpha-fetoprotein in patients with hepatocellular
cancer. Clin Cancer Res. 9:5902–5908. 2003.PubMed/NCBI
|
|
30
|
Butterfield LH, Ribas A, Dissette VB, et
al: A phase I/II trial testing immunization of hepatocellular
carcinoma patients with dendritic cells pulsed with four
alpha-fetoprotein peptides. Clin Cancer Res. 12:2817–2825. 2006.
View Article : Google Scholar
|
|
31
|
Greten TF, Forner A, Korangy F, et al: A
phase II open trial evaluating safety and efficacy of a telomerase
peptide vaccination in patients with advanced hepatocellular
carcinoma. BMC Cancer. 10:2092010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Capurro MI, Xiang YY, Lobe C and Filmus J:
Glypican-3 promotes the growth of hepatocellular carcinoma by
stimulating canonical Wnt signaling. Cancer Res. 65:6245–6254.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng M, Gao W, Wang R, et al:
Therapeutically targeting glypican-3 via a conformation-specific
single-domain antibody in hepatocellular carcinoma. Proc Natl Acad
Sci USA. 110:E1083–E1091. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nagao M, Nakajima Y, Hisanaga M, et al:
The alteration of Fas receptor and ligand system in hepatocellular
carcinomas: how do hepatoma cells escape from the host immune
surveillance in vivo? Hepatology. 30:413–421. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen C, Zhang C, Zhuang G, et al: Decoy
receptor 3 overexpression and immunologic tolerance in
hepatocellular carcinoma (HCC) development. Cancer Invest.
26:965–974. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu Y, Li H, Gao RL, Adeyemo O, Itkin M and
Kaplan DE: Expansion of interferon-gamma-producing multifunctional
CD4+ T-cells and dysfunctional CD8+ T-cells
by glypican-3 peptide library in hepatocellular carcinoma patients.
Clin Immunol. 139:302–313. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hailemichael Y, Dai Z, Jaffarzad N, et al:
Persistent antigen at vaccination sites induces tumor-specific
CD8(+) T cell sequestration, dysfunction and deletion. Nat Med.
19:465–472. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Woo SR, Turnis ME, Goldberg MV, et al:
Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate
T-cell function to promote tumoral immune escape. Cancer Res.
72:917–927. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Matsuzaki J, Gnjatic S, Mhawech-Fauceglia
P, et al: Tumor-infiltrating NY-ESO-1-specific CD8+ T
cells are negatively regulated by LAG-3 and PD-1 in human ovarian
cancer. Proc Natl Acad Sci USA. 107:7875–7880. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sierro SR, Donda A, Perret R, et al:
Combination of lentivector immunization and low-dose chemotherapy
or PD-1/PD-L1 blocking primes self-reactive T cells and induces
antitumor immunity. Eur J Immunol. 41:2217–2228. 2011. View Article : Google Scholar : PubMed/NCBI
|