Introduction
Cancer is described as one of the major world health
problems (1). Colorectal cancer is
the third most common malignancy worldwide and nearly 1.4 million
new cases were reported in 2012 (2). Accumulation of oncogenes and tumor
suppressor gene mutations can contribute to cancer development.
High-throughput DNA sequencing data suggested that thousands of
point mutations, translocations, amplifications, and deletions may
contribute to cancer development, and that the mutational range can
differ, even among tumors with identical histopathology (3). Therefore, any therapeutic strategy
designed to target individual signaling molecules has limitations
in improving current survival rates and novel strategies are
needed.
Metabolic reprogramming is one of the hallmarks of
cancer, in addition to gene mutation (4). To support rapid cell division and the
process of tumor progression, cancer cells need to generate energy
by reprogramming their metabolism. It is well established that
cancer cells can generate ATP through glycolysis rather than
oxidative phosphorylation, even in the presence of oxygen (the
Warburg effect) (5).
Characteristic metabolic reprogramming, including the Warburg
effect, is consistently seen in various cancers, despite numerous
gene mutations, indicating that cancer cell metabolic pathways
could be useful therapeutic targets. Hirayama et al
(6) and Soga et al
(7) reported metabolic profiling
of human colon and stomach cancers, and compared the levels of
metabolites in tumor and normal tissues using capillary
electrophoresis time-of-flight mass spectrometry (CE-TOFMS).
Recently, the use of metabolome analysis has remarkably developed
in various research fields, such as clinical research, cell
biology, and plant studies (8–10).
Metabolomics is the final step in the ‘omics’ cascade, of genomics,
transcriptomics, and proteomics, and can provide global information
on low-molecular-weight-metabolites (11,12).
Metabolome analysis could reveal the influences on cancer
metabolism of anticancer agents, and accelerate biomarker discovery
based on the determination of metabolomic differences between
normal and cancerous tissue.
Members of the cathelicidin family of antimicrobial
peptides are endogenous factors playing key roles in cancer
regulation (13). Human
cathelicidin antimicrobial protein, hCAP18, is the only member of
the cathelicidin family in human cells; its C-terminal domain,
LL-37, is released by proteolytic cleavage, and shows various
effects, such as antibacterial, antiviral, wound-healing, and
immunoregulatory effects (14,15).
LL-37 is expressed in epithelial cells of a number of organs
(16). A previous study showed
that the expression of LL-37 was markedly downregulated in human
colon cancer tissue, whereas exogenous LL-37 induced apoptotic cell
death in cultured colon cancer cells. In addition,
cathelicidin-deficient mice exhibited increased susceptibility to
azoxymethane-induced colon carcinogenesis (17).
We previously reported that a 27-residue analog of
the LL-37 peptide, FF/CAP18, induced apoptotic cell death, via
mitochondrial membrane depolarization and DNA fragmentation, in the
oral squamous cell carcinoma cell line SAS-H1, (18) and the colon carcinoma cell line
HCT116 (19). Although these
findings suggest that antimicrobial peptides have possible
anticancer effects and could be targeted for new therapeutic
strategies, the full mechanisms of their suppressive effects on
metabolic pathways are still largely unknown. In the present study,
using metabolome analysis by CE-TOFMS, we identified changes in
energy metabolism caused by FF/CAP18 during the process of
apoptosis in human colon cancer cells.
Materials and methods
Cell culture and peptides
The human HCT116 colon carcinoma-derived cell line
was provided by Dr Bert Vogelstein (Johns Hopkins University,
Baltimore, MD, USA). The cells were maintained in Dulbecco’s
modified Eagle’s medium (Nacalai Tesque, Kyoto, Japan) containing
10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA) and a 5%
antibiotic-antimycotic mixed stock solution (Nacalai Tesque) at
37°C and 5% CO2. Before being used for experiments,
cells were routinely maintained under exponential-proliferation
conditions. The cells were treated with a 0.25% trypsin-EDTA
solution (Nacalai Tesque) to dislodge them at each passage.
The primary structure of LL-37 is represented in a
single amino acid code as follows: LLGDFFRKSKEKIGKEFKRIV
QRIKDFLRNLVPRTES. To enhance antimicrobial activity, FF/CAP18 was
designed by the replacement of a glutamic acid residue and a lysine
residue with phenylalanine at positions 11 and 20, respectively, of
the 27mer (FRKSKEKIGKEFKRI VQRIKDFLRNLV) which resulted from the
removal of the first and last five amino acids of LL-37 (20). FF/CAP18 (FRKS
KEKIGKFFKRIVQRIFDFLRNLV) was synthesized by the method previously
described (18).
Detection of apoptosis using a combined
Annexin V-7-amino-actinomycin D (7-AAD) assay
One feature of the early stages of apoptosis is
externalization of plasma membrane phosphatidylserine to the cell
surface. Owing to this process, cells showing the early stages of
apoptosis can be identified via binding of Annexin V, which has
high affinity for phosphatidylserine, whereas cells in the late
stage of apoptosis or necrosis show no affinity for Annexin V.
Furthermore, 7-AAD, a fluorescent DNA-binding agent that
intercalates between cytosine and guanine, also allows the
distinction of cells that are alive, dead, or in the early or late
stages of apoptosis. The combination of these two reagents is
available as a powerful apoptosis-detection tool in the
Muse® Annexin V and Dead Cell assay kit (Merck
Millipore, Darmstadt, Germany). After incubation with FF/CAP18 for
96 h, cells were trypsinized, transferred into 1.5-ml microtubes,
and subjected to centrifugation at 800 × g for 5 min. Cell pellets
were resuspended in 100 μl of fresh medium, and the Muse Annexin V
and Dead Cell Dye assay kit reagent was added (100 μl to each tube)
with mixing. After incubation for 20 min at room temperature, cells
were applied to a Muse Cell Analyzer (Merck Millipore).
Metabolome extraction
Samples of cells and of conditioned medium were
obtained 96 h after administration of FF/CAP18. Cell samples were
washed twice with a 5% solution of mannitol and covered with
methanol. Cells were harvested after addition of the internal
standard solution (Human Metabolome Technologies, Tsuruoka, Japan),
and subjected to centrifugation for 5 min at 2,300 × g, 4°C. The
aqueous layers were collected into ultrafiltration units (EMD
Millipore, Billerica, MA, USA) and subjected to centrifugation for
2.5 h at 9,600 × g, 4°C. The conditioned medium from cell cultures
was directly collected to prepare medium samples. The sampled
medium was mixed with the internal standard solution, and subjected
to centrifugation for 2.5 h at 9,600 × g, 4°C.
Measurement of metabolites
CE-TOFMS was carried out using an Agilent 7100 CE
system equipped with an Agilent 6210 TOFMS system, Agilent 1100
high-performance liquid chromatography system with isocratic pump,
Agilent G1603A CE-Mass Spectrometry (MS) Adapter kit, and Agilent
G1607A CE Electrospray Ionization-MS Sprayer kit (Agilent
Technologies, Waldbronn, Germany). The systems were controlled
using the Agilent G2201AA ChemStation software, version B.03.01,
for CE (Agilent Technologies). Metabolites were analyzed using a
fused silica capillary (50 μm internal diameter × 80 cm length),
with commercial electrophoresis buffer (Solution ID: H3301-1001 for
cation analysis and I3302-1023 for anion analysis; Human Metabolome
Technologies) as the electrolyte. The sample was injected at a
pressure of 50 mbar for 10 sec (equivalent to ~10 nl) in the cation
analysis, and at 50 mbar for 25 sec (equivalent to ~25 nl) in the
anion analysis. The spectrometer was scanned across a
mass-to-charge ratio (m/z) of 50–1,000. Other conditions were as
previously described (21–23).
The peaks detected by CE-TOFMS were extracted using
MasterHands automatic integration software (Keio University,
Tsuruoka, Japan) in order to obtain peak information including m/z,
migration time (MT), and peak area (24). Signal peaks corresponding to
isotopomers, adduct ions, and other product ions of known
metabolites were excluded, and the remaining peaks were annotated
with putative metabolites from the Human Metabolome Technologies
metabolite database, based on their MT and m/z values. The
tolerance range for the peak annotation was configured at ±0.5 min
for MT and ±10 parts per million for m/z. In addition, peak areas
were normalized to those of the internal standards, and the
resultant relative area values were then further normalized to the
sample amount.
Hierarchical cluster analysis (HCA) and principal
component analysis (PCA) were carried out using the proprietary
software, PeakStat and SampleStat, respectively (Human Metabolome
Technologies). Detected metabolites were plotted on metabolic
pathway maps using the Visualization and Analysis of Networks
containing Experimental Data software (25).
Statistical analysis
In the combined Annexin V binding-7-AAD staining
assay, the ratio of cells at each apoptotic stage was expressed as
the mean ± standard deviation. Statistical differences were tested
with Student’s t-test. The statistical significance of differences
in the 177 intracellular metabolites and the 113 metabolites
detected in conditioned medium, between treated and untreated
cells, was determined using Welch’s t-test. A value of P<0.05
was considered significant.
Results
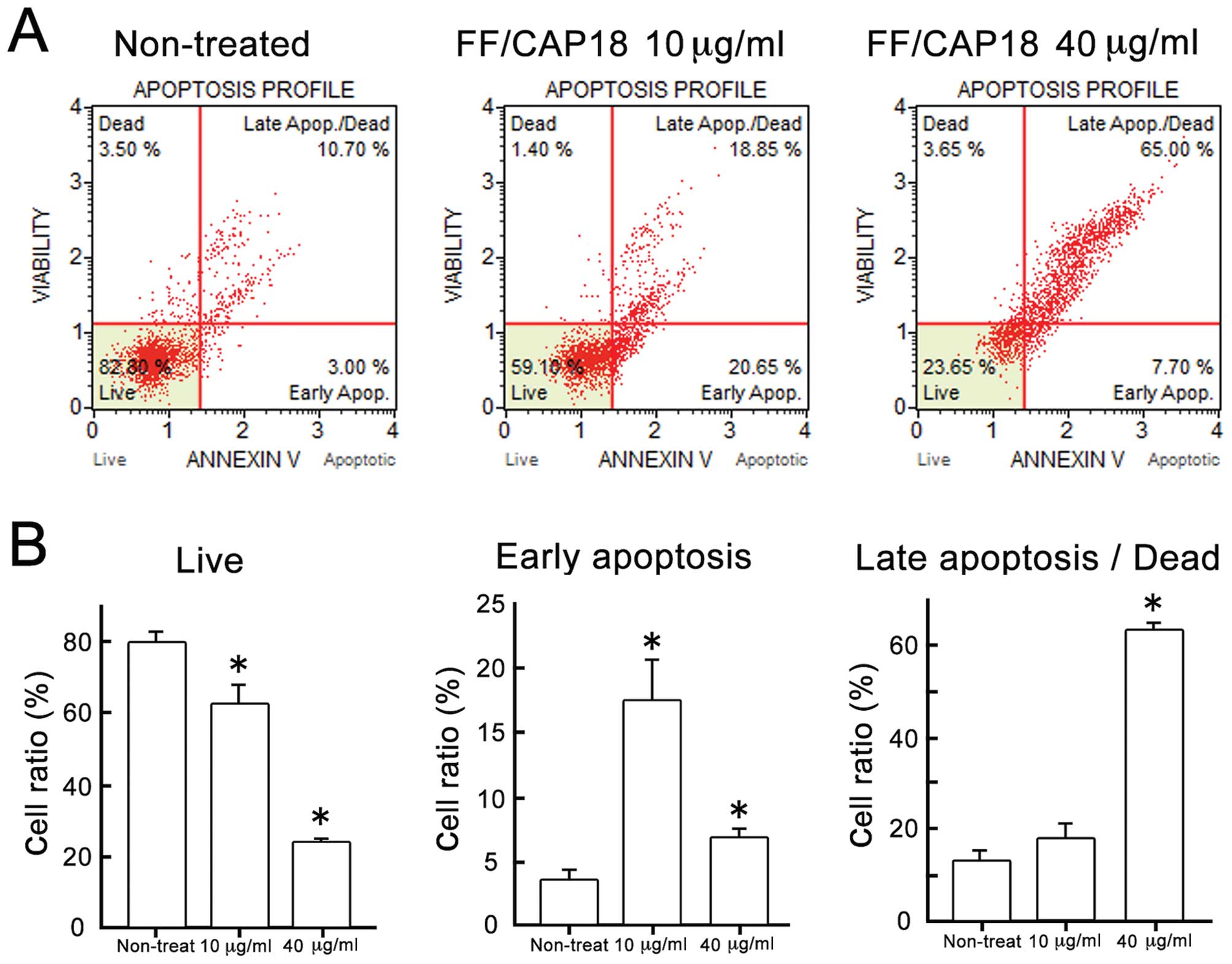
Apoptosis detection using the Annexin
V-7-AAD assay
Combined Annexin V and 7-AAD reactivity allowed
classification of cells into four groups, as follows: early
apoptotic cells [Annexin V (+) and 7-AAD (−)], late apoptotic or
dead cells [Annexin V (+) and 7-AAD (+)], dead cells [Annexin V (−)
and 7-AAD (+)], and live cells [Annexin V (−) and 7-AAD (−)]; see
the scatter plots in Fig. 1A.
Treatment of HCT116 cells with FF/CAP18 at 10 μg/ml induced high
affinity for Annexin V, as shown by the right shift of the scatter
plot compared with that of non-treated cells, indicating early
apoptosis (Fig. 1A, middle panel).
On the other hand, FF/CAP18 treatment at 40 μg/ml increased the
number of cells that were positive for Annexin V (+) and 7-AAD (+),
indicating that a high dose of FF/CAP18 induced apoptotic cell
death in HCT116 cells. The ratio of HCT116 cells at each stage of
apoptosis after treatment with the two different doses of FF/CAP18
is summarized in Fig. 1B. The
percentage of live cells decreased significantly in a
dose-dependent manner (Fig. 1B).
The percentage of cells in early apoptosis, however, significantly
increased with 10 μg/ml FF/CAP18 treatment, whereas, the percentage
of late apoptotic and dead cells only increased with 40 μg/ml
treatment (Fig. 1B). From these
results, we concluded that early-stage apoptosis was induced by a
comparatively low dose (10 μg/ml) of FF/CAP18, whereas high-dose
treatment (40 μg/ml) could cause late-stage apoptosis and cell
death.
Heat map and PCA representation of
metabolome data from HCT116 cells
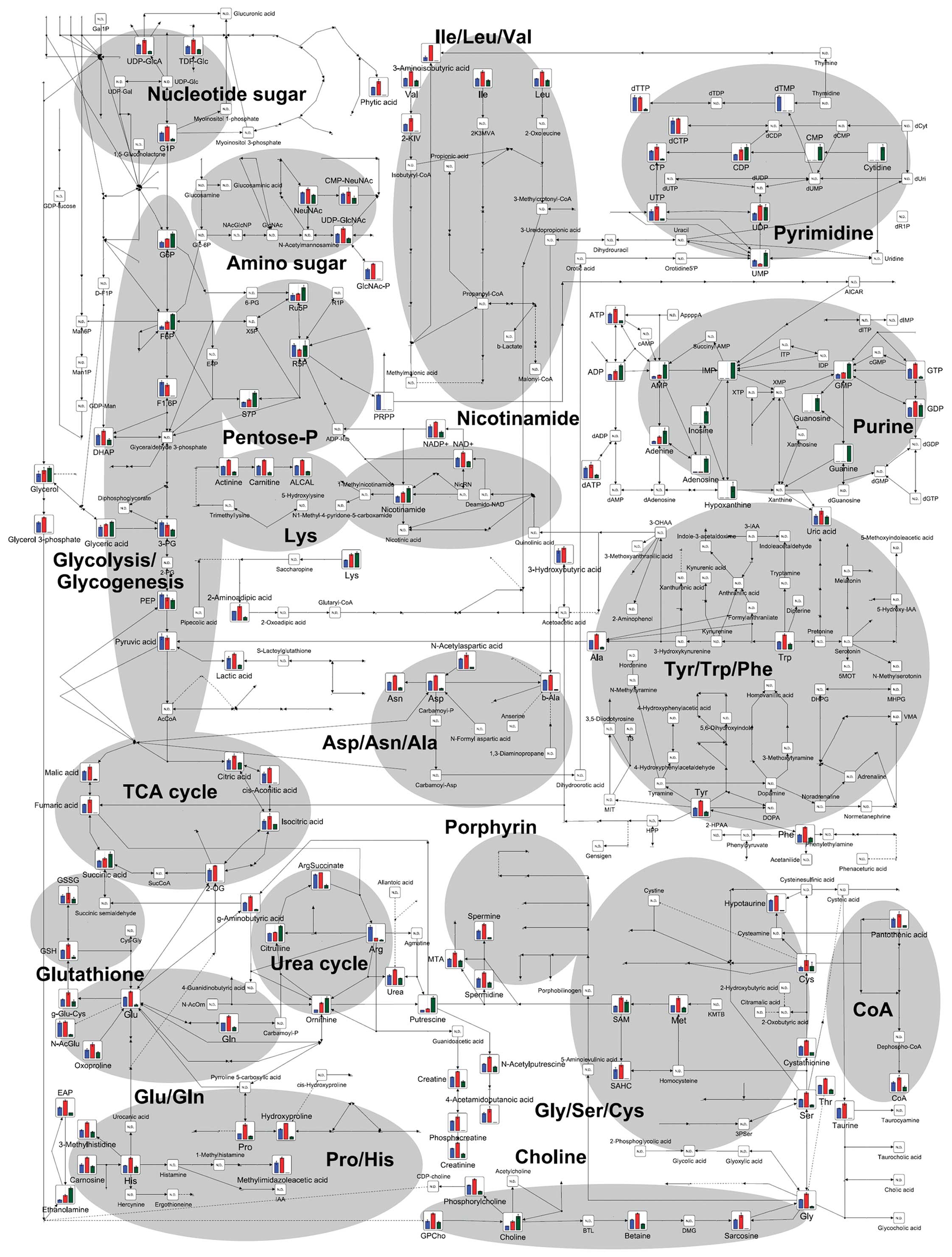
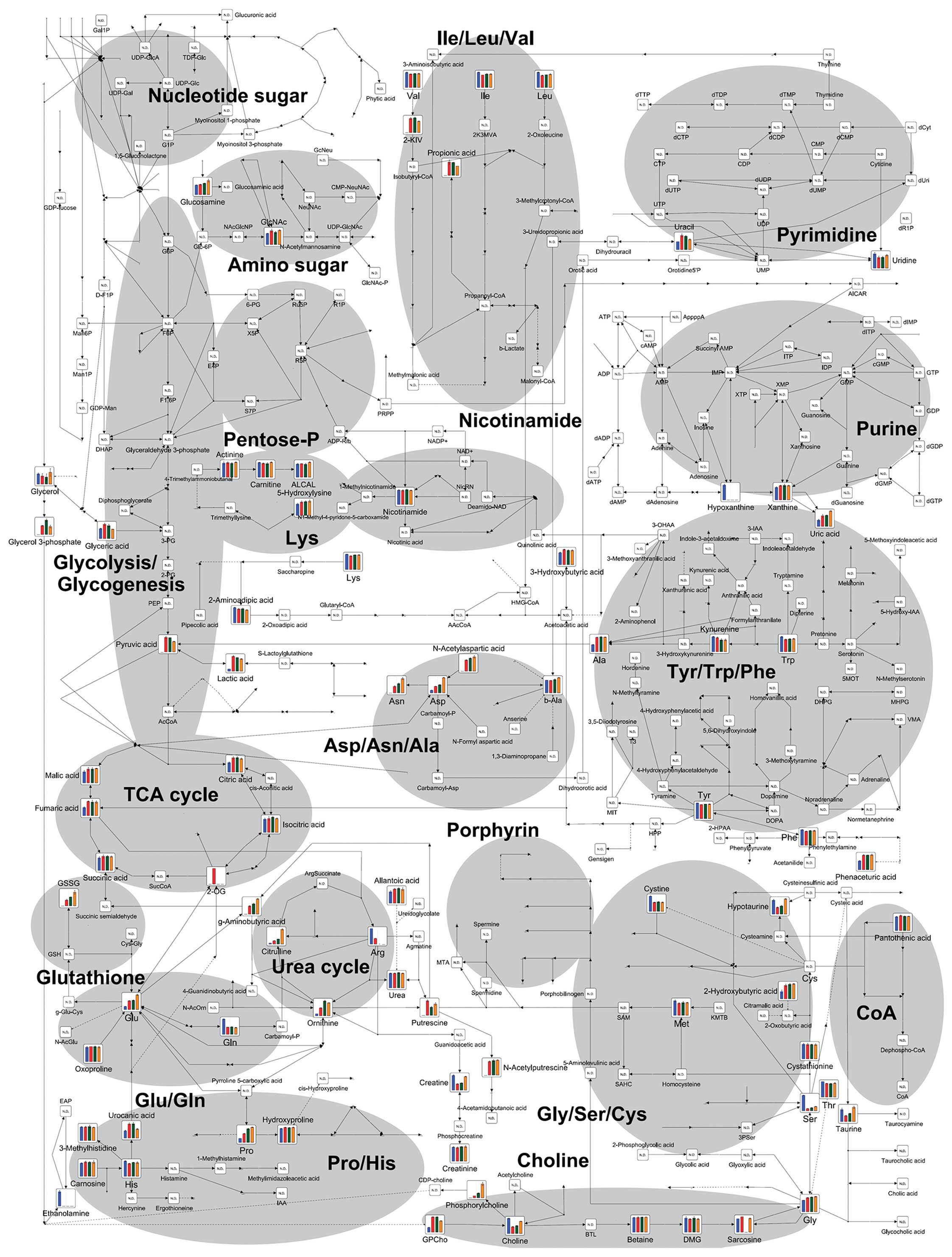
The 177 intracellular metabolites and 113
metabolites in conditioned medium were detected as peaks by
CE-TOFMS, and mapped onto metabolic pathways for ease of viewing,
as shown in Fig. 2 (cells) and
Fig. 3 (conditioned medium).
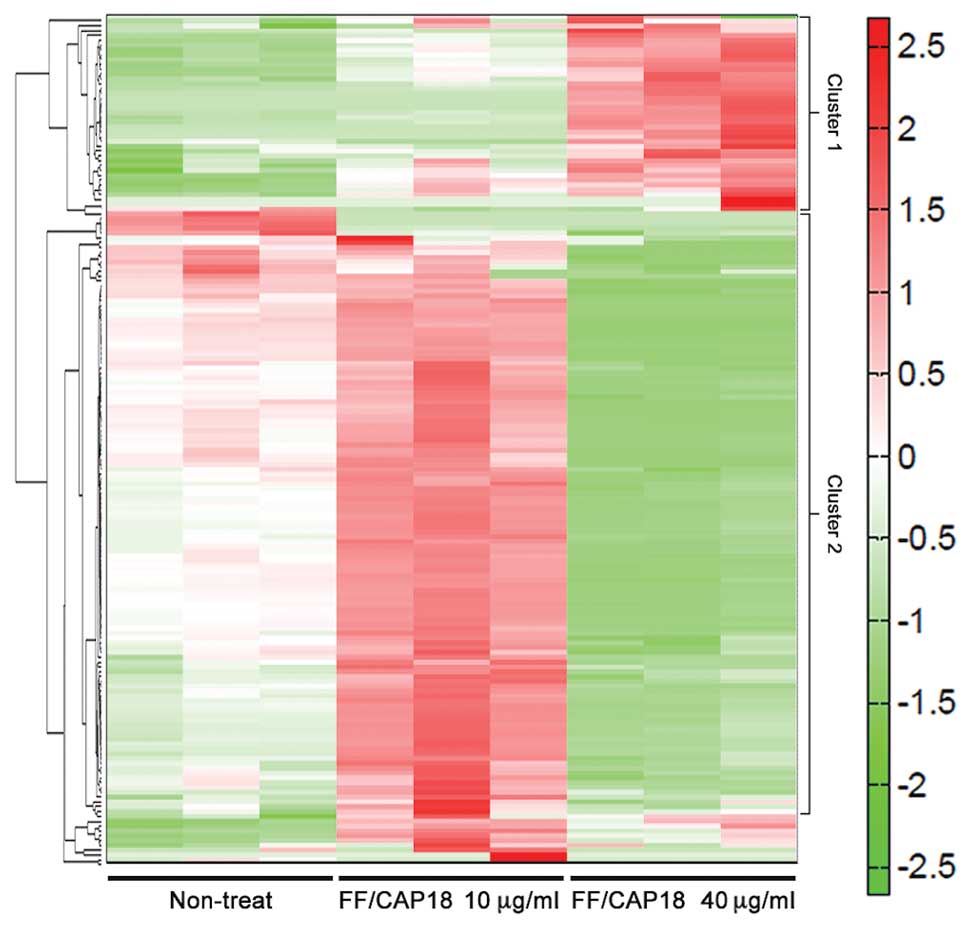
Overall trends of the intracellular metabolomic changes in HCT116
cells treated with FF/CAP18 and non-treated cells were analyzed by
Euclidean-distance-based HCA, and the results are presented as a
heat map (Fig. 4). The metabolomic
profile of HCT116 cells treated with 10 μg/ml FF/CAP18 showed high
values for the metabolites in cluster 2, including amino acids, and
tricarboxylic acid (TCA) cycle intermediates. In contrast, the
metabolomic profile of HCT116 cells treated with 40 μg/ml FF/CAP18
was reversed in comparison to the profile for treatment with 10
μg/ml FF/CAP18. We also confirmed that the metabolomic profiles of
HCT116 cells treated with FF/CAP18 at 10 or 40 μg/ml were reversed
for metabolites in cluster 1, including nucleotides and
nucleosides. These trends were made even clearer from the results
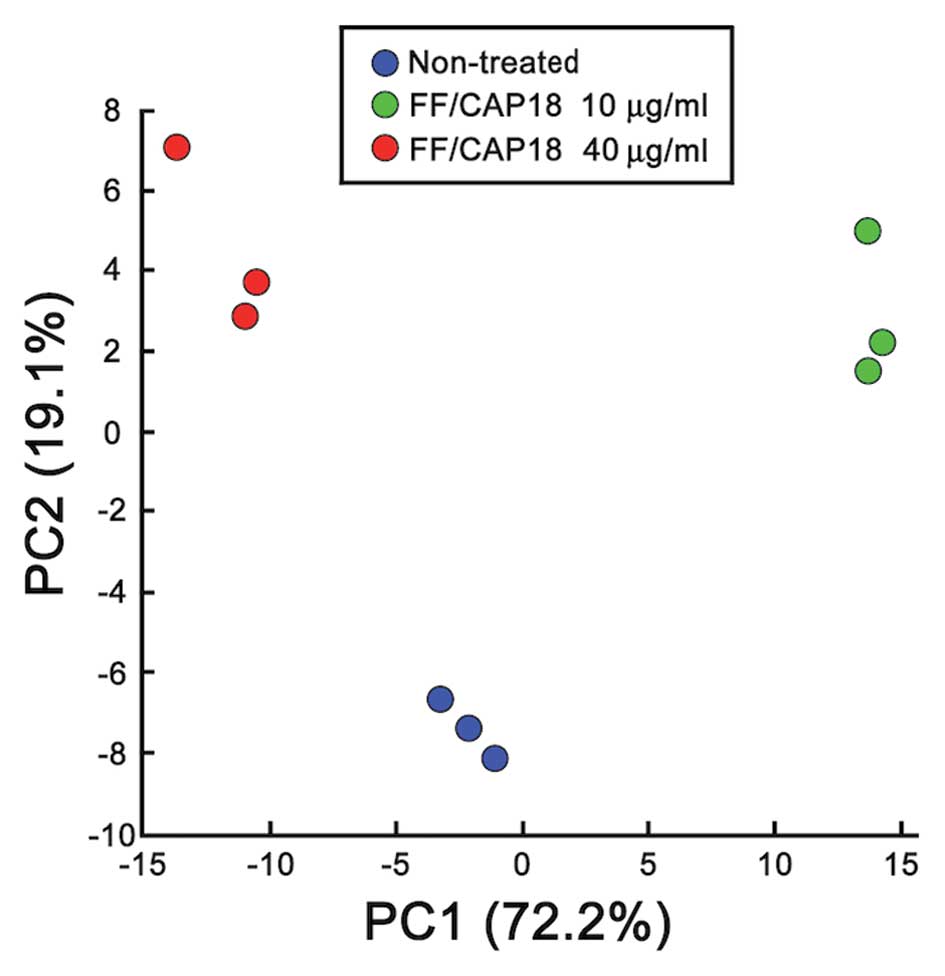
of the PCA of metabolome data for HCT116 cells treated with
FF/CAP18 (Fig. 5). The
concentration of FF/CAP18 was reflected in principal component 1;
principal component 2 demonstrated the difference between treatment
and non-treatment of cells with FF/CAP18. Therefore, treatment with
FF/CAP18 exerted a dramatic change on the metabolism of HCT116
cells, and that change depended on the concentration of
FF/CAP18.
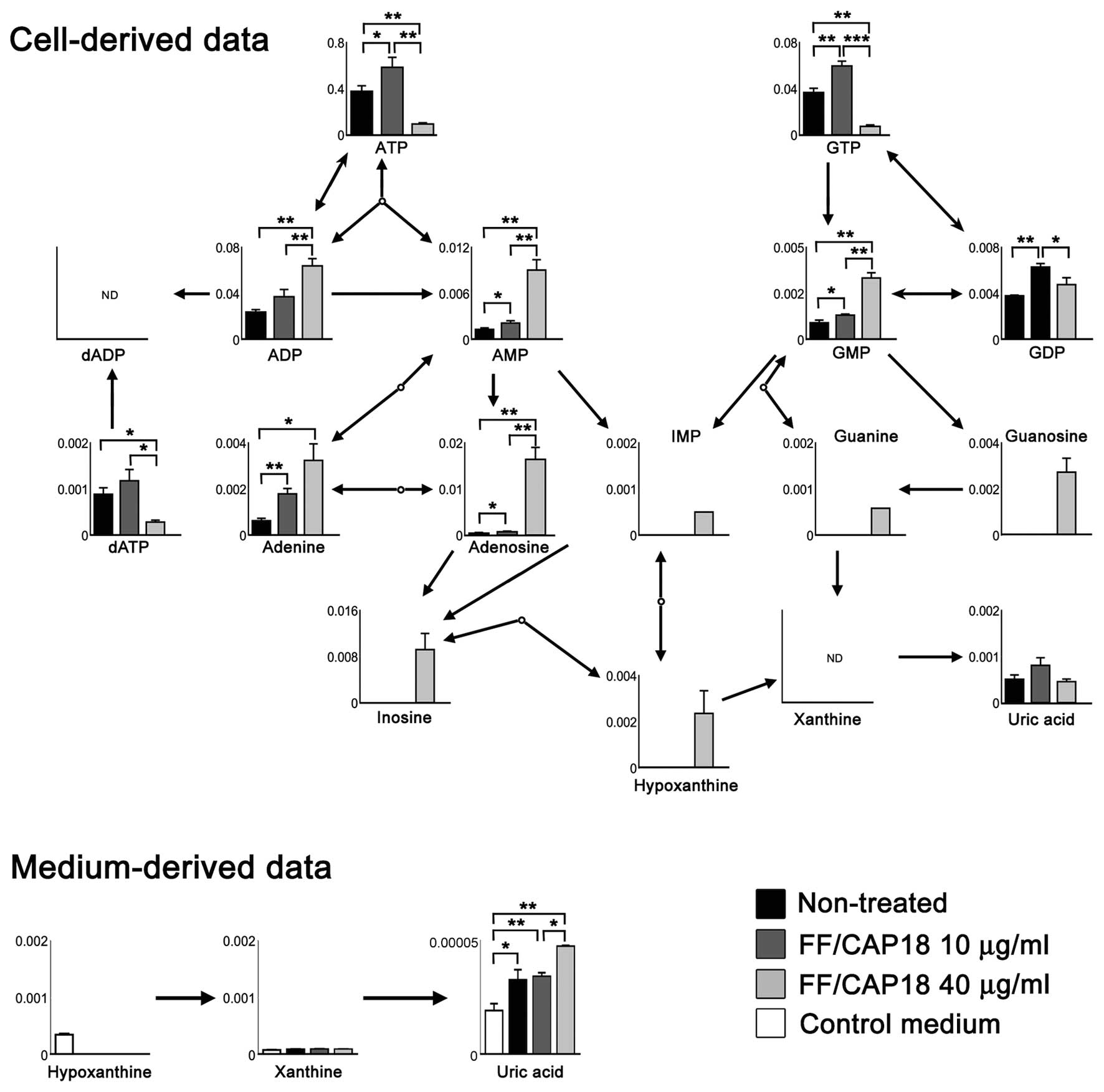
Purine metabolism
Purine metabolism is an important pathway in order
to supply nucleotides, such as 2′-deoxyadenosine triphosphate and
2′-deoxyguanosine triphosphate, for aggressive DNA synthesis in
cancer cells. Fig. 6 shows the
metabolic pathway of purine metabolism. The levels of adenosine
triphosphate (ATP) and guanosine triphosphate (GTP) measured in
HCT116 cells treated with FF/CAP18 at 10 μg/ml (dark grey bar) were
higher than in non-treated cells (black bar); simultaneously, a
slight increase was detected in the levels of adenosine diphosphate
(ADP), adenosine monophosphate (AMP), guanosine monophosphate
(GMP), and guanosine diphosphate (GDP). In contrast, in HCT116
cells treated with FF/CAP18 at 40 μg/ml (pale grey bar), there was
a substantial decrease in the levels of ATP and GTP, whereas the
levels of ADP, AMP, GMP, and GDP showed marked increases. These
results suggest a high-energy status in HCT116 cells treated with
the low dose (10 μg/ml) of FF/CAP18. However, HCT116 cells treated
with the high dose of FF/CAP18 (40 μg/ml) fall into an
energy-depleted status compared with non-treated and
low-dose-treated cells. We could also confirm the presence of
derivatives of purine, including adenine, hypoxanthine, and
guanine, in HCT116 cells treated with a high dose of FF/CAP18.
Additionally, increased uric acid was detected in conditioned
medium sampled after culturing HCT116 cells with a high dose of
FF/CAP18 (Fig. 6); uric acid is
the final product of purine metabolism, and these data indicate
that treatment of HCT116 cells with a high dose of FF/CAP18
facilitates the degradation of purine metabolites.
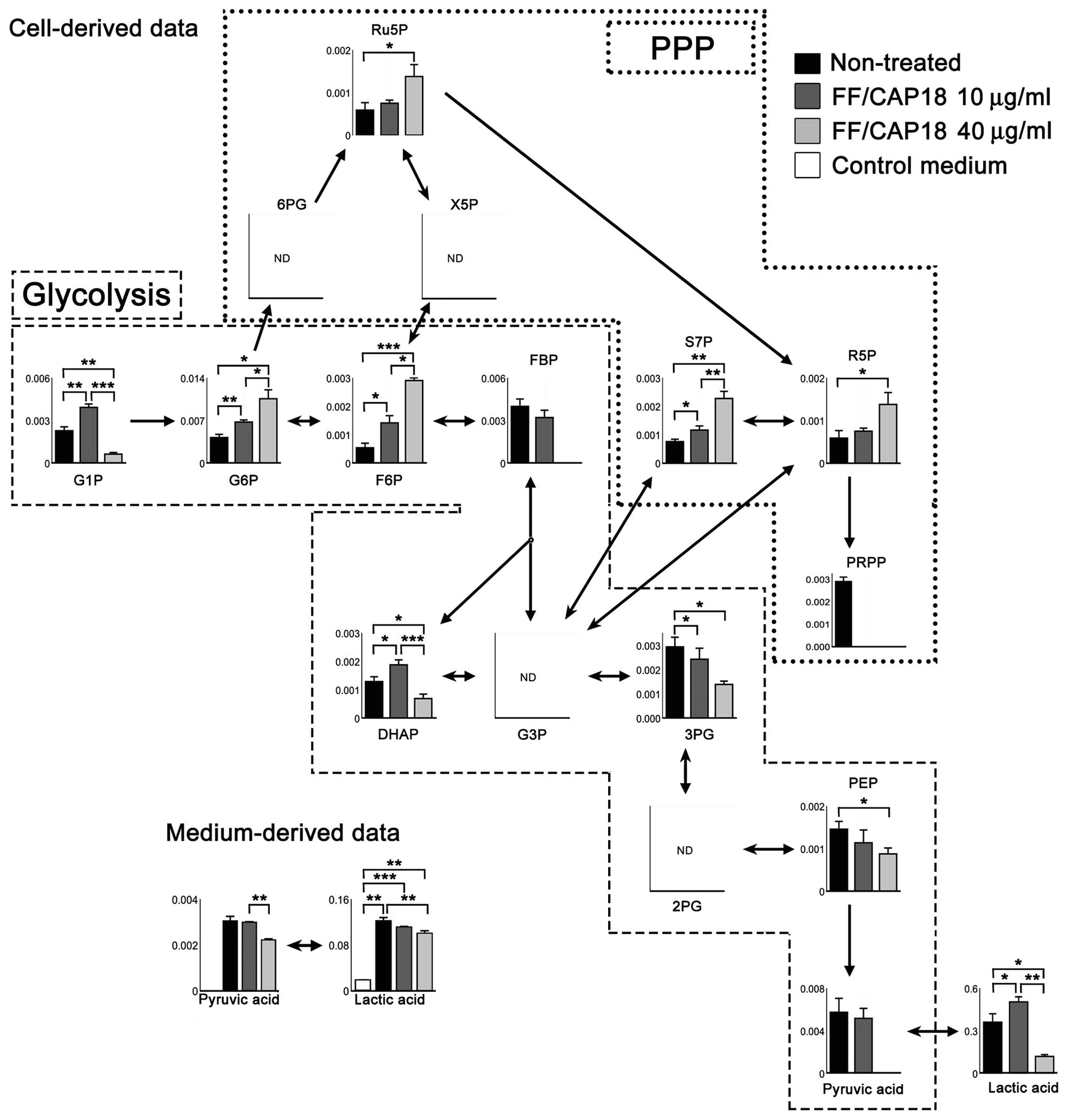
Glycolysis and the pentose phosphate
pathway (PPP)
Glycolysis is the key pathway for cancer cells to
generate the energy that is required to support rapid cell division
and cancer progression. We mapped the glycolysis pathway and a
metabolic pathway branching from glycolysis, the PPP, as shown in
Fig. 7. The levels of glucose
6-phosphate (G6P) and frucutose 6-phosphate (F6P) were increased
after treatment with FF/CAP18, in a dose-dependent manner. The
level of fructose 1,6-bisphosphate (FBP), however, tended to be
lower in cells treated with 10 μg/ml FF/CAP18 than in non-treated
cells, and FBP was not detectable in cells treated with FF/CAP18 at
40 μg/ml. Downstream metabolites, such as 3-phosphoglycerate,
phosphoenolpyruvate, pyruvic acid and lactic acid similarly showed
downregulation. Pyruvic acid and lactic acid in conditioned medium
also showed a slight decline in level in HCT116 cells treated with
FF/CAP18 (Fig. 7).
The PPP metabolites ribulose 5-phosphate (Ru5P),
ribose 5-phosphate (R5P), and sedoheptose 7-phosphate (S7P) were
upregulated in a dose-dependent manner, whereas 6-phosphogluconate
and xylurose 5-phosphate were not detected. These data indicate
that glycolysis in HCT116 cells was suppressed downstream of FBP in
the presence of FF/CAP18, and the cells had shifted their
metabolism towards the PPP.
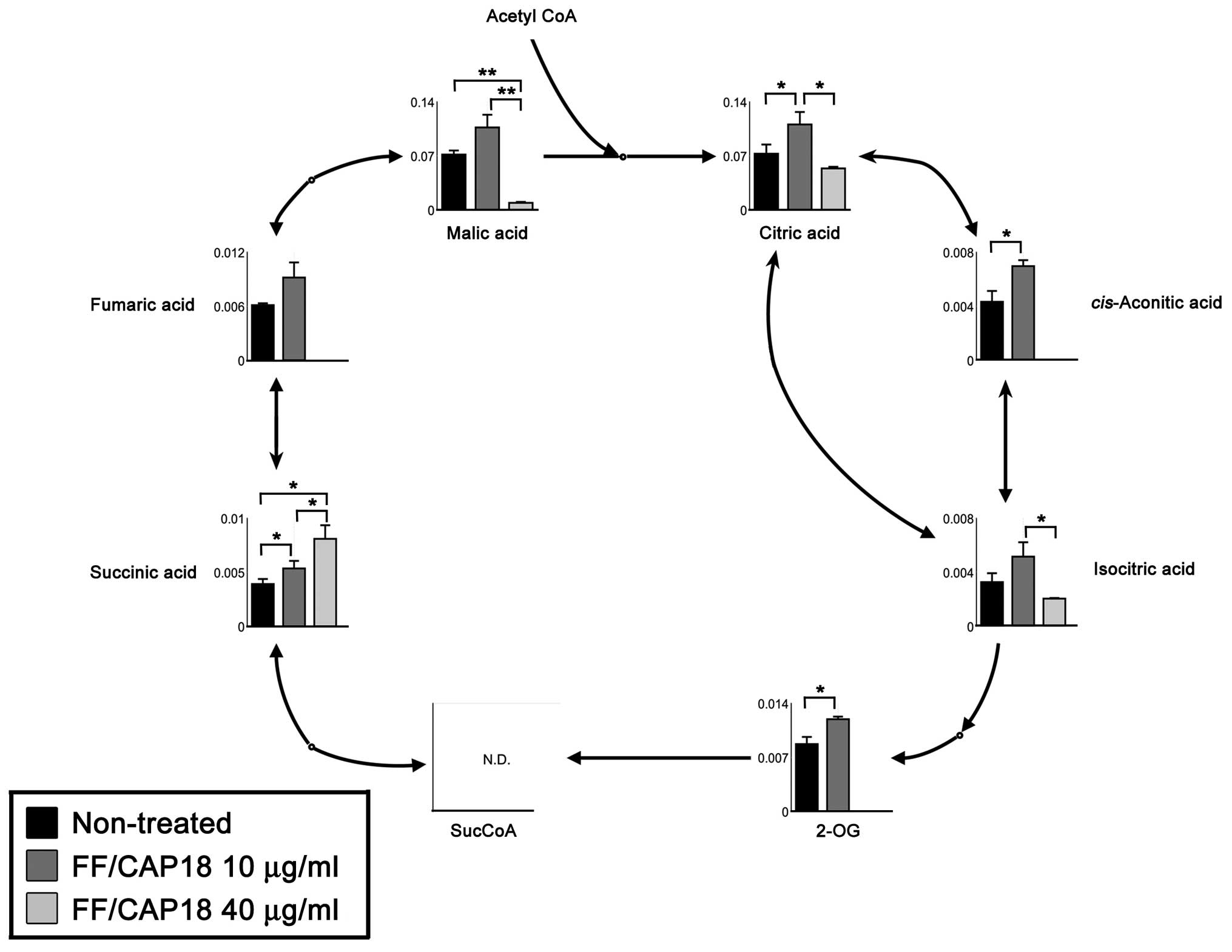
TCA cycle
We observed significant differences in the levels of
metabolites of the TCA cycle. While all metabolites detected in
this study were upregulated in HCT116 cells treated with 10 μg/ml
FF/CAP18, most metabolites of the TCA cycle were downregulated,
with the exception of succinic acid, and then only in HCT116 cells
treated with 40 μg/ml FF/CAP18 (Fig.
8); we could not confirm significant differences between HCT116
cells treated with FF/CAP18 at 10 and 40 μg/ml in the data for
conditioned medium (Fig. 3).
Therefore, the TCA cycle in HCT116 cells was likely to be
facilitated by the administration of FF/CAP18 at 10 μg/ml. On the
other hand, inactivation of the TCA cycle was induced through the
accumulation of succinic acid.
Discussion
Several studies have revealed anticancer activity of
antimicrobial peptides, as demonstrated by oncolytic properties
against many cancer cell types (26). However, the impact of antimicrobial
peptides on cell metabolism is poorly understood, although
metabolic reprogramming is a hallmark of cancer cells. Our study
demonstrates large-scale metabolic profiling of colon cancer cells
after treatment with an antimicrobial peptide, FF/CAP18, and
suggests that the apoptotic cell death of colon cancer cells
induced by FF/CAP18 is a result of dynamic levels of
metabolites.
In the present study, PCA and HCA of large-scale
metabolic profiling demonstrated remarkable differences between
non-treated HCT116 cells, cells treated with FF/CAP18 at 10 μg/ml,
and cells treated with 40 μg/ml FF/CAP18 (Figs. 4 and 5). A great number of metabolites,
including nucleotides, TCA cycle components, and amino acids in
HCT116 cells, increased in HCT116 cells after treatment with
FF/CAP18 at 10 μg/ml compared with non-treated cells, indicating
that FF/CAP18 could induce favorable metabolic conditions at this
comparatively low concentration. On the other hand, at a
concentration of 40 μg/ml, FF/CAP18 caused shortages of a number of
metabolites. In previous studies, including our own, the
cytotoxicity of various antimicrobial peptides towards cancer cells
was reported to be dose-dependent (18,19,27).
Thus, the results of the present study indicate that the levels of
metabolites in cells undergoing apoptotic cell death caused by
antimicrobial peptides do not show unidirectional movement in the
same way as the concentration-dependent effects previously
reported.
ATP generation occurs via glycolysis rather than
oxidative phosphorylation in cancer cells; defined as the Warburg
effect, this is a well-established metabolic characteristic of
cancer cells. We observed upregulation of the first three
intermediates in the glycolysis pathway [glucose 1-phosphate (G1P),
G6P, and F6P], or of only the second two intermediates (G6P and
F6P), after treatment of FF/CAP18 at 10 or 40 μg/ml, respectively
(Fig. 7). However, as the levels
of intermediates downstream of FBP were not increased in this
dose-dependent manner, conversion of F6P to FBP is the
rate-limiting step of glycolysis, and ATP generation via glycolysis
is suppressed by administration of FF/CAP18. When apoptosis is
induced in cells, their metabolism is regulated by various factors,
such as the TP53-inducible glycolysis and apoptosis regulator,
which blocks glycolysis at the stage of conversion of F6P to FBP
and promotes activity of the PPP (28). G1P and fructose 1-phosphate are
used to synthesize R5P, an intermediate of the PPP, as an
alternative route for glucose metabolism. In this study, the levels
of PPP intermediates (Ru5P, R5P, and S7P) were increased in
FF/CAP18-treated HCT116 cells compared with non-treated cells
(Fig. 7). Therefore, an
antimicrobial peptide, FF/CAP18, can shift glucose metabolism
towards the PPP and reduce the Warburg effect, resulting in slow
glycolysis.
Interestingly, intermediates in the TCA cycle were
increased by administration of FF/CAP18 at 10 μg/ml (Fig. 8), indicating that HCT116 cells
acquire ATP via mitochondrial respiration, not glycolysis, in the
early stage of apoptosis caused by FF/CAP18 treatment. On the other
hand, most of the TCA cycle intermediates in HCT116 cells treated
with FF/CAP18 at 40 μg/ml were downregulated (Fig. 8). This inverse relationship with
the concentration of FF/CAP18 occurred in connection with the
progression of apoptosis in treated HCT116 cells. Mitochondrial
depolarization is a major event in the progression of apoptosis,
and we confirmed that treatment with FF/CAP18 at 40 μg/ml induced
this process in HCT116 cells in a previous study (19), suggesting that down-regulation of
intermediates in the TCA cycle is potentially caused by this
alteration in mitochondrial function.
Apoptotic cell death requires ATP for the
progression of several steps, such as caspase activation, enzymatic
hydrolysis of macromolecules (29,30),
chromatin condensation (31), bleb
formation (32), and the formation
of apoptotic bodies (33).
Zamaraeva et al suggested that elevation of the ATP level is
a prerequisite for the apoptotic cell death process (34). FF/CAP18 treatment at 10 μg/ml could
trigger apoptosis of HCT116 cells via upregulation of ATP generated
by oxidative phosphorylation rather than glycolysis. Moreover,
increasing the FF/CAP18 concentration to 40 μg/ml induced
late-stage apoptosis that was accompanied by a reduction of the ATP
level via suppression of glycolysis and the TCA cycle and resulted
in conditions wherein most metabolites were depleted.
Owing to screening and/or improved treatment,
colorectal cancer mortality rates have been observed to be
decreasing in a large number of countries (35). However, the increasing prevalence
of obesity and decreasing levels of physical activity in many parts
of the world continue to contribute to the incidence of colorectal
cancer. In addition, sensitivity to chemotherapy and severe side
effects remain unresolved issues. Therefore, the discovery of novel
therapeutic strategies for colorectal cancer is the focus of
intense research efforts.
Antimicrobial peptides are an essential component of
the innate immunity of many organisms, and target a wide-range of
infectious disease agents, such as fungi, protozoa (36), the human immunodeficiency virus and
herpes viruses (37). Whereas
antimicrobial peptides have been studied as antimicrobial agents,
their potential as anticancer peptides in cancer therapy, either
alone or in combination with other conventional drugs, has been
regarded as a therapeutic strategy yet to be explored (26). Indeed, several recent studies have
suggested that antimicrobial peptides possibly exert cytotoxic
effects against colon cancer via apoptotic death (17,19,38).
Thus, our study, approaching this issue from the point of view of
metabolic changes, can support an understanding of the mechanisms
of anticancer agents such as antimicrobial peptides, which can in
turn facilitate the discovery of novel remedies or therapeutic
strategies for cancer.
Acknowledgements
We thank Dr Kenjiro Kami and Satsuki Yamaryo (Human
Metabolome Technologies Inc., Tsuruoka, Japan) for their technical
advice.
References
|
1
|
The global burden of disease: 2004 update.
WHO Library Cataloguing-in-Publication Data. 2008, http://www.who.int/healthinfo/global_burden_disease/2004_report_update/en/.
|
|
2
|
Cancer statistics: Worldwide. World Cancer
Research Fund International Web site. http://www.wcrf.org/int/cancer-facts-figures/worldwide-data.
|
|
3
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Phan LM, Yeung SC and Lee MH: Cancer
metabolic reprogramming: importance, main features, and potentials
for precise targeted anti-cancer therapies. Cancer Biol Med.
11:1–19. 2014.PubMed/NCBI
|
|
5
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hirayama A, Kami K, Sugimoto M, et al:
Quantitative metabolome profiling of colon and stomach cancer
microenvironment by capillary electrophoresis time-of-flight mass
spectrometry. Cancer Res. 69:4918–4925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Soga T, Baran R, Suematsu M, et al:
Differential metabolomics reveals ophthalmic acid as an oxidative
stress biomarker indicating hepatic glutathione consumption. J Biol
Chem. 281:16768–16776. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saito K: Phytochemical genomics - a new
trend. Curr Opin Plant Biol. 16:373–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mashego MR, Rumbold K, De Mey M, Vandamme
E, Soetaert W and Heijnen JJ: Microbial metabolomics: past, present
and future methodologies. Biotechnol Lett. 29:1–16. 2007.
View Article : Google Scholar
|
|
10
|
Spratlin JL, Serkova NJ and Eckhardt SG:
Clinical applications of metabolomics in oncology: a review. Clin
Cancer Res. 15:431–440. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oliver S: Guilt-by-association goes
global. Nature. 403:601–603. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fiehn O: Metabolomics - the link between
genotypes and phenotypes. Plant Mol Biol. 48:155–171. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu WK, Wang G, Coffelt SB, et al: Emerging
roles of the host defense peptide LL-37 in human cancer and its
potential therapeutic applications. Int J Cancer. 127:1741–1747.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zaiou M and Gallo RL: Cathelicidins,
essential gene-encoded mammalian antibiotics. J Mol Med (Berl).
80:549–561. 2002. View Article : Google Scholar
|
|
15
|
Zanetti M: The role of cathelicidins in
the innate host defenses of mammals. Curr Issues Mol Biol.
7:179–196. 2005.PubMed/NCBI
|
|
16
|
Bals R, Wang X, Zasloff M and Wilson JM:
The peptide antibiotic LL-37/hCAP-18 is expressed in epithelia of
the human lung where it has broad antimicrobial activity at the
airway surface. Proc Natl Acad Sci USA. 95:9541–9546. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren SX, Cheng AS, To KF, et al: Host
immune defense peptide LL-37 activates caspase-independent
apoptosis and suppresses colon cancer. Cancer Res. 72:6512–6523.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Okumura K, Itoh A, Isogai E, et al:
C-terminal domain of human CAP18 antimicrobial peptide induces
apoptosis in oral squamous cell carcinoma SAS-H1 cells. Cancer
Lett. 212:185–194. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kuroda K, Fukuda T, Yoneyama H, et al:
Anti-proliferative effect of an analogue of the LL-37 peptide in
the colon cancer derived cell line HCT116 p53+/+ and
p53. Oncol Rep. 28:829–834. 2012.PubMed/NCBI
|
|
20
|
Isogai E, Isogai H, Matuo K, et al:
Sensitivity of genera Porphyromonas and Prevotella to the
bactericidal action of C-terminal domain of human CAP18 and its
analogues. Oral Microbiol Immunol. 18:329–332. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Soga T and Heiger DN: Amino acid analysis
by capillary electrophoresis electrospray ionization mass
spectrometry. Anal Chem. 72:1236–1241. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Soga T, Ueno Y, Naraoka H, Ohashi Y,
Tomita M and Nishioka T: Simultaneous determination of anionic
intermediates for Bacillus subtilis metabolic pathways by capillary
electrophoresis electrospray ionization mass spectrometry. Anal
Chem. 74:2233–2239. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Soga T, Ohashi Y, Ueno Y, Naraoka H,
Tomita M and Nishioka T: Quantitative metabolome analysis using
capillary electrophoresis mass spectrometry. J Proteome Res.
2:488–494. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sugimoto M, Wong DT, Hirayama A, Soga T
and Tomita M: Capillary electrophoresis mass spectrometry-based
saliva metabolomics identified oral, breast and pancreatic
cancer-specific profiles. Metabolomics. 6:78–95. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Junker BH, Klukas C and Schreiber F:
VANTED: a system for advanced data analysis and visualization in
the context of biological networks. BMC Bioinformatics. 7:1092006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gaspar D, Veiga AS and Castanho MA: From
antimicrobial to anticancer peptides. A review. Front Microbiol.
4:2942013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu D, Gao Y, Qi Y, Chen L, Ma Y and Li Y:
Peptide-based cancer therapy: opportunity and challenge. Cancer
Lett. 351:13–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bensaad K, Tsuruta A, Selak MA, et al:
TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell.
126:107–120. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu Y, Benedict MA, Ding L and Nunez G:
Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated
caspase-9 activation and apoptosis. EMBO J. 18:3586–3595. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li P, Nijhawan D, Budihardjo I, et al:
Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9
complex initiates an apoptotic protease cascade. Cell. 91:479–489.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kass GE, Eriksson JE, Weis M, Orrenius S
and Chow SC: Chromatin condensation during apoptosis requires ATP.
Biochem J. 318:749–752. 1996.PubMed/NCBI
|
|
32
|
Barros LF, Kanaseki T, Sabirov R, et al:
Apoptotic and necrotic blebs in epithelial cells display similar
neck diameters but different kinase dependency. Cell Death Differ.
10:687–697. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Leist M, Single B, Castoldi AF, Kuhnle S
and Nicotera P: Intracellular adenosine triphosphate (ATP)
concentration: a switch in the decision between apoptosis and
necrosis. J Exp Med. 185:1481–1486. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zamaraeva MV, Sabirov RZ, Maeno E,
Ando-Akatsuka Y, Bessonova SV and Okada Y: Cells die with increased
cytosolic ATP during apoptosis: a bioluminescence study with
intracel-lular luciferase. Cell Death Differ. 12:1390–1397. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Center MM, Jemal A, Smith RA and Ward E:
Worldwide variations in colorectal cancer. CA Cancer J Clin.
59:366–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gentilucci L, Tolomelli A and Squassabia
F: Peptides and pepti-domimetics in medicine, surgery and
biotechnology. Curr Med Chem. 13:2449–2466. 2006. View Article : Google Scholar
|
|
37
|
Hancock RE and Diamond G: The role of
cationic antimicrobial peptides in innate host defences. Trends
Microbiol. 8:402–410. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ren SX, Shen J, Cheng AS, et al: FK-16
derived from the anti-cancer peptide LL-37 induces
caspase-independent apoptosis and autophagic cell death in colon
cancer cells. PLoS One. 8:e636412013. View Article : Google Scholar
|