Introduction
Malignant melanoma is the leading cause of skin
cancer-associated mortality, the median survival time of patients
with stage IV melanoma is less than 1 year and 5-year survival rate
is less than 10% (1). Incidence
and mortality rates of melanoma have increased steadily during the
past four decades (2,3). Patients with early stages of melanoma
are curable with surgery. However, once metastasis is established,
it is difficult to treat and the mortality rate is high due to the
lack of effective treatment. Despite significant improvement in
immunotherapy and targeted therapy in recent years, only a section
of patients with metastatic melanoma benefit for a short of period
of response because of drug resistance and high toxicity (4). Hence, identification of potential new
target drugs and development of effective therapeutic strategies
are the utmost importance to reduce melanoma-related mortality.
NF-κB represents a family of inducible transcription
factors involved in the maintenance of various cellular functions,
such as cell cycle progression, apoptosis, inflammatory and immune
responses (5). In cancer cells,
NF-κB is proposed to play an important role in tumorigenesis,
promoting cell proliferation, migration, angiogenesis and
anti-apoptotic effects. The regulation of NF-κB activation is
determined by its subcellular localization. In unstimulated cells,
NF-κB formed a complex in the cytoplasm with its inhibitor IκB.
Cell stimulation accompanied by IκB phosphorylated proteins and
ubiquitinated, thus liberating NF-κB from IκB complexes (6). The active NF-κB translocates into the
nucleus and activates the expression of specific κB enhancer gene
(5). Aberrant activation of NF-κB
has been reported in various types of cancer, including pancreatic,
prostate and colon cancer (7,8).
NF-κB is found constitutively activated in human melanoma cells,
and upregulation of the NF-κB levels promotes the progression of
melanoma and increases metastatic potential (9,10).
Inhibition of NF-κB activation in human melanoma cells enhanced
radio-sensitivity, induced apoptosis and inhibited invasion
(11,12).
Protein glycosylation plays an important role in
pathophysiological steps of tumor progression, including tumor
proliferation, invasion, metastasis and angiogenesis (13). Fucosylation is one of the important
steps in protein glycosylation. Fucosyltransferases (FUTs) are the
key enzymes catalyzing the synthesis of fucosylated glycans. At
least eight of 1, 3/4-FUT genes have been identified, they are
FUT3, 4, 5, 6, 7, 9, 10 and 11 (14,15).
Among these FUTs, FUT4 catalyzes the synthesis of α1,
3-fucosylation of Lewis Y. Several reports have shown that FUT4 is
overexpressed in many types of cancer, including colorectal,
gastric and lung cancer (16–18).
Furthermore, increased FUT4 expression has been implicated to
promote cell proliferation, metastasis and anti-apoptosis (19–21).
However, whether FUT4 is increased in melanomas, and whether FUT4
and NF-κB signaling pathway is involved in 20 (R)-Ginsenoside
Rg3-induced melanoma cell apoptosis are largely unknown.
Ginseng, is a well-known herbal medicine, used for
thousands of years alone or in combination with other herbal
ingredients, such as invigorant, cardiotonic, and drugs for
anti-inflammatory, antitumor and immune stimulation. Rg3 is a
monomer extracted from ginseng and has a broad spectrum of
antitumor activities. Rg3 inhibits cell proliferation and induces
apoptosis in colon and gastric cancer (22,23).
In addition, Rg3 enhances the chemosensitivity of patients to
docetaxel and cisplatin with prostate and colon cancer (24,25).
Moreover, Rg3 has been shown to inhibit tumor angiogenesis and
induces cancer cell apoptosis in liver carcinomas (26). However, the antitumor mechanism of
Rg3, FUT4 and NF-κB pathway on human melanoma remains unclear.
In the present study, we demonstrated, both in
vitro and in vivo, that Rg3 suppressed FUT4 expression
by inhibiting NF-κB signaling pathway, by which it induced melanoma
apoptosis. In conclusion, our results suggest that Rg3 deactivates
NF-κB signaling pathway to downregulate FUT4 playing an important
role in inhibiting melanoma progression.
Materials and methods
Ethics statement
All animal work performed in the present study was
approved by the Animal Ethics Committee of the Dalian Medical
University. The detail protocols and experimental processes
conformed to the Experimental Animal Management Regulations of
Dalian Medical University.
Antibodies and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), TRIzol and Lipofectamine™ 2000 reagents were
purchased from Invitrogen (Camarillo, CA, USA). Rg3 was provided by
Dalian Fusheng Pharmaceutical Co., Ltd. (Dalian, China). It was
diluted with cell culture media to final concentration. Cell
Counting kit-8 (CCK-8) was purchased from Dojindo Laboratories
(Kumamoto, Japan). Annexin V/FITC kit was purchased from Nanjing
KeyGen Biotech Co., Ltd. (Nanjing, Jiangsu, China). Antibodies
specific for PARP, caspase-3, -8, -9, Bcl-2, survivin, Bax, FUT4,
β-actin, HRP-conjugated anti-rabbit and anti-mouse antibodies were
purchased from Proteintech Group (Wuhan, China). Antibodies
specific for p-p65, p65, pIκBα, IκBα, pIKKα/β and IKKα/β were
purchased from Cell Signaling Technology (Boston, MA, USA). p65
siRNA was purchased from the Shanghai GenePharma Co. (Shanghai,
China). NF-κB inhibitor (Bay 11-7082) was purchased from Selleck
Chemicals (Houston, TX, USA). Nuclear extract kit and EMSA kit were
purchased from Thermo Fisher Scientific (Waltham, MA, USA). ChIP
kit was purchased from Millipore (Billerica, MA, USA). The enhanced
chemiluminescence (ECL) assay kit was purchased from Amersham
Biosciences (Pittsburgh, PA, USA).
Cell culture
Human melanoma cell lines A375P, A375M, C8161, Mevo
and SK-MEL-28 were gifted from Dr Mary Hendrix (Stanley Manne
Children’s Research Institute Northwestern University’s Feinberg
School of Medicine, Chicago, IL, USA) and were grown in DMEM with
10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin; maintained
at 37°C under 5% CO2 in humidified air.
Cell viability assay
Melanoma cells were plated at a density of 2,000
cells/well in 96-well plates, and treated with various
concentrations of Rg3 for 24 h the following day. Cell viability
was evaluated in cells using CCK-8 kit according to the
manufacturer’s instructions. Briefly, 10 μl of the CCK-8 solution
was added to cell cultures for the designated times. Plates were
incubated for 1 h with CCK-8 solution at 37°C. The optical density
(OD) was read at 450 nm absorbance on a microplate reader (Bio-Rad
Laboratories, Hercules, CA, USA).
Colony forming assay
Cells (1×103 cells/well) were plated in
6-well plates containing DMEM with 10% FBS at 37°C. After 24 h,
cells were treated with different concentrations of Rg3 (0, 25, 50,
75 and 100 μg/ml) for 24 h and then cells were allowed to grow for
10 days in the absence of Rg3. Cells were fixed and stained with
crystal violet (0.5%) for 20 min at room temperature. Images were
captured with the inverted microscope (Olympus IX71; Olympus,
Tokyo, Japan).
Apoptosis assay
Apoptosis was assessed by Annexin V-binding analysis
of flow cytometry. For flow cytometric analysis, cells
(1×105) were seeded in 6-cm dishes overnight before
treated with various concentrations of Rg3 for 24 h. Both adherent
and floating cells were harvested and combined, washed twice with
PBS, resuspended in 500 μl of binding buffer, and stained using an
Annexin V-FITC/PI kit according to the manufacturer’s instructions.
After incubation in the dark for 30 min, the cells were analyzed
using FACScalibur instrument (FACSCalibur; BD Biosciences, San
Jose, CA, USA). All experiments were performed in duplicate and
reproducibility was checked in three independent experiments.
Quantitative real-time PCR analysis
Total RNA was extracted using TRIzol (Invitrogen)
according to the manufacturer’s protocol. RNA was reverse
transcribed into cDNA using PrimeScript™ RT reagent kit (Takara,
Tokyo, Japan). The FUT4 primers were 5′-AAGGTCCAGGCCCACTGAAG-3′
(forward) and 5′-CAGTTCAGGTGACAGAGGCTCAA-3′ (reverse); the GAPDH
primers were 5′-ATGGGGAAGGTGAAGGTCG-3′ (forward) and
5′-GGGGTCATTGATGGCAACAATA-3′ (reverse). Real-time quantitative PCR
reactions were performed with Applied Biosystems StepOne Real-time
PCR system (Life Technologies, Carlsbad, CA, USA). Relative FUT4
mRNA levels were normalized with GAPDH and calculated using
2−ΔΔCT method.
Western blot analysis
Cells were washed with PBS (pH 7.4), and incubated
with 2× concentrated electrophoresis sample buffer (125 mM
Tris-HCl, pH 6.8, 5% glycerol, 2% SDS, 1% β-mercaptoethanol) for 30
min on ice. Protein concentration was determined with Coomassie
protein assay reagent using bovine serum albumin as a standard.
Total protein (50–70 μg) from the whole cell lysates was separated
by 12% SDS-PAGE and proteins separated in the gel were transferred
electrophoretically onto nitrocellulose membrane (Millipore,
Billerica, MA, USA), incubated with TTBS (50 mM Tris HCl, pH 7.5,
0.15 M NaCl, 0.1% Tween-20) containing 5% fat-free dry milk for 2 h
followed by overnight incubation with the appropriate primary
antibodies at the dilutions recommended by the suppliers at 4°C.
After incubation with an HRP-conjugated appropriate secondary
antibody, ECL (enhanced chemiluminescence) detection system
(Bio-Rad Laboratories) was used to visualize immunoreactive
bands.
Electrophoretic mobility shift assay
(EMSA)
Cells were treated with Rg3 (75 μg/ml) for 24 h. The
DNA binding activities of NF-κB in nuclear extracts were assessed
using the EMSA kit with biotin-labeled double-stranded NF-κB
oligonucleotides (Beyotime, Nantong, China). The sequences of the
oligonucleotides adopted were as follows:
5′-CGCTTGATGACTCAGCCGGAA-3′ and 3′-GCGAACTACTGAGTCGG CCTT-5′.
Briefly, nuclear extracts (8 μg/sample) were incubated with the
oligonucleotides in reaction buffer for 20 min. Protein DNA
complexes were separated on a 6.5% non-denaturing acrylamide gel,
transferred to positively charged nylon membranes, and immobilized
by UV cross-linking for 10 min. Band shifts were detected by
chemiluminescence method with a detection system (Bio-Rad
Laboratories).
Luciferase reporter assay
The NF-κB luciferase activity was determined in
A375P and C8161 cells after co-transfected, using Lipofectamine
2000 reagent, with 2 μg of NF-κB luciferase plasmids and 0.2 μg of
pGL3, which constitutively expressed Renilla luciferase.
Twenty-four hours after transfection, the cells were treated with
Rg3 (50 or 75 μg/ml as indicated) for 24 h. The luciferase activity
was assayed using the Dual Luciferase reporter assay system
Berthold Technologies (Bad Wildbad, Germany). Firefly luciferase
activity was measured and the reading was normalized to
Renilla luciferase activity, which served as an internal
control for transfection efficiency.
Chromatin immunoprecipitation
The chromatin immunoprecipitation (ChIP) assay was
performed according to the manufacturer’s instruction using cells
at ~80% confluent post Rg3 treatment for 24 h. In brief, 1%
paraformaldehyde was added to the cell-culture medium and incubated
at room temperature for 10 min to cross-link. The cells were then
washed twice in cold phosphate-buffered saline, scraped and lysed
in SDS lysis buffer containing protease inhibitor cocktail for 10
min at 4°C. The lysates were sonicated five times for 15 sec each
time, and the debris was removed by centrifugation. A total of 10
μl of the supernatant was used as an input sample, and the
remaining of the lysate were diluted 10-fold with a dilution buffer
containing protease inhibitor cocktail followed by incubation with
antibodies against specific transactivators or a non-immune rabbit
IgG control overnight at 4°C. Immunoprecipitated complexes were
collected using protein A/G plus agarose beads. The precipitates
were extensively washed and incubated in an elution buffer at room
temperature for 20 min. Cross-linking of protein-DNA complexes were
reversed at 65°C for 5 h, followed by treatment with 100 mg/ml
Proteinase K for 2 h at 45°C. DNA was extracted three times with
phenol/chloroform and precipitated with ethanol. The pellets were
resuspended in TE buffer and subjected to PCR amplification using
specific FUT4 promoter primers (forward primer,
CCATTCCCAGCACTGTCTATTTC and reverse primer,
CCTACGGGTTGAATTTGAATTTCT. The resulting product of FUT4 was
separated by 1.0% agarose gel electrophoresis.
Xenograft tumor mouse models
Male nude mice (Balb/c-nu/nu, 4–6 weeks old) were
obtained from the Animal Center (Dalian Medical University) and
maintained under sterile conditions during the entire experimental
period. Mice were divided into two groups (n=8/group) based on the
treatment. A375P cells (2×106) suspended in 0.2 ml PBS
were injected subcutaneously into the right flank. After 7 days of
tumor inoculation, mice were treated with Rg3 (20 mg/kg of body
weight) or its vehicle control subcutaneously for 3 weeks with time
interval of 48 h. Tumor volume was measured by Vernier calipers
every other day after tumor was palpable. The tumor volume was
calculated according to the formula (volume=1/2 length ×
width2). At the end of the experiment (at day 30), the
tumor was removed and weighed.
Statistical analysis
Each experiment was repeated 3 times and results are
presented as the mean ± SEM. Unpaired Student’s t-tests were used
to analyze differences between groups. P<0.05 was considered to
be significant. The statistical software SPSS ver. 17 was used for
analyzing the data.
Results
Rg3 inhibits human melanoma cell
proliferation
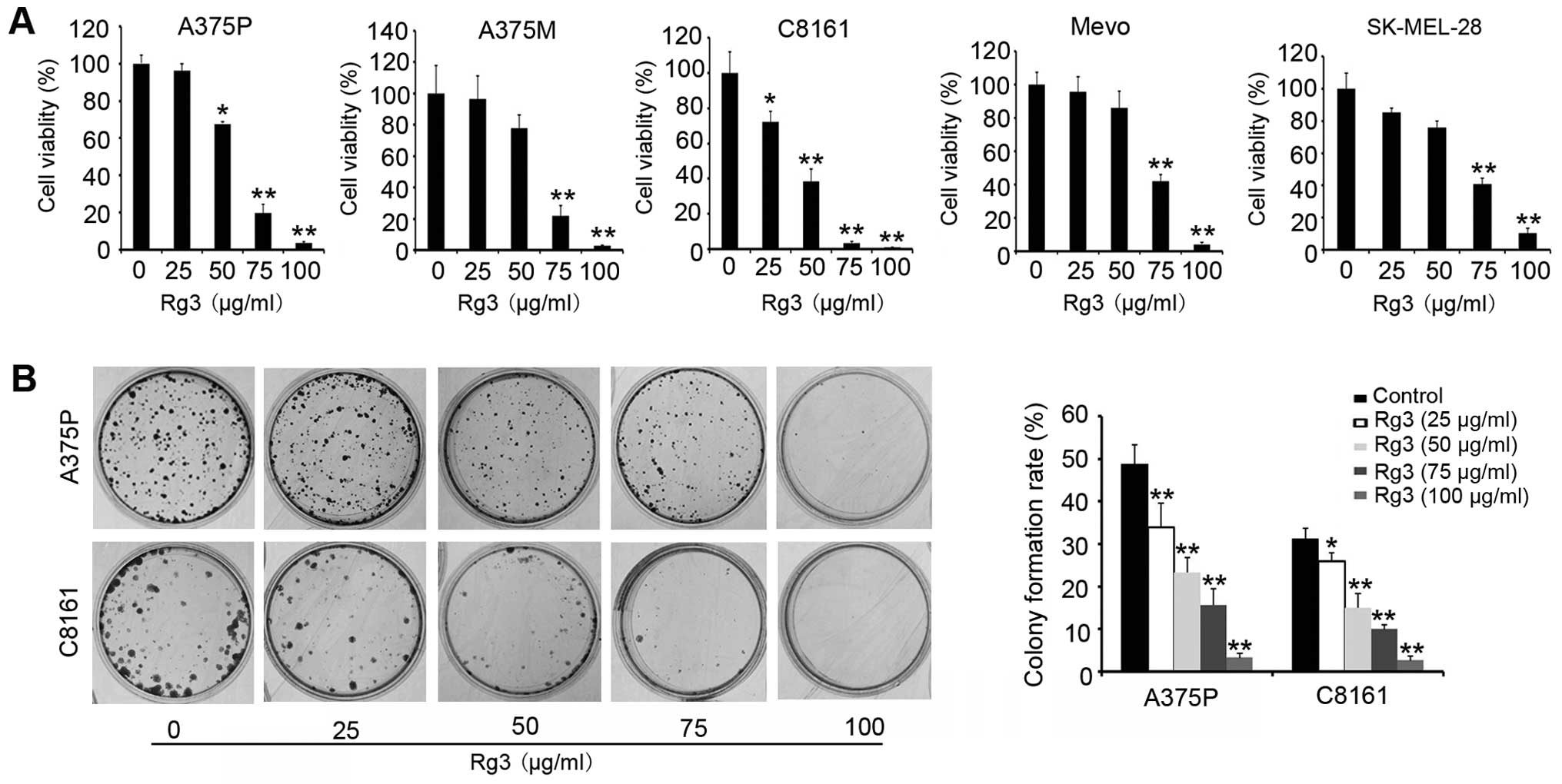
To evaluate the effect of Rg3 on melanoma cell
proliferation, we first treated various melanoma cells with
different concentrations of Rg3 (0, 25, 50, 75 and 100 μg/ml) for
24 h. CCK-8 proliferation assay showed that Rg3 treatment
significantly inhibited cell proliferation/viability in all cell
lines tested (Fig. 1A). The
IC50 values are in the range of 57–83 μg/ml. The
sensitivity of the cell responses to Rg3 was
C8161>A375P>A375M>SKMEL-28>Mevo. We selected the most
sensitive cell lines, A375P and C8161, for further study. Colony
forming assay revealed that Rg3 treatment led to a significant
decrease in the number of colonies (P<0.05) (Fig. 1B). The effect of Rg3 on cell growth
was dose-dependent. These results indicate that Rg3 has a cytotoxic
effect on melanoma cells and inhibits cell growth.
Rg3 induces human melanoma cell
apoptosis
We investigated whether the observed Rg3 antitumor
effect in human melanoma cells correlates with its effect on cell
apoptosis. Human melanoma cell lines A375P and C8161 were treated
with different concentrations of Rg3 for 24 h, followed by flow
cytometric analysis after staining with Annexin V-FITC/PI. The
results showed that treatment with Rg3 (0, 25, 50, 75 and 100
μg/ml) for 24 h led to concentration-dependent increases in
apoptotic cells in both A375P and C8161 cells, from 3.28 and 4.41%
in untreated control groups to 3.98 and 6.23% (25 μg/ml), 4.24 and
12.89% (50 μg/ml), 24.04 and 66.05% (75 μg/ml), and 57.75 and
86.51% (100 μg/ml), respectively (Fig.
2A). We further examined the expression of apoptotic marker
proteins. As shown in Fig. 2B, Rg3
treatment effectively activated the expression of pro-apoptotic
proteins (caspase-3, -8, -9 and Bax), decreased the expression of
anti-apoptosis proteins (Bcl-2 and survivin) in a dose-dependent
manner (Fig. 2B). In addition, Rg3
also activated PARP, a marker of apoptosis (Fig. 2B, top row). These results indicate
that Rg3 induces human melanoma cells apoptosis.
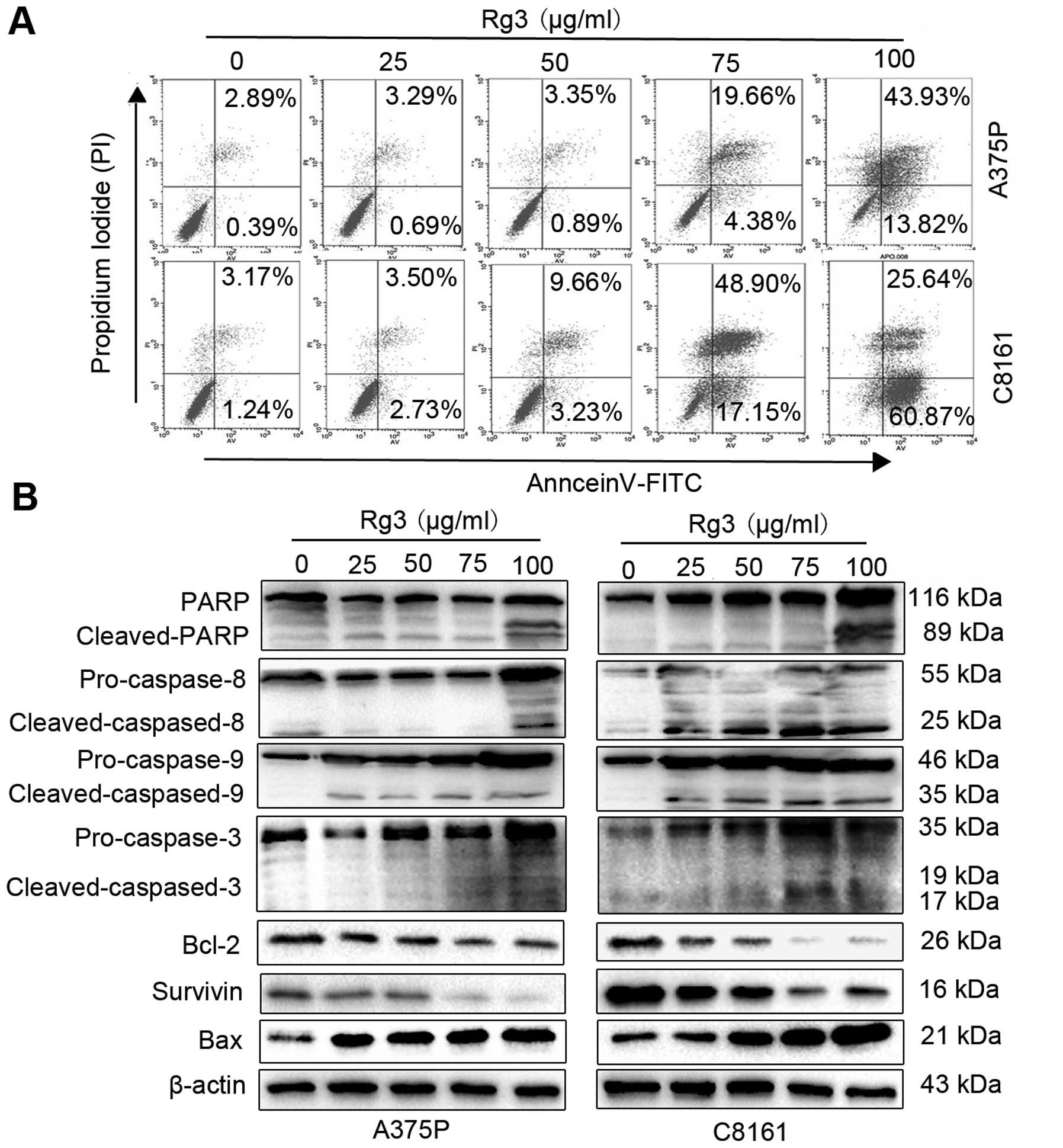 | Figure 2Rg3 induces human melanoma cell
apoptosis. (A) A375P and C8161 cells were double stained with
Annexin V/PI and analyzed by FACS analysis after 24-h treatment
with Rg3 (0, 25, 50, 75 and 100 μg/ml). The percentage of Annexin
V+/PI− (early apoptotic cells, lower right),
Annexin V+/PI+ (late apoptotic cells, upper
right), Annexin V−/PI− (viable cells, lower
left) and Annexin V−/PI+ (necrotic cells,
upper left), cells is shown. (B) A375P and C8161 cells were treated
with Rg3 (0, 25, 50, 75 and 100 μg/ml) for 24 h, the expression
levels of PARP, caspase-8, 9, 3, Bcl-2, survivin and Bax were
analyzed by western blot analysis. |
Rg3 inhibits the activation of the NF-κB
signaling pathway
Previous studies have demonstrated that NF-κB plays
a critical role in apoptosis and cell survival, it is possible that
NF-κB pathway might also be involved in Rg3-induced apoptosis in
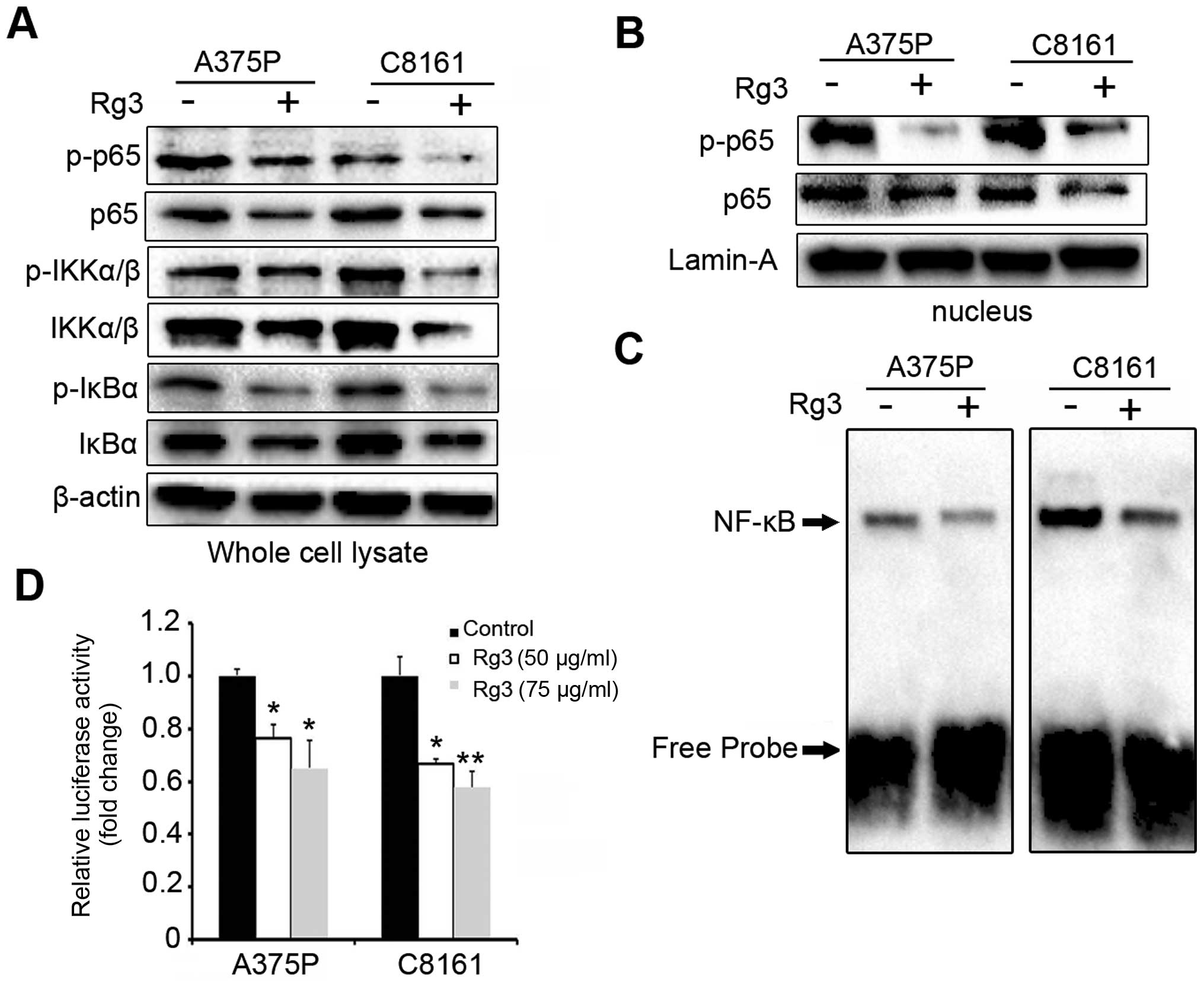
human melanoma cells. As shown in Fig.
3A, treatment with Rg3 (75 μg/ml) for 24 h led to a significant
decrease in the phosphorylation of NF-κB/p65, IκBα and IKKα/β in
both A375P and C8161 cells. Similarly, we also observed a decrease
in the basal level of NF-κB/p65, IκBα and IKKα/β after Rg3
treatment (Fig. 3A). Moreover, Rg3
decreased the nuclear level of NF-κB/p65 and its phosphorylation
(Fig. 3B), suggesting Rg3 also
inhibited the nuclear translocation of NF-κB/p65. Subsequently, we
investigated the effect of Rg3 on DNA-binding activity of
NF-κB/p65. By EMSA assays, we found dramatically reduced NF-κB DNA
binding in Rg3 (75 μg/ml) treated cells (Fig. 3C). To further investigate the
effects of Rg3 on NF-κB activity, NF-κB-Luc was transfected into
A375P and C8161 cells and the luciferase activity was measured. As
shown in Fig. 3D, preincubation
with Rg3 (50 and 75 μg/ml) significantly suppressed the NF-κB
transcription activity (P<0.05 or <0.01). These results
demonstrate the inhibitory effects of Rg3 on the NF-κB signaling
pathway and the NF-κB DNA binding/transcription activities.
Rg3 decreases FUT4 expression and
inhibits NF-κB binging to FUT4 promoter
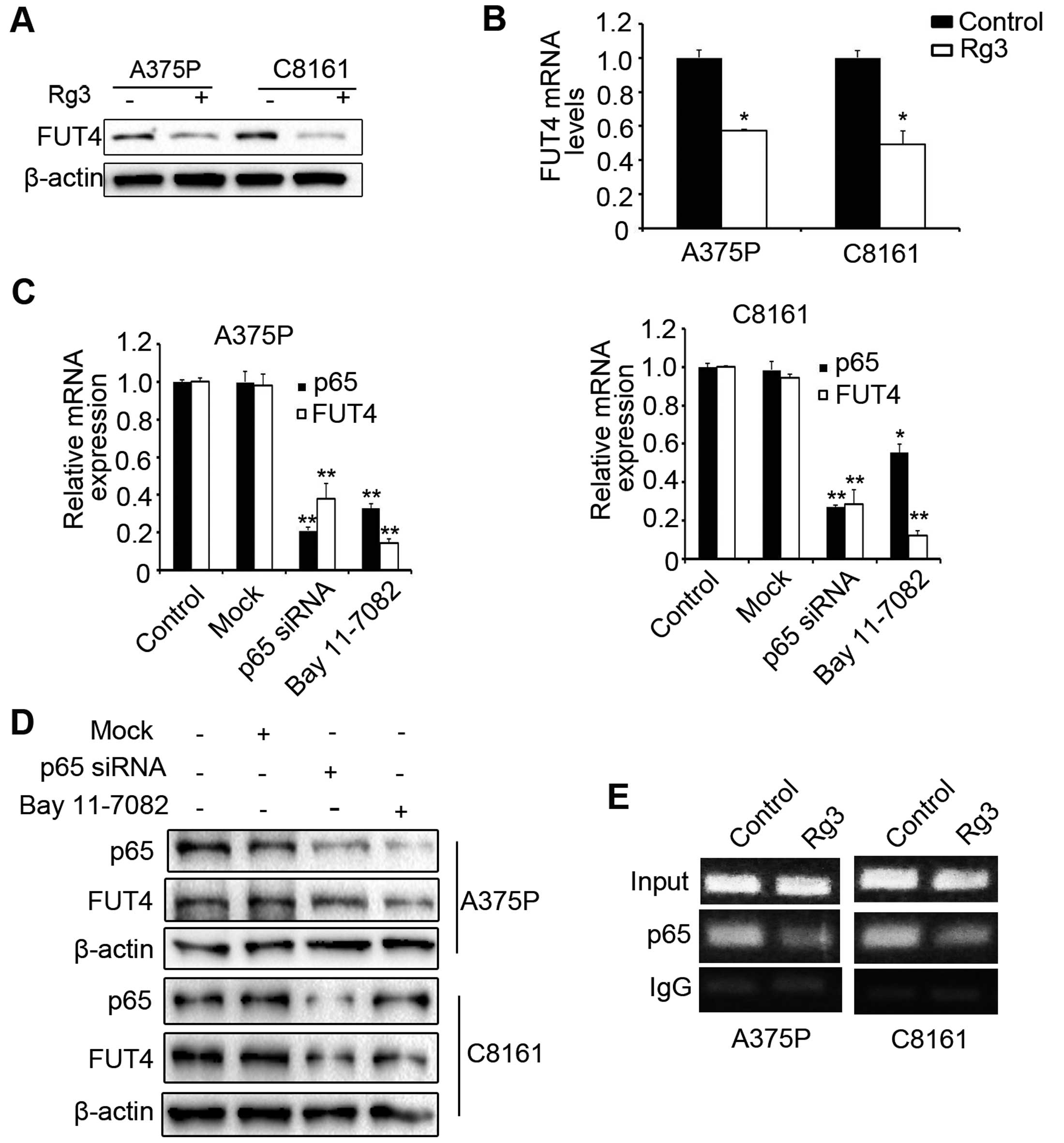
A previous study showed that FUT4 was an
anti-apoptosis protein. Increased expression of FUT4 inhibited
cyclophosphamide-induced apoptosis in A431 cells (21). To examine whether the effect of
Rg3-induced apoptosis was related to decreased expression of FUT4.
A375P and C8161 cells were treated with Rg3 (75 μg/ml) for 24 h and
FUT4 expression was examined. Results showed that treatment with
Rg3 suppressed FUT4 expression in both protein (Fig. 4A) and mRNA (Fig. 4B) levels. These results suggest
that Rg3 downregulates the expression of FUT4. To examine whether
FUT4 expression is regulated by NF-κB signaling pathway, NF-κB
signaling was disrupted by either knocking down p65 or treatment
with NF-κB inhibitor, Bay 11-7082 (10 μM). Results showed that
disrupting NF-κB downregulated the expression of FUT4 at both mRNA
(Fig. 4C) and protein (Fig. 4D) levels. We further evaluated the
effect of Rg3 on the binding activities of NF-κB to FUT4 promoter
by ChIP assay. Results showed that treatment of cells with Rg3 for
24 h markedly inhibited the binding of NF-κB p65 subunits to the
FUT4 chromatin structure as compared with the control (Fig. 4E). These results suggest that the
inhibition of FUT4 expression by Rg3 was modulated by the NF-κB/p65
signaling pathway and FUT4 is a possible downstream target of
NF-κB.
Rg3 induces apoptosis by inhibiting both
the NF-κB signaling and FUT4 expression
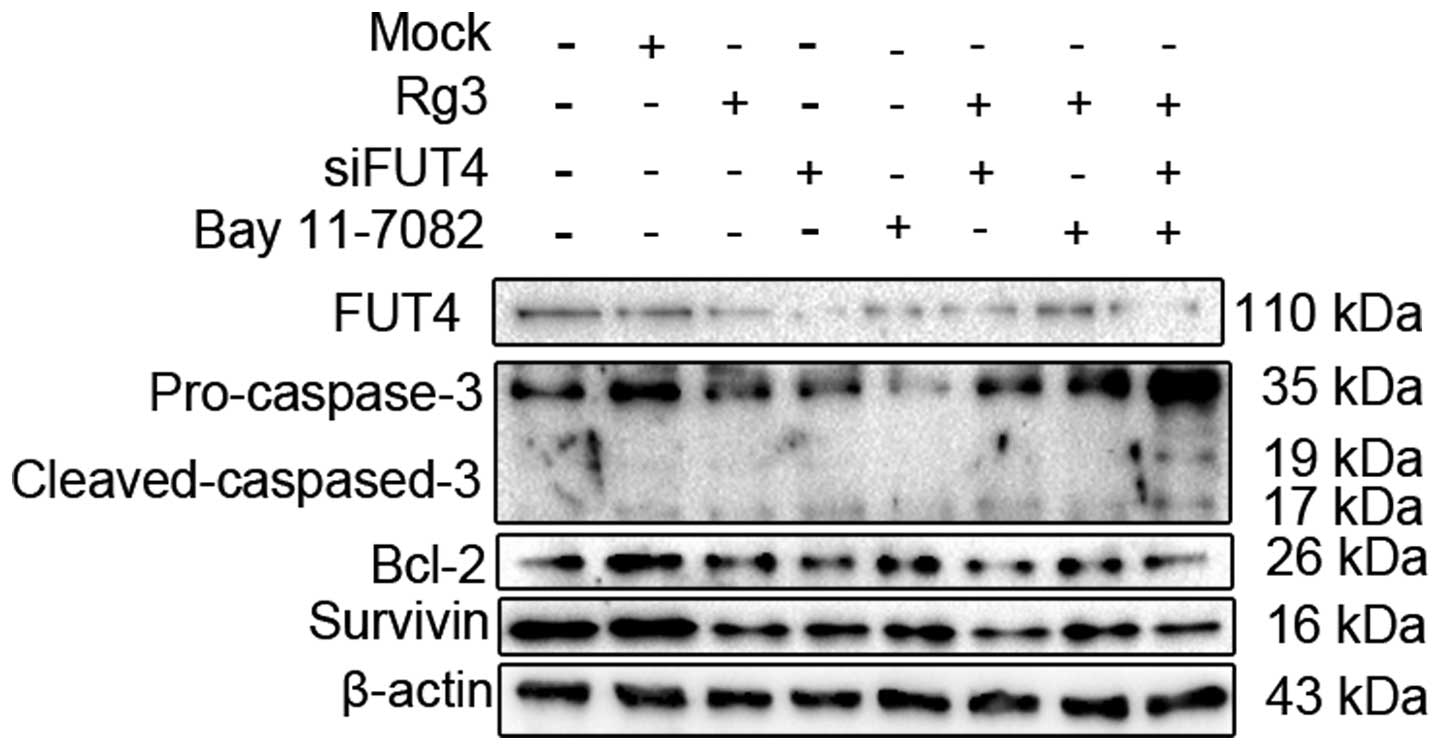
To determine the role of NF-κB signaling pathway and
the FUT4 expression in Rg3-induced apoptosis, A375P cells were
incubated with Rg3 (75 μg/ml), FUT4 siRNA and Bay 11-7082 (10 μM)
for 24 h, or preincubated with FUT4 siRNA 48 h, Bay 11-7082 (10 μM)
24 h, followed by Rg3 (75 μg/ml) treatment for 24 h. As shown in
Fig. 5, treatment with Rg3, FUT4
siRNA and NF-κB inhibitor resulted in the increased expression of
cleavage of caspase-3 by 2.0-, 2.5-and 1.2-fold, and decreased the
expression of anti-apoptosis proteins Bcl-2 and survivin. While Rg3
combined with FUT4 siRNA and Bay 11-7082 showed higher effect on
the expression of apoptosis-related proteins than either of them
alone. This result suggested that Rg3-induced melanoma cell
apoptosis was augmented by FUT4 siRNA and disrupted the NF-κB
signaling pathway with FUT4 downregulation. Moreover, NF-κB
signaling pathway blockage and FUT4 downregulation could be an
effective approach in sensitization of the antitumor efficacy of
Rg3 in human melanoma cells.
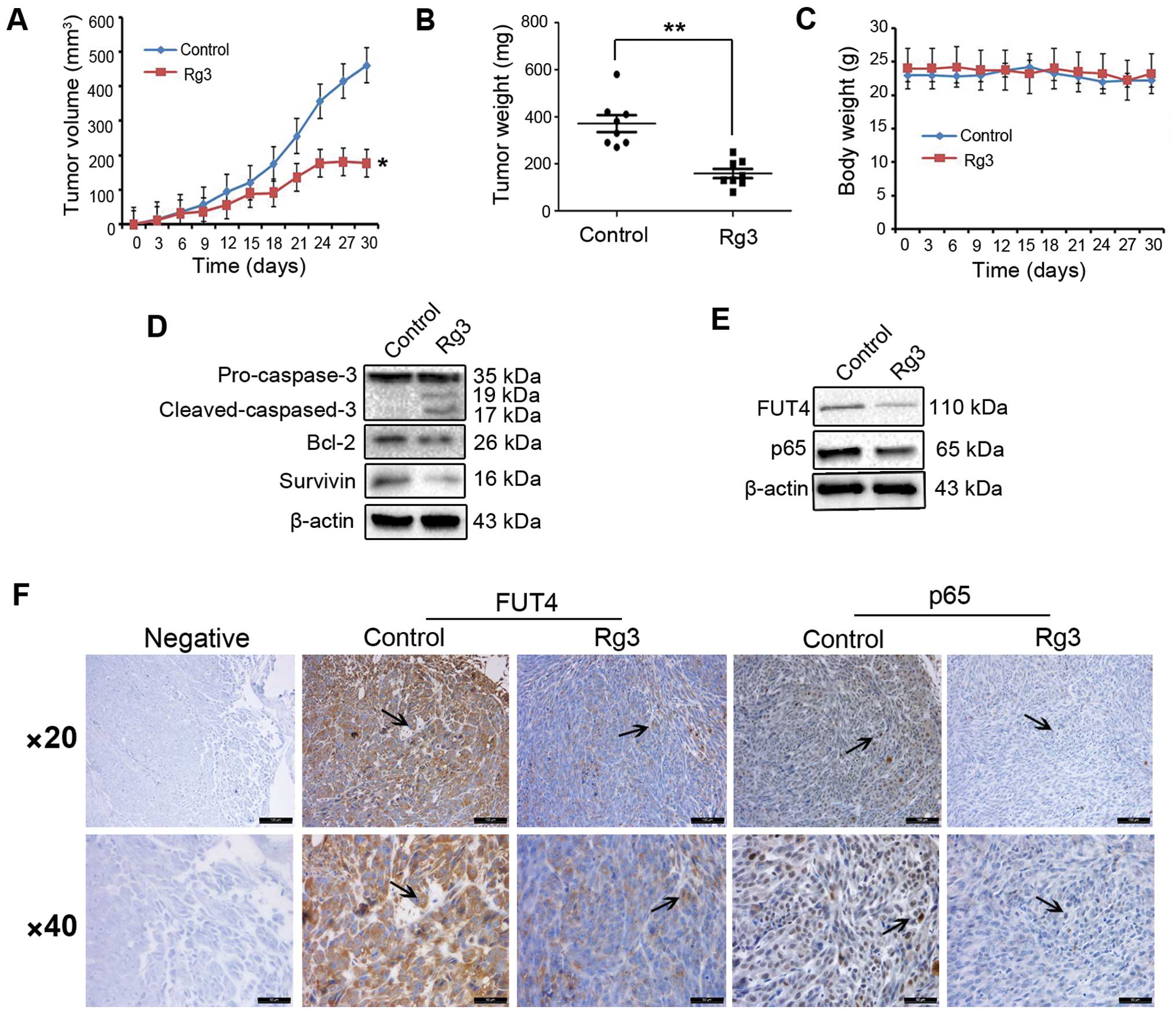
Rg3 suppresses the growth of human
melanoma xenografts
The anticancer activity of Rg3 was further evaluated
in A375P melanoma cell xenograft mouse model. As showed in Fig. 6A, treatment with Rg3 led to a
significant inhibition of tumor growth by 53.56% (P<0.05). Tumor
weight in groups treated with Rg3 (20 mg/kg) decreased by 52.86%
compared with the control group (Fig.
6B, P<0.01). All animals survived during the experimental
period, treatment with Rg3 (20 mg/kg) was well tolerated and there
was no significant loss of body weight or any observable toxic
effect in mice treated with Rg3 in comparison with the control
(Fig. 6C). The regulation of
apoptosis-related proteins in the xenograft tumor specimens was
also assessed. As shown in Fig.
6D, Rg3 treatment led to a significantly increased expression
of cleavage of caspase-3, and decreased expression of the
anti-apoptosis proteins Bcl-2 and survivin in comparison to
controls. Moreover, by western blot analysis and
immunohistochemical staining, we found that the expression levels
of FUT4 and p65 were significantly decreased with Rg3 treatment as
compared with the control group (Fig.
6E and F).
Discussion
It has been well established that Rg3 has strong
antitumor activity. The abnormal activation of signal pathways is
closely related to tumor cell growth, apoptosis and metastasis. Our
previous study showed that Rg3-induced EGFR/MAPK pathway
deactivation inhibiting melanoma cells proliferation via decreasing
FUT4/LeY expression (27). It has
been reported that Rg3 promotes apoptosis in human ovarian cancer
through the PI3K/Akt pathways (28). Rg3 also sensitizes prostate cancer
cells to docetaxel and other chemotherapeutics by inhibiting cell
growth and inducing apoptosis via inhibition of NF-κB signaling
pathway (24). Rg3 induced
apoptosis in MDA-MB-231 cells by blocking NF-κB signaling via
inactivation of ERK and Akt as well as destabilization of mutant
p53 (29). In the present study,
we demonstrated that Rg3 effectively inhibited melanoma cell growth
and induced apoptosis in a dose-dependent manner, with
IC50 range from 57 to 83 μg/ml. We discovered that Rg3
inhibited melanoma growth and induced apoptosis through inhibiting
NF-κB/FUT4 signaling pathway and activating the extrinsic and the
intrinsic apoptotic pathways both in vitro and in
vivo.
FUT4, as a key enzyme involved in LeY synthesis,
plays multiple roles in the regulation of various pathophysiologic
processes. Emerging evidence has shown that FUT4 is a potential
target to inhibit cell proliferation, survival, invasion and
metastasis (19,20,29).
Suppressing FUT1/4 expression blocked EGF-induced tyrosine
phosphorylation of EGFR and MAPK and inhibited cancer growth
(30); Yang et al (21) reported that FUT4 overexpression
inhibited cyclophosphamide-induced apoptosis through activating the
ERK/MAPK and PI3K/Akt signaling pathways. Moreover, FUT4 stimulates
epithelial-mesenchymal transition (EMT) to promote tumor metastasis
(20). FUT4 expression can be
regulated in different levels, including modulation of FUT4
promoter activity, interfering gene transcription and/or
translation. For example, in breast cancer, FUT4 transcription is
regulated by HSF1 and Sp1 to inhibit cell proliferation (31); in HaCaT cells, FUT4 level is lower
due to the higher methylation of CpG island in FUT4 promoter
(32). Knocking down FUT1/4
expression by short interfering RNA technique dramatically reduced
the expression of FUT1/4 and the end product of FUT4, LeY, by which
to inhibit cell proliferation (30). In the present study, we found that
NF-κB/p65 dependent transcriptional regulation of FUT4 inhibited
cell proliferation and induced apoptosis in human melanoma cells.
Suppression of p65 by either siRNA or the potent NF-κB inhibitor
Bay 11-7082 inhibited FUT4 expression. To the best of our
knowledge, this is the first report that NF-κB/p65 signaling
regulates the expression of FUT4. These results suggest that FUT4
is one of the target molecules regulated by NF-κB/p65.
The transcription factor, nuclear factor-κB (NF-κB)
plays significant roles in the regulation of cell growth, survival
and apoptosis. NF-κB activation allows cancer cells to escape
apoptosis, and attenuates the effect of chemotherapeutic
agent-induced apoptosis (11,33).
Previous studies have shown that abnormal activation of NF-κB
correlates with numerous human malignancies, such as pancreatic,
breast, colon cancer, T-cell leukemia and melanoma (34–37).
NF-κB stimulates the expression of anti-apoptotic proteins, such as
the inhibitor of apoptosis proteins and Bcl-2 family members,
thereby promoting cell survival (38). Suppression of NF-κB activity
significantly reduced cell growth, induced apoptosis and increased
sensitivity to radiation-induced cytotoxicity (11). In the present study, we found that
Rg3 not only inhibited the expression of NF-κB/p65, IκBα and IκB
kinase (IKKα/β) activation, but also inhibited NF-κB DNA binding
and transcription activity in human melanoma cells. Aberrant
expression of FUT4 is known to correlate with the activation of
NF-κB signaling pathway. Yang et al (20) showed that FUT4 knockdown led to
decreased nuclear expression of NF-κB in breast cell lines. Cheng
et al (39) found that, in
MDR hepatocellular carcinoma cells, interference or overexpression
of FUT4 decreased or increased the expression of NF-κB and NF-κB
DNA-binding, respectively. Therefore, we hypothesized that NF-κB
could also regulate FUT4 expression. In the present study, our data
showed that Rg3 inhibited NF-κB translocation from cytosol to
nuclear and the silencing of NF-κB/p65 not only significantly
downregulated p65 expression, but also suppressed FUT4 expression.
Moreover, Rg3 inhibited NF-κB/p65 binding to FUT4 promoter to
abrogate FUT4 transcriptional activation, by which to reduce FUT4
expression promoting apoptosis.
Collectively, the present study demonstrates that
Rg3 inhibits the growth of human melanoma cells and induces cell
apoptosis both in vitro and in vivo. Moreover,
downregulation of FUT4 expression by Rg3 induces cell apoptosis
through inhibiting NF-κB signaling pathway. These results suggest
that suppression of NF-κB/FUT4 activation by Rg3 may be a useful
strategy in the prevention or treatment of melanoma. These findings
provide strong evidence that Rg3 is a potential novel therapeutic
for treatment of melanoma.
Acknowledgements
The present study was supported by the funds from
the National Natural Science Foundation of China (nos. 30672753 and
31270866); the Program for Changjiang Scholars and the Innovative
Research Team in University of China (IRT13049).
Abbreviations:
|
FUT4
|
fucosyltransferase IV
|
|
NF-κB
|
nuclear factor-κB
|
|
LeY
|
Lewis Y
|
|
EMT
|
epithelial-mesenchymal transition
|
|
siRNA
|
small interfering RNA
|
|
PARP
|
poly ADP-ribose polymerase
|
|
DAPI
|
4,6-diamino-2-phenyl indole
|
|
EMSA
|
electrophoretic mobility shift
assay
|
|
ChIP
|
chromatin immunoprecipitation
|
References
|
1
|
McGettigan S: Dabrafenib: A new therapy
for use in BRAF-mutated metastatic melanoma. J Adv Pract Oncol.
5:211–215. 2014.PubMed/NCBI
|
|
2
|
Lens MB and Dawes M: Global perspectives
of contemporary epidemiological trends of cutaneous malignant
melanoma. Br J Dermatol. 150:179–185. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rigel DS, Russak J and Friedman R: The
evolution of melanoma diagnosis: 25 years beyond the ABCDs. CA
Cancer J Clin. 60:301–316. 2010. View Article : Google Scholar
|
|
4
|
Menaa F: Latest approved therapies for
metastatic melanoma: What comes next? J Skin Cancer.
2013:7352822013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Graham B and Gibson SB: The two faces of
NFkappaB in cell survival responses. Cell Cycle. 4:1342–1345. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kiernan R, Brès V, Ng RWM, Coudart MP, El
Messaoudi S, Sardet C, Jin DY, Emiliani S and Benkirane M:
Post-activation turn-off of NF-κB-dependent transcription is
regulated by acetylation of p65. J Biol Chem. 278:2758–2766. 2003.
View Article : Google Scholar
|
|
7
|
Doyle SL and O’Neill LA: Toll-like
receptors: From the discovery of NFkappaB to new insights into
transcriptional regulations in innate immunity. Biochem Pharmacol.
72:1102–1113. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perkins ND and Gilmore TD: Good cop, bad
cop: The different faces of NF-kappaB. Cell Death Differ.
13:759–772. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kashani-Sabet M, Liu Y, Fong S, Desprez
PY, Liu S, Tu G, Nosrati M, Handumrongkul C, Liggitt D, Thor AD, et
al: Identification of gene function and functional pathways by
systemic plasmid-based ribozyme targeting in adult mice. Proc Natl
Acad Sci USA. 99:3878–3883. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kashani-Sabet M, Shaikh L, Miller JR III,
Nosrati M, Ferreira CM, Debs RJ and Sagebiel RW: NF-kappa B in the
vascular progression of melanoma. J Clin Oncol. 22:617–623. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Munshi A, Kurland JF, Nishikawa T, Chiao
PJ, Andreeff M and Meyn RE: Inhibition of constitutively activated
nuclear factor-kappaB radiosensitizes human melanoma cells. Mol
Cancer Ther. 3:985–992. 2004.PubMed/NCBI
|
|
12
|
Ichikawa H, Takada Y, Shishodia S,
Jayaprakasam B, Nair MG and Aggarwal BB: Withanolides potentiate
apoptosis, inhibit invasion, and abolish osteoclastogenesis through
suppression of nuclear factor-kappaB (NF-kappaB) activation and
NF-kappaB-regulated gene expression. Mol Cancer Ther. 5:1434–1445.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fuster MM and Esko JD and Esko JD: The
sweet and sour of cancer: Glycans as novel therapeutic targets. Nat
Rev Cancer. 5:526–542. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Weston BW, Nair RP, Larsen RD and Lowe JB:
Isolation of a novel human alpha (1,3)fucosyltransferase gene and
molecular comparison to the human Lewis blood group alpha (1,3/1,4)
fucosyltransferase gene. Syntenic, homologous, nonallelic genes
encoding enzymes with distinct acceptor substrate specificities. J
Biol Chem. 267:4152–4160. 1992.PubMed/NCBI
|
|
15
|
Weston BW, Smith PL, Kelly RJ and Lowe JB:
Molecular cloning of a fourth member of a human alpha
(1,3)fucosyltransferase gene family. Multiple homologous sequences
that determine expression of the Lewis x, sialyl Lewis x, and
difucosyl sialyl Lewis x epitopes. J Biol Chem. 267:24575–24584.
1992.PubMed/NCBI
|
|
16
|
Petretti T, Schulze B, Schlag PM and
Kemmner W: Altered mRNA expression of glycosyltransferases in human
gastric carcinomas. Biochim Biophys Acta. 1428:209–218. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ito H, Hiraiwa N, Sawada-Kasugai M,
Akamatsu S, Tachikawa T, Kasai Y, Akiyama S, Ito K, Takagi H and
Kannagi R: Altered mRNA expression of specific molecular species of
fucosyl- and sialyl-transferases in human colorectal cancer
tissues. Int J Cancer. 71:556–564. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ogawa J, Inoue H and Koide S: Expression
of alpha-1,3-fucosyltransferase type IV and VII genes is related to
poor prognosis in lung cancer. Cancer Res. 56:325–329.
1996.PubMed/NCBI
|
|
19
|
Yang X, Zhang Z, Jia S, Liu Y, Wang X and
Yan Q: Overexpression of fucosyltransferase IV in A431 cell line
increases cell proliferation. Int J Biochem Cell Biol.
39:1722–1730. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang X, Liu S and Yan Q: Role of
fucosyltransferase IV in epithelial-mesenchymal transition in
breast cancer cells. Cell Death Dis. 4:e7352013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang X, Liu Y, Liu J, Wang X and Yan Q:
Cyclophosphamide-induced apoptosis in A431 cells is inhibited by
fucosyltransferase IV. J Cell Biochem. 112:1376–1383. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuan HD, Quan HY, Zhang Y, Kim SH and
Chung SH: 20(S)-Ginsenoside Rg3-induced apoptosis in HT-29 colon
cancer cells is associated with AMPK signaling pathway. Mol Med
Rep. 3:825–831. 2010.
|
|
23
|
Park EH, Kim YJ, Yamabe N, Park SH, Kim
HK, Jang HJ, Kim JH, Cheon GJ, Ham J and Kang KS: Stereospecific
anticancer effects of ginsenoside Rg3 epimers isolated from
heat-processed American ginseng on human gastric cancer cell. J
Ginseng Res. 38:22–27. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim SM, Lee SY, Cho JS, Son SM, Choi SS,
Yun YP, Yoo HS, Yoon Y, Oh KW, Han SB, et al: Combination of
ginsenoside Rg3 with docetaxel enhances the susceptibility of
prostate cancer cells via inhibition of NF-kappaB. Eur J Pharmacol.
631:1–9. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee CK, Park KK, Chung AS and Chung WY:
Ginsenoside Rg3 enhances the chemosensitivity of tumors to
cisplatin by reducing the basal level of nuclear factor erythroid
2-related factor 2-mediated heme oxygenase-1/NAD(P)H quinone
oxido-reductase-1 and prevents normal tissue damage by scavenging
cisplatin-induced intracellular reactive oxygen species. Food Chem
Toxicol. 50:2565–2574. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu Y, Zhang C, Liu L and Li X: Hepatic
arterial administration of ginsenoside Rg3 and transcatheter
arterial embolization for the treatment of VX2 liver carcinomas.
Exp Ther Med. 5:761–766. 2013.PubMed/NCBI
|
|
27
|
Shan X, Aziz F, Tian LL, Wang XQ, Yan Q
and Liu JW: Ginsenoside Rg3-induced EGFR/MAPK pathway deactivation
inhibits melanoma cell proliferation by decreasing FUT4/LeY
expression. Int J Oncol. 46:1667–1676. 2015.PubMed/NCBI
|
|
28
|
Wang JH, Nao JF, Zhang M and He P:
20(s)-ginsenoside Rg3 promotes apoptosis in human ovarian cancer
HO-8910 cells through PI3K/Akt and XIAP pathways. Tumour Biol.
35:11985–11994. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim BM, Kim DH, Park JH, Surh YJ and Na
HK: Ginsenoside Rg3 inhibits constitutive activation of NF-κB
signaling in human breast cancer (MDA-MB-231) cells: ERK and Akt as
potential upstream targets. J Cancer Prev. 19:23–30. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Z, Sun P, Liu J, Fu L, Yan J, Liu Y,
Yu L, Wang X and Yan Q: Suppression of FUT1/FUT4 expression by
siRNA inhibits tumor growth. Biochim Biophys Acta. 1783:287–296.
2008. View Article : Google Scholar
|
|
31
|
Yang X, Wang J, Liu S and Yan Q: HSF1 and
Sp1 regulate FUT4 gene expression and cell proliferation in breast
cancer cells. J Cell Biochem. 115:168–178. 2014. View Article : Google Scholar
|
|
32
|
Hongyan L, Shaoming T and Qiu Y:
Correlation between FUT4 expression and its promoter methylation in
HaCaT cells. Yi Chuan. 37:48–54. 2015.PubMed/NCBI
|
|
33
|
Kim MK, Jeon YK, Woo JK, Choi Y, Choi DH,
Kim YH and Kim CW: The C-terminal region of Bfl-1 sensitizes
non-small cell lung cancer to gemcitabine-induced apoptosis by
suppressing NF-κB activity and down-regulating Bfl-1. Mol Cancer.
10:982011. View Article : Google Scholar
|
|
34
|
Brandi G, Pantaleo MA, Biasco G and
Paterini P: Activated NF-κB in colorectal cancer: Predictive or
prognostic factor? J Clin Oncol. 26:1388–1389; author reply
1389–1390. 2008. View Article : Google Scholar
|
|
35
|
Richmond A: NF-κB, chemokine gene
transcription and tumour growth. Nat Rev Immunol. 2:664–674. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Karin M, Cao Y, Greten FR and Li ZW:
NF-kappaB in cancer: From innocent bystander to major culprit. Nat
Rev Cancer. 2:301–310. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Melnikova VO and Bar-Eli M:
Transcriptional control of the melanoma malignant phenotype. Cancer
Biol Ther. 7:997–1003. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Heon Seo K, Ko HM, Kim HA, Choi JH, Jun
Park S, Kim KJ, Lee HK and Im SY: Platelet-activating factor
induces up-regulation of antiapoptotic factors in a melanoma cell
line through nuclear factor-kappaB activation. Cancer Res.
66:4681–4686. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cheng L, Luo S, Jin C, Ma H, Zhou H and
Jia L: FUT family mediates the multidrug resistance of human
hepatocellular carcinoma via the PI3K/Akt signaling pathway. Cell
Death Dis. 4:e9232013. View Article : Google Scholar : PubMed/NCBI
|