Introduction
Ovarian cancer is the second most common and the
most lethal of malignancies arising in the female reproductive
system (1). Response rates to
first-line platinum-based therapy are >80%, but the overall
5-year survival rate for patients with advanced ovarian cancer is
only ~20% because of acquired drug resistance and adverse side
effects. Therefore, overcoming drug resistance is the key to
successful treatment of ovarian cancer, and the development of
novel therapeutic approaches is urgently needed (2).
Apoptosis is an active death process genetically
encoded that can be triggered by a wide variety of extra- and
intra-cellular stimuli. Recent evidence suggests that the failure
of drug-induced apoptosis may be an underlying cause of drug
resistance. Molecular mechanisms of failed apoptosis in
chemoresistant ovarian cancer cells include p53 mutations, abnormal
expression of the Bcl-2 family proteins and P-glycoproteins, and
upregulation of other inhibitors of apoptosis that block caspases
and stabilize the mitochondrial permeability pore. Triggering
apoptosis is one of the potential strategies to overcome
chemoresistance (3,4). The cell cycle is a series of events
that take place in a cell leading to its division and duplication
that produces two daughter cells. This process involves four
sequential phases that go from quiescence (G0 phase) to
proliferation (G1, S, G2, and M phases) and back to quiescence. A
disregulation of the cell cycle components may lead to tumor
formation. A number of potential molecular targets for novel
anticancer drug discovery have been identified in cell cycle
control mechanisms (5,6).
It is a common belief that compounds from fruits,
vegetables and other foods will help reduce the risk of various
chronic diseases including cancer, and have less side effects
(7). Black tea is the most widely
consumed beverage worldwide, and accounted for ~80% of the dried
tea manufactured annually (8). It
was reported that black tea extract possess potently
antiproliferative property against a variety of cancer cells in
vitro and in vivo (9,10).
In particular, the previous prospective cohort study showed that
black tea was a main dietary source of flavonols for US women, and
its intake was associated with lower risk of ovarian cancer
(11).
Theaflavins are major bioactive components in black
tea, and contribute importantly to properties of black tea
including its color, ‘mouth feel’ and extent of tea cream
formation. They possess a benzotropolone skeleton that is formed
from co-oxidation of appropriate pairs of catechins during the
production of black tea (12). The
major theaflavins in black tea are theaflavin (TF1),
theaflavin-3-gallate (TF2A), theaflavin-3′-gallate (TF2B) and
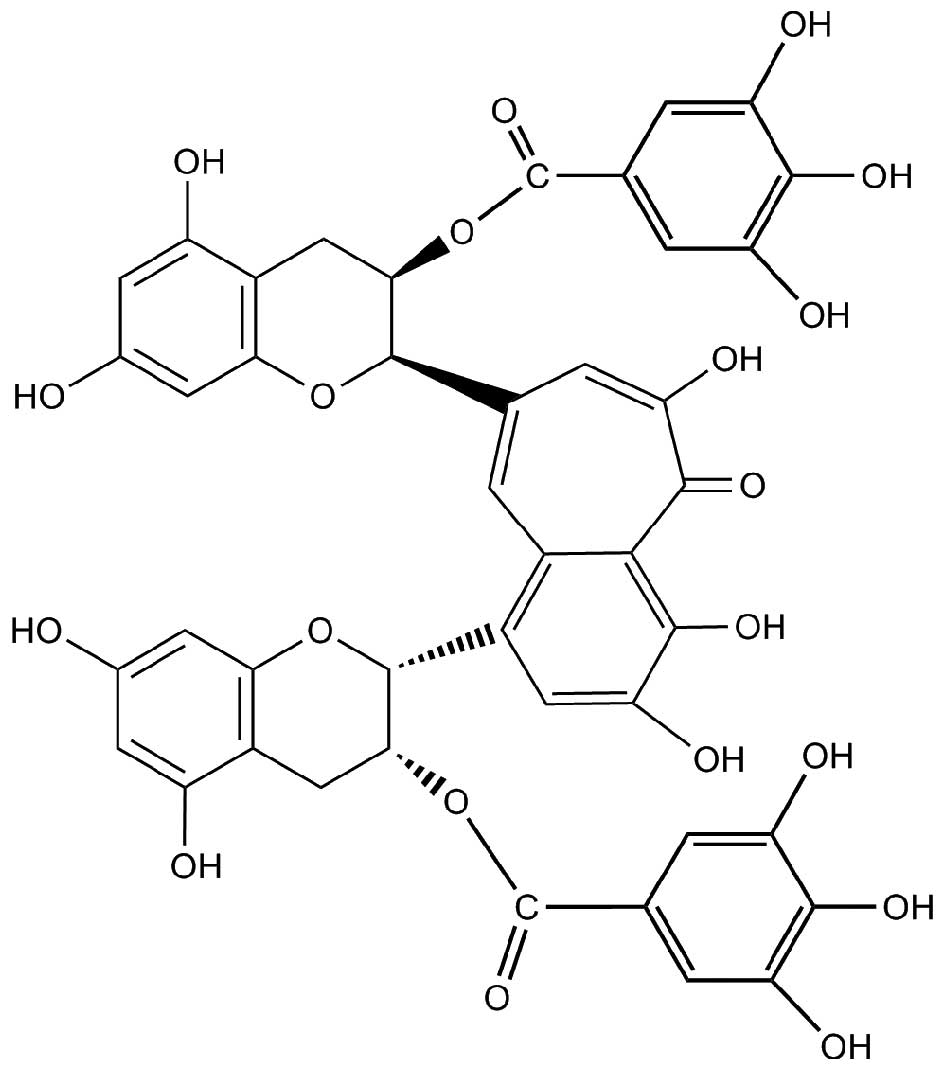
theaflavin-3, 3′-digallate (TF3), and TF3 (Fig. 1) is usually most abundant among
them (13). Theaflavins have been
demonstrated to inhibit proliferation and induce apoptosis in a
variety of cancer cells including human breast cancer MCF-7 cells,
malignant melanoma A375 cells and oral squamous carcinoma HSC-2
cells. In addition, theaflavins are responsible for the inhibition
of ROS-potentiated AH109A adhesion and invasion to the cultured rat
mesothelial cell monolayer (14–17).
Although theaflavins have received considerable
attention for their anticancer activity, their effect on the
ovarian cancer is still not clear. Therefore, the aim of this study
was to investigate the apoptotic and cell cycle arrest effects of
TF3 in the platinum-resistant ovarian cancer cell line A2780/CP70
and a normal ovarian surface epithelial cell line IOSE-364. The
possible mechanisms underlying these modulations of TF3 on the
ovarian cancer cells were also examined.
Materials and methods
Cell culture and reagents
The platinum-resistant human ovarian cancer cell
line A2780/CP70 (p53 wild-type) was a generous gift from Dr B.
Jiang at West Virginia University. IOSE-364, a normal ovarian
surface epithelial cell line, was presented by Dr N. Auersperg at
University of British Columbia, Canada. The cells were cultured in
RPMI-1640 medium (Sigma) supplemented with 10% fetal bovine serum
(FBS) (Invitrogen) at 37°C in a humidified incubator with 5%
CO2. Theaflavin-3, 3′-digallate (TF3, >90.0%) was
purchased from Sigma-Aldrich (St. Louis, MO, USA). The primary
antibodies against Bcl-xL, Bad, p21, p53, MDM2 and GAPDH were
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
The primary antibodies against caspase-8 and -9, Puma, Bax, cyclin
B1, phospho-cdc2 (Tyr15), cdc2, DR5, FADD, phospho-Akt
(Thr180/Tyr182) and total-Akt were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA).
Cell viability assay
The cell viability was assessed a using
3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)
assay. Briefly, the cells (1×104 cells per well) were
seeded in 96-well plate and incubated overnight. Then various
concentrations of TF3 (0–50 μM) were added, and an equal amount of
DMSO was used as control. After treatment for 24 h, 20 μl of MTT (5
mg/ml) was added to each well and incubated for an additional 4 h
at 37°C in the dark. The medium was discarded, and the formazan
crystals formed in the cells were dissolved in 200 μl DMSO. The
optical density was measured at 570 nm using a Synergy™ HT
Multi-Mode Microplate Reader (BioTek).
Flow cytometric analysis of apoptotic
cells
The apoptotic cell death was determined using an
Alexa Fluor® 488 Annexin V/ Dead Cell Apoptosis kit
(Invitrogen). After exposure to TF3 (0–20 μM) for 24 h, cells were
harvested, centrifuged for 10 min at 1500 rpm and then washed twice
with PBS. Cells were suspended in binding buffer with Alexa Fluor
488 Annexin V and propidium iodide (PI) for 15 min. The stained
cells were analyzed by flow cytometry (FACSCalibur system, BD
Biosciences), measuring the fluorescence emission at 530 and 575 nm
using 488 nm excitation.
Flow cytometry analysis of the cell
cycle
Cells treated with TF3 (0–20 μM) for 24 h were
digested with trypsin and harvested by 1500 rpm centrifugation for
10 min. The cell pellet was suspended with 70% ethanol at −20°C
overnight, washed with PBS, then incubated with 180 μg/ml RNase A
at 37°C for 15 min. After 15 min staining with 50 μg/ml PI in the
dark at 37°C, the samples were analyzed by flow cytometry
(FACSCalibur system, BD Biosciences). Data were plotted and
analyzed by using FCS Software (De Novo Software, Los Angeles, CA,
USA).
Caspase-3/7 assay
The caspase-3/7 activities in A2780/CP70 and
IOSE-364 cells were detected using a Caspase-Glo 3/7 Assay kit
(Promega). Cells were seeded in 96-well plates at 1×104
cells/well, incubated overnight, and treated with TF3 (0–20 μM) for
24 h. After treatment, 100 ml of caspase-3/7 reagent were added to
each well, mixed and incubated for 1 h at room temperature.
Luminescence was measured using a Synergy™ HT Multi-Mode Microplate
Reader (BioTek). Caspase-3/7 activities were normalized by total
protein levels, and were expressed as percentage of the untreated
control. The total protein levels were measured with a BCA assay
kit (Pierce).
Western blotting
A2780/CP70 ovarian cancer cells were seeded in 60-mm
dishes at 1×106 cells/dish, incubated overnight, and
treated with 0–20 μM TF3 for 24 h. Then cells were harvested with
M-PER Mammalian Protein Extraction Reagent (Pierce) supplemented
with Halt Protease and Phosphatase Inhibitor (Pierce), and total
protein levels were assayed with BCA Protein assay kit (Pierce).
Cell lysates were separated by SDS-PAGE and blotted onto
nitrocellulose membrane with a Mini-Protean 3 System (Bio-Rad). The
membrane was blocked with 5% skim milk, and then incubated with
specific primary antibodies and appropriate secondary antibodies
conjugated with horseradish peroxidase. The antigen-antibody
complex was visualized with the ECL kit (Pierce). Protein bands
were quantitated with NIH Image J software and normalized by GAPDH
bands for analysis.
Transfection with small interfering RNA
(siRNA)
A2780/ CP70 ovarian cancer cells were seeded in
60-mm dishes at 5×105 cells/dish, incubated overnight,
and then transfected with p53 siRNA (Santa Cruz) using
Lipofectamine 2000 transfection reagent (Invitrogen) according to
the manufacturer's protocol. Cells transfected with control siRNA
(Santa Cruz) were used as controls. After a 24-h transfection
period, cells were treated with TF3 (0–20 μM) for 24 h. Cell
lysates were collected for western blot analysis.
Statistical analysis
In this study, all samples were prepared and
analyzed in triplicate. The data are presented as means ± standard
deviations (SD). Multiple comparisons were performed by one-way
analysis of variance (ANOVA) followed with Student-Newman-Keuls
(SNK) test. Statistical differences between two groups were
evaluated using the Student's t-test. All statistical analyses of
data were performed using Statistical Analysis System (SAS) for
windows V8. A p<0.05 was considered statistically significant,
and p<0.01 was considered statistically highly significant.
Results
Effect of TF3 on A2780/CP70 and IOSE-364
cell viability
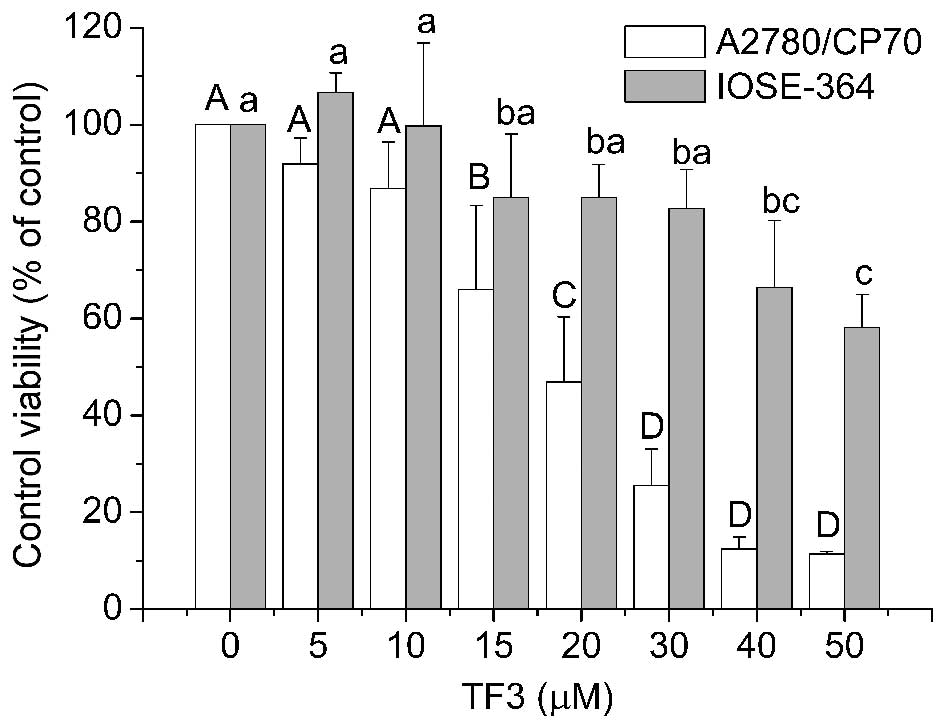
The cytotoxicity of TF3 on A2780/CP70 and IOSE-364
cells was analyzed using the MTT assay. As shown in Fig. 2, TF3 inhibited the growth of both
the cell types in a dose-dependent manner. The cell viability with
TF3 treatment (0–50 μM) for 24 h ranged from 100 to 11.38% for
A2780/CP70 cells, and from 100 to 58.14% for IOSE-364 cells. The
IC50 values of TF3 for A2780/CP70 and IOSE-364 cells
were estimated to be 23.81 and 59.58 μM, respectively. These
results indicated that TF3 had lower cytotoxic effect against
normal ovarian IOSE-364 cells than ovarian cancer A2780/CP70
cells.
TF3 induces apoptosis in A2780/CP70
cells
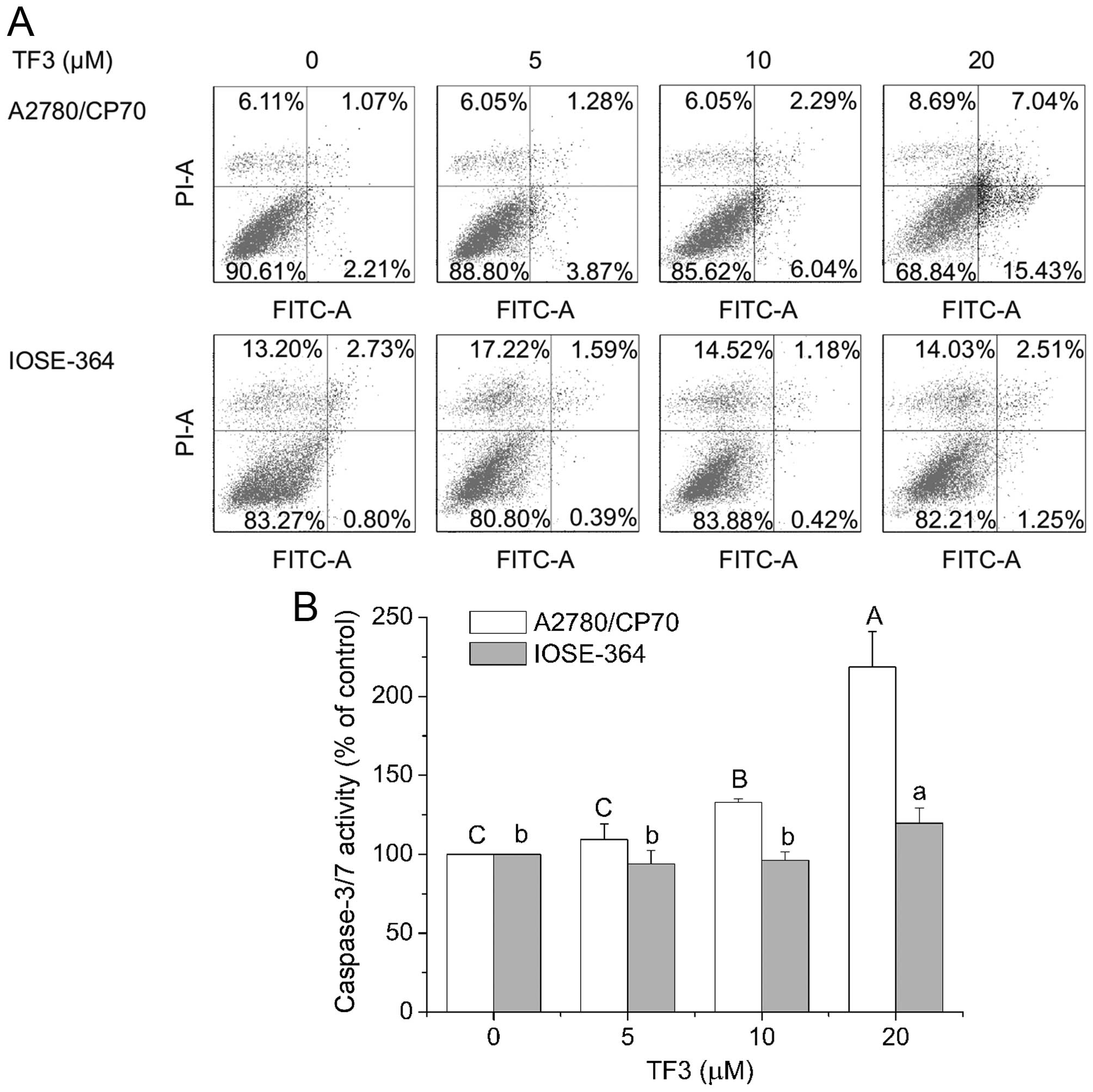
To investigate whether TF3 inhibited cell viability
by inducing apoptosis in A2780/CP70 and IOSE-364 cells, the cells
were treated with TF3 (0–20 μM) for 24 h and subsequently subjected
to double staining with Annexin V FITC and propidium iodide (PI)
followed by flow cytometry analysis. As shown in Fig. 3A, the treatment of TF3 increased
the total percent of apoptotic cells (upper right quadrant + low
right quadrant) from 3.28 to 22.47% in A2780/CP70 cells, but could
hardly induce apoptosis of IOSE-364 cells in the test range of 0–20
μM. To confirm that TF3 induced apoptosis, caspase-3/7 enzymatic
activities were evaluated in both cell lines. As shown in Fig. 3B, treatment with TF3 maximally
increased the caspase-3/7 enzymatic activity to 2.19- and 1.20-fold
of that in controls for A2780/CP70 and IOSE-364 cells, respectively
(p<0.05), indicating that TF3 had more potential to induce
apoptosis in the ovarian cancer cells than the normal ovarian cells
in this study.
TF3 induces G2 cell cycle arrest in
A2780/CP70 cells
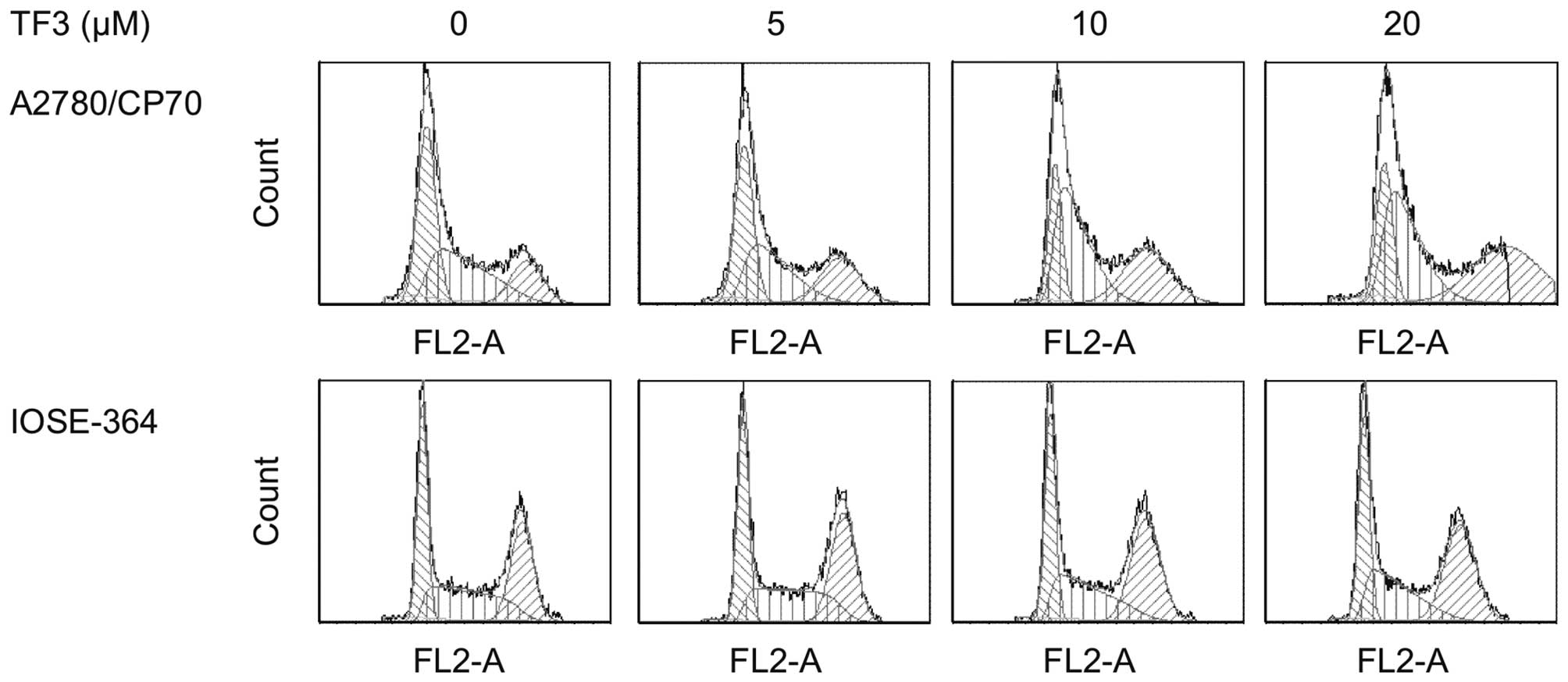
To determine whether the growth inhibitory effect of
TF3 is caused by specific perturbation of cell cycle-related
events, cell cycle phase distribution of cells treated with TF3
(0–20 μM) for 24 h was analyzed by flow cytometry after propidium
iodide staining. As shown in Fig.
4 and Table I, treatment of
A2780/CP70 and IOSE-364 cells with TF3 resulted in a significant
increase in the proportion of cells at the G2 phase in a
concentration-dependent manner (p<0.05). The G2 phase percentage
of the A2780/CP70 cells increased by 21.78%, while that of IOSE-364
cells increased by 6.68% after 20 μM TF3 treatment for 24 h. These
results suggested that TF3 had more potent capability to induce G2
cell cycle arrest for ovarian cancer cells than the ovarian normal
cells.
 | Table ICell cycle phase distribution of
A2780/CP70 and IOSE-364 cells with TF3 treatment. |
Table I
Cell cycle phase distribution of
A2780/CP70 and IOSE-364 cells with TF3 treatment.
| Cell cycle phase
distribution |
|---|
|
|
|---|
| Cell lines | TF3 (μM) | % G1 | % G2 | % S |
|---|
| A2780/CP70 | 0 |
42.47±1.33a |
18.00±1.22d |
39.53±2.65a,b |
| 5 |
39.32±2.71a |
25.04±0.79c |
35.64±1.87b |
| 10 |
26.57±1.26b |
29.08±0.86b |
44.35±3.37a |
| 20 |
21.81±1.94c |
39.78±1.73a |
38.41±2.83a,b |
| IOSE-364 | 0 |
32.93±0.85a |
32.91±1.47b |
34.16±1.93a |
| 5 |
31.52±2.23a |
35.35±2.39b |
33.14±2.61a |
| 10 |
29.55±2.71a |
39.08±1.24a |
31.37±1.06a |
| 20 |
30.32±1.64a |
39.59±1.67a |
30.08±2.27a |
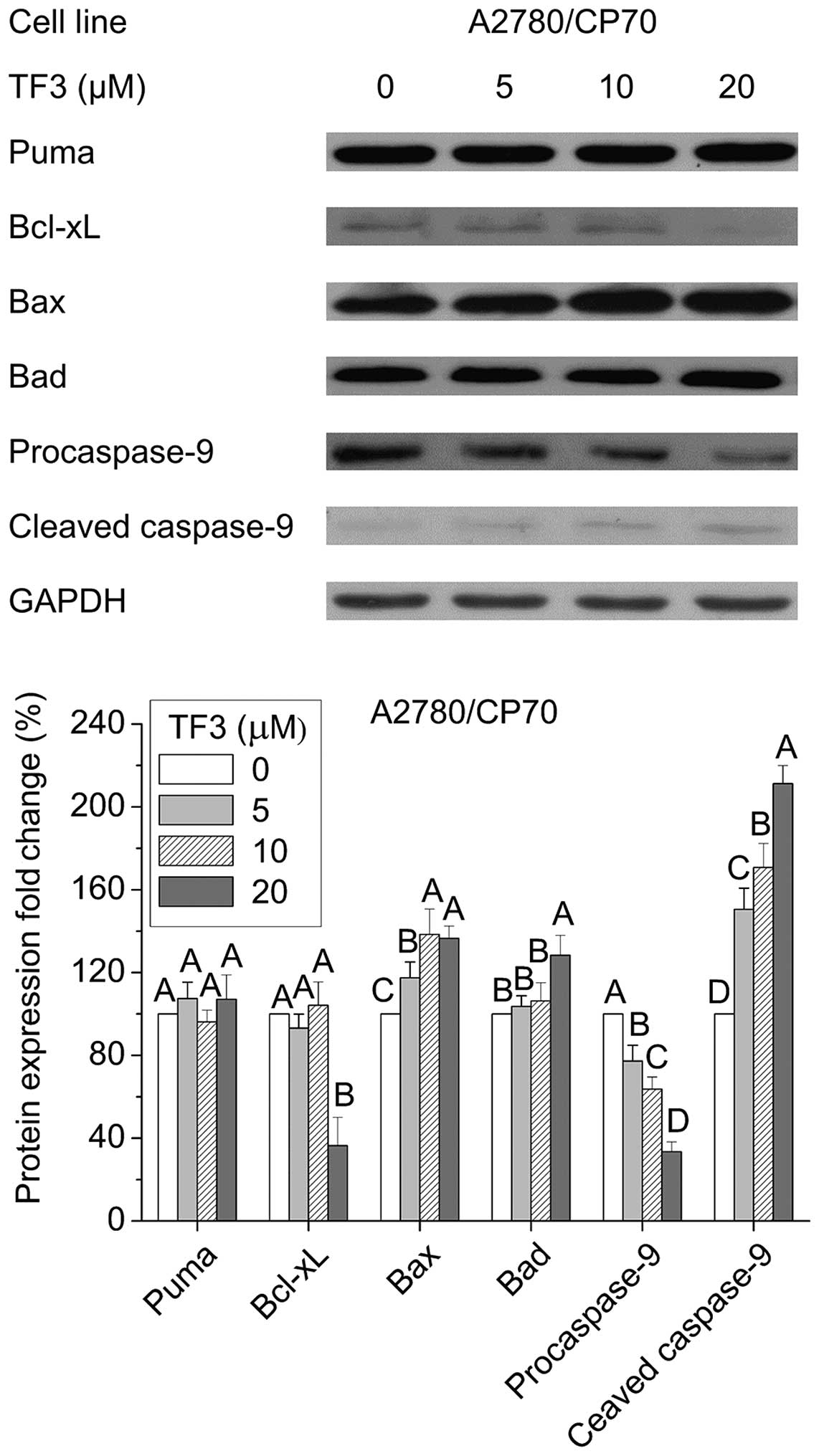
Effect of TF3 on the intrinsic apoptotic
pathway in A2780/ CP70 cells
Apoptosis can be initiated via two canonical
pathways: the intrinsic or mitochondrial pathway, and the extrinsic
or receptor-mediated pathway. To clarify whether the intrinsic
pathway is involved in TF3-induced apoptosis of A2780/CP70 cells,
we examined the protein expression of pro-apoptotic Bcl-2 family
proteins (Puma, Bax and Bad), anti-apoptotic Bcl-2 family protein
Bcl-xL, and caspase-9 by western blotting. Fig. 5 shows TF3 significantly increased
the protein levels of Bax, Bad and cleaved caspase-9, reduced
protein expressions of Bcl-xL and procaspase-9 (p<0.05), and had
no effect on Puma protein expression (p>0.05). These results
suggested that TF3 induced apoptosis in A2780/CP70 cells through
the intrinsic apoptotic pathway.
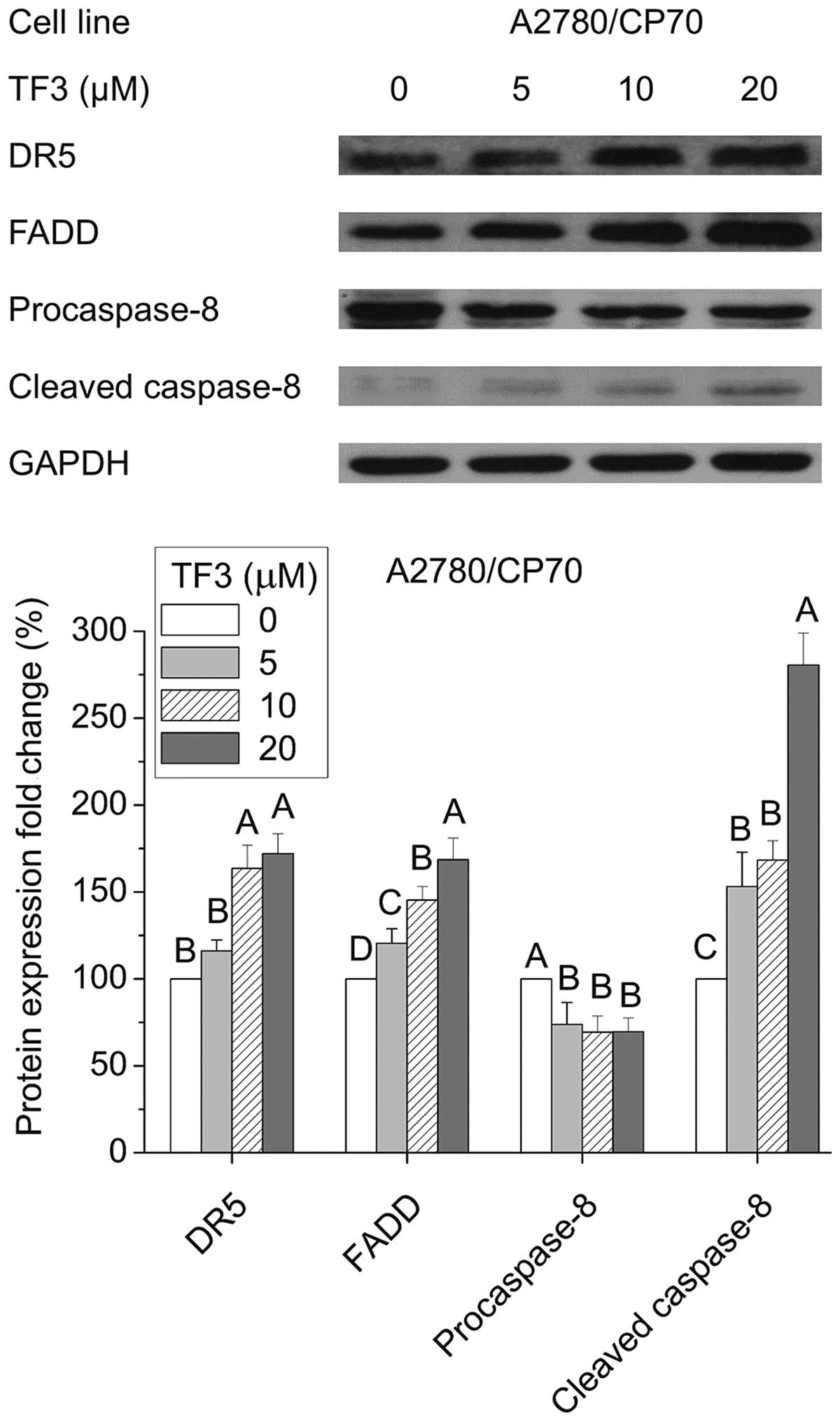
Effect of TF3 on the extrinsic apoptotic
pathway in A2780/ CP70 cells
Next, we investigated whether TF3 could activate the
extrinsic apoptotic pathway in A2780/CP70 cells. As shown in
Fig. 6, TF3 significantly
increased protein expression of DR5, FADD and cleaved caspase-8,
and decreased the protein level of procaspase-8 (p<0.05). These
results indicated that the extrinsic pathway was involved in
TF3-induced apoptosis of A2780/CP70 cells.
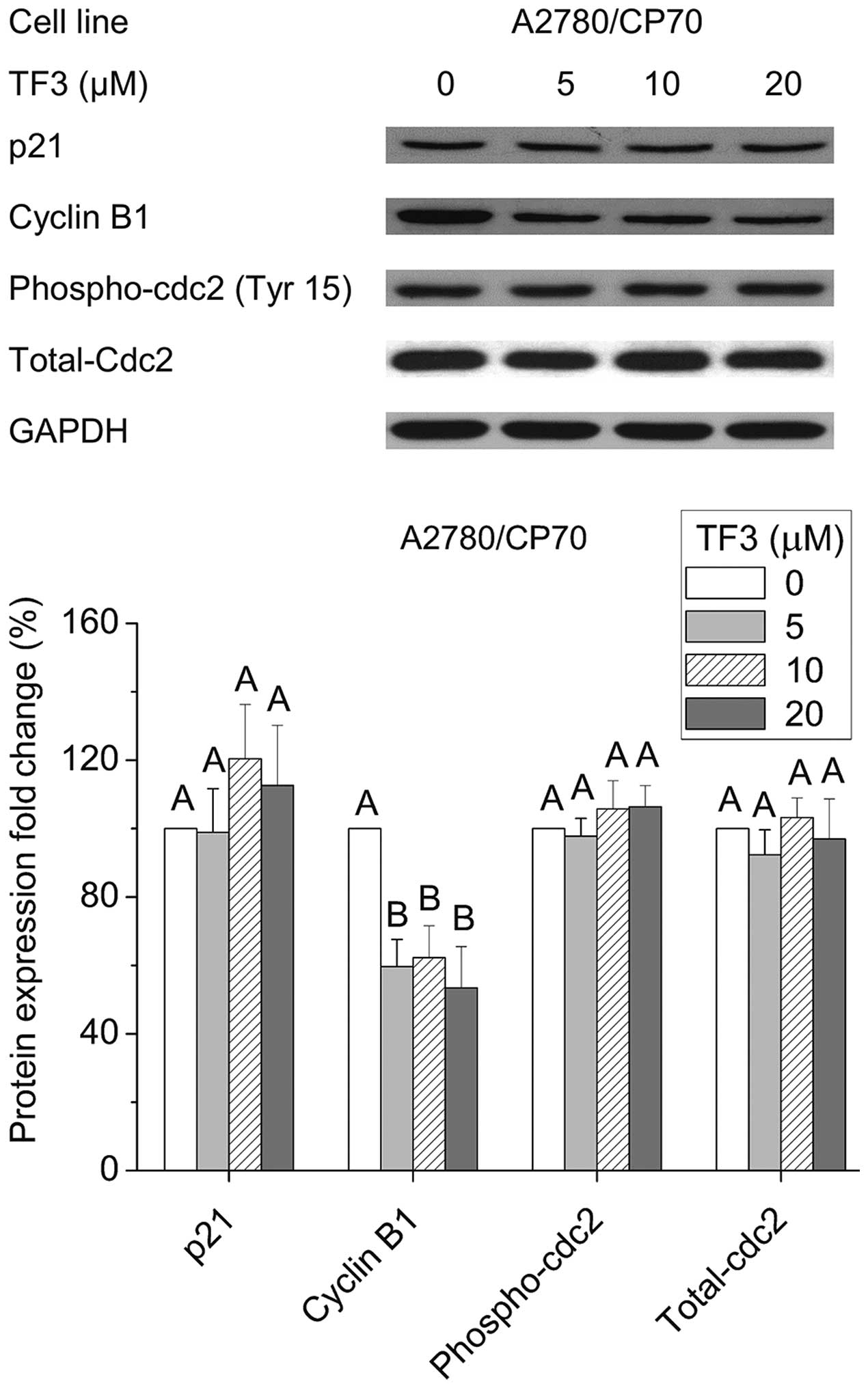
Effect of TF3 on cell cycle G2-related
proteins
To investigate the underlying mechanism of the G2
arrest induced by TF3 in A2780/CP70 cells, we evaluated the effect
of TF3 on cell cycle regulatory proteins that play important roles
in G2 cell cycle progression by western blot analysis. As shown in
Fig. 7, TF3 could effectively
suppress cyclin B1 expression (p<0.05), but had no effect on the
protein levels of p21, phospho-cdc2 (Tyr15) or total-cdc2
(p>0.05). These results implied that the down-regulation of
cyclin B1 protein expression may be responsible for the G2 growth
arrest induced by TF3 in A2780/CP70 cells.
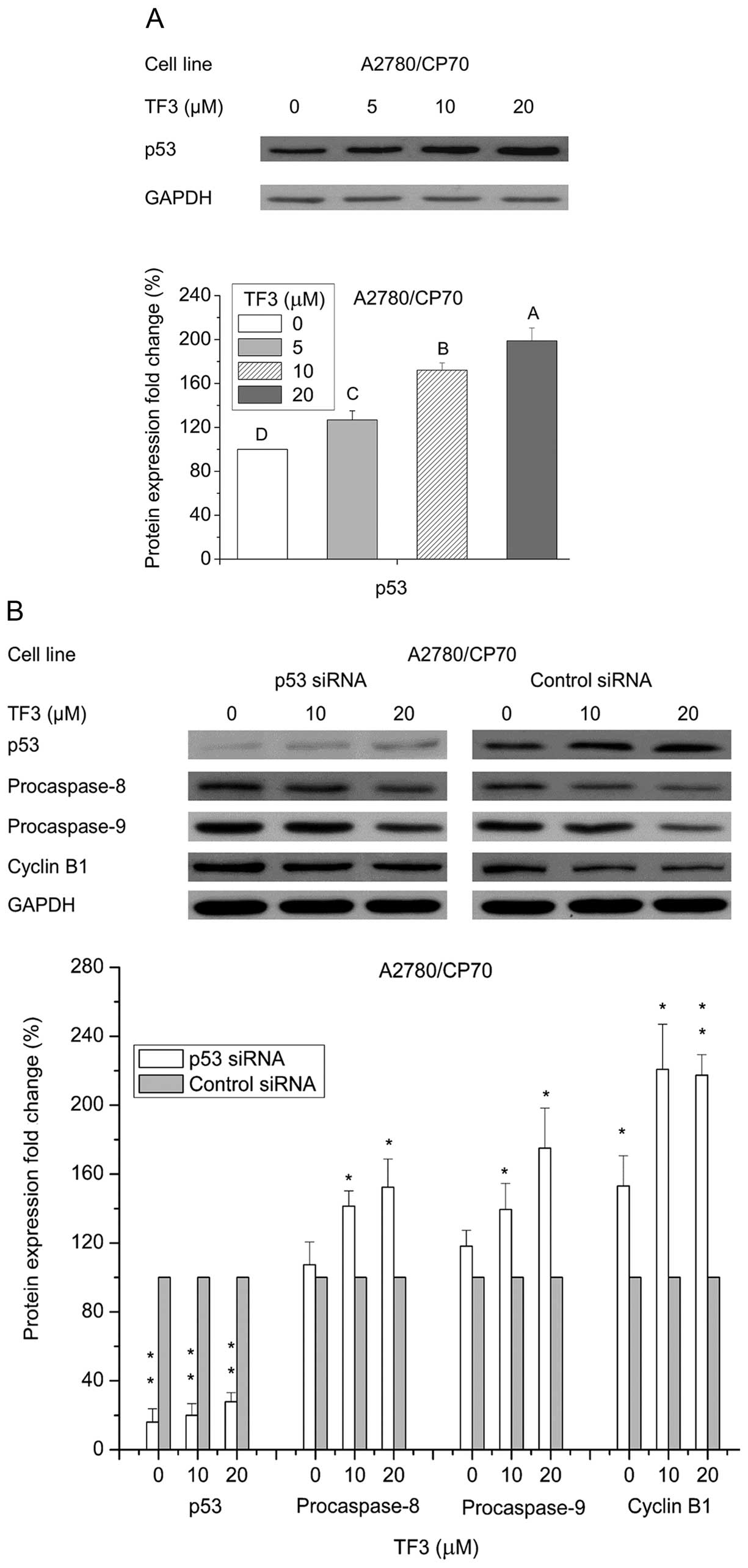
Role of p53 in TF3-induced apoptosis and
cell cycle arrest
The p53 signaling pathway is a well-known crucial
modulator of cell survival, apoptosis and cell cycle arrest
(18). To clarify the role of p53
in TF3-induced apoptosis and cell cycle arrest, p53 protein
expression was determined by western blotting. Fig. 8A shows that TF3 upregulated the
protein expression of p53 in A2780/CP70 cells (p<0.05). To
further analyze the role of p53 in TF3-induced cell death, p53 was
selectively knocked down by siRNA approach. Fig. 8B shows that p53 protein level was
obviously inhibited after treatment with 50 nM p53 siRNA
(p<0.01). This p53 depletion led to abrogation of TF3-induced
decrease in cyclin B1, procaspase-8 and procaspase-9 protein
expression (p<0.05). These results suggested that TF3 induced
apoptosis and G2 cell cycle arrest at least partly through p53 in
A2780/CP70 cells.
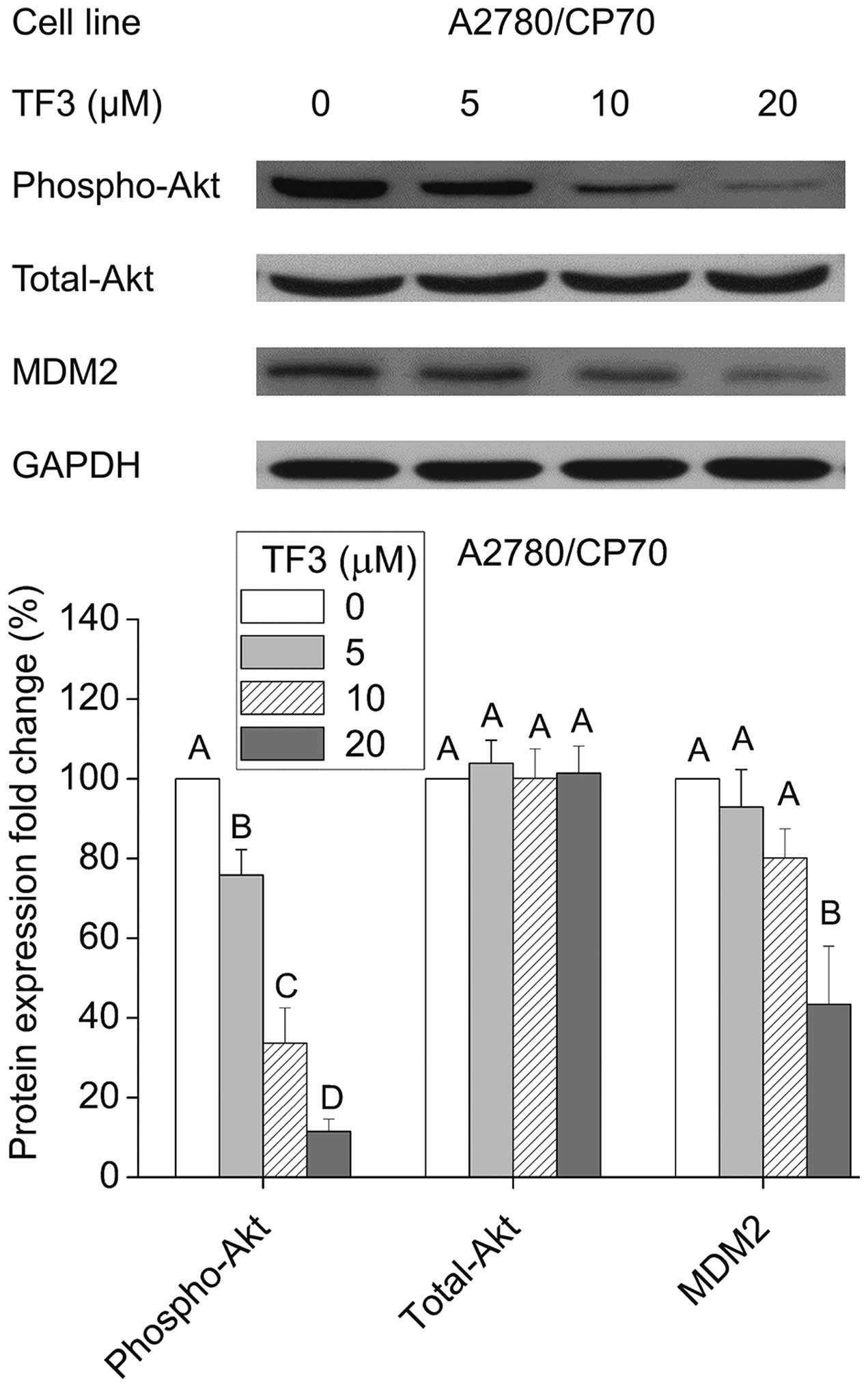
Regulation of phospho-Akt, total-Akt and
MDM2 protein expression by TF3
Akt is considered as a determinant of cisplatin
resistance in ovarian cancer cells (19). Murine double minute 2 (MDM2) is an
important negative regulator of p53 that directly inhibits p53
transcriptional activity and enhances p53 degradation via the
ubiquitin-proteasome pathway (20). Therefore, we tested the effects of
TF3 on the expression of Akt and its downstream effector MDM2. As
shown in Fig. 9, TF3 significantly
decreased the phosphorylation of Akt without the influence on total
Akt protein expression, and reduced the protein level of MDM2
(p<0.05).
Discussion
Although cisplatin-centered chemotherapy is the
first-line anticancer agent for human ovarian cancer,
chemoresistance and adverse side effects remain major hurdles to
successful treatment (21).
Natural products have recently received renewed focus as a rich
repository for drug discovery (22). (−)-Epigallocatechin-3-gallate
(EGCG), the most abundant catechin monomer in green tea, was
reported to potently suppress ovarian cancer cell growth alone, and
potentiate the inhibiting effect of sulforaphane on
paclitaxel-resistant ovarian cancer cell lines (23). TF3 is the oxidation product of
(−)-epicatechin gallate (ECG) and EGCG, and has been found to
possess stronger antioxidant and anti-inflammation activities
compared with EGCG (8,24). These findings stimulated our
interest in investigating the effect of TF3 on the ovarian cancer
cells.
In this study, we found that the IC50
value of TF3 for A2780/CP70 was 23.81 μM, which was around half of
the IC50 (59.58 μM) for TF3 in the IOSE-364 cells
(Fig. 2), and that (47.9 μM) for
cisplatin in the A2780/CP70 cells as reported in our previous study
(25). Flow cytometry analysis and
caspase-3/7 activity assay suggested that TF3 had more potential to
induce apoptosis and G2 cell cycle arrest of A2780/CP70 cells with
respect to IOSE-364 cells (Figs. 3
and 4), which provided a possible
explanation for the preferential cell growth inhibitory effect by
TF3.
A decreased susceptibility of ovarian cancer to
apoptosis was strongly associated with drug resistance (26). Agents that target the apoptotic
pathway have been shown to sensitize tumor cells to chemotherapy
and radiotherapy (27). Apoptosis
can be initiated via the intrinsic and extrinsic pathways, which
both ultimately activate the same effector caspases (3, 6 and 7)
and apoptosis effector molecules. The intrinsic apoptosis pathway
is triggered by intracellular signals such as cellular and DNA
damage. Key events are the depolarization of the mitochondrial
membrane potential and the release of mitochondrial factors such as
cytochrome c into the cytosol, which results in activation
of initiator caspase-9. Mitochondrial activation is critically
controlled by the family of Bcl-2 proteins, which consists of pro-
and anti-apoptotic members (28).
The extrinsic pathway implicates the activation of death receptors
at the plasma membrane level. Tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) interacts with the death
domain-containing receptors DR4/DR5 or Fas, resulting in the
recruitment of Fas-associated death domain (FADD), procaspase-8,
and procaspase-10 to form the death-inducing signaling complex
(DISC) (29). It was reported that
the differential induction of apoptosis in cancer versus normal
cells involves direct targeting of mitochondria associated with
alterations in the balance of Bcl-2 proteins (30). Another explanation is that normal
cells are resistant to the tumor necrosis factor related
apoptosis-inducing ligand (TRAIL) due to the high TRID levels,
whereas tumor lines carrying TRAIL-receptors in most cases can not
be protected (31). Theaflavins
have been reported to target the Fas/caspase-8 pathway to induce
apoptosis in human breast cancer cells, and influence Bcl-2 protein
expression in an experimental lung cancer mouse model (32,33).
In this study, treatment with TF3 significantly regulated the
protein expression of Bcl-2 family proteins (Bax, Bad and Bcl-xL),
DR5, FADD, caspase-8 and -9 (Figs.
5 and 6), indicating that TF3
induced apoptosis through both intrinsic and extrinsic pathways in
A2780/CP70 cells. Induction of apoptosis might account for the
preferential inhibitory effect of TF3 on ovarian cancer A2780/CP70
cells vs. normal ovarian IOSE-364 cells.
Cancer is a disease of inappropriate cell
proliferation. It was revealed that many cancer cells have
defective G1 checkpoint mechanisms, and depend on G2 checkpoint
during cell replication far more than normal cells. These insights
have given birth to the idea of cell cycle G2 checkpoint abrogation
as a cancer cell specific therapy (34). Our cell cycle analysis showed that
TF3 induced more prominent G2 arrest in A2780/ CP70 cells than in
IOSE-364 cells (Fig. 4 and
Table I), suggesting that in
addition to apoptosis, G2 cell cycle arrest was another factor that
contributed to the preferential cell growth inhibition. Transition
through G1 to S phase is regulated by the cyclin A-Cdk2 and cyclin
E-Cdk2 complexes. Inactivation of cyclin B-Cdc2 leads to the G2/M
arrest (35). The p21 protein is a
potent cyclin-dependent kinase inhibitor, and functions as a
regulator of cell cycle progression at G1 and G2 phase (36). A previous study showed that
theaflavins induced G2/M arrest by modulating expression of p21,
cdc25C and cyclin B in human prostate carcinoma PC-3 cell (37). However, our data showed that TF3
could not influence the p21, phospho-cdc2 (Tyr15) and total-cdc2
protein levels, but significantly decreased cyclin B1 expression
(Fig. 7), indicating cyclin B1
played an important role in the TF3-induced G2 arrest of A2780/CP70
cells.
The tumor suppressor protein, p53, transcriptionally
activates genes that control cell cycle arrest, apoptosis, and
other cellular processes that help to prevent tumor development.
p53 stimulates apoptosis by a wide network of signals including the
extrinsic pathway such as Killer/DR5 and Fas/APO-1, and intrinsic
pathway including proteins Bax, Puma and Noxa (38). In addition, p53 contributes to cell
cycle arrest, blocking the cell at the G1, S or G2/M phase by
inducing expression of p21 and the consequent inhibition of cyclin
D/Cdks, decreasing cyclin B1 transcription and synthesis, or
inactivates Cdc2 through Cdc25C (39). It is of particular interest in
ovarian cancer in view of the association of platinum sensitivity
and p53 pathway alterations. However, wild-type p53 status is
often, but not always, associated with cisplatin sensitivity
(40).
It is known that A2780/CP70 cell line has a
wild-type p53 gene sequence (41).
In this study, we found that TF3 significantly increased the p53
protein expression (Fig. 8A).
Furthermore, transfection with the p53 siRNA tended to attenuate
the reduction of procaspase-8, 9 and cyclin B1 protein expression
induced by TF3 (Fig. 8). These
results suggested that p53 played an important role in TF3 induced
apoptosis and G2 cell cycle arrest of A2780/CP70 cells, which was
consistent with the previous reports that p53 promoted
theaflavin-induced apoptosis in human breast and prostate cancer
cells through mitochondrial death cascade (42,43),
and prevented G2/M transition by decreasing intracellular levels of
cyclin B1 protein in ovarian cancer Ts-SKOV3 cells (44).
Akt is a serine/threonine kinase that plays a
critical role in the malignant transformation of human tumors and
their subsequent growth, proliferation, and metastasis. Targeted
inhibition of the Akt pathway is a promising strategy for cancer
therapy and can also be useful for overcoming chemotherapy
resistance (45). The functional
link between Akt-mediated chemoresistance and p53 has been found.
It was reported that Akt could induce an MDM2-dependent
downregulation of p53, and suppression of Akt sensitized
chemoresistant cells to cisplatin in a p53-dependent manner
(46). Our data showed that TF3
treatment significantly decreased the phosphorylation of Akt and
MDM2 protein level, indicating that TF3 might upregulate p53
expression through inactivation of Akt and MDM2 in A2780/CP70
cells. Further in vivo studies in animal models and human
patients are necessary to ascertain the anticancer effect of
theaflavins against ovarian cancer.
In conclusion, this study demonstrated that TF3 had
a potent and preferential cell growth inhibitory effect on the
cisplatin-resistant ovarian cancer cell line A2780/CP70 with
respect to normal cell line IOSE-364 via apoptosis and G2 cell
cycle arrest. TF3 induced apoptosis through both the intrinsic and
extrinsic pathways, and caused G2 cell cycle arrest via cyclin B1
in A2780/CP70 cells. These activities were at least partly mediated
by p53 upregulation dependent of Akt and MDM2. Based on these
results, TF3 would be a promising natural compound for prevention
and treatment of platinum-resistant ovarian cancer.
Acknowledgements
We thank Dr Kathy Brundage from the Flow Cytometry
Core at West Virginia University for providing technical help on
apoptosis and cell cycle analysis. We acknowledge financial support
from the West Virginia Higher Education Policy Commission/ Division
of Science Research, the National Natural Science Foundation of
China (grant no. 31501474), the Natural Science Foundation of
Zhejiang Province (grant no. LY15C200007), the Major Project of
Hubei Province for Science and Technology Development (grant no.
2013ABC002), and the Agricultural Science and Technology
Independent Innovation Foundation in Jiangsu Province [grant no.
CX(14)2122]. This study was also
supported by NIH grants P20RR016477 from the National Center for
Research Resources and P20GM103434 from the National Institute for
General Medical Sciences (NIGMS) awarded to the West Virginia IDeA
Network of Biomedical Research Excellence.
References
|
1
|
McLean K, VanDeVen NA, Sorenson DR, Daudi
S and Liu JR: The HIV protease inhibitor saquinavir induces
endoplasmic reticulum stress, autophagy, and apoptosis in ovarian
cancer cells. Gynecol Oncol. 112:623–630. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee RX, Li QQ and Reed E: β-elemene
effectively suppresses the growth and survival of both
platinum-sensitive and -resistant ovarian tumor cells. Anticancer
Res. 32:3103–3113. 2012.PubMed/NCBI
|
|
3
|
Yang YI, Kim JH, Lee KT and Choi JH:
Costunolide induces apoptosis in platinum-resistant human ovarian
cancer cells by generating reactive oxygen species. Gynecol Oncol.
123:588–596. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wahl H, Tan L, Griffith K, Choi M and Liu
JR: Curcumin enhances Apo2L/TRAIL-induced apoptosis in
chemoresistant ovarian cancer cells. Gynecol Oncol. 105:104–112.
2007. View Article : Google Scholar
|
|
5
|
Buolamwini JK: Cell cycle molecular
targets in novel anticancer drug discovery. Curr Pharm Des.
6:379–392. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diaz-Moralli S, Tarrado-Castellarnau M,
Miranda A and Cascante M: Targeting cell cycle regulation in cancer
therapy. Pharmacol Ther. 138:255–271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo H, Rankin GO, Li Z, Depriest L and
Chen YC: Kaempferol induces apoptosis in ovarian cancer cells
through activating p53 in the intrinsic pathway. Food Chem.
128:513–519. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sarkar A and Bhaduri A: Black tea is a
powerful chemopreventor of reactive oxygen and nitrogen species:
Comparison with its individual catechin constituents and green tea.
Biochem Biophys Res Commun. 284:173–178. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koňariková K, Ježovičová M, Keresteš J,
Gbelcová H, Ďuračková Z and Žitňanová I: Anticancer effect of black
tea extract in human cancer cell lines. Springerplus. 4:1272015.
View Article : Google Scholar
|
|
10
|
Qian Y, Zhu K, Wang Q, Li G and Zhao X:
Antimutagenic activity and preventive effect of black tea on buccal
mucosa cancer. Oncol Lett. 6:595–599. 2013.PubMed/NCBI
|
|
11
|
Cassidy A, Huang T, Rice MS, Rimm EB and
Tworoger SS: Intake of dietary flavonoids and risk of epithelial
ovarian cancer. Am J Clin Nutr. 100:1344–1351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin D, Xu Y, Mei X, Meng Q, Gao Y, Li B
and Tu Y: Antiobesity and lipid lowering effects of theaflavins on
high-fat diet induced obese rats. J Funct Foods. 5:1142–1150. 2013.
View Article : Google Scholar
|
|
13
|
Li B, Vik SB and Tu Y: Theaflavins inhibit
the ATP synthase and the respiratory chain without increasing
superoxide production. J Nutr Biochem. 23:953–960. 2012. View Article : Google Scholar
|
|
14
|
Bhattacharya U, Halder B, Mukhopadhyay S
and Giri AK: Role of oxidation-triggered activation of JNK and p38
MAPK in black tea polyphenols induced apoptotic death of A375
cells. Cancer Sci. 100:1971–1978. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Adhikary A, Mohanty S, Lahiry L, Hossain
DM, Chakraborty S and Das T: Theaflavins retard human breast cancer
cell migration by inhibiting NF-kappaB via p53-ROS cross-talk. FEBS
Lett. 584:7–14. 2010. View Article : Google Scholar
|
|
16
|
Schuck AG, Ausubel MB, Zuckerbraun HL and
Babich H: Theaflavin-3,3′-digallate, a component of black tea: An
inducer of oxidative stress and apoptosis. Toxicol In Vitro.
22:598–609. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang G, Miura Y and Yagasaki K:
Suppression of adhesion and invasion of hepatoma cells in culture
by tea compounds through antioxidative activity. Cancer Lett.
159:169–173. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sullivan KD, Gallant-Behm CL, Henry RE,
Fraikin JL and Espinosa JM: The p53 circuit board. Biochim Biophys
Acta. 1825.229–244. 2012.
|
|
19
|
Levine AJ, Hu W and Feng Z: The P53
pathway: What questions remain to be explored? Cell Death Differ.
13:1027–1036. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kao CL, Hsu HS, Chen HW and Cheng TH:
Rapamycin increases the p53/MDM2 protein ratio and p53-dependent
apoptosis by translational inhibition of mdm2 in cancer cells.
Cancer Lett. 286:250–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abedini MR, Muller EJ, Bergeron R, Gray DA
and Tsang BK: Akt promotes chemoresistance in human ovarian cancer
cells by modulating cisplatin-induced, p53-dependent ubiquitination
of FLICE-like inhibitory protein. Oncogene. 29:11–25. 2010.
View Article : Google Scholar
|
|
22
|
Miura K, Satoh M, Kinouchi M, Yamamoto K,
Hasegawa Y, Kakugawa Y, Kawai M, Uchimi K, Aizawa H, Ohnuma S, et
al: The use of natural products in colorectal cancer drug
discovery. Expert Opin Drug Discov. 10:411–426. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen H, Landen CN, Li Y, Alvarez RD and
Tollefsbol TO: Epigallocatechin gallate and sulforaphane
combination treatment induce apoptosis in paclitaxel-resistant
ovarian cancer cells through hTERT and Bcl-2 down-regulation. Exp
Cell Res. 319:697–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang Z, Jie G, Dong F, Xu Y, Watanabe N
and Tu Y: Radical-scavenging abilities and antioxidant properties
of theaflavins and their gallate esters in
H2O2-mediated oxidative damage system in the
HPF-1 cells. Toxicol In Vitro. 22:1250–1256. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li B, Gao Y, Rankin GO, Rojanasakul Y,
Cutler SJ, Tu Y and Chen YC: Chaetoglobosin K induces apoptosis and
G2 cell cycle arrest through p53-dependent pathway in
cisplatin-resistant ovarian cancer cells. Cancer Lett. 356(2 Pt B):
418–433. 2015. View Article : Google Scholar :
|
|
26
|
Hu W, Wang F, Tang J, Liu X, Yuan Z, Nie C
and Wei Y: Proapoptotic protein Smac mediates apoptosis in
cisplatin-resistant ovarian cancer cells when treated with the
anti-tumor agent AT101. J Biol Chem. 287:68–80. 2012. View Article : Google Scholar :
|
|
27
|
Wilson TR, Johnston PG and Longley DB:
Anti-apoptotic mechanisms of drug resistance in cancer. Curr Cancer
Drug Targets. 9:307–319. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brunelle JK and Letai A: Control of
mitochondrial apoptosis by the Bcl-2 family. J Cell Sci.
122:437–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim SY, Lee DH, Song X, Bartlett DL, Kwon
YT and Lee YJ: Role of Bcl-xL/Beclin-1 in synergistic apoptotic
effects of secretory TRAIL-armed adenovirus in combination with
mitomycin C and hyperthermia on colon cancer cells. Apoptosis.
19:1603–1615. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu T, Hannafon B, Gill L, Kelly W and
Benbrook D: Flex-Hets differentially induce apoptosis in cancer
over normal cells by directly targeting mitochondria. Mol Cancer
Ther. 6:1814–1822. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ozören N and El-Deiry WS: Cell surface
death receptor signaling in normal and cancer cells. Semin Cancer
Biol. 13:135–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lahiry L, Saha B, Chakraborty J, Adhikary
A, Mohanty S, Hossain DM, Banerjee S, Das K, Sa G and Das T:
Theaflavins target Fas/caspase-8 and Akt/pBad pathways to induce
apoptosis in p53-mutated human breast cancer cells. Carcinogenesis.
31:259–268. 2010. View Article : Google Scholar
|
|
33
|
Saha P, Banerjee S, Ganguly C, Manna S,
Panda CK and Das S: Black tea extract can modulate protein
expression of H-ras, c-Myc, p53, and Bcl-2 genes during pulmonary
hyperplasia, dysplasia, and carcinoma in situ. J Environ Pathol
Toxicol Oncol. 24:211–224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kawabe T: G2 checkpoint abrogators as
anticancer drugs. Mol Cancer Ther. 3:513–519. 2004.PubMed/NCBI
|
|
35
|
Duiker EW, Meijer A, van der Bilt AR,
Meersma GJ, Kooi N, van der Zee AG, de Vries EG and de Jong S:
Drug-induced caspase 8 upregulation sensitises cisplatin-resistant
ovarian carcinoma cells to rhTRAIL-induced apoptosis. Br J Cancer.
104:1278–1287. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu W, Park SK, Jia L, Tiwary R, Scott WW,
Li J, Wang P, Simmons-Menchaca M, Sanders BG and Kline K:
RRR-gamma-tocopherol induces human breast cancer cells to undergo
apoptosis via death receptor 5 (DR5)-mediated apoptotic signaling.
Cancer Lett. 259:165–176. 2008. View Article : Google Scholar
|
|
37
|
Prasad S, Kaur J, Roy P, Kalra N and
Shukla Y: Theaflavins induce G2/M arrest by modulating expression
of p21waf1/cip1, cdc25C and cyclin B in human prostate carcinoma
PC-3 cells. Life Sci. 81:1323–1331. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kuo YC, Kuo PL, Hsu YL, Cho CY and Lin CC:
Ellipticine induces apoptosis through p53-dependent pathway in
human hepatocellular carcinoma HepG2 cells. Life Sci. 78:2550–2557.
2006. View Article : Google Scholar
|
|
39
|
Pucci B, Kasten M and Giordano A: Cell
cycle and apoptosis. Neoplasia. 2:291–299. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Green JA, Berns EM, Coens C, van Luijk I,
Thompson-Hehir J, van Diest P, Verheijen RH, van de Vijver M, van
Dam P, Kenter GG, et al; EORTC Gynaecological Cancer Group.
Alterations in the p53 pathway and prognosis in advanced ovarian
cancer: A multi-factorial analysis of the EORTC Gynaecological
Cancer group (study 55865). Eur J Cancer. 42:2539–2548. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brown R, Clugston C, Burns P, Edlin A,
Vasey P, Vojtĕsek B and Kaye SB: Increased accumulation of p53
protein in cisplatin-resistant ovarian cell lines. Int J Cancer.
55:678–684. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lahiry L, Saha B, Chakraborty J,
Bhattacharyya S, Chattopadhyay S, Banerjee S, Choudhuri T, Mandal
D, Bhattacharyya A, Sa G, et al: Contribution of p53-mediated Bax
transactivation in theaflavin-induced mammary epithelial carcinoma
cell apoptosis. Apoptosis. 13:771–781. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kalra N, Seth K, Prasad S, Singh M, Pant
AB and Shukla Y: Theaflavins induced apoptosis of LNCaP cells is
mediated through induction of p53, down-regulation of NF-kappa B
and mitogen-activated protein kinases pathways. Life Sci.
80:2137–2146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Innocente SA, Abrahamson JL, Cogswell JP
and Lee JM: p53 regulates a G2 checkpoint through cyclin B1. Proc
Natl Acad Sci USA. 96:2147–2152. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mabuchi S, Kuroda H, Takahashi R and
Sasano T: The PI3K/ AKT/mTOR pathway as a therapeutic target in
ovarian cancer. Gynecol Oncol. 137:173–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim CW, Lu JN, Go SI, Jung JH, Yi SM,
Jeong JH, Hah YS, Han MS, Park JW, Lee WS, et al: p53 restoration
can overcome cisplatin resistance through inhibition of Akt as well
as induction of Bax. Int J Oncol. 43:1495–1502. 2013.PubMed/NCBI
|