Introduction
A recent article (2017) of the American Cancer
Society presenting key statistics for cutaneous melanoma reported
that among all types of malignancies, skin cancer is the most
common (1). Although squamous cell
carcinoma (SCC) and basal cell carcinoma (BCC) are more frequent,
melanoma, the so-called cancer of the Western world, accounts for
approximately 1% among all types of skin cancers. The
aggressiveness of this particular skin pathology is due to its
highly metastatic potential and increased resistance to
chemotherapeutic agents, events associated with a low survival of
affected patients (2,3). When localized to the primary site,
the chances of cure are high; however, after it develops and
reaches the lymph nodes, the prognosis becomes poor, with the
5-year survival rate being around 29%. The marked decrease in
survival presents once the major organs are invaded by malignant
cells, with the 5-year survival rate reaching only about 7%
(4). As regards therapeutic
strategies, the treatment approach is straightforward in the early
stages of the disease when the tumor is localized, and involves the
wide excision of the primary tumor site. The challenges in
treatment strategies present once metastasis occurs. Chemotherapy,
radiation therapy, immunotherapy and targeted therapy represent
current standard approaches for the management of this highly
aggressive disease, with varying results (5–7).
The high mortality rates associated with the disease
are due to its heterogenous molecular pattern based on multiple
progressive mutations that can occur. The most frequent subtype of
malignant melanoma, found in 40–50% of cases, harbors the oncogene
mutation, B-RAF V600, which is related to sun exposure. It is
followed by a mutation of the oncogene N-RAS, detected in 15–20% of
patients. However, although this mutation exhibits significant
frequency, its' effect has not yet been widely studied and the
response to the few treatment options is relatively poor (8–10).
Based on these data, in this study, in order to investigate
possible targeted new therapeutics, we decided to investigate the
SK-MEL-2 melanoma cell line that harbors two mutant genes, N-RAS
and TP53.
Currently, for the development of treatment options,
natural compounds are intensively investigated for their
chemopreventive and anti-invasive potential, in addition to lower
secondary effects (11,12). Angiogenesis, the process through
which new blood vessels are formed from pre-existing ones, is one
of the key steps in tumor growth and metastasis. This process is
being highly investigated as a possible therapeutic approach in
cancer, after Folkman et al (13) introduced the concept of tumor
angiogenesis.
The discovery of novel drugs from natural sources
targeting cancer and angiogenesis was first based on the
traditional practices of treating vascular-dependent pathologies
(14). As an unbalanced type of
diet is increasingly associated with cancer pathologies, a growing
number of natural compounds found in healthy diet foods is being
evaluated as anticancer agents (15). An important class of phytochemicals
with demonstrated anticancer potential is represented by the
triterpenoids. Triterpenoid compounds are secondary metabolites
widely distributed in plants. Structurally, they are formed by 30
atoms of carbon, organized in isoprene units. Important
triterpenoids are derivatives of the pentacyclic carbon skeleton,
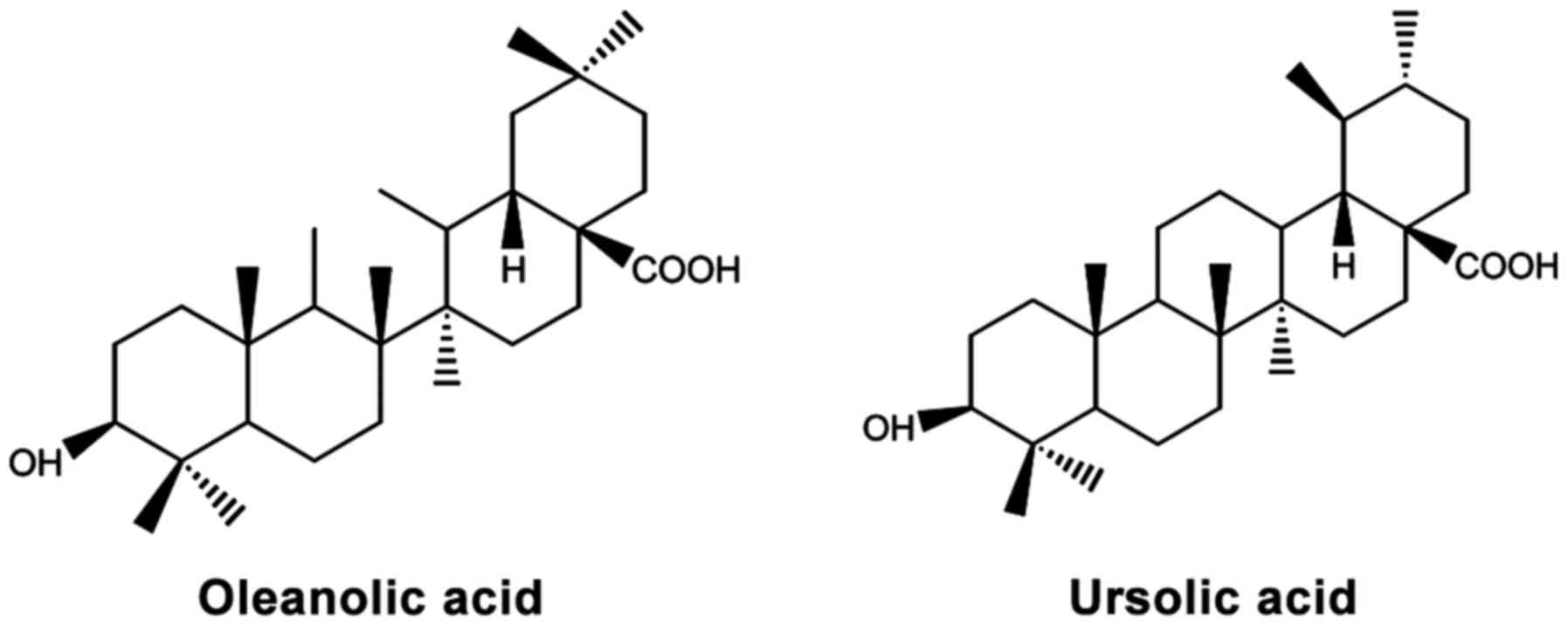
including lupane (e.g., betulinic acid), oleane [e.g., oleanolic
acid (OA)] and ursane [e.g., ursolic acid (UA)]. Several
representatives are known for their potential therapeutic benefits
as antioxidant, anti-inflammatory, anti-bacterial, anti-malaria and
anti-viral agents. Importantly, anticancer properties have been
attributed to these compounds in various types of cancer cell
lines, in which they have been shown toexert anti-proliferative,
pro-apoptotic and tumor anti-invasive effects (16–19).
Among the triterpenoids OA
(3-beta-3-hydroxy-olean-12- ene-28-oic-acid) and its isomer, UA
(3-beta-3-hydroxy-urs-12- ene-28-oic-acid) (Fig. 1), are promising therapeutic
candidates. They are highly abundant in edible plant foods, such as
apples, pears, olives or aromatic plants from the Lamiaceae family,
such as oregano, basil, rosemary or lemon balm (20–23).
The pharmacological value of the two compounds is demonstrated both
by the multiple pharmacological targets, but also by their low
toxicity (24,25). To date, the various pharmacological
effects of UA and OA exerted via multiple mechanisms are not yet
fullycompletely understood. Thus, they are the subject of current
research.
Despite structural similarities, the effectiveness
of their anticancer activity differs. Some studies have reported an
increased antitumor activity both in vitro and in
vivo for UA, but OA has also been assigned with anticancer
properties (26,27). Hence, the selection of one of the
two triterpenic compounds for cancer chemoprevention should be
carried out taking into account the involved cancerous cell line
and/or target molecules engaged in each type of cancer (28). Some available data suggest the
potential benefits of OA and UA in the management of melanoma.
In vitro, various IC50/EC50 values for
OA and UA have been obtained for several melanoma cell lines
(29–31).
MAP/extracellular signal-regulated kinase (MEK)
protein kinases form a family of proteins that next to mammalian
target of rapamycin (mTOR), are involved in cell proliferation,
survival, differentiation and angiogenesis, and are considered
interesting synergistic targets for N-RAS mutated cancers (8). Modern approaches for the discovery of
active compounds involve the virtual screening of large natural
compound databases against druggable targets in cancer (32–34).
In this study, for the selection of the two triterpenic compounds,
we considered the availability from natural sources and their
promising anticancer activities. Moreover, recent studies have
indicated the possible role of UA and OA in modulating some of the
deregulated kinases involved in the progression of various types of
cancer (35,36). However, the mechanisms of action
and the potential effects in vivo in N-RAS-mutated melanoma
of the two compounds are not yet fully understood.
The tumor microenvironment, as regards
tumor-associated inflammation and angiogenesis, as well as specific
targets, has been intensively studied (18,19,37–39)
in order to elucidate the mechanism(s) of action of triterpenic
compounds. In the present study, we aimed to test the in
vitro and in vivo anti-invasive and anti-metastatic
activity of OA and UA to determine their possible use as
chemopreventive or therapeutic agents in melanoma. For the in
vitro experiments, the anti-proliferative activity of the
triterpenic compounds on SK-MEL-2 melanoma cells was examined. The
anti-invasive potential was assessed by examining the effects of
the active compounds on vascular cell adhesion molecule (VCAM) and
intercellular adhesion molecule (ICAM) adhesion to melanoma cells.
Normal and tumor angiogenesis was evaluated in vivo by
chicken embryo chorioallantoic membrane (CAM) assay.
Materials and methods
In vitro analysis
Cell culture
The SK-MEL-2 human melanoma cells (ATCC, Manassas,
VA, USA) were cultured in Eagle's minimum essential medium (EMEM),
containing 10% fetal bovine serum (FBS) (both from ATCC) and 1%
penicillin-streptomycin (Pen/Strep, 10,000 IU/ml; PromoCell,
Heidelberg, Germany). The cells were maintained in an atmosphere of
5% CO2 at 37°C.
In vitro cell proliferation
assay:AlamarBlue assay
The SK-MEL-2 cells were seeded in 96-well
microplates (5,000 cells/plate) and incubated overnight in order to
allow attachment of the cells to the bottom of the plate.
Subsequently, 150 µl of fresh EMEM medium containing UA or
OA was added and the cells were incubated for 48 h. UA or OA was
added at concentrations between 25 and 100 µM. After 48 h,
15 µl of AlamarBlue solution were added and the cells were
incubated for 4–10 h at 37°C. The samples were
spectrophotometrically analyzed at 570 and 600 nm using an xMark™
Microplate Spectrophotometer (Bio-Rad, Hercules, CA, USA). Wells
with untreated cells were used as controls. The test substances
were dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich,
Ayrshire, UK) and stock solutions were stored at 2–8°C. The final
concentrations were prepared by diluting the stock solution with
the EMEM growth medium. The highest DMSO concentration (0.1%) of
the medium did not exert any significant effect on cell
proliferation. The experiments were performed using microplates
with at least 4 parallel wells. The results are presented as the
mean value ± standard deviation. One-way ANOVA was used to
determine statistically signficant differences between various
experimental groups.
Cell cycle analysis
The SK-MEL-2 human melanoma cells were seeded in 25
cm2 plates (106 cells/plate) and treated with
UA and OA (30, 50, or 75 µM). After 48 h of treatment, the
cells were collected, fixed with cold 70% ethanol and stored for 30
min at 4°C. Following centrifugation (1,500 rpm, 22°C), cold PBS
was used to wash the cells. Subsequently, 50 µl of propidium
iodide (concentration, 50 µM) (BD Pharmingen; BD
Biosciences, San Diego, CA, USA) were added to the cells and the
cells were incubated for 10 min in the dark, in order to stain the
DNA. A FACSCalibur flow cytometer (Becton-Dickinson, Franklin
Lakes, NJ, USA) was used to perform the DNA content analysis. The
percentage of cells present in the different cell cycle phases was
determined using Modfit software.
Flowchamber assay
The SK-MEL-2 tumor cell lines were cultured
according to the protocol described above. The cells were treated
with 30 and 50 µM of UA. After 48 h of treatment, the cells
were harvested by trypsinization and the adhesive capacity of the
cells exposed to mechanical stress (shearstress) on the VCAM and
ICAM substrates (R&D Systems, Abingdon, UK) was evaluated using
the flow chamber method.
Flowchamber analysis follows several consecutive
steps beginning with the coating of the flowchamber channels with
adhesion molecules (VCAM and ICAM), used at a concentration of 2
µl/ml, 30 µl/channel 15 min prior to the beginning of
the experiment (3 channels for each adhesion molecule). The cells
were trypsinized and incubated while the peristaltic pump (Ismatec
- IPC High Precision Multichannel Dispenser (IDEX Corporation,
Glattbrugg, Switzerland) was prepared. The preparation of the
peristaltic pump consists of assembling the tubulature, passing the
HBSS medium (100 ml PBS with Ca2+ and Mg2+,
50 ml DMEM high glucose medium, 0.75 ml BSA 20%) through the tubes
and fixating the flowchamber to the microscope connected to the
camera. The tube is fixated in the 'OUT' position and to the
'waste' recipient. The supernatant in the 'IN' position is
discarded.
In continuation, 100 µl well omogenized
cellular suspension (105 cells) was inserted at the 'IN'
end of the channel, after which the tube connecting to the pump was
inserted. The cells were allowed 3 min to adhere to the substrate,
after which, using a Leica ICC50HD camera (Leica, Bucharest,
Romania) an image was acquired representing the START moment. After
that moment, shearstress was introduced following this schedule:
0.35 dyne/cm2 - 1 min; 2 dyne/cm2 - 30 sec; 5
dyne/cm2 - 30 sec; 8 dyne/cm2 - 30 sec; 15
dyne/cm2 - 30 sec. Following each time-speed period, an
image was acquired using a Leica ICC50HD camera and Leica DMD108
microscope (Leica). At the end of the experiment, all images were
analyzed in order to count the adhered cells at the initial moment
and the remaining cells throughout the experiment. The cell number
was correlated to the increase of shear stress. Variations of at
least 15% from the total number of cells were considered
significant in comparison to the control cells for the same flux
values.
In vivo analysis
CAM assay
Fertilized eggs (Gallus gallus domesticus),
obtained from a local poultry farm, were disinfected with 70°
ethanol, dated and subsequently incubated in a horizontal position,
at constant humidity and 37°C. On the third day of incubation, 3–4
ml of albumen was removed, so that the developing chorioallantoic
membrane could be detached from the eggshell, and the blood vessels
from the extra-embryonic vascular plexus could be easily observed.
On the 4th day of incubation, a window was cut, resealed with
adhesive tape, and set to incubate until the beginning of the
experimental procedures, as previously described (40).
Evaluation of angiogenesis and tumor
angiogenesis on the chorioallantoic membrane
The present study was conducted in ovo, beginning on
the 7th day of incubation of the chick embryos. The effect of UA
and OA on the process of angiogenesis of the chick embryo CAM was
firstly evaluated in the absence of tumor cells between the 7th and
the 11th embryonic days. During this interval, the vascular network
is in a rapid growing phase, and the endothelial cells exhibit a
high mitotic rate, similar to that of tumor-associated angiogenesis
(41). The samples were tested at
a concentration of 30 mM, using 3% DMSO. Three doses of the test
samples and blank solutions containing only 3% DMSO were applied in
triplicate daily for 5 days. Volumes of 5 ml were added inside a
plastic ring with a diameter of 5 mm previously placed on the CAM
surface in vascularized areas and the specimens were returned for
incubation. Evaluation was performed daily by means of a
stereomicroscope and relevant images were captured for further
analysis. Finally, on day 12 of incubation, the specimens were
sacrificed and the fine CAMs were harvested.
The SK-MEL-2 cells were cultured according to the
protocol described above. After harvesting the cells from the
culture plate through trypsinization, the cells were resuspended in
the culture medium until reaching the final concentration of
105/3 µl. On the 10th day of incubation, 3
µl of the SK-MEL-2 melanoma cell suspension were inoculated
inside a sterile ring previously placed on the CAM. The control
samples were only inoculated with 3 µl of cell culture
medium.
UA, OA and blank (DMSO) solutions were added in
volumes of 5 µl inside the rings 1 day after the inoculation
of the cells. Samples were applied daily for 5 days. The process
was dynamically examined and relevant captures were saved. On the
final day of the experiment, all the specimens were sacrificed and
prepared for further histological analysis.
Zeiss Axio V16 stereomicroscope was used for the in
ovo examination of the specimens. Images were registered by means
of the Zeiss Axio Cam equipment and image analysis was done using
Zeiss ZEN and Image J softwares. Morphometric analysis was applied
on the stereomicroscopic photographs of the tested CAMs, using an
arbitrary 0–5 scale that scores the intensity of vascular density
on the area of application. Low values are correlated with a
reduced angiogenic process, while the higher scores indicate an
activated process. The results are expressed as mean values ±
standard deviation. Statistical analysis was performed using SPSS
software (IBM SPSS Statistics for Windows, version 20.0; IBM Corp.,
Armonk, NY, USA).
Results
Effects of UA and OA on the proliferation
of SK-MEL-2 melanoma cells
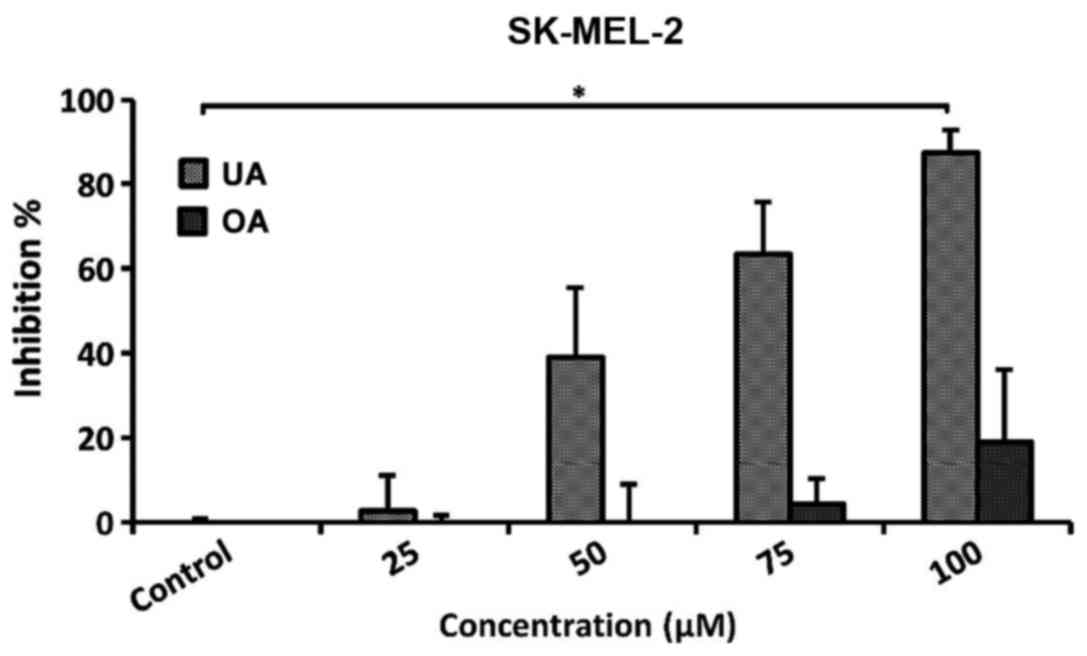
Cell proliferation experiments demonstrated that UA
exerted an inhibitory and dose-dependent effect on SK-MEL-2 human
melanoma cell growth (Fig. 2),
with an IC50 value of 58.43 µM. Specifically, the
lowest concentration presented an insignificant inhibitory index of
only 2.68±8.35%, with a gradual increase until reaching a value of
87.34±5.50% for the highest tested concentration (100 µM)
(p<0.05). As regards OA, a tendency of the inhibition of cell
proliferation was observed (Fig.
2), which did not however, reach statistical significance
(p=n.s.).
Effects of UA on SK-MEL-2 cell cycle
progression
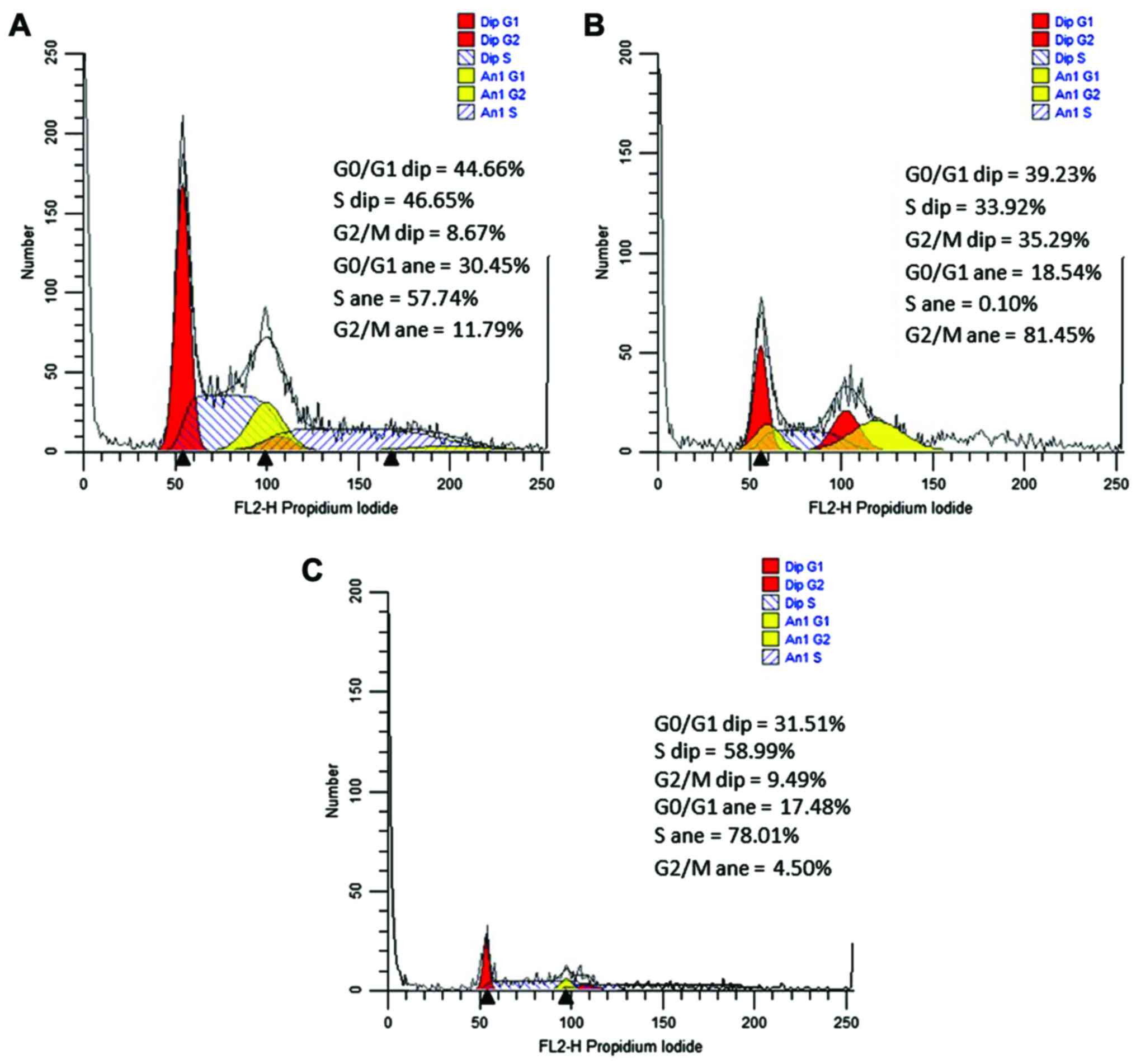
The aneuploid character of the SK-MEL-2 cells was
demonstrated utilizing flow cytometric analysis, which indicated
that the diploid cells represented 65% of the total number of cells
and the aneuploid cells represented 35% of the total cell
population. Therefore, the cell cycle passage of both the aneuploid
and diploid cell populations is separately presented and
demonstrated in Table I and
Fig. 3 (dip, diploid cells; ane,
aneuploidy cells). There was a significant shift in the cell cycle
in both the diplod and the aneploid cells as compared with the
controls. Of note, UA exerted effects on the cell cycle passage of
aneuploid and diploid cells in a distinct manner. Thus, at 30
µM, there was an even distribution of diploid cells among
the G0/G1, S and G2/M phase, whereas there was a massive
distribution of aneuploid cells in the G2/M phase. An interesting
fact is that for both the diploid and aneuploid populations, the
exposure to 50 µM UA resulted in S phase cell cycle arrest
(p<0.05) (Table I). Exposure of
the SK-MEL-2 cells to the concentration of 75 µM led to the
destruction of the cells and their detachment from the culture
plate, not allowing for cell cycle analysis.
 | Table IEffect of ursolic acid on cell cycle
phases in the SK-MEL-2 melanoma cells. |
Table I
Effect of ursolic acid on cell cycle
phases in the SK-MEL-2 melanoma cells.
Cell cycle
distribution (%)
|
|---|
| Treatment | SK-MEL-2 cells
|
|---|
| G0/G1 dip | S dip | G2/M dip | G0/G1 ane | S ane | G2/M ane |
|---|
| 0 | 44.66 | 46.65 | 8.67 | 30.45 | 57.74 | 11.79 |
| UA 30
µM | 39.23 | 33.92 | 35.29 | 18.54 | 0.10 | 81.45 |
| UA 50
µM | 31.51 | 58.99a | 9.49 | 17.48 | 78.01a | 4.50 |
Effects of UA on SK-MEL-2 melanoma cell
adhesion in the flow chamber assay
The flow chamber method represents an in
vitro model which simulates the exposure of cells to the
dynamical flux of fluids in a physiological environment (42). During the assay, the cells are
subjected to mechanical stress (shear stress) of fluids. The method
allows for the evaluation of theadhesive capacity to key molecules
correlated with cancer progression, such as ICAM-1 and V-CAM 1, and
enables the quantification of the cancer metastasis patterns of
tumor cells (43,44).
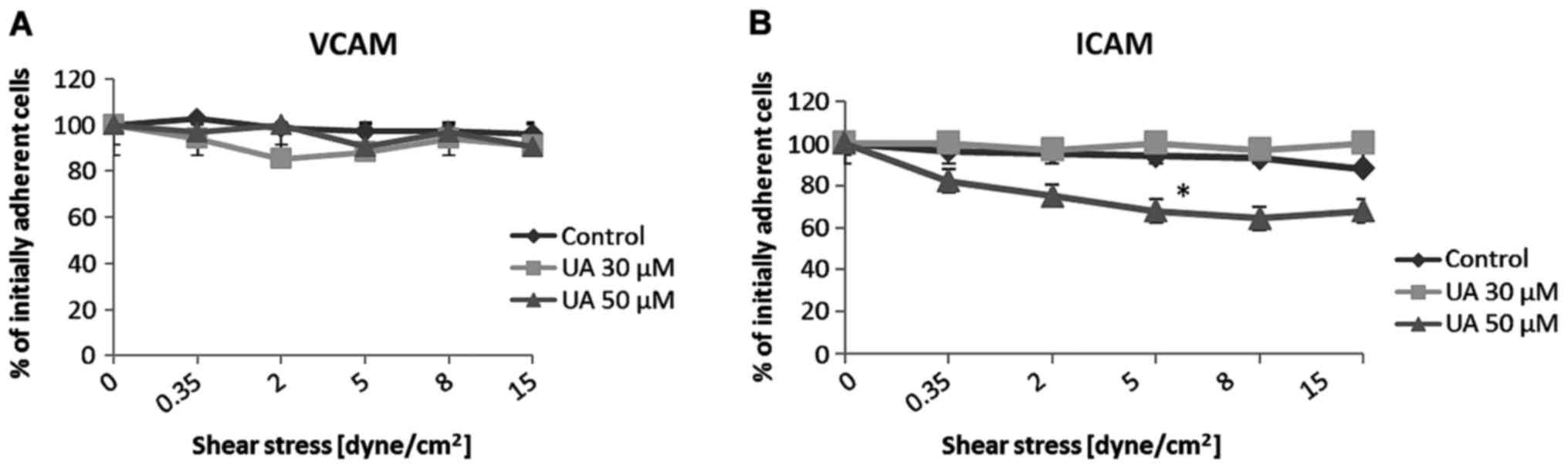
As shown in Fig.
4A, flow chamber analysis of the SK-MEL-2 cells demonstrated
that the untreated cells remained attached to the VCAM substrate at
the end of progressive exposure to shear stress at a percentage of
95.90%. Data collected for the cells exposed to UA demonstrated
slight alterations in the cell adhesive capacity (Fig. 4A). These alterations were not
considered significant and, upon completing the experiment, a
difference of only 4.70% compared with the adhesive ability of the
controls was shown for a concentration of 30 µM and only
5.30% for a concentration of 50 µM UA.
Likewise, treatment with 30 µM UA did not
affect the ability of the cells to attach to the ICAM substrate
(p=n.s.) (Fig. 4B). Significant
changes in cell adhesive ability were noted for the cells treated
with 50 µM UA. Indeed, upon the increase in shear stress
(from 0.35 dyne/cm2 to 15 dyne/cm2) there was
a proportional decrease in the number of attached cells with a
difference of 24% lower cell number as compared with the controls
(p<0.05).
Effects of UA and OA on angiogenesis
utilizing CAM assay
The anti-angiogenic potential of UA and OA was
assessed in an in vivo protocol by CAM assay. Normal
angiogenesis during the high mitotic phase of endothelial cells and
tumor-associated angiogenesis were investigated using the SK-MEL-2
melanoma cells. The evaluation was performed in ovo by means of
stereomicroscopy.
The chicken embryo specimens (inoculated or not with
tumor cells) exhibited good viability and survival rates with the
test compounds, which were similar to those of the controls, e.g.,
around ED13 for all specimens. For the assessment of normal
angiogenesis, modifications of the vascular plexus were observed
throughout the experiment, and a gradual increase in the effects
was noted, as shown in Fig. 5A–F.
After 5 days of treatment, relevant images indicating the affected
vessel architecture were registered and analyzed. Both OA and UA
induced changes in the vascular network inducing a decrease in
vessel number inside the application area, as compared with the
blank specimens. A more prominent effect was noted with OA, which
induced a reduction in vascular density and several areas with very
low number of fine capillaries were noted inside the ring (Fig. 5E and F).
 | Figure 5Effects of UA and OA on CAM assay.
(A-C) Normal angiogenesis, with two doses of the test samples on
ED9. (A) Blank (DMSO, 3%), (B) UA, (C) OA. (D-F) Normal
angiogenesis, with four doses of the test samples on ED11. (D)
Blank (DMSO, 3%), (E) UA, (F) OA. (G-I) SK-MEL-2 melanoma cells,
with four doses of the test samples on ED13. (G) Control (cell
medium), (H) SK-MEL-2 + UA, (I) SK-MEL-2 + OA. UA, ursolic acid;
OA, oleanolic acid; CAM, chorioallantoic membrane; DMSO, dimethyl
sulfoxide. |
An investigation of the angiogenic modulatory
effects of the two triterpenoid compounds was also conducted in a
melanoma model upon SK-MEL-2 cell inoculation to the CAM. The
process was stereomicroscopically examined from the first day of
the triterpene application onto previously inoculated cells to the
CAM. After 4 doses of triterpene solutions, relevant changes in the
vascular arrangement surrounding the tumor cell implants were
noted, accompanied by some alterations in the process of tumor
growth. In correlation with the effects on the normal developing
vascular plexus, OA exerted a significant effect on
melanoma-associated angiogenesis, which seemed to have contributed
to the growth of the tumor area inside the ring (Fig. 5I). The angiogenic reaction in the
OA-treated CAMs was clearly reduced compared to both the control
and the UA samples (Fig. 5G–I).
The 'spokes wheel' type of vascular reaction can be observed
converging towards the tumor site for the control and UA samples,
but not for cells treated with OA (Fig. 5I). Conversely, UA did not severely
impair the tumor-associated capillary density, the growth of
melanoma cells was not inhibited (Fig.
5H), and the invasiveness of the melanoma cells was higher.
Although the anti-angiogenic effects were more potent with OA than
with UA, the metastatic potential was not inhibited with either
agent. For the applied experimental conditions, the two triterpenes
influenced, but did not inhibit the invasiveness of the SK-MEL-2
melanoma cells.
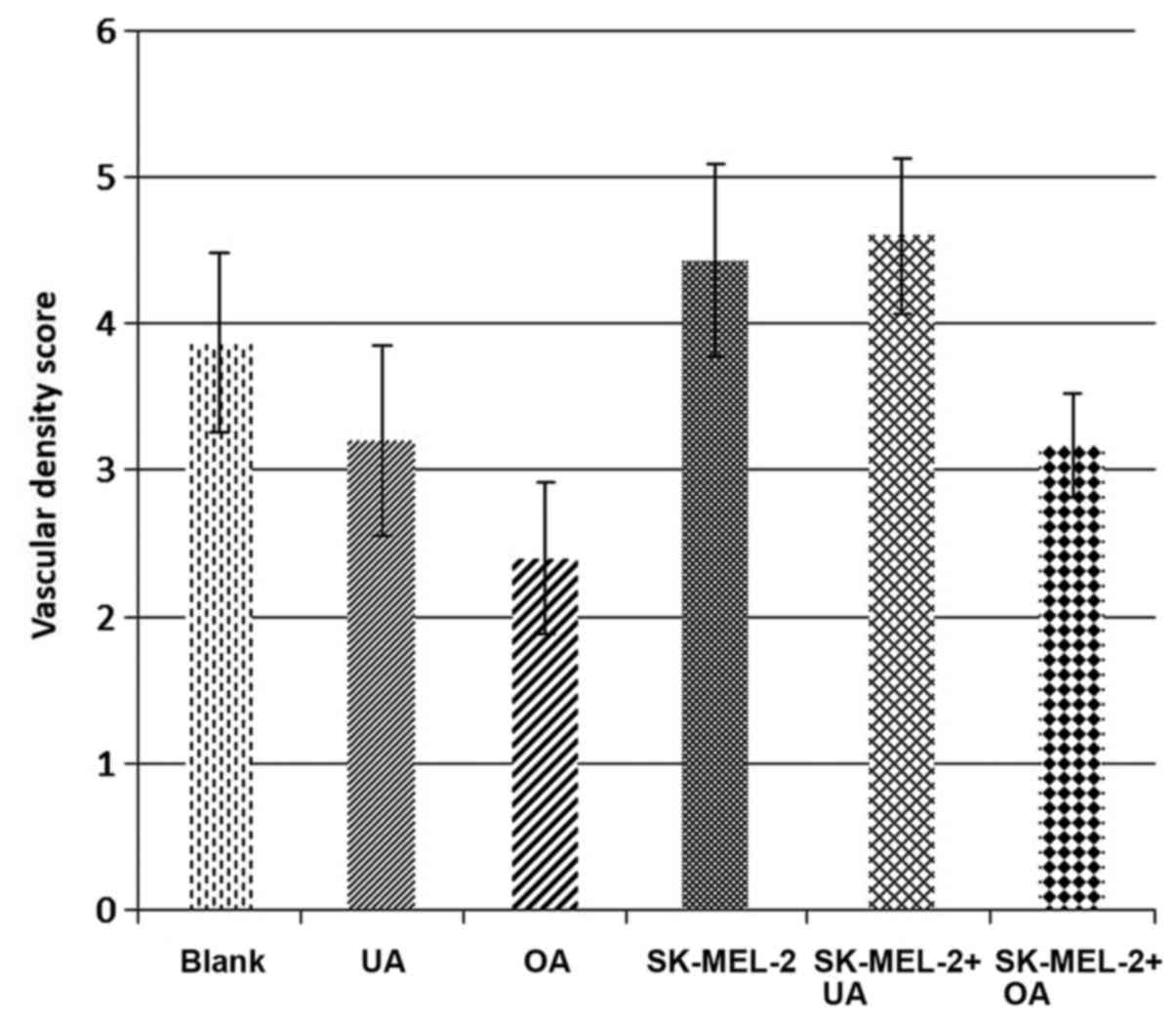
The morphometric evaluation of the angiogenic
process by applying a 0–5 scale in correlation with the degree of
vascular density also indicated that vessel growth was inhibited to
a greater extent by OA during the rapid growing stage of the
embryo, as well during the tumor growth process and related
angiogenesis, using the SK-MEL-2 cell line. Treatment with OA led
to a lower vascular density for the assessment of melanoma-induced
angiogenesis, while UA induced very similar or slightly higher
values than those of the control specimens with only melanoma cells
(Fig. 6).
Discussion
Proliferation and apoptosis constitute a very
precise equilibrium in the healthy human body. Solid tumors
represent a cluster of cells originating from a transformed founder
cell with an increased capacity of proliferation, a decreased rate
of apoptosis, an augmented metabolic rate, a high invasive capacity
and an altered morphology (45,46).
The proliferative ability of cancer cells at a distant site is
essential for metastasis, and difficulties in establishing
secondary growth may explain why <0.01% of circulating tumor
cells (CTCs) actually form metastases (47).
In this study, we evaluated the potential use of two
widely distributed natural triterpenoids as chemopreventive agents
in the management of N-RAS-mutated melanoma, obtaining new data on
the possible effects of UA and OA on the deregulated pathways of
this type of melanoma not related to sun exposure. UA is known to
induce apoptosis through the inhibition of the MEK and
phosphoinositide 3-kinase (PI3K)/mTOR pathways, involved in a
synergistic manner in N-RAS-mutated cancers (8), as demonstrated in prostate cancer
(48) and leukemia cells (49), but not in melanoma cells. Less data
are available on the modulation of these involved tyrosine kinase
receptors by OA. Our results are indicative of the potential
benefits of both compounds possibly in combination, for targeting
the affected signaling keys of the N-RAS type of melanoma.
Our results revealed an intense anti-proliferative
activity of UA in vitro on the human melanoma cell line,
SK-MEL-2, as compared to the effects of OA, whose
anti-proliferative activity was lower. This difference in potency
was also highlighted by other research groups. Meng et al
(48) reported a strong
anti-proliferative potential for UA compared to UA in the case of
Caco2 colon cancer cells with iC50 values of 200
µM for OA and 70 µM for UA. Fulias et al
(50) demonstrated that UA exerted
a significant apoptotic activity against A2058 human melanoma
cells, an effect that was not observed in the case of OA. In a
separate study, our research group reported a superior activity of
UA on several breast cancer tumor cell lines, as opposed to OA to
which cancer cells were resistant (51).
The cytotoxic activity of triterpenoid compounds on
in vitro melanoma assays shown in other studies has
exhibited variability in potency when tested on different cell
lines. Both OA and UA were shown to exert more potent effects on
B164A5 cells as compared to the A431 and A375 melanoma cells
(52). Due to their very low
solubility in water, detrimental to their bioavailability,
triterpenic acids were also investigated in different
pharmaceutical formulations utilized to improve these parameters.
Gamma-cyclodextrincomplexes with UA and OA were also tested for
their biological activities, indicating better results than for the
pure compounds on three melanoma cell lines (52). Some other triterpenoids have been
tested under the form of myristoyl ester derivatives, which
exhibited increased cytotoxicity values as compared to the
compounds alone, on both A431 and A375 melanoma cells (53).
In normal cells, the cell cycle is tightly
regulated. In cancer cells, due to genetic alterations, this
process becomes disrupted, resulting in uncontrolled growth. Thus,
the cell cycle represents the basis of tumor pathology and
compounds proven to be active on cell cycle regulation may be
candidates in anticancer therapy (54,55).
Taking into account reports that show an
anti-proliferative effect of UA on melanoma cells, which may be
caused by alterations in cell cycle phases, we performed cell cycle
analysis of SK-MEL-2 melanoma cells treated with UA, as previously
described (56). In the present
study, OA did not exert a significant decrease in cell
proliferation; therefore, the activity on cell cycle events was
investigated only for UA. Our study confirmed that the
anti-proliferative effects of UA were mediated by alterations in
the cell cycle, with exposure to 50 µM of UA inducing arrest
in the S phase. It has been previously demonstrated that UA induces
cell cycles arrest at the G1 phase in MCF-7 and pC-3 cells
(57,58). Others have reported the involvement
of OA in the G2/M phase of the cell cycle by decreasing Cyclin
Bi/cdc2 activity in HepG2 cells (18).
Although metastasis is widely regarded as an
inefficient process, the majority of cancer patients succumb to the
disease due to metastases rather than from their primary tumors. In
the metastasis cascade, the adhesion of cancer cells to vascular
endothelial cells through adhesion molecules is a crucial step
(47). This intercellular
interaction takes place in the context of a permanent mechanical
shear stress of the bloodstream, where a balance between the
hydrodynamic friction forces and the intercellular liaisons (cancer
cells, endothelium) is established (59). Due to these pressures, only a small
percentage of CTCs survives in order to metastasize (60). This is why the inhibition of
intercellular interaction has become an important therapeutic
target in the attenuation of metastases (61). Thus, tests which measure cell
adhesion to surfaces/substrates are useful in the characterization
of cellular superficial interactions (62). Investigations into tumor
cell-endothelial contact formations have been based on similarities
to the leukocyte-endothelial cell interactions during inflammation
(61).
Since cell adhesion and cell interaction are key
steps in the metastatic process, in this study, we aimed to examine
the influence of UA, the most active compound, on cell adhesive
capacity. Using the flow chamber protocol, we observed that
incubating the SK-MEL-2 cells with high concentrations of UA (50
µM) led to a decreased adhesion capacity to the ICAM
substrate, without however, influencing the adherence capacity of
the cells to VCAM.
VCAM 1 (also known as CD106) is an adhesion molecule
which mediates the adhesion of leukocytes to the endothelium. It is
expressed by endothelial cells following stimulation by cytokines.
It has been reported that colorectal cancer cells adhere to the
vascular endothelium by binding to VCAM-1 (63). In addition, some types of melanoma
cells are known to adhere to the endothelium through VCAM-1
(64). ICAM-1 (also known as CD54)
is a transmembrane glycoprotein, part of the immunoglobulin family,
and is widely expressed by hematopoietic and non-hematopoietic
cells, including endothelial cells, leukocytes, fibroblasts or
cancer cells; however, under cytokine stimulation it may be
expressed on every human cell type (65). It is known that the inhibition of
ICAM-1 expression on melanoma cells may reduce their metastatic
potential (66).
In order to obtain more data regarding the possible
implication of the two selected triterpenoid compounds on the
aggressive metastatic process induced by melanoma cells with an
N-RAS mutation, in conjunction to the in vitro studies, we
chose to apply an in vivo method, CAM assay. The choice of
the protocol was based on multiple advantages, such as low costs,
consumed time and number of sacrificed animals that render this
assay effective for pre-screening model studies (67).
Following stereomicroscopic evaluation, we observed
that the two analyzed compounds functioned differently on the in
vivo developing vascular system and the tumor microenvironment.
OA exerted more potent effects as compared to UA, on the normal
process of angiogenesis, but without influencing the viability of
the embryos. Inside the application ring, OA induced a decrease in
the number of newly formed capillaries during a highly angiogenic
interval (i.e., EDD7-EDD11) (68).
The vascular branching pattern was also influenced outside the
application site, but to a lower extent compared to UA.
The two triterpenoids also differently influenced
the development of in vivo SK-MEL-2 melanoma on the chick
embryo CAM. UA induced a lower impairment of the tumor progress,
without important limitations in tumor growth, which was in
contrast with the in vitro results of the SK-MEL-2
cytotoxicity assay. Moreover, tumor angiogenesis was not inhibited.
Therefore, the invasiveness of the tumor was not restricted,
showing extended areas of secondary tumors outside the application
spot. At a concentration of 30 µM, it seemed that UA had
stimulatory implications on tumor progression, compared with the
untreated control CAMs. Conversely, OA, the compound considered to
exert less potent effects on tumor growth in vitro on the
SK-MEL-2 cells, limited to a greater extent the growth of tumor
cells inside the ring, in vivo.
Other studies have reported the anti-angiogenic
effects of the two compounds. Cárdenas et al (69) reported the in vitro effects
of UA on angiogenesis, proving that it inhibited certain stages of
angiogenesis (proliferation, migration and endothelial cell
differentiation), while stimulating other stages (extracellular
matrix degradation by MMP-2 and urokinases). Still, both compounds
were shown to inhibit the production of vascular endothelial growth
factor (VEGF), one of the factors responsible for angiogenesis.
Lin et al (70) reported the anti-angiogenic effects
of UA in vitro and in vivo in colorectal cancer. It
reduced intratumoral microvascular density in mice with colorectal
cancer, as well as the number of blood vessels in the CAM pattern,
and it also inhibited the expression of certain angiogenic factors.
Both UA and OA exerted anti-angiogenic effects on liver cancer
cells (71).
It has been reported that VEGF-induced angiogenesis
can be modulated by ICAM (72), a
cellular adhesion molecule with high impact in developing
recurrence, invasion and metastatic process (73), its production being inhibited by
the two studied acids (28). As we
only tested UA for this effect, and it showed an anti-adhesive
potential, particularly towards I-CAM molecules in vitro, we
considered that the invasive pattern of the SK-MEL2 cells incubated
with UA on the CAM assay was rather determined by a different
pathway.
As regards the anti-angiogenic effects of 30
µM UA and OA on normal CAM, our results obtained for UA were
contrary to those reported by Cárdenas et al (69), who observed a reduction in
angiogenesis in 50% of the eggs treated with 20 µmol UA. In
addition, Sohn et al (74)
reported an increased anti-angiogenic potential for UA, compared to
OA, with iC50 values of 5 and 20 µM for UA and
OA, respectively. However, both studies were performed using a
bovine aortic endothelial cell CAM model. On the other hand, Kiran
et al (75) reported an
impairment of angiogenic modulators after the administration of UA
at concentrations >10 µM, while lower concentrations did
not affect the angiogenesis of human umbilical vein endothelial
cells (HUVECs). No data were found regarding the effects of OA or
UA in a SK-MEL-2 melanoma model on the CAM assay.
In conclusion, the two tested triterpenoid acids, UA
and OA, exerted differential effects in vitro and in
vivo on SK-MEL-2 melanoma cells. UA exerted a significant
dose-dependent anti-proliferative effect in vitro, compared
to OA. The cytotoxic effects in vitro on the melanoma cells
were determined by the alterations in the cell cycle phases by UA
that induced cell arrest in the S phase. Moreover, UA may
contribute to the capacity of SK-MEL-2 melanoma cell invasiveness
by limiting the cell adhesion capacity to ICAM molecules, but not
influencing the adhesion to VCAM.
Assessing the effects of the two triterpenoids in
vivo using SK-MEL-2 melanoma cells on a CAM model, our results
revealed the potential impairment of the invasive and angiogenic
tumor process to a greater extent for OA compared to UA. These
elements are suggestive of testing the two compounds as a mixture,
with possible synergic or additive effects on blood vessels and
tumor cells. Even though further studies are warranted to confirm
our effects, the easily available compounds from accessible natural
sources, UA and OA, may be considered effective chemopreventive
agents for possible use in the management of N-RAS-mutated
melanoma.
Acknowledgments
This study was supported by an internal grant at
UMFT 'Victor Babes', Grant III-C5-PCFI-2017/2018-04 ROINEXTRAMAM;
Project director, Avram Stefana.
References
|
1
|
American Cancer Society: Key statistics
for melanoma skin cancer. https://www.cancer.org/cancer/melanoma-skin-cancer/about/key-statistics.html.
Accessed March 17, 2017.
|
|
2
|
Schadendorf D, Fisher DE, Garbe C,
Gershenwald JE, Grob JJ, Halpern A, Herlyn M, Marchetti MA,
McArthur G, Ribas A, et al: Melanoma. Nat Rev Dis Primers.
1:150032015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Danciu C, Oprean C, Coricovac DE, Andreea
C, Cimpean A, Radeke H, Soica C and Dehelean C: Behaviour of four
different B16 murine melanoma cell sublines: C57BL/6J skin. Int J
Exp Pathol. 96:73–80. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu S and Singh RK: Resistance to
chemotherapy and molecularly targeted therapies: rationale for
combination therapy in malignant melanoma. Curr Mol Med.
11:553–563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mouawad R, Sebert M, Michels J, Bloch J,
Spano JP and Khayat D: Treatment for metastatic malignant melanoma:
Old drugs and new strategies. Crit Rev Oncol Hematol. 74:27–39.
2010. View Article : Google Scholar
|
|
6
|
Khan MK, Khan N, Almasan A and Macklis R:
Future of radiation therapy for malignant melanoma in an era of
newer, more effective biological agents. Onco Targets Ther.
4:137–148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Finn L, Markovic SN and Joseph RW: Therapy
for metastatic melanoma: The past, present, and future. BMC Med.
10:232012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Posch C, Moslehi H, Feeney L, Green GA,
Ebaee A, Feichtenschlager V, Chong K, Peng L, Dimon MT, Phillips T,
et al: Combined targeting of MEK and PI3K/mTOR effector pathways is
necessary to effectively inhibit NRAS mutant melanoma in vitro and
in vivo. Proc Natl Acad Sci USA. 110:4015–4020. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Johnson DB and Sosman JA: Update on the
targeted therapy of melanoma. Curr Treat Options Oncol. 14:280–292.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Posch C, Vujic I, Monshi B, Sanlorenzo M,
Weihsengruber F, Rappersberger K and Ortiz-Urda S: Searching for
the Chokehold of NRAS Mutant Melanoma. J Invest Dermatol.
136:1330–1336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang Y-L and Liu Z-P: Natural products as
anti-invasive and anti-metastatic agents. Curr Med Chem.
18:808–829. 2011. View Article : Google Scholar
|
|
12
|
Cragg GM and Pezzuto JM: Natural products
as a vital source for the discovery of cancer chemotherapeutic and
chemopreventive agents. Med Princ Pract. 25(Suppl 2): 41–59. 2016.
View Article : Google Scholar
|
|
13
|
Folkman J, Merler E, Abernathy C and
Williams G: Isolation of a tumor factor responsible for
angiogenesis. J Exp Med. 133:275–288. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ejaz S, Anwar K, Taj R and Ashraf M: A
novel link between angiogenesis and natural products:
Anti-angiogenic effects of Opuntia dillenii. Cent Eur J Biol.
9:298–308. 2013.
|
|
15
|
Li WW, Li VW, Hutnik M and Chiou AS: Tumor
angiogenesis as a target for dietary cancer prevention. J Oncol.
2012:8796232012. View Article : Google Scholar
|
|
16
|
Hata K, Hori K and Takahashi S:
Differentiation- and apoptosis-inducing activities by pentacyclic
triterpenes on a mouse melanoma cell line. J Nat Prod. 65:645–648.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Allouche Y, Warleta F, Campos M,
Sánchez-Quesada C, Uceda M, Beltrán G and Gaforio JJ: Antioxidant,
antiproliferative, and pro-apoptotic capacities of pentacyclic
triterpenes found in the skin of olives on MCF-7 human breast
cancer cells and their effects on DNA damage. J Agric Food Chem.
59:121–130. 2011. View Article : Google Scholar
|
|
18
|
Zhang W, Men X and Lei P: Review on
anti-tumor effect of triterpene acid compounds. J Cancer Res Ther.
10(Suppl 1): 14–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chudzik M, Korzonek-Szlacheta I and Król
W: Triterpenes as potentially cytotoxic compounds. Molecules.
20:1610–1625. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mencherini T, Picerno P, Scesa C and
Aquino R: Triterpene, antioxidant, and antimicrobial compounds from
Melissa officinalis. J Nat Prod. 70:1889–1894. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wójciak-Kosior M, Sowa I, Kocjan R and
Nowac R: Effect of different extraction techniques on
quantification of oleanolic and ursolic acid in Lamii albi flos.
Ind Crops Prod. 44:373–377. 2013. View Article : Google Scholar
|
|
22
|
J C Furtado NA, Pirson L, Edelberg H, M
Miranda L, Loira-Pastoriza C, Preat V, Larondelle Y and André CM:
Pentacyclic triterpene bioavailability: An overview of in vitro and
in vivo studies. Molecules. 22:E4002017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Žiberna L, Šamec D, Mocan A, Nabavi SF,
Bishayee A, Farooqi AA, Sureda A and Nabavi SM: Oleanolic acid
alters multiple cell signaling pathways: implication in cancer
prevention and therapy. Int J Mol Sci. 18:E6432017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ikeda Y, Murakami A and Ohigashi H:
Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol Nutr
Food Res. 52:26–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bishayee A, Ahmed S, Brankov N and Perloff
M: Triterpenoids as potential agents for the chemoprevention and
therapy of breast cancer. Front Biosci (Landmark Ed). 16:980–996.
2011. View Article : Google Scholar
|
|
26
|
Meng QX, Roubin RH and Hanrahan JR:
Ethnopharmacological and bioactivity guided investigation of five
TCM anticancer herbs. J Ethnopharmacol. 148:229–238. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Guo WJ and Yang QY: Effects of
ursolic acid and oleanolic acid on human colon carcinoma cell line
HCT15. World J Gastroenterol. 8:493–495. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan SL, Huang CY, Wu ST and Yin MC:
Oleanolic acid and ursolic acid induce apoptosis in four human
liver cancer cell lines. Toxicol In Vitro. 24:842–848. 2010.
View Article : Google Scholar
|
|
29
|
Lee IK, Kim DH, Lee SY, Kim KR, Choi SU,
Hong JK, Lee JH, Park YH and Lee KR: Triterpenoic acids of Prunella
vulgaris var. lilacina and their cytotoxic activities in vitro.
Arch Pharm Res. 31:1578–1583. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
George VC, Naveen Kumar DR, Suresh PK and
Kumar A: Oleanolic acid inhibits cell growth and induces apoptosis
in A375 melanoma cells. Biomed Prev Nutr. 4:95–99. 2014. View Article : Google Scholar
|
|
31
|
Oprean C, Mioc M, Csányi E, Ambrus R,
Bojin F, Tatu C, Cristea M, Ivan A, Danciu C, Dehelean C, et al:
Improvement of ursolic and oleanolic acids' antitumor activity by
complexation with hydrophilic cyclodextrins. Biomed Pharmacother.
83:1095–1104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Avram S, Avram S, Crisan L, Pacureanu L,
Kurunczi L and Bora A: Self-Organizing Map Classif Model. Rev Roum
Chim. 60:167–173. 2015.
|
|
33
|
Avram SI, Pacureanu LM, Bora A, Crisan L,
Avram S and Kurunczi L: ColBioS-FlavrC: A collection of
bioselective flavonoids and related compounds filtered from
high-throughput screening outcomes. J Chem Inf Model. 54:2360–2370.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bora A and Avram S, Ciucanu I, Raica M and
Avram S: Predictive models for fast and effective profiling of
kinase inhibitors. J Chem Inf Model. 56:895–905. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mu DW, Guo HQ, Zhou GB, Li JY and Su B:
Oleanolic acid suppresses the proliferation of human bladder cancer
by Akt/mTOR/S6K and ErK1/2 signaling. Int J Clin Exp Pathol.
8:13864–13870. 2015.
|
|
36
|
Rufino-Palomares EE, Pérez-Jiménez A,
Reyes-Zurita FJ, García-Salguero L, Mokhtari K, Herrera-Merchán A,
Medina PP, Peragón J and Lupiáñez JA: Anti-cancer and
anti-angiogenic properties of various natural pentacyclic
tri-terpenoids and some of their chemical derivatives. Curr Org
Chem. 19:919–947. 2015. View Article : Google Scholar
|
|
37
|
Laszczyk MN: Pentacyclic triterpenes of
the lupane, oleanane and ursane group as tools in cancer therapy.
Planta Med. 75:1549–1560. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shanmugam MK, Nguyen AH, Kumar AP, Tan BKH
and Sethi G: Targeted inhibition of tumor proliferation, survival,
and metastasis by pentacyclic triterpenoids: Potential role in
prevention and therapy of cancer. Cancer Lett. 320:158–170. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shan J, Xuan Y, Zhang Q, Zhu C, Liu Z and
Zhang S: Ursolic acid synergistically enhances the therapeutic
effects of oxaliplatin in colorectal cancer. Protein Cell.
7:571–585. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ribatti D: The chick embryo
chorioallantoic membrane in the study of tumor angiogenesis. Rom J
Morphol Embryol. 49:131–135. 2008.PubMed/NCBI
|
|
41
|
Ribatti D: The Chick Embryo
Chorioallantoic Membrane in the Study of Angiogenesis and
Metastasis. 1st edition. Springer; Netherlands: 2010, View Article : Google Scholar
|
|
42
|
Bacabac RG, Smit TH, Cowin SC, Van Loon
JJ, Nieuwstadt FT, Heethaar R and Klein-Nulend J: Dynamic shear
stress in parallel-plate flow chambers. J Biomech. 38:159–167.
2005. View Article : Google Scholar
|
|
43
|
Zhang Y and Neelamegham S: An analysis
tool to quantify the efficiency of cell tethering and firm-adhesion
in the parallel-plate flow chamber. J immunol Methods. 278:305–317.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao L, Liao FL, Han D and Zhou H:
Application of parallel-plate flow chamber in cancer research. Bull
Acad Mil Med Sci. 33:482–485. 2009.
|
|
45
|
Griffiths AJF, Miller JH, Suzuki DT,
Lewontin RC and Gelbart WM: Cancer: The Genetics Of Aberrant Cell
Control An introduction to Genetic Analysis. 7th edition. W.H.
Freeman & Co; New York, NY: 2000
|
|
46
|
Pfarr K, Danciu C, Arlt O, Neske C,
Dehelean C, Pfeilschifter JM and Radeke HH: Simultaneous and dose
dependent melanoma cytotoxic and immune stimulatory activity of
betulin. PLoS One. 10:e01188022015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wong SY and Hynes RO: Lymphatic or
hematogenous dissemination: How does a metastatic tumor cell
decide? Cell Cycle. 5:812–817. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Meng Y, Lin ZM, Ge N, Zhang DL, Huang J
and Kong F: Ursolic acid induces apoptosis of prostate cancer cells
via the PI3K/Akt/mTOR pathway. Am J Chin Med. 43:1471–1486. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Beagle B and Fruman DA: The PI3K-AKT-mTOR
signaling network in AML. Targeted Therapy of Acute Myeloid
Leukemia. Andreeff M: Springer; New York, NY: pp. 335–362. 2015
|
|
50
|
Fuliaş A, Ledeţi I, Vlase G, Vlase T,
Şoica C, Dehelean C, Oprean C, Bojin F, Şuta LM, Bercean V and
Avram S: Thermal degradation, kinetic analysis, and apoptosis
induction in human melanoma for oleanolic and ursolic acids. J
Therm Anal Calorim. 125:759–768. 2016. View Article : Google Scholar
|
|
51
|
Oprean C, Zambori C, Borcan F, Soica C,
Zupko I, Minorics R, Bojin F, Ambrus R, Muntean D, Danciu C, et al:
Anti-proliferative and antibacterial in vitro evaluation of the
polyurethane nanostructures incorporating pentacyclic triterpenes.
Pharm Biol. 54:2714–2722. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Trandafirescu C, Antal D, Soica C, Zupko
I, Minorics R, Ambrus R, Borcan F, Oprean C, Danciu C, Avram S, et
al: Cyclodextrin complexes of oleanolic and ursolic acid
physico-chemical and biological preliminary evaluation. Rev Chim.
65:1163–1167. 2014.
|
|
53
|
Pinzaru I, Trandafirescu C, Szabadai Z,
Mioc M, Ledeti I, Coricovac D, Ciurlea S, Ghiulai RM, Crainiceanu Z
and Simu G: Synthesis and biological evaluation of some pentacyclic
lupane triterpenoid esters. Rev Chim. 65:848–851. 2014.
|
|
54
|
wiseGEEK: What is cell cycle arrest?
http://www.wisegeek.com/what-is-cell-cycle-arrest.htm.
Accessed Aug 15, 2017.
|
|
55
|
Cyclacel: Cell cycle in cancer. http://www.cyclacel.com/research_science_cell-cycle.shtml.
Accessed Aug 17, 2017.
|
|
56
|
Chakravarti B, Maurya R, Siddiqui JA, Bid
HK, Rajendran SM, Yadav PP and Konwar R: In vitro anti-breast
cancer activity of ethanolic extract of Wrightia tomentosa: Role of
pro-apoptotic effects of oleanolic acid and urosolic acid. J
Ethnopharmacol. 142:72–79. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Es-Saady D, Simon A, Jayat-Vignoles C,
Chulia AJ and Delage C: MCF-7 cell cycle arrested at G1 through
ursolic acid, and increased reduction of tetrazolium salts.
Anticancer Res. 16:481–486. 1996.PubMed/NCBI
|
|
58
|
Park JH, Kwon H-Y, Sohn EJ, Kim KA, Kim B,
Jeong SJ, Song JH, Koo JS and Kim S-H: Inhibition of Wnt/β-catenin
signaling mediates ursolic acid-induced apoptosis in pC-3 prostate
cancer cells. Pharmacol Rep. 65:1366–1374. 2013. View Article : Google Scholar
|
|
59
|
Lawrence MB, Mcintire LV and Eskin SG:
Effect of flow on polymorphonuclear leukocyte/endothelial cell
adhesion. Blood. 70:1284–1290. 1987.PubMed/NCBI
|
|
60
|
Mitchell MJ and King MR: Computational and
experimental models of cancer cell response to fluid shear stress.
Front Oncol. 3:442013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Bendas G and Borsig L: Cancer cell
adhesion and metastasis: selectins, integrins, and the inhibitory
potential of heparins. Int J Cell Biol. 2012:6767312012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Olivier LA and Truskey GA: A numerical
analysis of forces exerted by laminar flow on spreading cells in a
parallel plate flow chamber assay. Biotechnol Bioeng. 42:963–973.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen C, Zhang Q, Liu S, Parajuli KR, Qu Y,
Mei J, Chen Z, Zhang H, Khismatullin DB and You Z: IL-17 and
insulin/iGF1 enhance adhesion of prostate cancer cells to vascular
endothelial cells through CD44-VCAM-1 interaction. Prostate.
75:883–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Eibl RH and Benoit M: Molecular resolution
of cell adhesion forces. IEE Proc, Nanobiotechnol. 151:128–132.
2004. View Article : Google Scholar
|
|
65
|
Paschos KA, Canovas D and Bird NC: The
role of cell adhesion molecules in the progression of colorectal
cancer and the development of liver metastasis. Cell Signal.
21:665–674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang P, Goodrich C, Fu C and Dong C:
Melanoma upregulates ICAM-1 expression on endothelial cells through
engagement of tumor CD44 with endothelial E-selectin and activation
of a PKCα-p38-Sp-1 pathway. FASEB J. 28:4591–4609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Nowak-Sliwinska P, Segura T and
Iruela-Arispe ML: The chicken chorioallantoic membrane model in
biology, medicine and bioengineering. Angiogenesis. 17:779–804.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Deryugina Ei and Quigley JP: Chick embryo
chorioallantoic membrane model systems to study and visualize human
tumor cell metastasis. Histochem Cell Biol. 130:1119–1130. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cárdenas C, Quesada AR and Medina MA:
Effects of ursolic acid on different steps of the angiogenic
process. Biochem Biophys Res Commun. 320:402–408. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lin J, Chen Y, Wei L, Hong Z, Sferra TJ
and Peng J: Ursolic acid inhibits colorectal cancer angiogenesis
through suppression of multiple signaling pathways. Int J Oncol.
43:1666–1674. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lin CC, Huang CY, Mong MC, Chan CY and Yin
MC: Antiangiogenic potential of three triterpenic acids in human
liver cancer cells. J Agric Food Chem. 59:755–762. 2011. View Article : Google Scholar
|
|
72
|
Sun JJ, Zhou XD, Liu YK, Tang ZY, Feng JX,
Zhou G, Xue Q and Chen J: Invasion and metastasis of liver cancer:
Expression of intercellular adhesion molecule 1. J Cancer Res Clin
Oncol. 125:28–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gho YS, Kim PN, Li HC, Elkin M and
Kleinman HK: Stimulation of tumor growth by human soluble
intercellular adhesion molecule-1. Cancer Res. 61:4253–4257.
2001.PubMed/NCBI
|
|
74
|
Sohn KH, Lee HY, Chung HY, Young HS, Yi SY
and Kim KW: Anti-angiogenic activity of triterpene acids. Cancer
Lett. 94:213–218. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Kiran MS, Viji RI, Sameer Kumar VB and
Sudhakaran PR: Modulation of angiogenic factors by ursolic acid.
Biochem Biophys res Commun. 371:556–560. 2008. View Article : Google Scholar : PubMed/NCBI
|