Introduction
Head and neck squamous cell carcinoma (HNSCC) is the
sixth most common cancer worldwide and the eighth most common
cancer in the USA, with an incidence that is ever-increasing
(1). The majority of these cancer
cases (60%) are identified within the oral cavity (oral squamous
cell carcinoma or OSCC), usually in the base of the tongue or the
floor of the mouth (1). In total
~70% of newly diagnosed OSCC cases are late-stage disease (stage
III and IV) with a 5-year overall survival rate reportedly as low
as 34% (1). Therefore, novel
therapeutic strategies to target advanced OSCC are required to
treat this deadly disease. Thus, the aim of the present study was
to synthesize novel polygodial analogs and investigate their
cytotoxic effects against multiple drug-resistant cancer cell lines
in vitro (2). Polygodial, a
known transient receptor potential vanilloid subtype 1 (TRPV1)
agonist, is an unsaturated 1,4-dialdehyde sesquiterpene and the
pungent component of peppers such as the Dorrigo pepper, mountain
pepper and water pepper (3-6).
Polygodial has a wide range of pharmacological activities including
anti-inflammatory, anti-microbial and anticancer effects (6-9).
Previously described was the synthesis of a C12-Wittig
derivative of polygodial, herein termed P3, which demonstrated
antiproliferative potencies that were 4-fold higher than those of
polygodial against a panel of cancer cell lines (A549, SKMEL-28,
MCF-7, U373 and Hs683) (2).
Notably, these antiproliferative effects were via a
TRPV1-independent mechanism of action that has yet to be
identified. In the present study, an additional analog, P27, was
synthesized that has more marked potency compared with P3 in
vitro, and the antitumor efficacy of P3 and P27 against OSCC
xenografts in athymic nude mice was investigated. In order to
identify the antiproliferative mechanism of action, the effects of
P3 and P27 on cell cycle distribution, mitochondrial transmembrane
potential and apoptosis in vitro were investigated. To
identify potential TRPV1-associated adverse effects, the effects of
P3 and P27 on TRPV1 channel activation in vitro and
nocifensive behaviors elicited in a rat model of orofacial pain
were investigated.
Materials and methods
Plant material
Fresh Tasmannia lanceolata leaf material was
provided by Essential Oils of Tasmania, Kingston, Tasmania,
Australia (www.eotasmania.com.au) and dried at 35°C for 18 h
before use. This leaf material originated from trees cultivated as
commercial T. lanceolata crops grown in Tasmania, Australia.
Polygodial was isolated from T. lanceolata following a
procedure described previously (10).
Synthesis
Compound P3 was prepared as described previously
(2). Compound P27 was prepared
using the following procedure (Fig.
1A): To a solution of tetraethyl methylenediphosphonate (36.9
mg, 0.128 mmol) in tetrahydrofuran (THF; 2 ml), 1.6 M
n-butyllithium (80 µl, 0.128 mmol)was added at −78°C, which
was then stirred for 20 min. A solution of polygodial (10 mg,
0.0427 mmol) in THF (1 ml) (10)
was then added dropwise at −78°C. The resultant mixture was allowed
to warm up to 0°C and stirred for 2 h. Following completion, the
reaction was quenched with saturated NH4Cl and then
extracted with ethyl acetate. The organic layer was washed with
water and dried over anhydrous Na2SO4, and
then concentrated to give a crude residue. The crude product was
purified by preparative thin-layer chromatography using a 60% ethyl
acetate/hexanes solvent system and P27 was obtained with a 95%
yield as a mixture of C9-epimers (14.9 mg). The
previously synthesized compound P3 and the novel compound P27 were
each >95% pure on the basis of the NMR spectra.
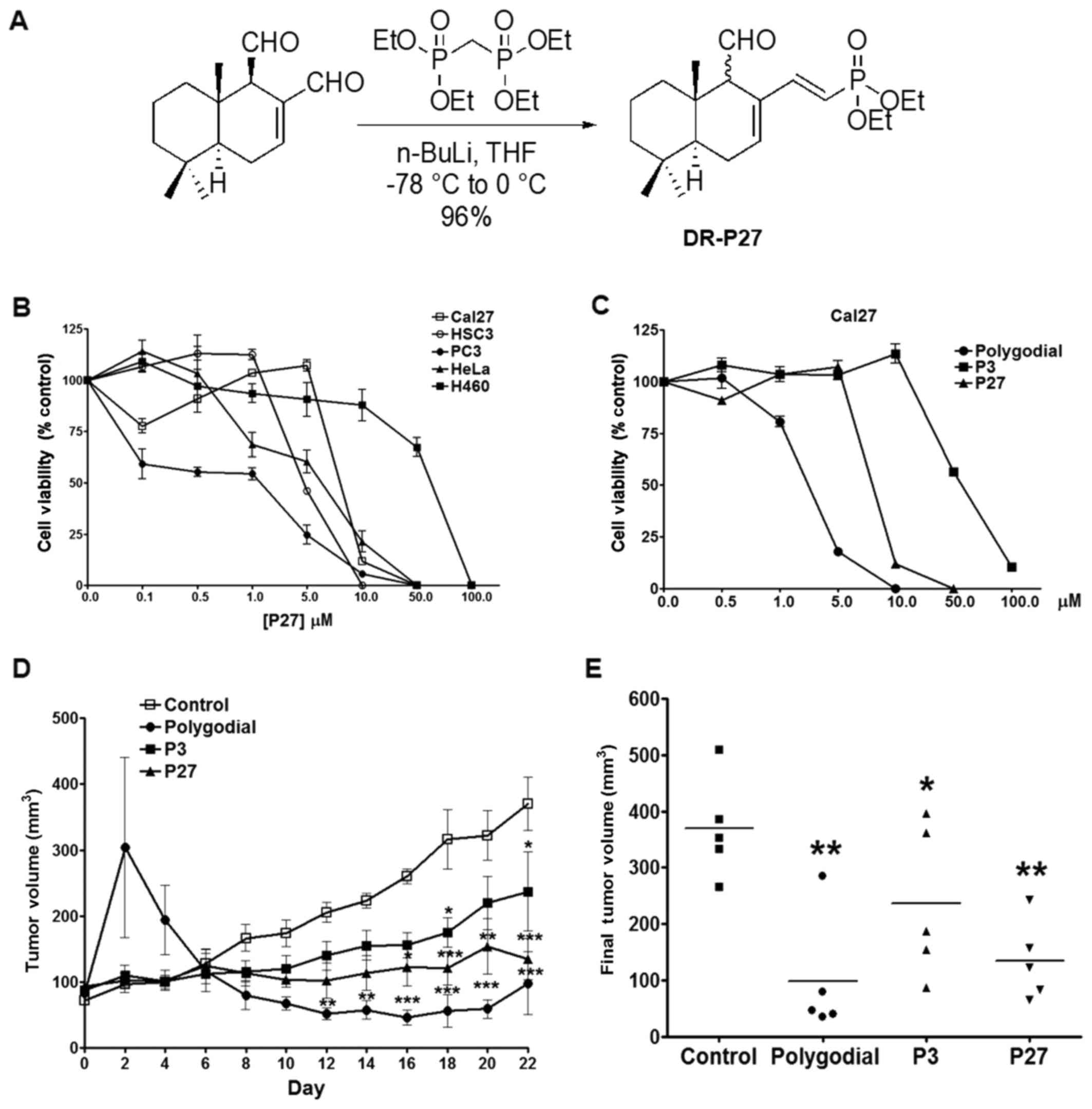 | Figure 1Effects of polygodial analogs on
cancer cell proliferation in vitro and tumor growth in
vivo. (A) Schematic diagram of P27 synthesis. (B) MTS viability
assay in cancer cell lines treated with P27 for 48 h (n=4 per
group). (C) MTS viability assays of Cal27 cells treated with
polygodial, P3 or P27 for 48 h. (D) Tumor volumes (mean ± standard
deviation) of Cal27-derived xenografts treated with 120 µg
polygodial, P3 or P27 every other day for 22 days. Significant
decreases in tumor volumes were observable by day 12 for
polygodial, day 16 for P27 and day 18 for P3 (n=5 per group;
*P<0.05, **P<0.01 and ***P<0.001 vs.
vehicle control). (E) Scatter plot of Cal27 tumor volumes. The mean
volume for vehicle control-treated tumors was 370 mm3,
whereas the mean tumor volumes for polygodial, P3 and P27 treated
tumors were 98, 237 and 135 mm3, respectively (n=5 per
group; *P<0.05 and **P<0.01). Et,
ethyl; n-BuLi, n-butyllithium; THF, tetrahydrofuran. |
Characterization data
1H NMR (400 MHz,
C2HCl3) δ 9.57 (1 H, d, J 5.0 Hz),
9.47 (0.5 H, d, J 4.8 Hz), 7.23-7.00 (1.5 H, m), 6.51-6.41
(1.5 H, m), 5.57 (1 H, t, J 17.4 Hz), 5.26 (0.5 H, t,
J 17.1 Hz), 4.12-3.94 (6 H, m), 2.88-2.71 (1.5 H, m), 2.49
(1 H, dt, J 20.2, 5.0 Hz), 2.31 (0.5 H, dt, J 19.8,
4.8 Hz), 2.27-2.13 (1.0 H, m), 1.89-1.80 (0.5 H, m), 1.78-1.64 (3
H, m), 1.63-1.55 (1.5 H, m), 1.54-1.45 (3 H, m), 1.35-1.25 (9 H,
m), 1.23-1.14 (3 H, m), 1.02-0.87 (13.5 H, m); 13C NMR
(100 MHz, C2HCl3) δ 205.4, 201.7, 150.6,
150.5, 150.0, 149.9, 141.2, 141.2, 141.0, 140.9, 130.9, 130.7,
129.3, 129.0, 113.13, 113.1, 111.3, 111.2, 62.6, 62.6, 62.2, 62.1,
61.5 (m) 60.4, 48.6, 44.6, 42.1, 41.8, 40.2, 37.7, 37.4, 36.6,
33.2, 33.1, 32.7, 25.4, 25.46, 24.7, 24.7, 22.2, 21.9, 21.2, 21.0,
18.4, 18.0, 16.4 (m), 15.5, 14.2; high-resolution mass spectrometry
m/z (electrospray ionization) for
C20H34O4P (M+H+)
calculated, 369.2195; found, 369.2193.
Cell lines
All cell lines, namely Cal27 (OSCC), PC3 (prostate
cancer), HeLa (cervical cancer) and H460 (non-small cell lung
cancer; NSCLC), were obtained from the American Type Culture
Collection (Manassas, VA, USA). HSC3 cells were provided by Dr.
Brian Schmidt (New York University College of Dentistry, New York,
NY, USA). Genetica DNA Laboratories (Cincinnati, OH, USA)
authenticated all cell lines prior to use in the present study.
Cells were maintained in Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum and 1%
penicillin/streptomycin, and maintained at 37°C under 5%
CO2. In order to determine if P3 and P27 interact with
TRPV1 channels, CHO cells overexpressing TRPV1 (CHO-TRPV1) were
used for calcium imaging studies. CHO-TRPV1 cells were kindly
provided by Dr. Ardem Patapoutian at the Scripps Research
Institute, and cultured as previously described at 37°C under 5%
CO2 (11).
MTS cell viability assays
Cell viability was assessed using a Cell Titer
96® Aqueous Non-Radioactive Cell Proliferation assay
(Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocol. For concentration curves, cells were
plated and treated for 48 h with increasing concentrations of
polygodial, P3 or P27 diluted in DMEM at the indicated
concentrations, as described previously (12). Pretreatments were performed using
capsazepine (CPZ; 1 µM) or N-acetylcysteine (NAC; 10 mM) for
30 min, followed by co-treatment with 25 µM polygodial, P3
or P27 for 24 h. Final ethanol concentrations were maintained at
0.1%. Absorbance values of the test groups were compared with those
of the vehicle-treated controls (n=4 per group).
Antitumor evaluations
All mouse studies were approved by the University of
Texas Health Science Center at San Antonio (UTHSCSA) Institutional
Animal Care and Use Committee (IACUC). All animal studies followed
the inter-national guidelines on animal welfare and were in
accordance with the National Institutes of Health (NIH) guide for
the care and use of laboratory animals. In addition, all animal
studies complied with the Animal Research: Reporting of In
Vivo Experiments (ARRIVE) guidelines and the 2013 American
Veterinary Medical Association (AVMA) euthanasia guide-lines. A
total of 20 6-week-old female athymic nude mice, each weighing
approximately 16 g (Envigo Laboratories, Indianapolis, IN, USA),
were used in a laminar airflow cabinet under pathogen-free
conditions. Mice were provided with a 12-h light/12-h dark schedule
at a controlled temperature and humidity, with food and water
available ad libitum. Mice were acclimated for 1 week before
study initiation.
Mice were subcutaneously injected in the right flank
with 3×106 Cal27 cells in 0.1 ml sterile PBS. At 2-4
weeks post-inoculation, tumors grew to an average volume of 100
mm3 and mice were stratified into four experimental
groups (n=5 per group), receiving one of the following treatments
via intratumor injection: Vehicle control (100 µl 0.25%
ethanol diluted in sterile saline), 120 µg polygodial, P3 or
P27 diluted in 100 µl sterile saline (final concentration of
0.25% ethanol). Treatments were repeated every other day for 3
weeks. Mice were monitored daily for tumor growth (using digital
calipers), cachexia and weight loss. Tumor volumes were calculated
using the elliptical formula: 1/2 (length × width2) and
the maximum allowable tumor volume was 1.5 cm3; however
the largest tumor volume in this study was 510 mm3
(13). At the conclusion of the
experiment, tumors were formalin-fixed and processed for
histological analysis. Hematoxylin and eosin (H&E) staining,
terminal deoxynucleotidyl transferase dUTP nick end labeling
(TUNEL) and Ki-67 staining (Ventana Medical Systems, Inc., Tucson,
AZ, USA) were performed to detect apoptotic figures and
proliferating cells respectively; these analyses were conducted by
the UTHSCSA South Texas Reference Laboratory Histopathological Core
Facility using standard methods.
Calcium imaging
Calcium imaging was performed using a FLIPR Calcium
6 Evaluation Kit (Molecular Devices, LLC, Sunnyvale, CA, USA),
according to the manufacturer's protocol. CHO-TRPV1 cells
(9×103) were seeded into a 384-well plate, loaded with
Calcium 6 dye for 2 h and assessed using a PHERAstar FS multimode
plate reader (BMG Labtech, Cary, NC, USA). The effects of 5
µM P3 and P27 (n=3 per group) on calcium influx were
determined according to alterations in fluorescent intensity (535
nm) and compared with that of 100 nM capsaicin (positive
control).
Eye-wipe testing
All procedures for the rat studies were approved by
the UTHSCSA IACUC and followed the NIH Guidelines for the Care and
Use of Laboratory Animals. In addition, all rat studies complied
with the ARRIVE guidelines and the 2013 AVMA euthanasia guidelines.
Six-week-old male Sprague-Dawley rats, weighing approximately 300 g
(Envigo Laboratories), were provided with a 12-h light/12-h dark
schedule at a controlled temperature and humidity, with food and
water available ad libitum. Rats were acclimated for two
weeks prior to study initiation. Rats were placed in a
temperature-controlled (22-25°C) behavioral laboratory in
individual mirrored testing boxes (30×30×30 cm) in which they were
allowed to acclimate for at least 1 h. One drop (40 µl) of a
solution of 0.01% (w/v) capsaicin, P3, or P27 in sterile saline,
was dropped onto one eye of each freely moving animal (n=6 per
group), as described previously (14,15).
The time spent grooming or closing the affected eye was recorded
for a total of 2 min, with the observers blinded to the treatment
allocation groups.
Cell cycle distribution
Cal27 cells were cultured to 50% confluency and
treated with 25 µM polygodial, P3 or P27 for 24 h. Cells
were harvested and fixed in 70% ethanol followed by treatment with
RNase A and staining with propidium iodide. Fluorescence-activated
cell sorting (FACS) analysis of DNA profiles in terms of the
proportion of cells in the G1, S and G2/M
phases of the cell cycle was performed.
JC-1 assay
Cal27 cells were plated at a density of
5×105 in 60-mm dishes and grown overnight. Medium was
replaced the following day with phenol red-free DMEM supplemented
with 20 mM 4-(2-hydroxyethyl)-1-piperazine-ethanesulfonic acid
buffer to maintain a proper pH of 7.4. Cells were then treated for
2 h with polygodial, P3, P27, or vehicle control (n=3 per group) at
the indicated concentrations. As a positive control, cells were
also treated with 15 µM carbonyl cyanide m-chlorophenyl
hydrazide (CCCP). Cells were then stained with 1 µM JC-1 dye
(Thermo Fisher Scientific, Inc.) for 15 min at 37°C, washed, and
assessed via FACS analysis.
Western blot analysis
Cal27 cells were treated for 48 h with the vehicle
control, or with 10, 25 and 50 µM polygodial, P3 or P27
(final ethanol concentration of 0.1%), respectively, and then
harvested and lysed in Laemmli lysis buffer. The cell lysate
concentration was determined at 570 nm reading using a
bicinchoninic acid protein assay (Pierce; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Cell lysates
containing equal concentrations of protein (40 µg) were
separated by SDS-PAGE (10% gel), before being transferred onto a
PVDF membrane and blocked in 5% milk solution in PBS with 0.05%
Tween-20. The membrane was incubated overnight at 4°C with
anti-cleaved poly(ADP-ribose) polymerase (c-PARP) rabbit polyclonal
antibody (cat. no. 5625S; dilution, 1:1,250), anti-α/β-tubulin
rabbit monoclonal antibody (cat. no. CS2148; dilution, 1:1,000) and
anti-GAPDH mouse monoclonal antibody (cat. no. 97166S; dilution,
1:1,000; all from Cell Signaling Technology, Inc., Danvers, MA,
USA) in a total of 6 ml diluent (1% milk in PBS containing 0.1%
Tween-20), as previously described (12). The membrane was washed five times
with PBS containing Tween-20 and incubated with Enhanced
Chemiluminescence Plus detection solution (GE Healthcare, Chicago,
IL, USA) for 1 min. Signals were detected by exposure to
radiographic film for 30 sec.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism (version 4; GraphPad Software, Inc., La Jolla, CA, USA).
Results from cell viability, calcium influx, nocifensive behavior
and mitochondrial transmembrane potential assays, in addition to
Ki-67 staining and TUNEL assays (n=3 per group; 3 fields per
section), were analyzed using one-way analysis of variance (ANOVA)
and Bonferroni's post-hoc test. Statistical analyses of tumor
growth were conducted using two-way ANOVA and repeated measures
with Bonferroni's post-hoc test. Cell cycle distribution was
analyzed using Student's t-test. All data are expressed as the mean
± standard deviation, except for the cell cycle distribution
results, which are expressed as the mean ± standard error of the
mean. P<0.05 was considered to indicate a statistically
significant difference.
Results
P27, a novel polygodial analog exhibits
potent antiproliferative effects in vitro
P27 was synthesized and purified to 95%, as outlined
in Fig. 1A, prior to analysis for
its effects against multiple cancer types using cell viability
assays: Cal27 (OSCC), HSC3 (OSCC), PC3 (prostate cancer), HeLa
(cervical cancer) and H460 (NSCLC). It was identified previously
that the antiproliferative effects of polygodial and P3 are not
cancer-type-specific (2).
Likewise, P27 was efficacious against multiple cancer types
resulting in a concentration-dependent decrease in cell viability
following 48 h of treatment, with the concentration of drug causing
50% decrease in proliferation of cancer cells (GI50)
ranging between 2 and 8 µM in all cell lines except for H460
cells, which had a GI50 of 70 µM (Fig. 1B).
P27 and P3 exhibit significant
antiproliferative effects in Cal27 OSCC cells with no observable
adverse side effects in tumor-bearing mice
To determine whether the antiproliferative effects
observed in vitro translated in vivo, polygodial, P3
and P27 were investigated in Cal27 cells in culture and in
Cal27-derived tumors xenografted in athymic nude mice. A marked
decrease in Cal27 cell viability was noted following 48 h of
treatment with polygodial, P3 and P27, with GI50 values
of 4, 75 and 10 µM, respectively (Fig. 1C). Similarly, Cal27-derived tumors
exhibited significant decreases in growth when treated with
polygodial, P3 and P27 every other day (Fig. 1D). Notably, polygodial treatment
initially induced a significant increase in tumor volume on day 2
that subsided by day 6. This increase was due to an inflammatory
response at the injection site; however, no changes in activity or
body weight were observed. One mouse exhibited some tissue damage
(hemorrhage) following initial treatment with polygodial, which was
resolved by day 6. By day 12, polygodial treatment had induced a
significant decrease in tumor growth (P<0.01), compared with the
vehicle control, an effect that persisted throughout the 22-day
study period; final average tumor volumes were 98 mm3
and 370 mm3, respectively (Fig. 1E).
P3 and P27 treatments each led to significant
decreases in tumor growth, with final average tumor volumes of 237
mm3 and 135 mm3, respectively (Fig. 1D and E). Similar to the in
vitro results, P27 was more potent compared with P3 and
exhibited significant decreases in tumor volumes by day 18
(P<0.001). There was no statistically significant difference
between P27 and polygodial antitumor effects; the treatments
yielded significantly decreased tumor volumes compared with the
vehicle control, and were equally efficacious. However, P27 did not
elicit the inflammatory response that was consistently observed
with polygodial, nor did it cause any observable tissue damage.
Although P3 was comparably the least effective, it did
significantly decrease tumor growth by day 22 when compared with
the vehicle-treated controls (P<0.05); P3 treatment also had no
detectable adverse effects.
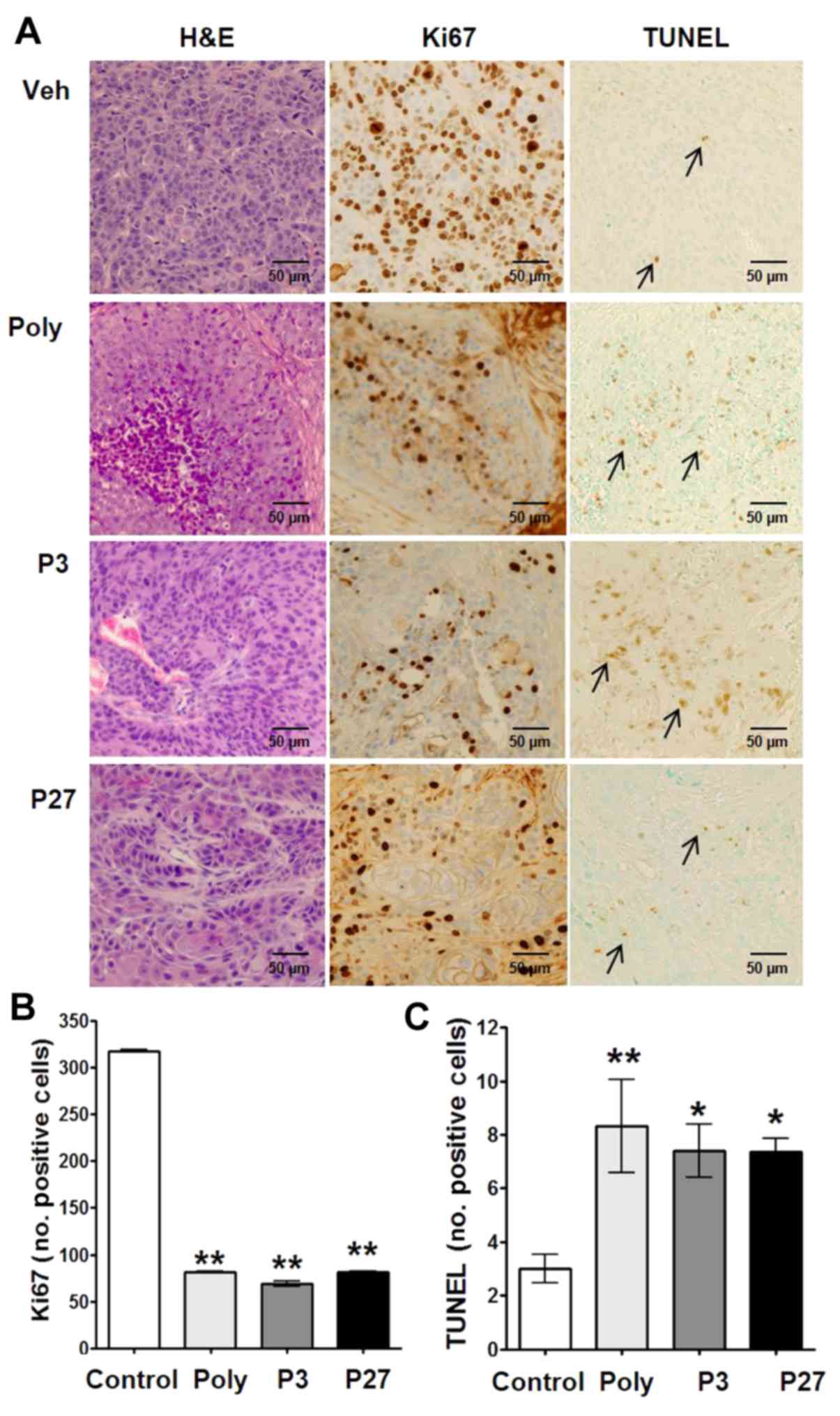
Polygodial, P3, and P27 decrease the
number of proliferating tumor cells and induce apoptosis in
vivo
Histopathological analysis of H&E-stained
Cal27-derived tumors from mice treated with polygodial, P3 or P27
revealed large necrotic cores with viable cells along the tumor
margins (Fig. 2A). Ki-67 staining
and TUNEL assays were performed and quantified within the growing
tumor front to assess changes in cell proliferation and apoptosis,
respectively (Fig. 2A). All
compounds significantly decreased the number of proliferating cells
(P<0.01; Fig. 2B). In addition,
a significant increase in the number of apoptotic cells, compared
with the vehicle control, was observed with polygodial, P3 and P27
(Fig. 2C). Whereas polygodial
yielded the highest number of apoptotic cells, there was no
significant difference between polygodial, P3 and P27.
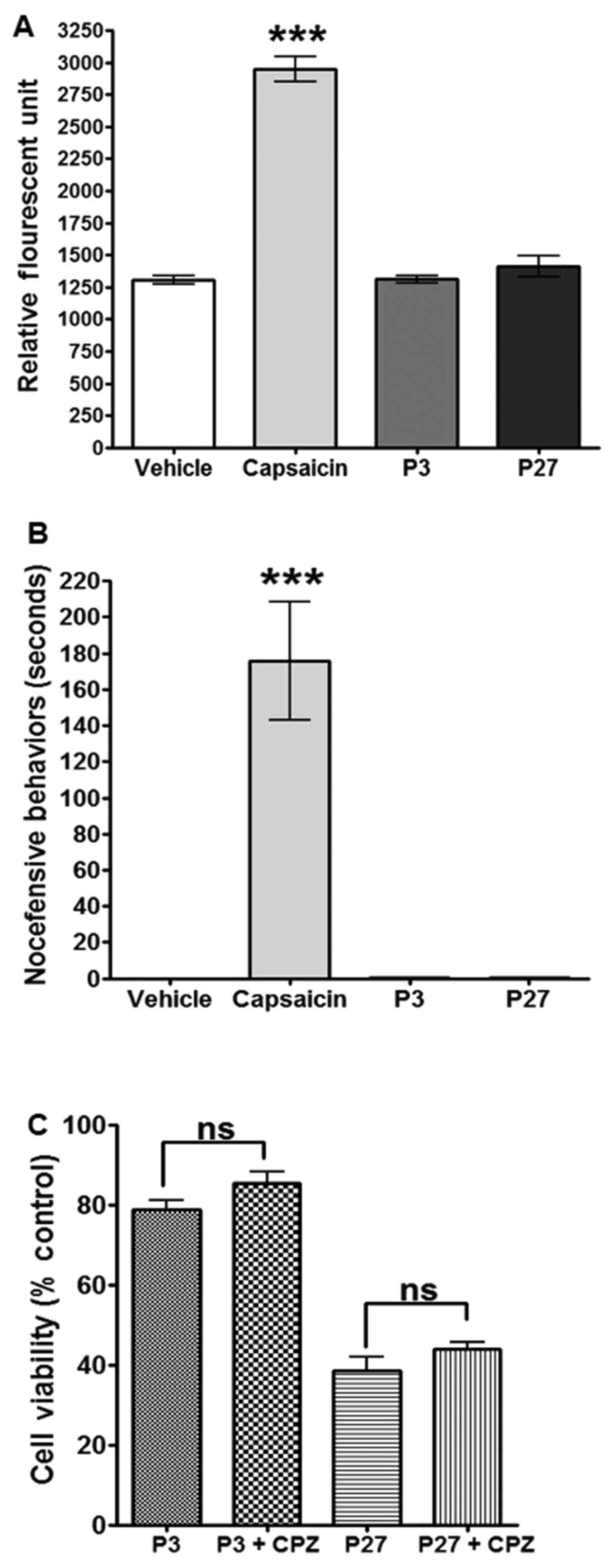
P3 and P27 fail to activate TRPV1 in
vitro and to induce nocifensive behavior in a rat model of
orofacial pain
Our previous study indicated that P3 does not
interact with TRPV1, whereas polygodial is a known TRPV1 agonist
(2). Therefore, the potential
TRPV1 interactions with P27 were compared with those with P3. Using
CHO cells that overexpress TRPV1, calcium imaging analysis was
performed, which confirmed that P3 does not induce calcium influx
(Fig. 3A). Similarly, P27 failed
to induce calcium influx (Fig.
3A). We hypothesized that these compounds may be antagonizing
TRPV1, and, thus, Pretreated CHO-TRPV1 cells with either P3 or P27,
followed by the standard TRPV1 agonist capsaicin; however, no TRPV1
antagonism was identified using P3 or P27 (data not shown). These
results were recapitulated in vivo using a rat model of
orofacial pain. Rats were treated with the vehicle control, P3, P27
or capsaicin, and nocifensive behavior was determined for 5 min.
Capsaicin induced nocifensive eye-wiping and blinking for 160±40
sec. As with the vehicle control, P3 and P27 failed to induce
nocifensive behavior (Fig.
3B).
Finally, to confirm that TRPV1 activation does not
regulate the antiproliferative effects of P3 and P27, Cal27 cells
were Pretreated with CPZ (1 µM), followed by treatment with
P3 or P27, and the effects on cell proliferation were analyzed. CPZ
failed to reverse the antiproliferative effects of all treatments,
and no additive effects were noted (Fig. 3C).
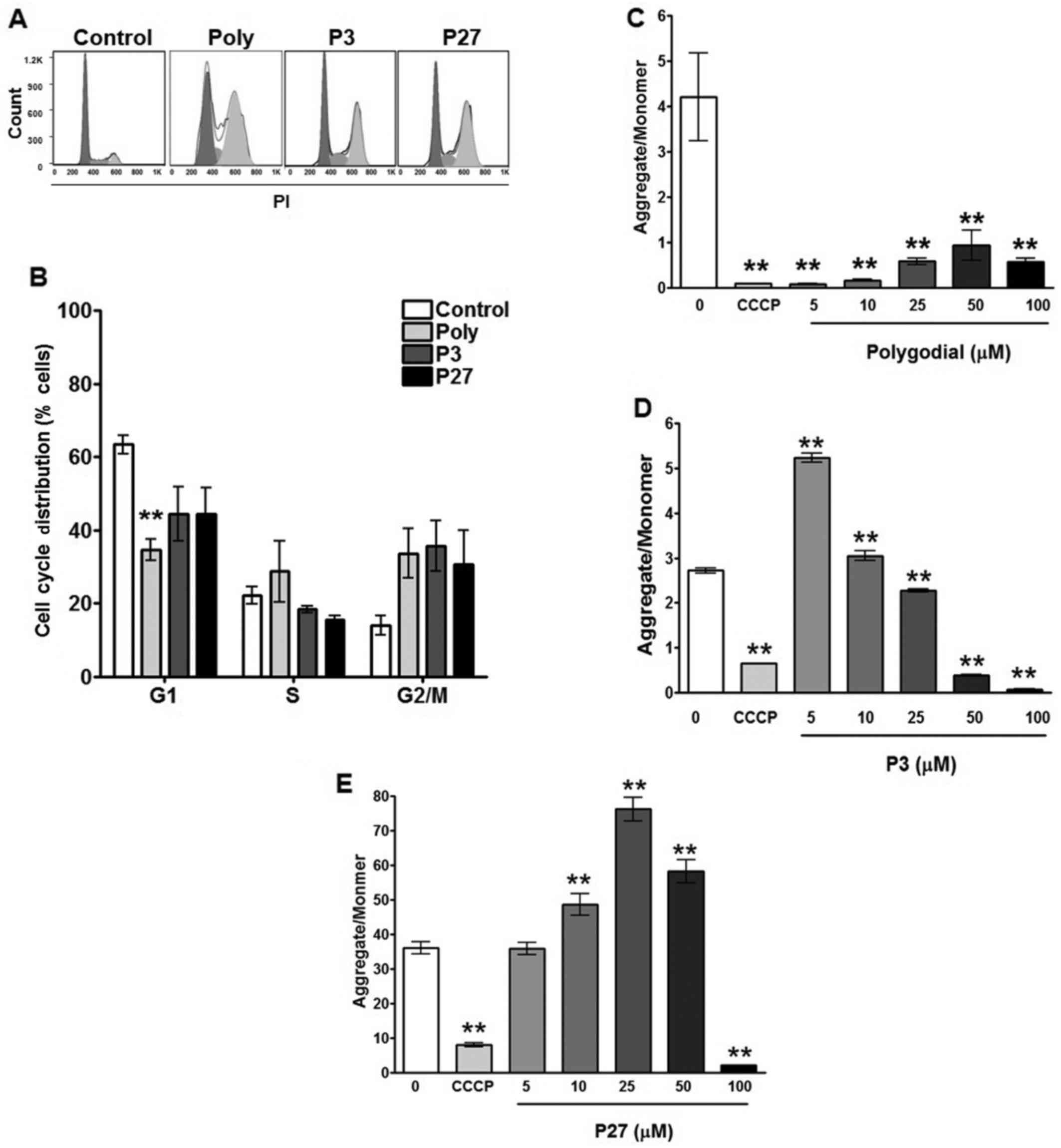
Polygodial, P3 and P27 induce a
G2/M phase block and mitochondrial dysfunction in OSCC
cells
In order to assess the TRPV1-independent
mechanism(s)-of-action, cell cycle distribution and mitochondrial
function assays were performed. Cell cycle distribution analysis in
Cal27 cells revealed that polygodial, P3 and P27 cause a
G2/M phase block (Fig.
4A). Quantification of the cell cycle distribution revealed a
significant decrease in the number of polygodial-treated cells in
the G1 phase, with corresponding increases in the S and
G2/M phases (Fig. 4B).
P3 and P27 induced a decrease in the percentage of cells in the
G1 phase and an increase in the percentage of cells in
the G2/M phase, compared with the control; however,
these differences were not statistically significant. Mitochondrial
transmembrane depolarization was measured in response to increasing
concentrations of polygodial, P3 and P27, using JC-1 dye. The
aggregate to monomer ratio, representative of mitochondrial
transmembrane potential, was immediately diminished with even the
lowest concentration of polygodial (5 µM), indicating a
significant decrease in transmembrane potential (P<0.01;
Fig. 4C). Similarly, P3 and P27
treatments each elicited significant decreases in mitochondrial
transmembrane potential, but at higher concentrations compared with
polygodial (25 and 100 µM, respectively; Fig. 4D and E).
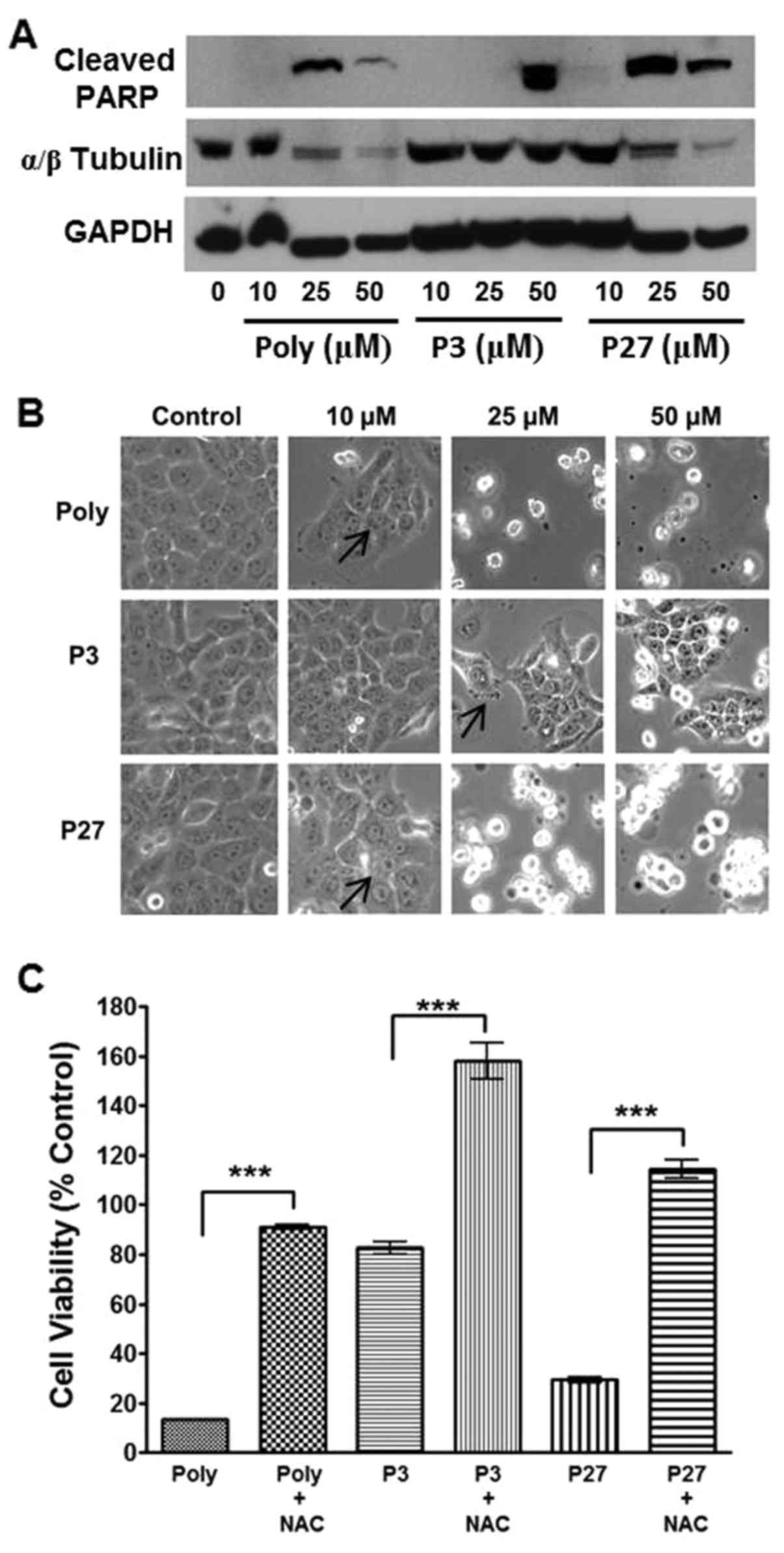
In order to determine whether the decrease in
membrane potential paralleled the induction of apoptosis, western
blot analysis of c-PARP in Cal27 cells was performed. Indeed, a
concentration-dependent induction of c-PARP was present with all
treatments (Fig. 5A). Similarly,
photomicrographs of cells treated with polygodial, P3 and P27
identified membrane blebbing, apoptotic figures and apoptosis with
increasing concentrations that corresponded to the induction of
c-PARP (Fig. 5B; arrows). For
example, polygodial and P27 induced c-PARP at concentrations of 25
µM, whereas P3 required a higher concentration (50
µM) to induce this effect within the same timeframe
(Fig. 5A). In addition, increased
c-PARP levels were correlated with increased numbers of apoptotic
cells as presented in the photomicrographs at 25 µM for
polygodial and P27, and at 50 µM for P3.
Cell viability assays confirmed that the antioxidant
NAC reversed the anti-proliferative effects of polygodial, P3 and
P27 (P<0.001; Fig. 5C),
implicating reactive oxygen species (ROS) as potential mediators of
mitochondrial dysfunction and cell apoptosis. Additionally, a
concentration-dependent decrease in tubulin (initially used as a
loading control) was observed with polygodial and P27 treatments
(Fig. 5A). Therefore, equal
protein loading concentrations were established with GAPDH, which
did not exhibit alterations in expression following these
treatments, thus confirming the changes in tubulin levels in
response to polygodial and P27 treatments.
Discussion
Numerous sesquiterpenes, including polygodial, are
reported to have medicinal and antifeedant properties. Polygodial
is an unsaturated sesquiterpene dialdehyde that is non-mutagenic
with significant antimicrobial and cytotoxic activities against
multiple cancer types (9,16). Structure activity association
studies of unsaturated sesquiterpene dialdehydes reveal that
specific structural features of the dialdehyde functional group are
essential for cytotoxicity (2,16-18).
For example, it was identified that the removal of the aldehyde
groups, through either their reduction to the diol or incorporation
into the pyridazine ring, led to the loss of activity (2,18).
However, conversion of the α,β-unsaturated aldehyde into an
α,β-γ,δ-unsaturated ester resulted in a more potent analog, P3
(2). The present study involved
the synthesis of the polygodial analog P27, which contains an
α,β-γ,δ-unsaturated phosphonate group in place of the aldehyde
(Fig. 1A) and maintains superior
antiproliferative effects while avoiding the undesired TRPV1
agonist property associated with polygodial. On the basis of our
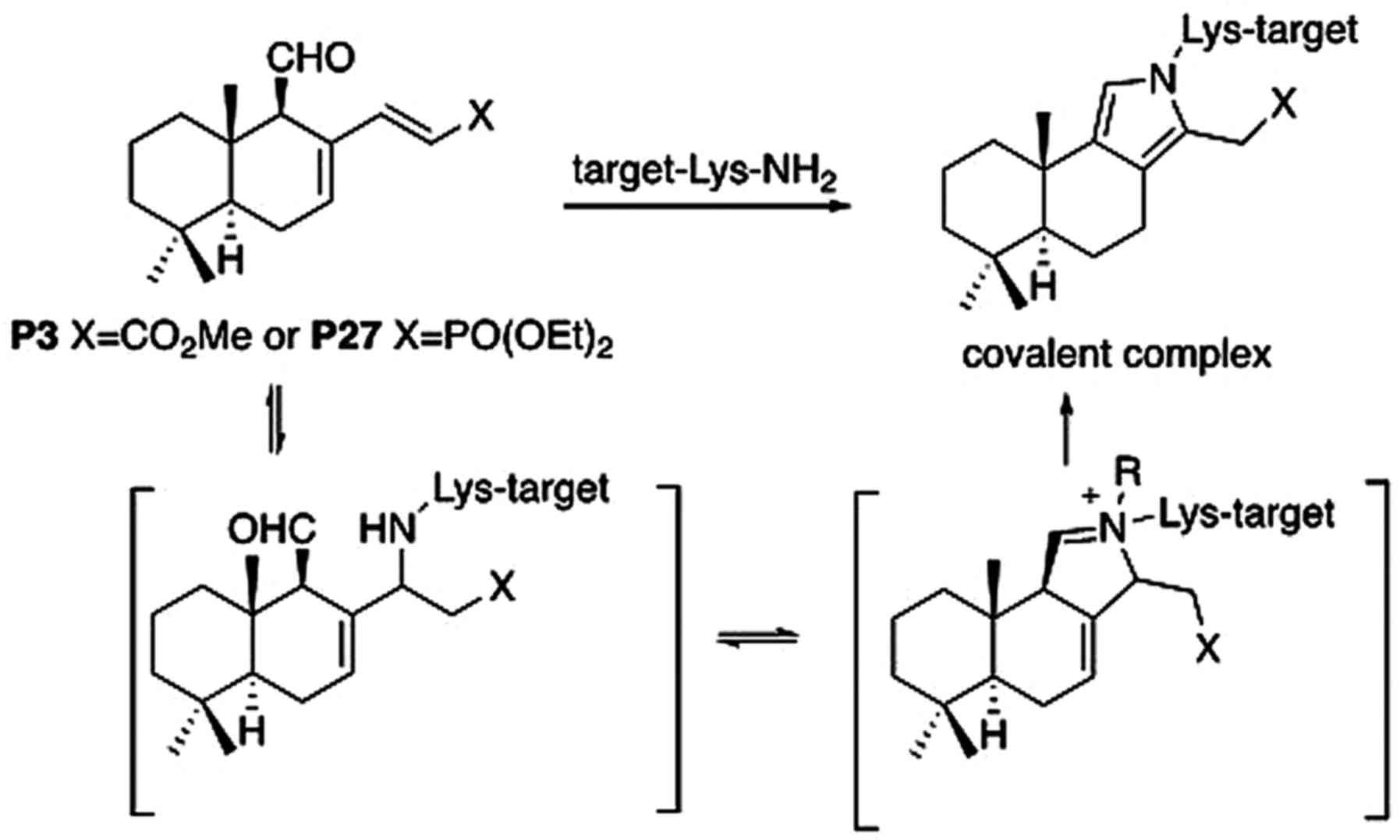
chemical studies (2), we
previously proposed that P3 exerts its biological activities
through the formation of a covalent complex with a lysine group of
its intracellular target (Fig. 6).
Since similar chemistry should apply to P27 (Fig. 6), it could also be hypothesized
that the reaction of P27 with a lysine residue to irreversibly form
a stable pyrrole ring is responsible for its anticancer effects.
This hypothesis requires further study.
As with polygodial, P27 cytotoxicity is not specific
to any cancer type tested; rather, P27 is highly efficacious
against multiple cancer types, indicating that it may have a
universal anticancer mechanism of action. However, the present
mechanistic study focused specifically on oral cancer. P27
significantly decreased the viability of two OSCC cell lines, Cal27
and HSC3. Comparative analyses performed in Cal27 cells
demonstrated that P27 was more potent compared with P3, and less
potent compared with polygodial, in vitro. However, P27 was
equipotent with polygodial against OSCC tumor growth in athymic
nude mice revealing significant decreases in tumor volumes and
proliferating cells, and significant increases in the number of
apoptotic cells, compared with vehicle control-treated tumors. P3
was also identified to be efficacious against OSCC xenografts;
however, similar to the cell based assays, P3 remained the least
potent of the three compounds.
Polygodial consistently induced a marked yet
transitory inflammatory response at the injection site in all mice,
which was evident by day 2 and had subsided by day 6. No
respiratory distress was observed, and no changes in activity
levels or body weight were noted with any of the treatments.
However, one mouse did develop a hemorrhage at the injection site
following administration of the first polygodial treatment, which
healed and did not recur. Presumably, the inflammatory response and
soft tissue damage was due to TRPV1 activation in adjacent
non-cancerous tissues. P3 and P27 did not induce any observable
adverse side effects in adjacent non-malignant tissues, respiration
or neuromuscular function. Furthermore, behavioral studies using a
rat orofacial pain model revealed that P3 and P27 do not elicit
nocifensive behaviors, such as eye-wiping, blinking or closing of
the eye, that are elicited with the TRPV1 agonist capsaicin.
Polygodial was not analyzed in this assay because it is a known
TRPV1 agonist and not a standard positive control for these
behavioral studies. The aim is to develop novel therapeutic
compounds based on the polygodial pharmacophore that do not elicit
polygodial-associated adverse side effects. Indeed, calcium imaging
confirmed that P3 and P27 do not activate or antagonize TRPV1
channels. However, future studies evaluating efficacy and toxicity
via systemic administration are required to elucidate other
potential adverse side effects.
In order to assess the anticancer
mechanism(s)-of-action, cell cycle distribution analysis was
performed. Notably, polygodial, P3 and P27 all caused a
G2/M phase arrest. Although not the focus of the present
study, it was also observed that polygodial and P27 significantly
decreased tubulin protein levels, whereas P3 had no effect. The
antifungal activity of polygodial is attributed, in part, to its
binding to thiol groups found on fungal microtubules (19). Thus, future studies evaluating P27
effects on microtubule polymerization and cell cycle regulators in
cancer are required in order to delineate this potential anticancer
activity.
Polygodial also inhibits fungal mitochondrial ATP
synthesis (7). Castelli et
al (20) demonstrated that
polygodial inhibits electron transport and ATPase activity in rat
liver cells and bovine cardiac mitochondria in vitro.
Therefore, the effects of polygodial, P3 and P27 on OSCC cell
mitochondrial membrane potential were investigated as a measure of
mitochondrial function. Mitochondrial transmembrane polarization is
essential to drive ATP synthesis. Cancer cells are generally
hyperpolarized, compared with non-cancerous cells, due to their
high rate of respiration. Furthermore, high cellular respiration
rates produce increased levels of ROS due to inefficiencies in the
electron transport chain. We hypothesize that these compounds
disrupt mitochondrial function, leading to high levels of ROS that
surpass homeostatic thresholds and induce apoptosis. Indeed, the
antioxidant NAC completely reversed the antiproliferative effects
of polygodial, P3 and P27, implicating the induction of ROS as
drivers of apoptosis.
Depolarization of the mitochondrial transmembrane
potential is one of the key types of mitochondrial dysfunction that
leads to apoptosis (21). It was
identified that polygodial, P3 and P27 all induce depolarization of
the mitochondrial transmembrane potential and cell apoptosis;
however, the mechanisms may differ between polygodial and the
analogs. Initially, P3 and P27 induced an increase in transmembrane
potential followed by a concentration-dependent decrease in
polarization. Calcium homeostasis is a regulator of cell death with
the endoplasmic reticulum (ER) being the primary store of calcium
in the cell, and the mitochondria the main effectors of calcium
mobilization from the ER. These initial apparent increases in
transmembrane potential are consistent with ER stress and the
subsequent release of high calcium levels into the cytosol, thus
interfering with the accuracy of dyes that provide indirect
measures of mitochondrial pH (22). Furthermore, ROS are drivers of ER
stress-associated calcium release, resulting in increased
mitochondrial calcium levels and the production of additional ROS,
thus creating a positive feed-back loop that assures apoptosis
(23-25). Therefore, P3-and P27-induced ROS
may potentiate the apoptotic pathway via ER stress and subsequent
mitochondrial transmembrane depolarization. Ultimately, all
compounds tested induced mitochondrial transmembrane depolarization
that corresponded with increased PARP cleavage and cell apoptosis;
however, polygodial elicited the most notable depolarization of the
mitochondrial membrane at even the lowest concentration tested (5
µM). Polygodial has been identified to inhibit mitochondrial
ATP synthase and ATPase; thus the marked depolarization of
mitochondrial transmembrane potential may be due to polygodial's
direct inhibition of ATP synthesis in addition to its promoting ROS
production (7,20). Conversely, higher concentrations of
P3 and P27 were required to induce the same effect on mitochondrial
transmembrane potential. Given that P27 is equipotent to polygodial
in vivo, it may have a second mechanism of action that is
responsible for its antitumor effects, which may include prompting
ER stress. Studies are underway to evaluate the interactions OF P27
with the ER, electron transport complexes and microtubules in order
to fully ascertain its anticancer mechanism(s)-of-action.
Taken together, we hypothesize that the shared
anticancer effects of polygodial, P3 and P27 are due to the
disruption of mitochondrial function, the generation of ROS and
cell apoptosis. However, the novel compound P27 may have additional
mechanism(s) that enhance its antitumor affects compared with P3;
and with no observable adverse side effects that are present with
polygodial. Additional studies evaluating higher doses and
different routes of administration are underway in order to better
target metastatic OSCC and to fully determine P27 efficacy as a
single-agent therapy. Furthermore, P27 may prove to be useful as an
adjuvant therapy to standard treatments, including chemotherapy and
radiation therapy.
Abbreviations:
|
OSCC
|
oral squamous cell carcinoma
|
|
TRPV1
|
transient receptor potential channel
vanilloid subtype 1
|
Acknowledgments
The authors would like to thank Karla M. Gorena for
her assistance with the FACS analysis at the CTRC Flow Cytometry
Core Facility.
Funding
The present study was supported by the University of
Texas Health Science Center of San Antonio (UTHSCSA), Cancer
Therapy and Research Center (CTRC). FACS analysis was performed at
the CTRC Flow Cytometry Core Facility supported by a National
Cancer Institute (NCI) Cancer Center Support Grant (grant no. P30
CA054174). Medicinal chemistry was supported by the NCI (grant no.
CA186046-01A1). The study performed in the Center for Innovative
Drug Discovery High Throughput Screening Facility was supported by
the National Center for Advancing Translational Sciences, National
Institutes of Health (grant no. UL1 TR001120).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors listed made substantial contributions to
conception and design, acquisition of data, or analysis and
interpretation of data. All authors were also involved in drafting
the manuscript and have given final approval of the version to be
published.
Ethics approval and consent to
participate
All mouse and rat studies were approved by the
University of Texas Health Science Center at San Antonio (UTHSCSA)
Institutional Animal Care and Use Committee (IACUC). All animal
studies followed the international guidelines on animal welfare and
were in accordance with the National Institutes of Health (NIH)
guide for the care and use of laboratory animals. In addition, all
animal studies complied with the Animal Research: Reporting of
In Vivo Experiments (ARRIVE) guide-lines and the 2013
American Veterinary Medical Association (AVMA) euthanasia
guidelines. All procedures for the rat studies were approved by the
UTHSCSA IACUC and followed the NIH Guidelines for the Care and Use
of Laboratory Animals. In addition, all rat studies complied with
the ARRIVE guidelines and the 2013 AVMA euthanasia guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cancer Facts Figures: American Cancer
Society. 2016, https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2016.html,
Accessed on 11-13-17.
|
|
2
|
Dasari R, De Carvalho A, Medellin DC,
Middleton KN, Hague F, Volmar MN, Frolova LV, Rossato MF, De La
Chapa JJ, Dybdal-Hargreaves NF, et al: Wittig derivatization of
sesquiterpenoid polygodial leads to cytostatic agents with activity
against drug resistant cancer cells and capable of pyrrolylation of
primary amines. Eur J Med Chem. 103:226–237. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
da Cunha FM, Fröde TS, Mendes GL,
Malheiros A, Cechinel Filho V, Yunes RA and Calixto JB: Additional
evidence for the anti-inflammatory and anti-allergic properties of
the sesquiterpene polygodial. Life Sci. 70:159–169. 2001.
View Article : Google Scholar
|
|
4
|
McCallion RF, Cole AL, Walker JR, Blunt JW
and Munro MH: Antibiotic substances from New Zealand plants. II.
Polygodial, an anti-Candida agent from Pseudowintera colorata.
Planta Med. 44:134–138. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barnes CS and Loder JW: Structure of
polygodial-a new sesquiterpene dialdehyde from polygonum hydropiper
L. Aust J Chem. 15:322–327. 1962. View Article : Google Scholar
|
|
6
|
André E, Campi B, Trevisani M, Ferreira J,
Malheiros A, Yunes RA, Calixto JB and Geppetti P: Pharmacological
charac-terisation of the plant sesquiterpenes polygodial and
drimanial as vanilloid receptor agonists. Biochem Pharmacol.
71:1248–1254. 2006. View Article : Google Scholar
|
|
7
|
Kubo I, Fujita K and Lee SH: Antifungal
mechanism of polygodial. J Agric Food Chem. 49:1607–1611. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leonard CM and Viljoen AM: Warburgia: A
comprehensive review of the botany, traditional uses and
phytochemistry. J Ethnopharmacol. 165:260–285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Montenegro I, Tomasoni G, Bosio C,
Quiñones N, Madrid A, Carrasco H, Olea A, Martinez R, Cuellar M and
Villena J: Study on the cytotoxic activity of drimane
sesquiterpenes and nordrimane compounds against cancer cell lines.
Molecules. 19:18993–19006. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Just J, Jordan TB, Paull B, Bissember AC
and Smith JA: Practical isolation of polygodial from Tasmannia
lanceolata: A viable scaffold for synthesis. Org Biomol Chem.
13:11200–11207. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rooney L, Vidal A, D'Souza AM, Devereux N,
Masick B, Boissel V, West R, Head V, Stringer R, Lao J, et al:
Discovery, optimization, and biological evaluation of
5-(2-(trifluoromethyl) phenyl)indazoles as a novel class of
transient receptor potential A1 (TRPA1) antagonists. J Med Chem.
57:5129–5140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gonzales CB, De La Chapa JJ, Saikumar P,
Singha PK, Dybdal-Hargreaves NF, Chavez J, Horning AM, Parra J and
Kirma NB: Co-targeting ALK and EGFR parallel signaling in oral
squamous cell carcinoma. Oral Oncol. 59:12–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jensen MM, Jørgensen JT, Binderup T and
Kjaer A: Tumor volume in subcutaneous mouse xenografts measured by
microCT is more accurate and reproducible than determined by
18F-FDG-microPET or external caliper. BMC Med Imaging. 8:162008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Austah ON, Ruparel NB, Henry MA, Fajardo
RJ, Schmitz JE and Diogenes A: Capsaicin-sensitive innervation
modulates the development of apical periodontitis. J Endod.
42:1496–1502. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karai L, Brown DC, Mannes AJ, Connelly ST,
Brown J, Gandal M, Wellisch OM, Neubert JK, Olah Z and Iadarola MJ:
Deletion of vanilloid receptor 1-expressing primary afferent
neurons for pain control. J Clin Invest. 113:1344–1352. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Anke H and Sterner O: Comparison of the
antimicrobial and cytotoxic activities of twenty unsaturated
sesquiterpene dialdehydes from plants and mushrooms. Planta Med.
57:344–346. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Forsby A, Walum E and Sterner O: The
effect of six sesquiter-penoid unsaturated dialdehydes on cell
membrane permeability in human neuroblastoma SH-SY5Y cells. Chem
Biol Interact. 84:85–95. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dasari R, De Carvalho A, Medellin DC,
Middleton KN, Hague F, Volmar MN, Frolova LV, Rossato MF, De La
Chapa JJ, Dybdal-Hargreaves NF, et al: Synthetic and biological
studies of sesquiterpene polygodial: Activity of 9-epipolygodial
against drug-resistant cancer cells. ChemMedChem. 10:2014–2026.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kiso T, Fujita K, Ping X, Tanaka T and
Taniguchi M: Screening for microtubule-disrupting antifungal agents
by using a mitotic-arrest mutant of Aspergillus nidulans and novel
action of phenylalanine derivatives accompanying tubulin loss.
Antimicrob Agents Chemother. 48:1739–1748. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Castelli MV, Lodeyro AF, Malheiros A,
Zacchino SA and Roveri OA: Inhibition of the mitochondrial ATP
synthesis by polygodial, a naturally occurring dialdehyde
unsaturated sesqui-terpene. Biochem Pharmacol. 70:82–89. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boland ML, Chourasia AH and Macleod KF:
Mitochondrial dysfunction in cancer. Front Oncol. 3:2922013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Perry SW, Norman JP, Barbieri J, Brown EB
and Gelbard HA: Mitochondrial membrane potential probes and the
proton gradient: A practical usage guide. Biotechniques. 50:98–115.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Haynes CM, Titus EA and Cooper AA:
Degradation of misfolded proteins prevents ER-derived oxidative
stress and cell death. Mol Cell. 15:767–776. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Malhotra JD, Miao H, Zhang K, Wolfson A,
Pennathur S, Pipe SW and Kaufman RJ: Antioxidants reduce
endoplasmic reticulum stress and improve protein secretion. Proc
Natl Acad Sci USA. 105:18525–18530. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zeeshan HM, Lee GH, Kim HR and Chae HJ:
Endoplasmic Reticulum Stress and Associated ROS. Int J Mol Sci.
17:3272016. View Article : Google Scholar : PubMed/NCBI
|