Introduction
Oral squamous cell carcinoma (OSCC) is one of the
most aggressive neoplasms among head and neck malignant tumors
(1). Smoking and alcohol
consumption are the primary environmental factors affecting the
development of OSCC. Annually, ~540,000 new cases are diagnosed as
OSCC, and mortality rates have not reduced significantly in recent
years (2). Although advances have
been made in its treatment, the prognosis of OSCC remains poor,
with a 5-year survival ratio of nearly 50% in USA in 2009 (3). Improved understanding of the genetic
and molecular disorders of the disease is the key to early
diagnosis, appropriate treatment and improved prognosis of patients
with OSCC.
Long non-coding RNAs (lncRNAs) are a class of poorly
conserved endogenous RNAs >200 nucleotides in length that do not
encode proteins, but regulate gene expression (4). They are involved in gene regulation
by titration of transcription factors, splicing alteration,
sponging of microRNAs and recruitment of chromatin modifying
enzymes (5-7). Additionally, dysregulated lncRNAs can
influence carcinogenesis and chemoresistance in patients with
cancer (8). Increasing evidence
has demonstrated that a number of lncRNAs, including HOX transcript
antisense RNA, H19 and PVT1, are dysregulated and serve a role in
the processes of cell proliferation, differentiation, apoptosis and
cancer metastasis (9). However, it
remains unknown why lncRNAs are upregulated and which signaling
pathways are regulated by these lncRNAs.
The discovery of dysregulated lncRNAs represents a
new layer of complexity in the molecular architecture of human
disease (10). However, numerous
gaps in our current understanding of lncRNA function remain.
Placenta-specific protein 2 (PLAC2; ENSG00000223573), also known as
TINCR, is a 3.7-kb lncRNA located in Chromosome 19p13.3 (11), and no reports have examined the
role of lncRNA PLAC2 in OSCC. PLAC2 is upregulated in a number of
other cancer types, including gastric, bladder and breast cancer
(12). However, whether PLAC2 is
dysregulated and regulates the carcinogenesis of OSCC remains
unknown.
Currently, the underlying mechanism of lncRNA
dysregulation in cancer remains not defined. Histone acetylation,
such as histone H3 on lysine 27 (H3K27) acetylation (H3K27ac), is
one of the most common types of histone post-translational
modifications (13). H3K27ac was
first discovered in yeast, and is present in animals and plants
(14). H3K27ac is frequently
associated with active enhancer regulatory elements (15), and genes associated with these
enhancers are expressed at increased levels, compared with those
lacking H3K27ac. Previously, Ding et al (16) demonstrated that upregulation of the
lncRNA gastric carcinoma proliferation enhancing transcript 1
(GHET1) is due to the H3K27ac modification at the promoter region
of the GHET1 gene. However, whether other activated lncRNAs,
including PLAC2, are also induced by H3K27ac remains unknown.
In the present study, the aim was to identify the
expression level of PLAC2 in OSCC and reveal the underlying
mechanism that caused the dysregulation of PLAC2. It was determined
that PLAC2 is upregulated and activated by H3K27ac in OSCC.
Furthermore, enhanced PLAC2 promotes OSCC progression by regulating
the Wnt/β-catenin pathway.
Materials and methods
Clinical samples and ethics
statement
The present study included OSCC tissues samples from
48 patients (21 females and 27 males; age range, 35-67 years;
median age, 47 years) with OSCC who underwent partial or total
surgical resection at the Department of Oral and Maxillofacial Head
and Neck Oncology, Ninth People's Hospital, Shanghai JiaoTong
University School of Medicine (Shanghai, China) between June 2010
and July 2012. Primary cancer tissues and adjacent non-tumor
tissues (>2 cm distal from the cancer area) were collected by
surgical resection (no biopsy samples) and were used to investigate
the clinical diagnostic and prognostic role of PLAC2. No patients
were received radiotherapy or chemotherapy prior to surgical
resection. Tissue samples were immediately snap-frozen in liquid
nitrogen upon resection and then stored at -80°C until use. The
present study was approved by Research Scientific Ethics Committee
of Ninth People's Hospital, Shanghai JiaoTong University School of
Medicine. All participants signed informed consent prior to using
the tissues for scientific research.
Cell culture and regents
A total of 2 human OSCC cell lines SCC-9 and CAL-27
were purchased from Chinese Academy of Sciences (Shanghai, China).
The human normal oral epithelial keratinocytes (HOK) were purchased
from ScienCell Research Laboratories, Inc. (cat. no. 2610; San
Diego, CA, USA). CAL-27 and SCC-9 cells were cultured in Dulbecco's
modified Eagle's medium: nutrient Mixture F-12 (DMEM/F12; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 10% fetal
bovine serum (FBS; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
HOK was cultured in DMEM supplemented with 10% FBS and
penicillin-streptomycin (100 U/ml and 100 µg/ml,
respectively). The culture condition was 37°C in an atmosphere
containing 5% CO2. C646 (Selleck Chemicals, Houston, TX,
USA) was used at the concentration of 10 µmol/l for 24 h as
required.
Vector construction and cell
transfection
The synthetic silencing oligonucleotides used for
interfering PLAC2 (si-PLAC2) and cAMP-response element binding
protein-binding protein (CBP; si-CBP) were synthesized by Guangzhou
RiboBio Co., Ltd. (Guangzhou, China). Negative control si-NC was
purchased from Invitrogen (cat. no. 12935-110; Thermo Fisher
Scientific, Inc.). Additionally, the overexpression plasmid
containing oligonucleotides of PLAC2 (p-PLAC2) and negative control
vector (p-vector) was generated by Guangzhou RiboBio Co., Ltd. and
were loaded into lentivirus vectors (Lv-PLAC2 and Lv-NC,
respectively). Cells were transfected with DNA plasmids using
TransFast transfection reagent (Promega Corporation, Madison, WI,
USA). A total of 5×105 cells was seeded into each well
of a 6-well plate and transfected with respective
oligoribonucleotides upon reaching 70-80% confluence. For
transfection, ~4 µg RNA for each well was used and the final
transfection concentration was 100 nM. The transfection effects
were assessed by RT-qPCR after transfection for 24 h. The cells
were then subjected to further functional assays. The
oligonucleotides sequences used for silencing are: si-PLAC2:
5′-UAUUCCUUCAGCCAGUACCCAGGUC-3′; and si-CBP:
5′-CGGCACAGCCTCTCAGTCA-3′.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from OSCC cells using a
RNeasy plus mini kit (Qiagen, Inc., Valencia, CA, USA), according
to the manufacturer's protocol. The 20 µl RT reactions were
performed using a PrimeScript® RT reagent kit (Takara
Biotechnology Co., Ltd., Dalian, China), and incubated for 30 min
at 37°C and 5 sec at 85°C. For RT-qPCR, 2 µl diluted RT
product was mixed with 23 µl reaction buffer from a
SYBR® Green PCR kit (Takara Biotechnology Co., Ltd.) to
a final volume of 25 µl. All reactions were conducted using
an Eppendorf Mastercycler EP Gradient S (Eppendorf, Hamburg,
Germany) under the following conditions: 95°C for 30 sec, followed
by 45 cycles of 95°C for 5 sec and 60°C for 30 sec. The expression
levels of detected RNAs were normalized to GAPDH using the
comparative 2-ΔΔCq method (17). The primer sequences for RT-qPCR are
as follows: PLAC2 (sense) 5′-TGT GGCCCAAACTCAGGGATACAT-3′, and
(antisense) 5′-AGATGACAGTGGCTGGAGTTGTCA-3′; CBP (sense)
5′-GTGCTGGCTGAGACCCTAAC-3′, and (antisense)
5′-GGCTGTCCAAATGGACTTGT-3′; and GAPDH (sense)
5′-GCACCGTCAAGGCTGAGAAC-3′, and (antisense)
5′-ATGGTGGTGAAGACGCCAGT-3′.
Bioinformatics analysis
The putative binding site of H3K27ac at the promoter
region of PLAC2 gene was predicted by UCSC Genome Browser
(http://genome.ucsc.edu/). The full sequence of
PLAC2 gene can be accessed at NCBI Reference Sequence: NC_000019.10
(https://www.ncbi.nlm.nih.gov/nuccore/NC_000019.10?report=fasta&from=5558167&to=5568034&strand=true).
Cell proliferation assay
The altered cell proliferation following
transfection with si-PLAC2 was assayed using an MTT kit (cat. no.
2348-71-2; Dojindo Molecular Technologies, Inc., Rockville, MD,
USA). Cells were seeded onto 96-well plates at a density of
3×103 cells/well and cultured in 200 µl DMEM/F12
cell culture medium followed by treatment with si-PLAC2 for 12, 24,
36 or 72 h at the concentration of 100 µM under room
temperature. A total of 10 µl MTT (5 mg/ml; pH 7.4; prepared
with PBS) was added to culture the cells for 2 h at 37°C in an
atmosphere containing 5% CO2. After the medium was
removed, the precipitate was diluted in 100 µl dimethyl
sulfoxide. An enzyme-linked immunosorbent plate reader was utilized
to determine the absorbance of each well.
Cell migration and invasion assay
A wound-healing assay was used here to determine the
migration ability of OSCC cells. In brief, cells were seeded at a
density of 5×105 cell/well onto 6-well plates. After 12
h with respective vectors, the layer of cells was scratched to form
wounds using a sterile 20 µl pipette tip. The non-adherent
cells were washed away with culture medium (DMEM/F12) under room
temperature, and then the cells were further cultured for 48 h
under room temperature followed by imaging with an inverted light
confocal microscope at a ×20 magnification (Leica Microsystems
GmbH, Wetzlar, Germany) to identify the gap area. Cell invasive
ability was evaluated using a Matrigel invasion assay with Boyden
chambers (BD Biosciences; Becton, Dickinson and Company, Franklin
Lakes, NJ, USA) that had 8-μm pore size membranes with Matrigel.
1×105 cells in the serum-free Opti-MEM media (Gibco;
Thermo Fisher Scientific) were placed into the upper chamber of an
insert. DMEM/F12 culture medium containing 10% FBS was added to the
lower chamber. After 12 h of incubation, the cells that had invaded
through the membrane were stained with methanol (15 min) and 0.1%
crystal violet (5 min) under room temperature and imaged using an
inverted light confocal microscope (Leica Microsystems GmbH).
Immunofluorescence
Cells on slides were permeabilized with 0.3% Triton
X-100 (Beyotime Institute of Biotechnology, Shanghai, China) for 15
min after being fixed with 4% paraformaldehyde for 15 min under
room temperature. The cells were blocked using 5% goat serum
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) for 1 h at room temperature followed by incubation with
anti-Ki67 antibody (cat. no. ab15580) or anti-CBP antibody (cat.
no. ab2832) or anti-β-catenin (cat. no. ab32572) (all from Abcam,
Cambridge, MA, USA; all 1:100) overnight at 4°C. The slides were
then incubated with anti-rabbit Alexa Fluor 488 (cat. no.
111-545-003; 1:200; Jackson ImmunoResearch Laboratories, Inc., West
Grove, PA, USA) for 1 h at room temperature. DAPI was used for
nuclear counterstaining at room temperature for 1 h. The samples
were observed under a fluorescence microscope (×40; IX73; Olympus
Corporation, Tokyo, Japan).
Signal transduction reporter array
The Cignal 45-pathway Reporter ArrayTM (Qiagen,
Inc.; cat. no. CCA-901L) was used to determine the potential
activity alterations of the 45 untranslated regions in response to
overexpression of PLAC2, according to the manufacturer's protocols.
Cells (5×105) were suspended in Opti-MEM supplemented
with 10% of FBS and 0.1 mM non-essential amino acids
(Sigma-Aldrich; Merck KGaA), and then 50 µl of the cell
suspension, equivalent to 1×104 cells, was added into
each plate well and mixed with DNA resident (Qiagen, Inc.; cat. no.
CCA-901L). Cells were transfected with PLAC2-overexpression plasmid
(Guangzhou RiboBio Co., Ltd.) for 24 h and were subsequently
transfected with a mixture of a transcription factor-responsive
firefly luciferase reporter and a constitutively expressing
Renilla construct (Qiagen, Inc.) using the TransFast
transfection reagent. The cells were incubated for 48 h at 5%
CO2 and 37°C. The relative activity of each pathway was
decided by luciferase/Renilla, normalized by untreated controls and
measured with a Luciferase Reporter Assay System (Promega
Corporation).
Cytosolic/nuclear fraction and RNA
florescent in situ hybridization (RNA FISH)
The cellular fraction was isolated to locate the
sublocation of PLAC2. Briefly, 1×107 cells were
harvested, resuspended in 1 ml ice-cold RNase-free PBS, 1 ml buffer
C1 (1.28 M Sucrose, 40 mM Tris-HCl, pH 7.5, 20 mM MgCl2
and 4% Triton X-100) and 3 ml RNase-free water, and incubated for
15 min on ice. The cells were then centrifuged for 15 min at 3,000
× g at 4°C, and the supernatant containing the cytoplasmic
constituents and the nuclear pellet were retained for RNA
extraction.
For RNA FISH, OSCC cells were seeded and fixed with
4% paraformaldehyde for 15 min at 4°C, treated with 0.5% Triton in
PBS for 10 min at room temperature, followed by pre-hybridization.
They were then hybridized with the PLAC2 probe (5 µM)
overnight at 4°C. The PLAC2 probes were synthesized by Sangon
(Shanghai Biotech Co., Ltd., Shanghai, China). The cells were
visualized under a light confocal microscope at ×40 magnification
(Leica Microsystems GmbH).
Chromatin immunoprecipitation (ChIP)
An EZ-Magna ChIP kit (EMD Millipore, Billerica, MA,
USA) was used for the ChIP experiment, according to the
manufacturer's protocol. OSCC cells were treated with formaldehyde
and incubated for 10 min at room temperature to generate
DNA-protein cross-links. Cell lysates were then sonicated to
generate chromatin fragments of 200-300 bp and immunoprecipitated
for 1 h at room temperature with antibodies, including anti-H3K27ac
(Abcam; 1:100; cat. no. ab4729), anti-CBP (Abcam; 1:100; cat. no.
ab2832) or normal mouse IgG polyclonal antibody (EMD Millipore;
1:100; cat. no. 12-371). Precipitated chromatin DNA was recovered
and analyzed by RT-qPCR as aforementioned.
Western blots and antibodies
Radioimmunoprecipitation assay buffer (Sigma
Aldrich; Merck KGaA) was used for the cell lysis to obtain total
protein-containing lysates. Protein concentration was measured by
using the bicinchoninic acid method (Sigma Aldrich; Merck KGaA).
Subsequently, quantified protein (25 µg) was transferred
onto polyvinylidene fluoride (PVDF) membranes following the
separation process by performing a routine 10% gel electrophoresis.
The membrane was then blocked with TBS with 10% Tween-20 solution
containing 5% non-fat dry milk for 1 h at room temperature followed
by treatment with respective primary antibodies (1:1,000) at 4°C
overnight. Subsequently, they were incubated with a goat
anti-rabbit polyclonal horseradish peroxidase-conjugated secondary
antibody (1:5,000; Abcam; cat. no. ab7090) for 1 h at room
temperature. The primary antibodies are as follows:
anti-transcription factor 4 (TCF-4) antibody (Abcam; cat. no.
ab76151), anti-matrix metallopeptidase-7 (MMP-7) antibody (Abcam;
cat. no. ab205525), anti-CyclinD1 antibody (Abcam; cat. no.
ab16663), anti-MMP-9 antibody (Abcam; cat. no. ab76003),
anti-β-catenin antibody (Abcam; cat. no. ab8932) and anti-GAPDH
antibody (Invitrogen; Thermo Fisher Scientific, Inc.; cat. no.
PA1-987). Protein bands were detected using enhanced
chemiluminescence reagent (GE Healthcare, Chicago, IL, USA) and
exposed on an X-ray film. Gray analysis by image analysis was
performed using the software Gel-Pro Analyzer version 4 (United
States Biochemical, Salem, MA, USA) following scanning. The
semi-quantitative analysis was performed according to the relative
expression of objective protein and internal control protein GAPDH,
namely the ratio of objective protein and GAPDH.
In vivo tumorigenesis assay
A total of 12 male BALB/c nude mice (19-22 g; 6
weeks old) were obtained from Animal Center of the Chinese Academy
of Science (Shanghai, China). They were randomly divided into two
groups of 6 mice and housed 3 per cage. All mice were housed and
maintained under specific pathogen-free conditions at 18-22°C, with
20% humidity, a 12/12 h light/dark cycle and feeding ad
libitum. The experimental protocol was approved by the
Committee on the Animal Ethics of Shanghai Tenth People's Hospital
(Shanghai, China). PLAC2 stably expressing cell lines were
established by infecting SCC-9 cells with lentivirus as
aforementioned and selected by puromycin (Sigma-Aldrich; Merck
KGaA). OSCC cells were subcutaneously injected into right side of
the armpit regions of mice. The mice were housed for 5 weeks, then
were sacrificed and the formed tumors were stripped, and tumor mass
was measured. Xenograft volumes were evaluated by caliper
measurements of two perpendicular diameters and calculated as
formula: Volume = a x b2/2, where ‘a’ represents the
length and ‘b’ represents the width). The length of the largest
subcutaneous tumor observed was 13.1 mm, and maximum tumor burden
was 6.14% (tumor mass/body weight, 1.35/22.0 g). No multiple tumors
were exhibited in any mouse.
Another 12 male BALB/c nude mice (19-22 g; 6 weeks
old) were also obtained from Animal Center of the Chinese Academy
of Science (Shanghai, China). They were randomly divided into two
groups of 6 mice and housed 3 per cage with the same conditions
aforementioned. Experimental lung metastases were induced by
injections of a single-cell suspension (2×106 cells in
100 µl) into the mouse lateral tail vein. Cells were stably
infected with Lv-PLAC2 or Lv-NC as aforementioned, and all cell
injections were administered in a total volume of 500 µl PBS
containing 0.1% bovine serum albumin (Gibco; Thermo Fisher
Scientific, Inc.) over 60 sec (18). At 5 weeks after intravenous
injection into the nude mice, mice were sacrificed, then the lungs
were isolated and tumor nodule numbers and areas were analyzed in a
blinded fashion. For luciferase imaging, mice were injected with
luciferin (25 mg/ml in 0.1 ml PBS). At 15 min after injection,
animals were imaged using an IVIS-100 system at the UAB Small
Animal Imaging Core Facility (Birmingham, AL, USA).
All surgeries (subcutaneous or lateral tail vein
cell injection, tumor volume measurements and luciferase imaging)
were performed under sodium pentobarbital anesthesia (75 mg/kg),
and all efforts were made to minimize suffering. During anesthesia
(15-20 min) and while recovering, mice were kept warm under a red
heat lamp. To prevent the cornea from drying out, sterile eye
ointment (Viscotears®; Novartis International AG, Basel,
Switzerland) was carefully applied to the eyes during all
anesthesia interventions. The endpoint criteria for removing the
mice before finishing the study were approved by the Committee on
the Ethics of Shanghai Tenth People's Hospital. If one of the
endpoint criteria was observed the mouse was sacrificed. For our
experiments the following endpoint criteria was used: i) weight
loss >15% from the initial weight; ii) ruffled fur, hunched
back, closed eyes for more than one day; iii)
immobilization/disability because of tumor size or tumor volume
>2×103 mm3; iv) breathing difficulties; v)
no contact reactions; vi) no palm gripping reflex; and vii) signs
of infection.
All mice were sacrificed using CO2
euthanization methods. Briefly, the mice were placed in a
transparent polycarbonate shoebox euthanasia chamber
(44.0×23.5×21.0 cm). The euthanasia chamber was covered with an
acrylic lid, with ports for gas inlet and outlet. Chamber air was
replaced with CO2 at a rate of 30% of the chamber volume
per minute. Compressed CO2 gas was provided from a
cylinder (Weiler Welding, Moraine, OH) and controlled by a
CO2-specific regulator (Weiler Welding). A total of 3
mice were euthanized each time. The CO2 inflation time
was 6-8 min until the mice were confirmed as dead. The death of the
mice was verified comprehensively by the stop of breathing and
heartbeat, and the disappearance of pupil and nerve reflex.
Immunohistochemistry staining
Anti-β-catenin antibody (rabbit, polyclonal
antibody, 1:100; cat. no. ab32572; Abcam) was used for the
immunohistochemistry detection of protein expression in the tumor
tissues from mice (4 µm) at 4°C overnight. Endogenous
peroxidase was inhibited by incubation with freshly prepared 3%
hydrogen peroxide with 0.1% sodium azide for 1 h at room
temperature. Non-specific staining was blocked with 0.5% casein and
5% normal serum (Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature. The tissue samples were incubated with horseradish
peroxidase goat anti-rabbit antibody (Absin, Shanghai, China; cat.
no. abs957) for 1 h at room temperature. Staining was developed
with diaminobenzidine substrate and the sections were
counterstained with hematoxylin for 15 sec at room temperature.
Images were visualized using a Nikon ECLIPSE Ti (Nikon Corporation,
Tokyo, Japan) microscope system at a ×10 or ×40 magnification and
processed using Nikon software version 3.0.
Statistical analysis
The Kolmogorov-Smirnov test was applied for data
analysis with the distribution of each group samples. Data were
presented as the median (interquartile range). Mann-Whitney U test
was used for the comparison of datasets containing two groups. The
Kruskal-Wallis test (post-hoc Mann-Whitney U test with
Bonferroni's) was used for analyzing statistical difference among
multiple groups. Receiver Operation Characteristic (ROC) analysis
was used to evaluate the diagnostic performance of PLAC2. The
correlations between PLAC2 and CBP expression, and PLAC2 and
β-catenin expression were analyzed using Spearman's correlation
test. Kaplan-Meier analysis followed by log-rank test was performed
to determine the prognostic performance of PLAC2. P<0.05 was
considered to indicate a statistically significant difference.
Statistical analysis was performed using GraphPad Prism 5 (GraphPad
Software, Inc., La Jolla, CA, USA).
Results
PLAC2 is transcriptionally activated by
H3K27ac in OSCC
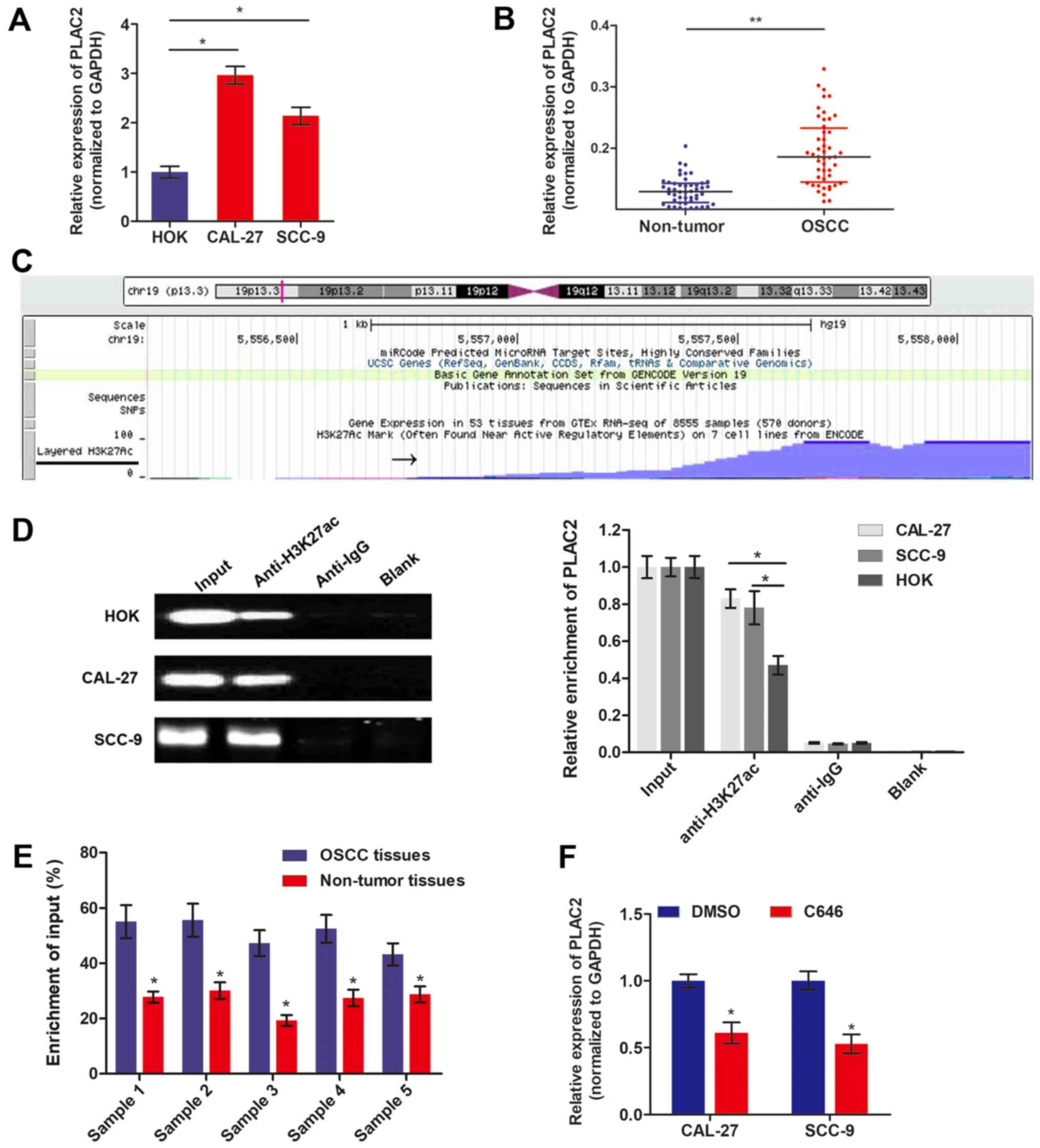
The expression level of PLAC2 in OSCC was measured
by RT-qPCR. Fig. 1A demonstrates
that PLAC2 expression is significantly increased in OSCC cells,
CAL-27 and SCC-9, compared with HOK. Additionally, PLAC2 was also
significantly increased in 48 OSCC tissues, compared with the
paired precancerous tissues (Fig.
1B). Recent research demonstrated that aberrant expression of
lncRNAs is attributed to transcriptionally activation, including
acetylation (19). To understand
the cause of PLAC2 upregulation in OSCC, the probable mechanisms
were investigated by genome bioinformatics analysis. Fig. 1C demonstrates that the promoter
region (~1,000 bp) of PLAC2 was predicted with high density of
H3K27ac enrichment. To validate the histone acetylation at the
promoter region of PLAC2, ChIP assay was performed using an
anti-H3K27ac antibody. As depicted in Fig. 1D, H3K27ac was enriched at the
promoter region of PLAC2 in both OSCC cells and normal epithelial
HOK cells. Furthermore, the enrichment level of H3K27ac was
significantly increased in both OSCC cell lines, compared with HOK
cells. ChIP assay was further conducted using 5 paired tissues,
which were identified with significantly increased PLAC2 level in
tumor tissues, compared with non-tumor tissues. As expected, the
H3K27ac enrichment level was significantly upregulated in tumor
tissues, compared with the paired non-tumor tissues (Fig. 1E). Furthermore, when OSCC cells
were treated with histone acetyltransferase (HAT) inhibitor C646,
the expression level of PLAC2 was significantly downregulated
(Fig. 1F).
CBP facilitate the H3K27ac activation of
PLAC2
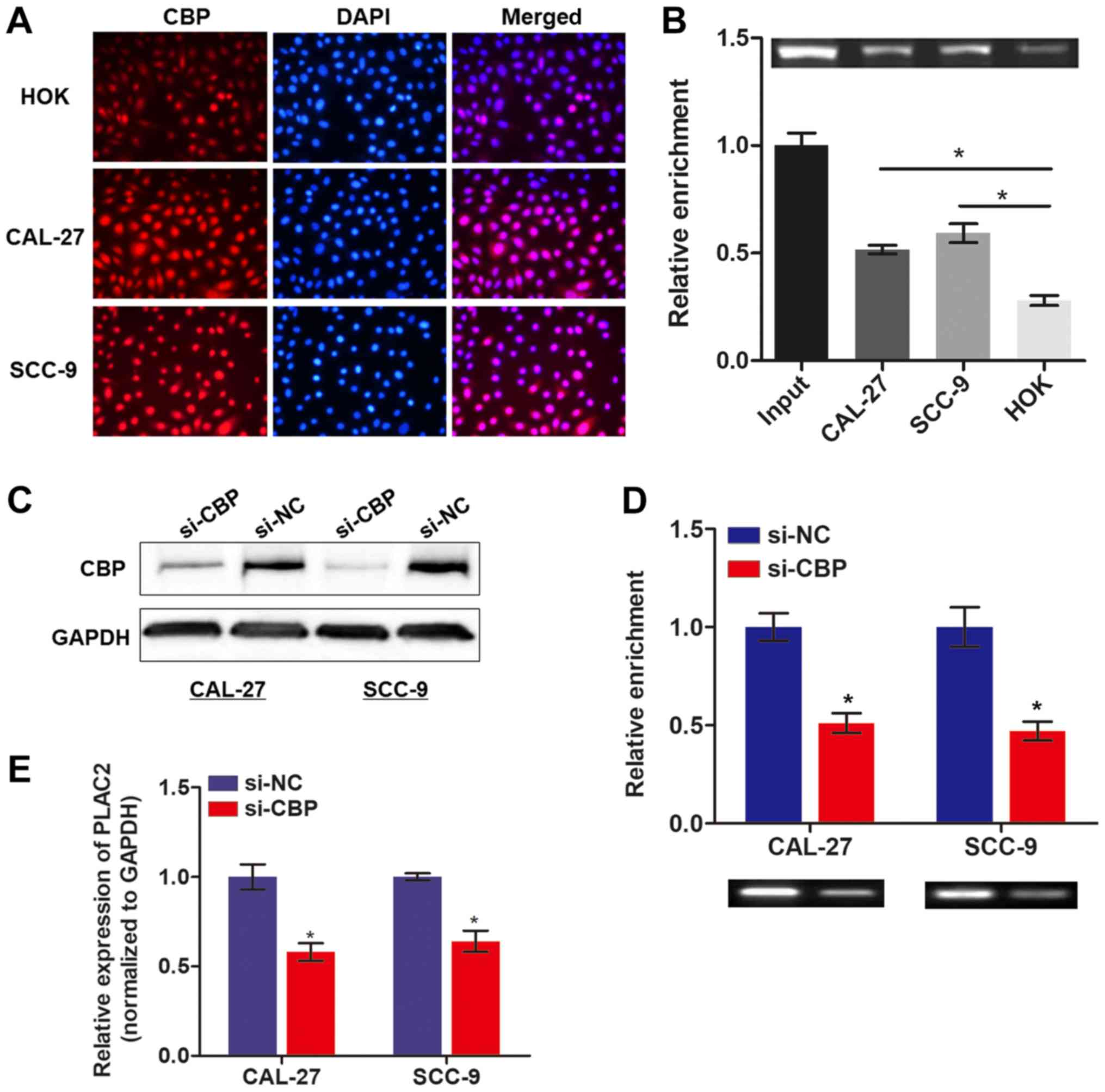
Subsequently, whether other key enzymes participate
in the process of H3K27ac was investigated. As CBP is essential for
chromatin acetylation (20),
therefore it was hypothesized that CBP may be important for the
enhanced acetylation. To test this hypothesis, immunofluorescence
was conducted to detect the CBP expression in OSCC cells and normal
HOK cells. As expected, the expression level of CBP was increased
in both OSCC cell lines, compared with HOK cells (Fig. 2A). Notably, CBP protein was
enriched by the PLAC2 gene and the enrichment level was increased
in OSCC cells, compared with normal HOK cells (Fig. 2B). Following knockdown of the CBP
gene with specific siRNA (Fig.
2C), a ChIP assay was conducted to assess the influence of CBP
on H3K27ac enrichment. As expected, the enriched H3K27ac with PLAC2
gene was significantly decreased upon knockdown of CBP (Fig. 2D). Furthermore, knockdown of CBP
also caused a significantly decreased expression of PLAC2 in OSCC
cells (Fig. 2E). Collectively, the
results indicate that upregulated PLAC2 in OSCC is attributed to
the CBP-mediated H3K27ac at the promoter region of the PLAC2
gene.
Knockdown of PLAC2 inhibits proliferation
and invasion of OSCC cells
As CAL-27 cells exhibited a relatively high
expression of PLAC2 while SCC-9 cells exhibited a low expression
level, the PLAC2-overexpression vector was built in SCC-9 cells and
the PLAC2-knockdown vector in CAL-27 cells (Fig. 3A). Subsequently, an MTT assay was
conducted to assess the effects of PLAC2 on cell proliferation over
6 days. As expected, knockdown of PLAC2 significantly suppressed
CAL-27 cell proliferation while overexpression of PLAC2 promoted
proliferation of SCC-9 cells (Fig.
3B). Additionally, an immunofluorescence assay was performed by
an antibody against Ki-67 protein, which is a well-known
proliferation marker (21).
Fig. 3C demonstrates that
knockdown of PLAC2 in CAL-27 cells suppressed the expression level
of Ki-67, whereas overexpression of PLAC2 in SCC-9 cells
demonstrated the opposite effect. Furthermore, a wound-healing
assay indicated that knockdown of PLAC2 significantly suppressed
the cell migration ability, while overexpression of PLAC2 increased
cell migration (Fig. 3D).
Similarly, a Matrigel assay demonstrated a decreased invasion
ability caused by PLAC2 knockdown and an increased invasion ability
induced by PLAC2 overexpression (Fig.
3E). These results indicate that PLAC2 may serve an oncogenic
role in OSCC.
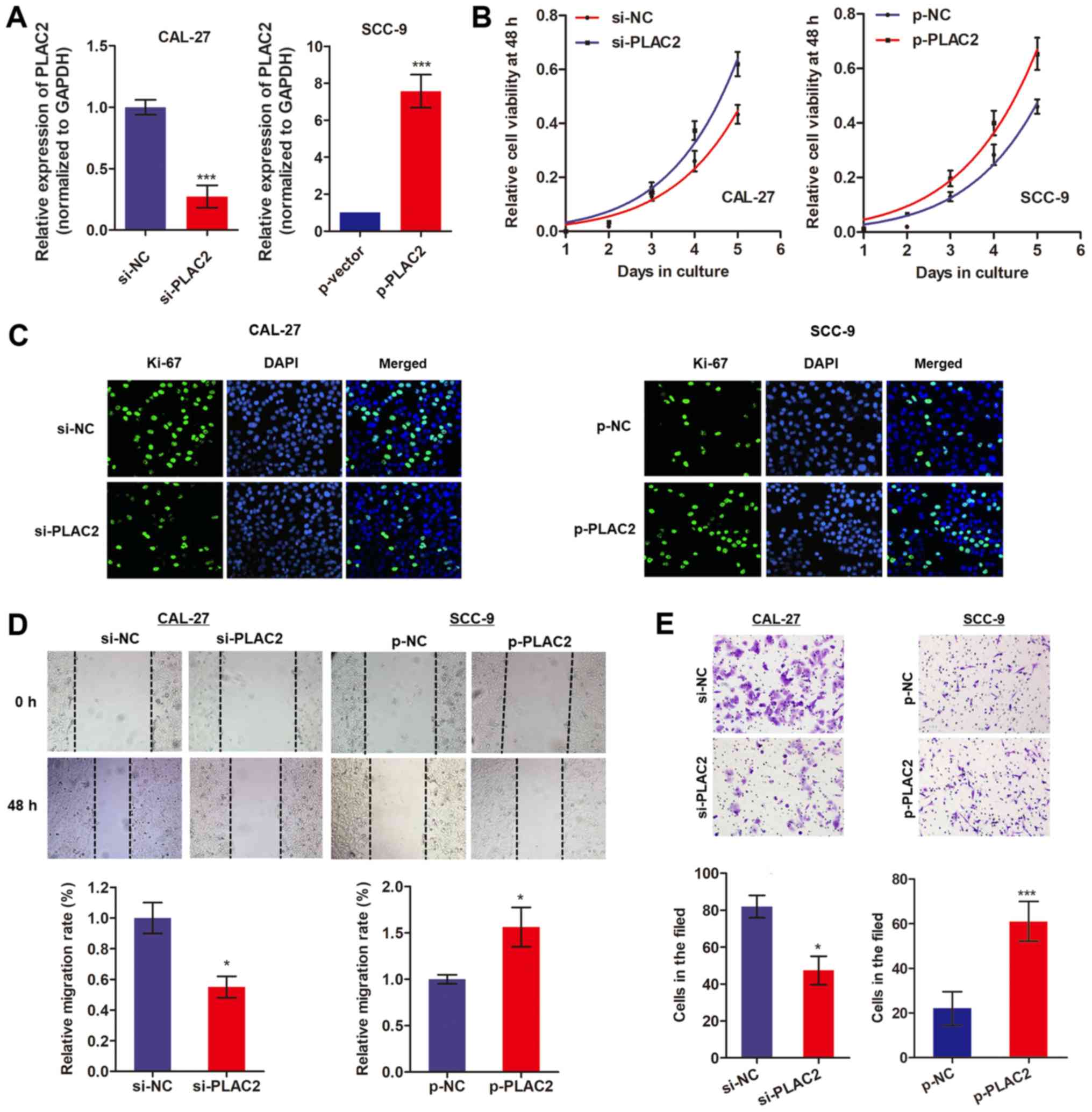 | Figure 3PLAC2 promotes proliferation and
invasion of OSCC cells. (A) The transfection efficiency of
knockdown or overexpression of PLAC2 was determined via reverse
transcription-quantitative polymerase chain reaction.
***P<0.001, compared with the control groups. (B)
Cell viability was assessed with an MTT assay in OSCC cells.
*P<0.05, compared with the control groups. (C) Ki-67
protein level was measured in OSCC cells with an immunofluorescence
assay. (D and E) Cell migration and invasion abilities were
assessed in OSCC cells using wound-healing and Matrigel assays,
respectively. *P<0.05, ***P<0.001,
compared with the control groups. PLAC2, placenta-specific protein
2; OSCC, oral squamous cell carcinoma; si, small interfering; NC,
negative control; p-NC, negative control overexpressed plasmid;
p-PLAC2, PLAC2 overexpressed plasmid. |
PLAC2 promotes OSCC malignant progression
via the activation of the Wnt/β-catenin pathway
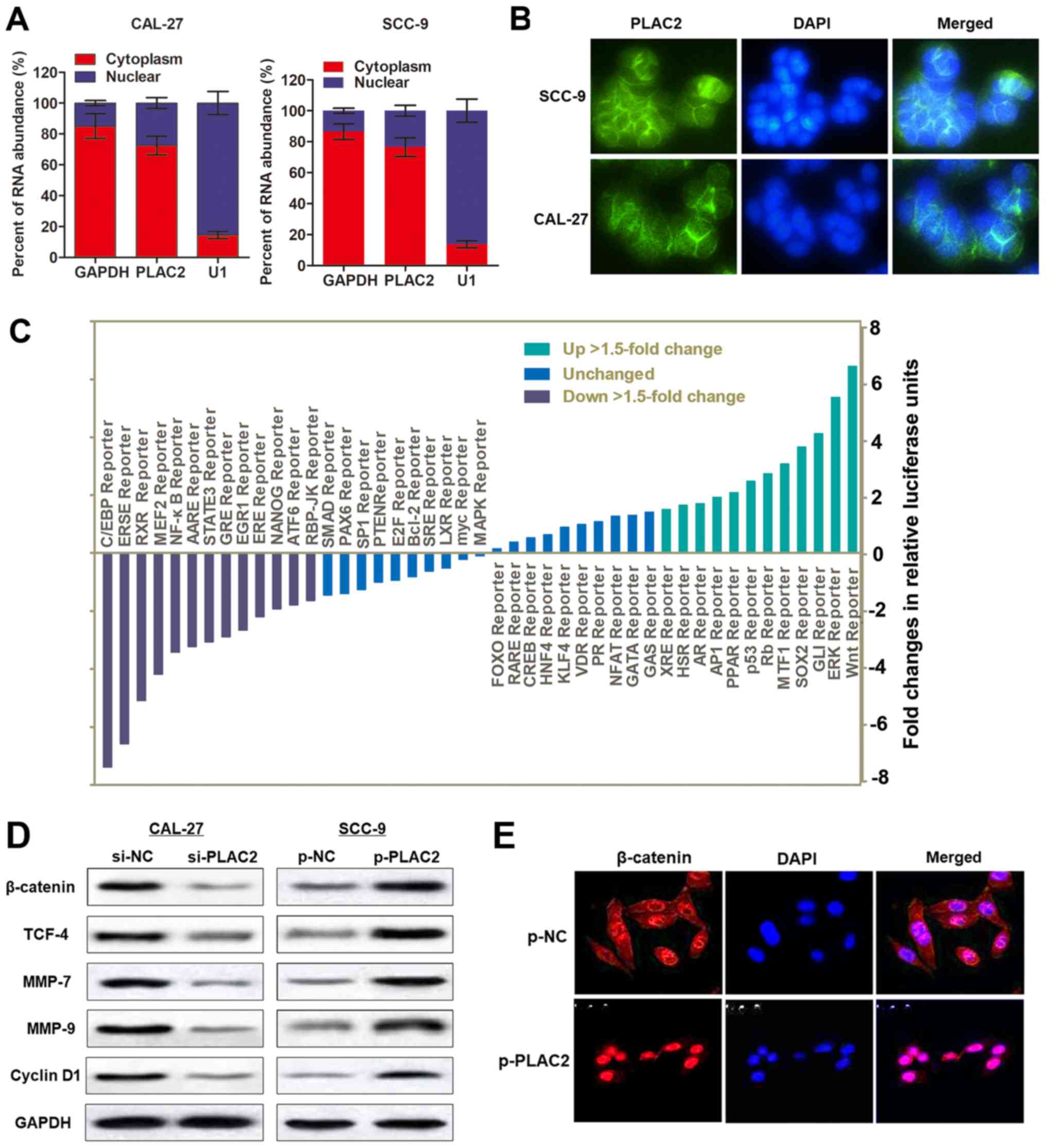
The subcellular localization of an lncRNA is
associated with its biological mechanism. To verify how PLAC2
contributes to OSCC progression, a cellular fractionation assay and
RNA-FISH were performed. It was demonstrated that PLAC2 was
distributed primarily in the cytoplasm of SCC-9 cells (Fig. 4A and B). Subsequently, a Signal
Reporter Array was conducted to simultaneously investigate the
potential signaling pathways in SCC-9 cells affected by
overexpression of PLAC2. Notably, the Wnt reporter pathway was
reported as one of the most activated pathways upon overexpression
of PLAC2 (Fig. 4C). It is
previously reported that CBP could interact with Wnt/β-catenin and
may serve as a co-activator of Wnt/β-catenin-mediated transcription
(22). Furthermore, activation of
the Wnt/β-catenin pathway could enhance the proliferation and
invasion of cancer cells (23).
Therefore, it was hypothesized that PLAC2 may promote malignant
progression of OSCC via activating the Wnt/β-catenin signaling
pathway. To test this hypothesis, western blot analysis was
conducted. The results demonstrated that overexpression of PLAC2 in
SCC-9 cells increased the expression of proteins in the
Wnt/β-catenin pathway, including β-catenin, TCF-4, MMP-7, MMP-9 and
CyclinD1, while knockdown of PLAC2 in CAL-27 cells demonstrated the
opposite effect (Fig. 4D).
Additionally, the results of the immunofluorescence staining
demonstrated that β-catenin trans-located from the cytoplasm to the
nucleus in SCC-9 cells upon overexpression of PLAC2 (Fig. 4E). Collectively, the results
indicated that PLAC2 may promote OSCC malignant development through
activating the Wnt/β-catenin pathway.
PLAC2 promotes tumor growth and
metastasis in vivo
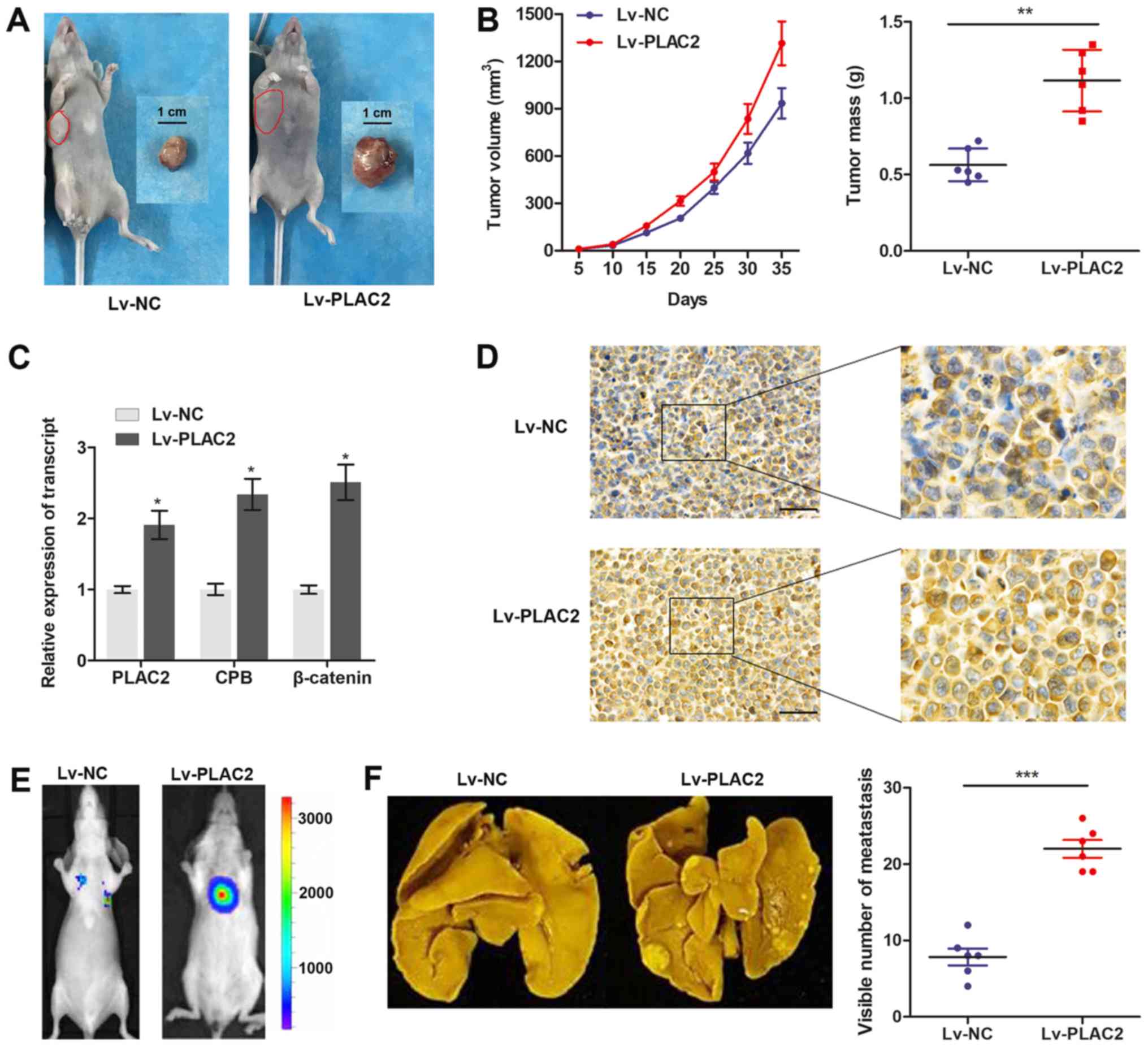
Subsequently, an in vivo xenograft model was
constructed in male BALB/C nude mice by planting SCC-9 cells, which
were stably infected with Lv-PLAC2 or Lv-NC. The mice bearing
xenografts were fed for 5 weeks and then tumor xenografts were
stripped from nude mice (Fig. 5A).
By measuring the volume and mass of the obtained xenograft, it was
verified that enhanced PLAC2 expression notably promoted the tumor
growth, compared with cells infected with Lv-NC (Fig. 5B). Furthermore, a RT-qPCR assay
revealed that the transcript expression of PLAC2, CBP and β-catenin
in implanted OSCC tissues were significantly increased in the
Lv-PLAC2 group, compared with the Lv-NC group (Fig. 5C). Immunohistochemistry analysis
further revealed a significant upregulation of β-catenin in the
tumor tissues from the Lv-PLAC2 group, compared with the Lv-NC
group (Fig. 5D). Furthermore, a
lung-metastasis model was established by injections of a
single-cell suspension into the mouse lateral tail vein. As
depicted in Fig. 5E and F, the
luciferase flux count of lung metastases and visible number of
metastases on lung surface were significantly increased in the
Lv-PLAC2 group, compared with the Lv-NC group. To conclude, the
results confirmed the functional role of PLAC2 in an in vivo
xenograft.
Upregulated expression of PLAC2 is
associated with poor prognosis of OSCC patients
Based on the aforementioned observations, the role
of PLAC2 expression in the diagnosis and prognosis of OSCC was
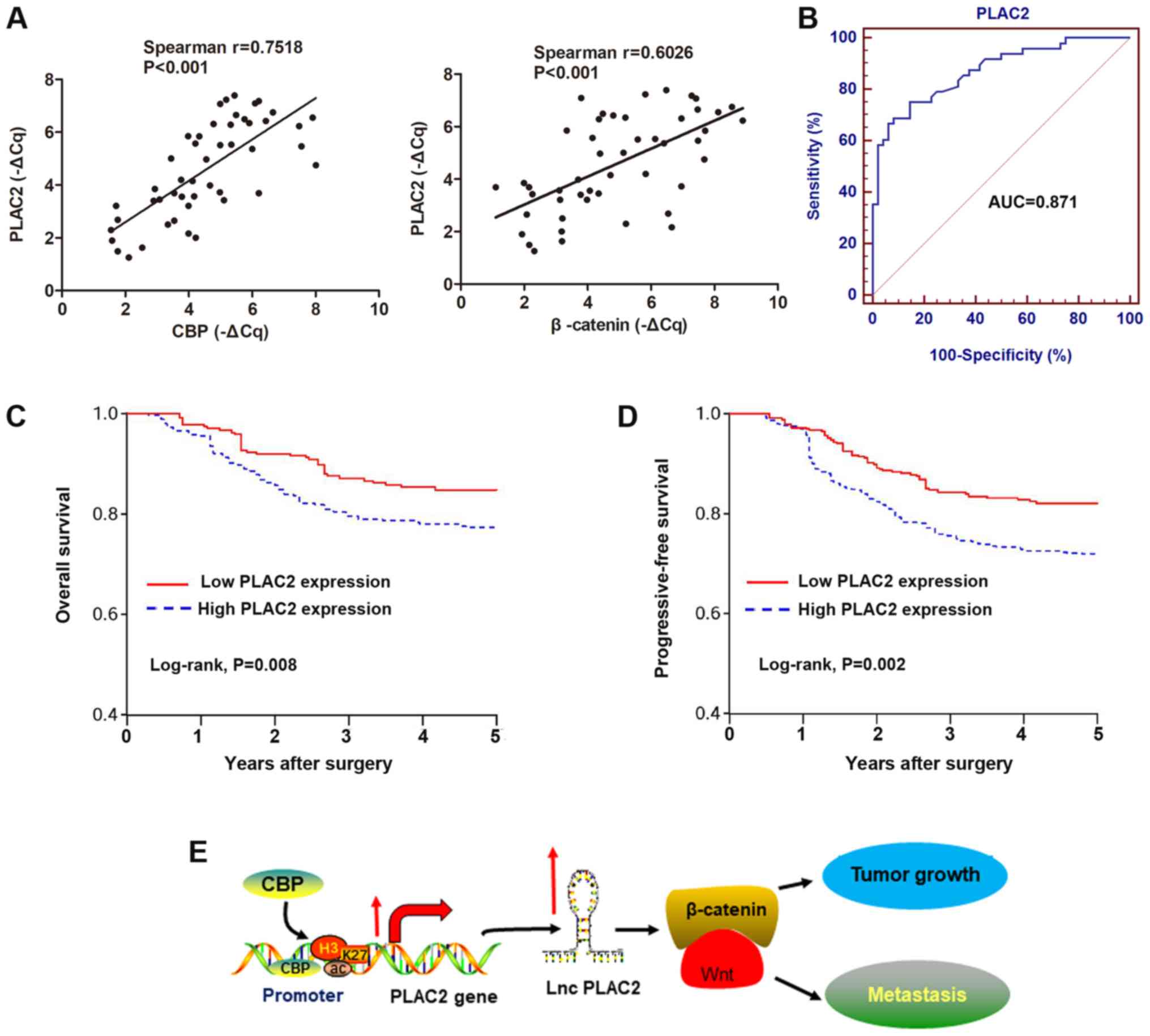
investigated. By performing Spearman's correlation test, positive
correlations were identified between the transcript expression of
PLAC2 and CBP, or PLAC2 and β-catenin in 48 OSCC tissues (Fig. 6A). Subsequently, the diagnostic
role of PLAC2 was investigated by establishing ROC curves using the
48 paired OSCC and adjacent non-tumor tissues (Fig. 6B). The area under the curve of the
ROC curve was 0.871, with a diagnostic sensitivity and specificity
reaching 75.0 and 85.4% (95% confidence interval, 0.787-0.930),
respectively. Analysis of association between PLAC2 expression and
clinical pathological factors revealed that PLAC2 expression is
positively associated with tumor size and Tumor-Node-Metastasis
stage, but not with other factors, including sex, age and
histological grade (Table I). By
using the median value of PLAC2 as a cut-off, patients with OSCC
were stratified into a high and a low PLAC2 expressing group.
Kaplan-Meier analysis of the survival data demonstrated that
patients with high PLAC2 expression were associated with reduced
overall survival and progressive-free survival rates, compared with
patients with low expression of PLAC2 (Fig. 6C and D). This indicates a potential
prognostic significance of PLAC2 for patients with OSCC.
 | Table IClinical characteristics of 48
patients and the expression of PLAC2 in primary OSCC tissues. |
Table I
Clinical characteristics of 48
patients and the expression of PLAC2 in primary OSCC tissues.
| Characteristics
(46) | Case | PLAC2 median
(range) | P-value |
|---|
| Age (years) | | | 0.830 |
| <55 | 27 | 0.15
(0.10-0.32) | |
| ≥55 | 21 | 0.16
(0.11-0.28) | |
| Sex | | | 0.185 |
| Male | 31 | 0.17
(0.10-0.32) | |
| Female | 17 | 0.15
(0.10-0.30) | |
| Tumor size | | | 0.036 |
| <5 cm | 32 | 0.13
(0.10-0.29) | |
| ≥5 cm | 16 | 0.18
(0.12-0.32) | |
| Histological
grade | | | 0.087 |
| Well | 10 | 0.14
(0.11-0.31) | |
| Moderate | 27 | 0.16
(0.10-0.30) | |
| Poor | 11 | 0.17
(0.14-0.32) | |
| TNM stage | | | 0.005 |
| I-Ⅱ | 22 | 0.12
(0.10-0.26) | |
| III-IV | 26 | 0.19
(0.14-0.32) | |
Collectively, the present study demonstrates that
the H3K27ac-induced upregulation of PLAC2 promotes OSCC cell growth
and metastasis via the activation of Wnt/β-catenin signaling
pathway (Fig. 6E). Therefore,
PLAC2 may be a potential prognostic indicator of OSCC.
Discussion
Efforts have been devoted to improve the
understanding of molecular characteristics and functional roles of
lncRNAs in cancer progression; however, their expression levels and
specific roles remain largely unknown in cancer, including OSCC
(24). Therefore, identification
of the expression level role and functional of lncRNAs, and
developing lncRNA-based targeted therapy, may be important for
improving the clinical outcome of OSCC. In the present study, the
upregulation of lncRNA PLAC2 was verified and the regulatory
mechanism that induced the upregulation PLAC2 was revealed.
Additionally, it the oncogenic role of PLAC2 in OSCC via the
activation of Wnt/β-catenin pathway was also demonstrated.
The prognosis of patients with advanced OSCC remains
dismal, and an improved understanding of the underlying mechanisms
is critical for identifying effective targets with therapeutic
potential to improve the survival of patients with OSCC. Currently,
a number of lncRNAs have been studied in OSCC, including HOX
transcript antisense RNA (25),
metastasis associated lung adenocarcinoma transcript 1 (26) and protein disulfide isomerase
family A member 3 (27); however,
further investigation is required. The underlying mechanism that
regulates the dysregulation of lncRNAs is largely unknown, which
makes it difficult to develop effective targeted therapies. In the
present study, it was identified that PLAC2 was upregulated in OSCC
tissue specimens and cell lines, and further uncovered the
epigenetic regulation of PLAC2.
The epigenetic factors, including chromatin
modifications, have not only been implicated in the spatio-temporal
regulation of gene expression, but also influences how these
transcripts are processed (28).
H3K27ac was first discovered in yeast (29), and previous advancements in DNA
sequencing technology have enabled the analysis of histone
acetylation distribution patterns through the whole genome
(30). The dysregulation of
histone H3K27ac is associated with cancer initiation and malignant
progression (13). Notably, lncRNA
GHET1 is upregulated and activated in hepatocellular carcinoma due
to the H3K27ac modification at the promoter region (16). Therefore, the promoter region of
PLAC2 was analyzed by genome bioinformatics analysis and a
high-level enrichment of H3K27ac was identified. Histone proteins
have long flexible N-terminal tails that are subject to a number of
covalent modifications, including acetylation (31). Additionally, numerous Lys residues
of histones are involved in interacting with DNA, and this
acetylation neutralizes the positive charge of Lys, resulting in
the weakening of the DNA-histone interaction and subsequent
activation of transcription (32).
This indicates that PLAC2 may be transcriptionally activated by the
modification of histone acetylation. To test this hypothesis, a
ChIP assay was performed and enriched H3K27ac at PLAC2 promoter
region was identified. Furthermore, the enrichment level was
notably increased in both OSCC cell lines, compared with normal HOK
cell lines, which is further confirmed by an increased enrichment
of H3K27ac in tumor tissues, compared with non-tumor tissues.
Enzymes that catalyze the reversible
post-translational modification of histones on lysine residues by
acetylation have received attention as potential drug targets in
cancer, aging and embryonic development (33). The histone acetylation process is
controlled by HATs and histone deacetylases (34). It is considered that acetylated
histones are associated with active transcription, whereas the
hypoacetylated histones are involved in gene repression (35). CBP is a HAT that primarily
acetylates H3 histones and has a strong association with tumor
initiation and progression (36,37).
Previously, it was reported that blocking HAT activity of CBP
offers a valid target for rationale-based designing of chemical
modulators, which could regulate p53 transcriptional functions
(38). By using steady-state HAT
assays, Bose et al (20)
demonstrated that a RNA binding region in the HAT domain of CBP, a
regulatory motif unique to CBP, allows RNA to stimulate CBP's HAT
activity. Therefore, whether CBP is involved in the H3K27ac
modification of PLAC2 promoter was investigated. As expected,
knockdown of CBP induced a decreased enrichment of H3K27ac and
thereafter a decreased expression of PLAC2.
Following verification of PLAC2 upregulation, its
function in OSCC malignant progression was investigated. As
expected, increased expression of PLAC2 notably promoted cell
proliferation, migration and invasion, whereas decreased PLAC2
demonstrated the opposite effect. The function of PLAC2 has been
investigated in other cancer types and was identified as oncogenes
in gastric cancer (39), breast
cancer (40) and hepatocellular
carcinoma (41). Notably, PLAC2
were verified as an oncogene in colorectal and lung cancer by Zheng
et al (42) and Zhu et
al (43), respectively.
However, a contradictory conclusion was developed by Zhang et
al (44) and Liu et al
(45), which indicates a
tumor-suppressive role of PLAC2 in colorectal cancer and lung
cancer, respectively. This suggests that the functional role of
PLAC2 requires further studies. The present study demonstrated that
PLAC2 served an oncogenic function in OSCC. Notably, PLAC2 promotes
progression of OSCC by the activation of Wnt/β-catenin signaling
pathway as indicates with a Signal Transduction Reporter array
followed by western blot analysis. Additionally, miRNA-relevant
experiments were not performed; however, it was considered that
miRNA could participate in the cancer initiation and progression.
Therefore, this hypothesis will be investigated in a follow-up
study.
In conclusion, it was demonstrated that the
CBP-mediated H3K27ac modification at the promoter region of PLAC2
gene induced an upregulation of PLAC2. Furthermore, enhanced PLAC2
expression promotes OSCC malignant progression via activation of
the downstream Wnt/β-catenin signaling pathway. Therefore, PLAC2
may be a potential biological biomarker for OSCC progression.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FC and SQ acquired the data and created a draft of
the manuscript. XY, XZ and JW collected clinical samples and
performed the in vitro and in vivo assays. FC, XZ and
RW analyzed and interpreted the data, and performed statistical
analysis. FC, XZ and RW reviewed the manuscript, figures, and
tables. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocol of animal study was
approved by the Committee on the Animal Ethics of Shanghai Tenth
People's Hospital (Shanghai, China). The clinical study protocol
was approved by the Clinical Research Ethics Committee of Ninth
People's Hospital, Shanghai JiaoTong University School of Medicine.
Written informed consent was obtained from each participant prior
to tissue collection.
Patient consent for publication
All patients enrolled in this study have approved
the publication of this study.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Binmadi NO and Basile JR: Perineural
invasion in oral squamous cell carcinoma: A discussion of
significance and review of the literature. Oral Oncol.
47:1005–1010. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yan L, Chen F, Liu F, Qiu Y, Wang J, Wu J,
Bao X, Hu Z, Peng X, Lin X, et al: Differences in modifiable
factors of oral squamous cell carcinoma in the upper and lower of
oral fissure. Oncotarget. 8:75094–75101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Glazer CA, Chang SS, Ha PK and Califano
JA: Applying the molecular biology and epigenetics of head and neck
cancer in everyday clinical practice. Oral Oncol. 45:440–446. 2009.
View Article : Google Scholar
|
|
4
|
Loewen G, Jayawickramarajah J, Zhuo Y and
Shan B: Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol.
7:902014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nishizawa Y, Konno M, Asai A, Koseki J,
Kawamoto K, Miyoshi N, Takahashi H, Nishida N, Haraguchi N, Sakai
D, et al: Hypoxia stimulates the cytoplasmic localization of
oncogenic long noncoding RNA LINC00152 in colorectal cancer. Int J
Oncol. 52:453–460. 2018.PubMed/NCBI
|
|
6
|
Zhang W, Cai X, Yu J, Lu X, Qian Q and
Qian W: Exosome-mediated transfer of lncRNA RP11 838N2.4 promotes
erlotinib resistance in non-small cell lung cancer. Int J Oncol.
53:527–538. 2018.PubMed/NCBI
|
|
7
|
Zhang X, Feng W, Zhang J, Ge L, Zhang Y,
Jiang X, Peng W, Wang D, Gong A and Xu M: Long non coding RNA PVT1
promotes epithelial mesenchymal transition via the TGF β/Smad
pathway in pancreatic cancer cells. Oncol Rep. 40:1093–1102.
2018.PubMed/NCBI
|
|
8
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar
|
|
9
|
Bassett AR, Akhtar A, Barlow DP, Bird AP,
Brockdorff N, Duboule D, Ephrussi A, Ferguson-Smith AC, Gingeras
TR, Haerty W, et al: Considerations when investigating lncRNA
function in vivo. eLife. 3:e030582014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hu YW, Kang CM, Zhao JJ, Nie Y, Zheng L,
Li HX, Li X, Wang Q and Qiu YR: LncRNA PLAC2 down-regulates RPL36
expression and blocks cell cycle progression in glioma through a
mechanism involving STAT1. J Cell Mol Med. 22:497–510. 2018.
View Article : Google Scholar
|
|
12
|
Kretz M: TINCR, staufen1, and cellular
differentiation. RNA Biol. 10:1597–1601. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang S, Zang C, Xiao T, Fan J, Mei S, Qin
Q, Wu Q, Li X, Xu K, He HH, et al: Modeling cis-regulation with a
compendium of genome-wide histone H3K27ac profiles. Genome Res.
26:1417–1429. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zubritskiy A and Medvedeva YA: DNA
sequence features in the establishing of H3K27ac. F1000 Res.
7:1652018. View Article : Google Scholar
|
|
15
|
Creyghton MP, Cheng AW, Welstead GG,
Kooistra T, Carey BW, Steine EJ, Hanna J, Lodato MA, Frampton GM,
Sharp PA, et al: Histone H3K27ac separates active from poised
enhancers and predicts developmental state. Proc Natl Acad Sci USA.
107:21931–21936. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ding G, Li W, Liu J, Zeng Y, Mao C, Kang Y
and Shang J: LncRNA GHET1 activated by H3K27 acetylation promotes
cell tumorigenesis through regulating ATF1 in hepatocellular
carcinoma. Biomed Pharmacother. 94:326–331. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tuomi JM, Voorbraak F, Jones DL and
Ruijter JM: Bias in the Cq value observed with hydrolysis probe
based quantitative PCR can be corrected with the estimated PCR
efficiency value. Methods. 50:313–322. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Minn AJ, Gupta GP, Siegel PM, Bos PD, Shu
W, Giri DD, Viale A, Olshen AB, Gerald WL and Massagué J: Genes
that mediate breast cancer metastasis to lung. Nature. 436:518–524.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jain AK, Xi Y, McCarthy R, Allton K,
Akdemir KC, Patel LR, Aronow B, Lin C, Li W, Yang L, et al:
LncPRESS1 Is a p53-Regulated LncRNA that Safeguards Pluripotency by
Disrupting SIRT6-Mediated De-acetylation of Histone H3K56. Mol
Cell. 64:967–981. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bose DA, Donahue G, Reinberg D,
Shiekhattar R, Bonasio R and Berger SL: RNA Binding to CBP
Stimulates Histone Acetylation and Transcription. Cell.
168:135–149.e22. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miller I, Min M, Yang C, Tian C, Gookin S,
Carter D and Spencer SL: Ki67 is a Graded Rather than a Binary
Marker of Proliferation versus Quiescence. Cell Reports.
24:1105–1112.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arensman MD, Telesca D, Lay AR, Kershaw
KM, Wu N, Donahue TR and Dawson DW: The CREB-binding protein
inhibitor ICG-001 suppresses pancreatic cancer growth. Mol Cancer
Ther. 13:2303–2314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han F, Xu Q, Zhao J, Xiong P and Liu J:
ERO1L promotes pancreatic cancer cell progression through
activating the Wnt/catenin pathway. J Cell Biochem. 119:8996–9005.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian T, Wang M, Lin S, Guo Y, Dai Z, Liu
K, Yang P, Dai C, Zhu Y, Zheng Y, et al: The Impact of lncRNA
Dysregulation on Clinicopathology and Survival of Breast Cancer: A
Systematic Review and Meta-analysis. Mol Ther Nucleic Acids.
12:359–369. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chang SM and Hu WW: Long non-coding RNA
MALAT1 promotes oral squamous cell carcinoma development via
microRNA-125b/STAT3 axis. J Cell Physiol. 233:3384–3396. 2018.
View Article : Google Scholar
|
|
26
|
Zhang Wu Y, Zhang L, Wang L, Li Y, Ren H,
Wei X, Yu F, Liu W, Wang TX, et al: Long non-coding RNA HOTAIR
promotes tumor cell invasion and metastasis by recruiting EZH2 and
repressing E-cadherin in oral squamous cell carcinoma. Int J Oncol.
46:2586–2594. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun CC, Zhang L, Li G, Li SJ, Chen ZL, Fu
YF, Gong FY, Bai T, Zhang DY, Wu QM, et al: The lncRNA PDIA3P
Interacts with miR-185-5p-to Modulate Oral Squamous Cell Carcinoma
Progression by Targeting Cyclin D2. Mol Ther Nucleic Acids.
9:100–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Piunti A and Pasini D: Epigenetic factors
in cancer development: Polycomb group proteins. Future Oncol.
7:57–75. 2011. View Article : Google Scholar
|
|
29
|
Suka N, Suka Y, Carmen AA, Wu J and
Grunstein M: Highly specific antibodies determine histone
acetylation site usage in yeast heterochromatin and euchromatin.
Mol Cell. 8:473–479. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tie F, Banerjee R, Conrad PA, Scacheri PC
and Harte PJ: Histone demethylase UTX and chromatin remodeler BRM
bind directly to CBP and modulate acetylation of histone H3 lysine
27. Mol Cell Biol. 32:2323–2334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tak YG, Hung Y, Yao L, Grimmer MR, Do A,
Bhakta MS, O'Geen H, Segal DJ and Farnham PJ: Effects on the
transcriptome upon deletion of a distal element cannot be predicted
by the size of the H3K27Ac peak in human cells. Nucleic Acids Res.
44:4123–4133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kouzarides T: Chromatin modifications and
their function. Cell. 128:693–705. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Benton CB, Fiskus W and Bhalla KN:
Targeting Histone Acetylation: Readers and Writers in Leukemia and
Cancer. Cancer J. 23:286–291. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pradeepa MM, Grimes GR, Kumar Y, Olley G,
Taylor GC, Schneider R and Bickmore WA: Histone H3 globular domain
acetylation identifies a new class of enhancers. Nat Genet.
48:681–686. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gräff J and Tsai LH: Histone acetylation:
Molecular mnemonics on the chromatin. Nat Rev Neurosci. 14:97–111.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hamzaoui H, Rizk-Rabin M, Gordon J, Offutt
C, Bertherat J and Bouizar Z: PTHrP P3 p romoter activity in breast
cancer cell lines: Role of Ets1 and CBP (CREB binding protein). Mol
Cell Endocrinol. 268:75–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang Z, Yu W, Zhang C, Zhao S, Yu Z, Xiao
X, Tang R, Xuan Y, Yang W, Hao J, et al: CREB-binding protein
regulates lung cancer growth by targeting MAPK and CPSF4 signaling
pathway. Mol Oncol. 10:317–329. 2016. View Article : Google Scholar
|
|
38
|
Hsu CH, Chang MD, Tai KY, Yang YT, Wang
PS, Chen CJ, Wang YH, Lee SC, Wu CW and Juan LJ: HCMV IE2-mediated
inhibition of HAT activity downregulates p53 function. EMBO J.
23:2269–2280. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu TP, Wang YF, Xiong WL, Ma P, Wang WY,
Chen WM, Huang MD, Xia R, Wang R, Zhang EB, et al: E2F1 induces
TINCR transcriptional activity and accelerates gastric cancer
progression via activation of TINCR/STAU1/CDKN2B signaling axis.
Cell Death Dis. 8:e28372017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y, Du Y, Hu X, Zhao L and Xia W:
Up-regulation of ceRNA TINCR by SP1 contributes to tumorigenesis in
breast cancer. BMC Cancer. 18:3672018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tian F, Xu J, Xue F, Guan E and Xu X:
TINCR expression is associated with unfavorable prognosis in
patients with hepato-cellular carcinoma. Biosci Rep. 37:372017.
View Article : Google Scholar
|
|
42
|
Zheng Y, Yang C, Tong S, Ding Y, Deng W,
Song D and Xiao K: Genetic variation of long non-coding RNA TINCR
contribute to the susceptibility and progression of colorectal
cancer. Oncotarget. 8:33536–33543. 2017.PubMed/NCBI
|
|
43
|
Zhu ZJ and He JK: TINCR facilitates
non-small cell lung cancer progression through BRAF-activated MAPK
pathway. Biochem Biophys Res Commun. 497:971–977. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang ZY, Lu YX, Zhang ZY, Chang YY, Zheng
L, Yuan L, Zhang F, Hu YH, Zhang WJ and Li XN: Loss of TINCR
expression promotes proliferation, metastasis through activating
EpCAM cleavage in colorectal cancer. Oncotarget. 7:22639–22649.
2016.PubMed/NCBI
|
|
45
|
Liu X, Ma J, Xu F and Li L: TINCR
suppresses proliferation and invasion through regulating
miR-544a/FBXW7 axis in lung cancer. Biomed Pharmacother. 99:9–17.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pollaers K, Hinton-Bayre A, Friedland PL
and Farah CS: AJCC 8th Edition oral cavity squamous cell carcinoma
staging - Is it an improvement on the AJCC 7th Edition? Oral Oncol.
82:23–28. 2018. View Article : Google Scholar : PubMed/NCBI
|