Introduction
Ovarian cancer (OC) is an aggressive gynecological
cancer with a high propensity for postmenopausal women (1). In the United States, ~22,240 women
were diagnosed with OC, and 63% of those patients were expected to
succumb to mortality from this disease (2). Surgery and platinum-based
chemotherapy remain the standard treatment for patients with
advanced stage III ovarian cancer; however, the clinical outcomes
are unsatisfactory, which is mainly due to the late diagnosis and
resistance of the cancer cells to chemotherapeutic agents (1,3,4).
Therefore, there is an urgent need for novel therapies that can
enhance the sensitivity of ovarian cancer cells to
chemotherapy.
Autophagy is a cellular catabolic process in which
autophagosomes are formed, and proteins, organelles and the cytosol
undergo lysosomal digestion and recycling (5). Autophagy serves a key role in various
physiopathological processes, including oncogenesis, cellular
development, apoptosis and survival (6,7).
Accumulating evidence indicates that autophagy, which occurs in
response to stressful conditions and specific environmental
factors, including nutrient deprivation, pathogen infection and
chemotherapeutic agents, can promote cell survival (8,9).
Despite being a protective response to stimuli, the
self-degradation undertaken via autophagy can also damage critical
cellular components, leading to autophagic cell death (type 2 cell
death; apoptosis is considered as type 1 cell death) (10). The various outcomes of autophagy
are associated with specific circumstances and certain molecular
pathways (11).
The phosphoinositide 3-kinase/protein kinase
B/mammalian target of rapamycin (PI3K/AKT/mTOR) signaling pathway
is a major regulator of autophagy in eukaryote cells, and is
involved in cell growth, viability, migration and apoptosis,
particularly during cancer development, metastasis and
chemotherapy-resistance (12,13).
PI3K activation stimulates its downstream target AKT to activate
mTOR, leading to suppression of autophagy (14). Conversely, inactivation of AKT/mTOR
signaling promotes dissociation of the Unc-51 like autophagy
activating kinase 1-autophagy-related (ATG) protein 13-FAK
family-interacting protein of 200 kDa complex from mTOR1 and induce
the autophagy process (15).
Recently, several PI3K/AKT/mTOR-targeted compounds have been
reported in problems affecting the application of chemotherapy
agents, including drug resistance, undesirable pharmacokinetics and
toxicity (16). The class-III-PI3K
inhibitor, 3-methyladenine suppresses autophagy in colon cancer and
esophageal squamous cell carcinoma (ESCC) cells, and thereby
increases their sensitivity to the chemotherapeutic agents
fluorouracil and cisplatin, respectively (17,18).
Rottlerin promotes the apoptosis of human pancreatic cancer stem
cells by inducing autophagy via the suppression of the
PI3K/AKT/mTOR signaling pathway (19).
Icariin is a flavonoid extract obtained from several
Epimedium species; it prevents neuroinflammation and
decreases cellular damage produced by oxidative stress (20). Icariin demonstrates its
antiapoptotic and anti-autophagic effects in bone marrow-derived
endothelial progenitor cells by promoting cell migration and
capillary tube formation (21).
Additionally, icariin decreases oxygen-glucose deprivation and
reperfusion-induced autophagy in rat pheochromocytoma (PC12) cells
by promoting cross talk between autophagy and apoptosis-associated
pathways mediated by B-cell lymphoma-2 (Bcl-2) (22). It also inhibits tumor oncogenesis
and the development of human ESCC by inducing stress signaling in
the endoplasmic reticulum (23).
In SKVCR cells, a potential anticancer function of icariin has been
associated with dysregulation of miR-21, phosphatase and tensin
homolog, reversion-inducing-cysteine-rich protein with kazal motifs
and Bcl-2 (24). Cisplatin, a
platinum-containing chemotherapeutic drug, is one of the most
effective agents against a wide variety of solid tumors, including
ovarian, lung, breast and colon tumors (25). In our previous study, we concluded
that icariin can serve an anti-cancerous role by inhibiting
autophagy (26); however, the
specific mechanism remains unknown.
In the present study, we report the novel finding
that icariin attenuates autophagy in SKVCR cells, which leads to an
exacerbation of cisplatin-induced cell growth inhibition by
activation of the PI3K/AKT/mTOR pathway. Improving understanding
into the biological functions of autophagy and pharmacological
regulators of autophagy may provide a basis for treating cisplatin
resistance in OC.
Materials and methods
Drug and reagents
Icariin and cisplatin were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). A Cell Counting
Kit-8 (CCK-8) was purchased from Dojindo Molecular Technologies,
Inc. (Kumamoto, Japan). Antibodies against Bax (SC-7480), caspase-3
(SC-7148), p62 (rabbit polyclonal), and Beclin-1 (rabbit
polyclonal) were obtained from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Antibody against microtubule-associated protein
1 light chain 3β (LC3B; L7543) was purchased from Sigma-Aldrich
(Merck KGaA). Antibodies against cleaved caspase-3 (cat. no. 9661),
Akt (cat. no. 4691), phosphorylated (p)-Akt (Ser473), mTOR (cat.
no. 2972), p-mTOR (Ser2448), ATG5 (8540S), and GAPDH (cat. no.
2118) were obtained from Cell Signaling Technology, Inc. (Danvers,
MA, USA). Annexin V‑fluorescein isothiocyanate (FITC) and propidium
iodide (PI) were purchased from Sigma-Aldrich (Merck KGaA).
Cell culture and drug treatment
The human multidrug-resistant phenotype OC cell line
SKVCR (SKVCR0.015) was obtained from the Cell Bank of the Chinese
Academy of Sciences (Shanghai, China). The OC cells were cultured
in α-minimum essential medium supplemented with 10% fetal bovine
serum (FBS, Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) in a humidified atmosphere containing 5% CO2 at
37°C. The experiments involved five groups of cells that were
treated as follows: i) A control group with no drug treatment
(blank); ii) an icariin treatment group (10, 20 and 30
μg/ml); iii) a cisplatin treatment group (10 μM); iv)
a group treated with cisplatin followed by icariin 1 h later
[cisplatin (5 μg/ml) + icariin (20 μg/ml) group]; and
v) a group treated with cisplatin + icariin followed by treatment
with rapamycin [cisplatin + icariin + rapamycin (0.1 μM)
group]. All cells were maintained for 48 h at 37°C in a humidified
atmosphere containing 5% CO2.
Plasmids and transfection
The cDNA sequence of ATG5 was synthesized based on
its mRNA sequences by Sangon Biotech Co. Ltd. (Shanghai, China)
with restriction sites (KpnI and BamHI) and inserted
into a pcDNA3.1 plasmid to construct a recombinant pcDNA3.1-ATG5
vector. Cells were grown in 6-well plates to 60% confluence, and
then transfected with pcDNA3.1-ATG5 or empty pcDNA plasmid as a
negative control using Lipofectamine® 2000 according to
the manufacturer's protocols (Invitrogen; Thermo Fisher Scientific,
Inc.). The mass concentration of all transfectants was 100
ng/ml.
Cell viability assay
Cells were seeded into 96-well plates at a density
of 1x104 cells/ml and incubated for 24 h at 37°C in a
humidified atmosphere containing 5% CO2; after which,
they were treated with icariin or cisplatin. Then, 10 μl of
CCK-8 solution was added to each well, and the plates were
incubated for 2 h in an incubator at 37°C. The optical density
value of each well was measured at 450 nm with a microplate reader
(BioTek Instruments, Inc., Winooski, VT, USA).
Flow cytometric analysis of the cell
cycle and apoptosis
Cells that received the different drug treatments
were harvested and fixed overnight at 4°C with 70% ethanol; after
which, they were resuspended in 500 μl PBS. For the cell
cycle assay, 10 μl RNase (10 mg/ml) was added to an aliquot
of cultured cells, and the cells were incubated for 30 min at 37°C.
The cells were then stained with 10 μl PI (1 mg/ml), and
their DNA content was analyzed with a flow cytometer (BD
Biosciences, San Jose, CA, USA). For cell the apoptosis assay,
cells were harvested, washed twice with cold PBS, and then
incubated with 100 μl binding buffer containing 5 μl
Annexin V-FITC and 1 μl PI working solution (100
μg/ml) for 30 min at room temperature in the dark. The cells
were then analyzed with a flow cytometer (BD Biosciences).
Apoptosis was analyzed using FlowJo 7.6 software (FlowJo LLC,
Ashland OR, USA), and cell cycle was processed by ModFit software
version 3.2 (Verify Software House, Inc., Topsham, ME, USA). The
apoptotic rate was calculated as follows: Apoptotic rate = early
apoptotic rate (right lower percentage) + later apoptotic rate
(right upper percentage).
Transmission electron microscopy (TEM)
analysis TEM was used to detect autophagosomes as previously
described (27,28)
Briefly, cells were fixed in 2.5% glutaraldehyde and
0.1 M cacodylate buffer for 2 h at 4°C. Following digestion with
trypsin (1:250) for 1 min at 37°C, the cells were rinsed twice with
precooled PBS, and posted‑fixed in 1% osmium tetroxide at room
temperature for 1 h. The fixed cells were then washed with PBS
buffer solution, and dehydrated with gradient alcohol (50, 70, 90
and 100%), and embedded in epoxy resin. The ultrastructure of cells
undergoing autophagy were observed and imaged under a transmission
electron microscope (JEM-1200; JEOL, Ltd., Tokyo, Japan) performed
at 80 kV.
Immunofluorescence assay
Cell autophagy was assessed by immunofluorescence
staining. Briefly, cells were fixed in 4% paraformaldehyde for 5
min at 4°C and washed three times with PBS (5 min per wash). After
washing, the cells were blocked with 5% non-fat milk in 0.1% Triton
X-100 at room temperature for 1 h, and then incubated with
anti-LC3B primary antibodies (1:1,000; Sigma-Aldrich; Merck KGaA)
for 1 h at 4°C. Then, the cells were incubated with Alexa
Fluor-labeled secondary antibodies (1:500; Sigma-Aldrich; Merck
KGaA) for 1 h and washed with PBS. DAPI was used as a counterstain
to identify the nucleus at 4°C for 10 min. The stained cells were
visualized using immunofluorescence microscopy (BX60; Olympus
Corporation, Tokyo, Japan) under five non‑overlapping fields
(magnification, x200).
Western blot analysis
Total cellular proteins were extracted using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen China) and quantified by a Bicinchoninic Acid
method. Aliquots containing ~30 μg total proteins were
subjected to SDS-PAGE, and the separated protein bands were
transferred onto a polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). Subsequently, the membrane was
blocked with 5% nonfat milk for 1 h at 4°C and then incubated with
several primary antibodies, including anti-Bax, anti-caspase-3,
anti-LC3B, anti-p62, anti-Beclin-1, anti-Akt, anti-p-Akt,
anti-mTOR, anti‑p‑mTOR, anti‑ATG5 and anti‑GAPDH at 4°C overnight.
Following washing with PBS, the membranes were incubated with a
horseradish peroxidase-conjugated secondary antibody (1:5,000;
Beijing Zhongshan Jinqiao Biotechnology Co., Ltd., Beijing, China)
for 2 h at room temperature. The chemiluminescent staining signals
were detected using an enhanced chemiluminescence detection system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). GAPDH was used as
an internal control. This experiment was repeated three times
independently.
Statistical analysis
All data are expressed as the mean ± standard
deviation. All statistical parameters were calculated using
GraphPad Prism 6.01 software (GraphPad Software Inc., La Jolla, CA,
USA). One-way analysis of variance followed by a Tukey's post-hoc
test was used for data analysis. P<0.05 was considered to
indicate a statistically significant difference. In experiments
were repeated three times independently.
Results
Icariin treatment suppresses cell
viability and cell cycle progression, and activates apoptosis and
autophagy in OC cells
To investigate whether icariin could exert antitumor
activity in SKVCR cells, the CCK-8 assay was used to evaluate cell
viability following treatment with different concentrations of
icariin. As presented in Fig. 1A,
icariin treatment significantly decreased the percentage of viable
SKVCR cells in a dose-dependent manner, and the half-maximal
inhibitory concentration (IC50) value of icariin was ~60
μg/ml. Based on this IC50 value, the effects of
icariin on cell cycle progression and apoptosis by were analyzed
flow cytometry. Furthermore, icariin treatment significantly
increased the percentage of apoptotic cells, including cells in
early apoptosis or late apoptosis (P<0.001; Fig. 1B). As presented in Fig. 1C, the percentage of SKVCR cells in
G0/G1 phase significantly increased following icariin treatment,
which was accompanied with a reduction in the number of S phase
cells in the icariin group when compared with the control group
(P<0.001). TEM analysis was performed to observe whether icariin
affected the ultrastructure of SKVCR cells. As presented in
Fig. 1D, autophagic vacuoles were
detected in the blank control group; however, fewer were observed
following icariin treatment. We further confirmed the occurrence of
autophagy via an immunofluorescence assay using staining with
anti‑LC3B. When compared with the blank group, the ratio LC3B I/II
was significantly lower in the icariin group, suggesting that
icariin treatment could significantly reduce the occurrence of
autophagy (Fig. 1E and F).
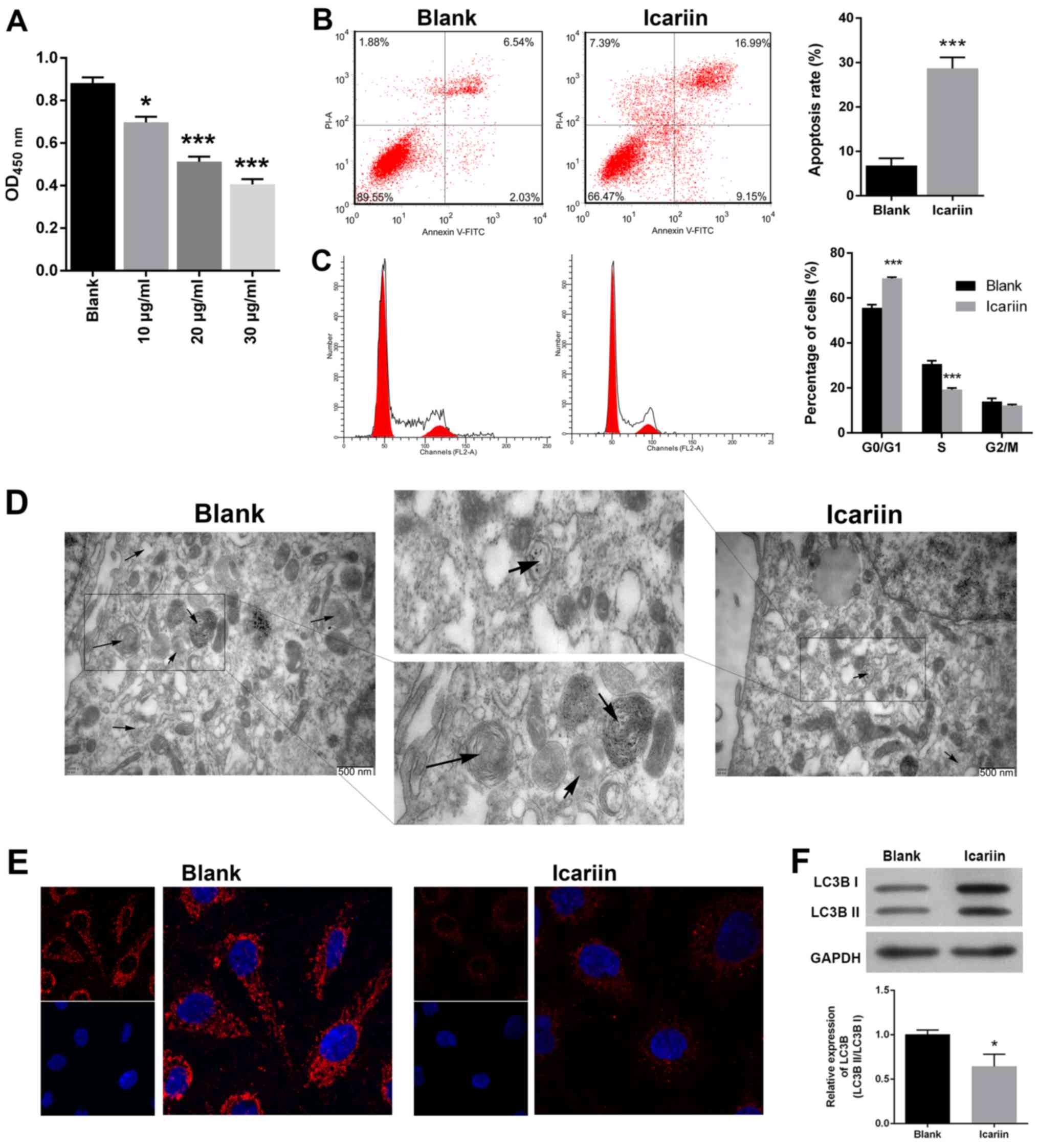 | Figure 1Effects of icariin on cell viability,
cell cycle distribution, apoptosis and autophagy in SKVCR cells.
(A) Cell Counting Kit-8 assay was used to determine the
proliferative ability of SKVCR cells treated with icariin (10, 20,
and 30 μg/ml, respectively). (B) Flow cytometry combined
with Annexin V-FITC and PI staining was used to analyze cell
apoptosis in SKVCR cells treated with icariin (20 μg/ml).
(C) Flow cytometry combined with PI staining was used to analyze
the cell cycle distribution of SKVCR cells treated with icariin (20
μg/ml). (D) A transmission electron microscope image of cell
autophagosomes (20 μg/ml). Scale bar, 500 nm. (E and F) An
immunofluorescence assay was used to examine LC3B expression in
SKVCR cells treated with icariin (20 μg/ml). Magnification,
x200. *P<0.05, ***P<0.001 vs. Blank
group. FITC, fluorescein isothiocyanate; LC3B,
microtubule‑associated protein 1 light chain 3β; PI, prop-idium
iodide; OD, optical density. |
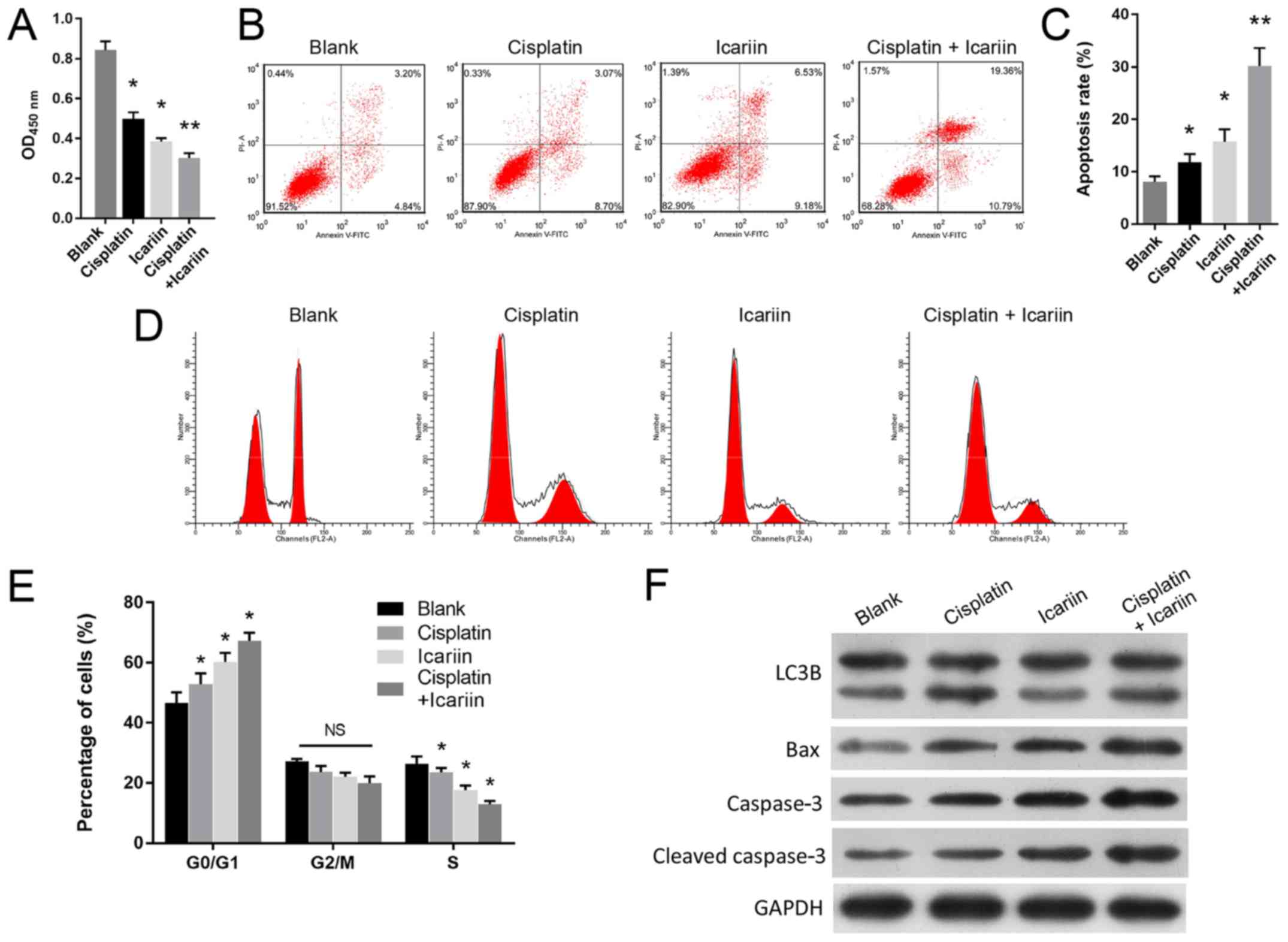
Icariin treatment sensitizes OC cells to
cisplatin
In addition, how icariin may mediate the viability
of SKVCR cells treated with cisplatin was investigated. A CCK-8
assay revealed that combined treatment with icariin and cisplatin
significantly suppressed the viability of SKVCR cells when compared
with cisplatin treatment alone (P<0.001; Fig. 2A). This indicated that icariin
enhanced the inhibitory effects of cisplatin on SKVCR cell
viability. Additionally, icariin treatment significantly induced
cell apoptosis (P<0.01; Fig. 2B and
C) and cycle arrest at the G0/G1 phase (P<0.001; Fig. 2D and E) in cisplatin-treated SKVCR
cells. Western blot analysis suggested that the expression levels
of Bax and caspase-3 proteins were notably upregulated (Fig. 2F). Furthermore, LC3B II was notably
downregulated by icariin compared with the blank group, and cells
treated with cisplatin and icariin presented markedly higher LC3B
II expression compared with cells treated with icariin (Fig. 2F).
Enhanced autophagy reduces the
sensitivity of ovarian cancer cells to icariin
The aforementioned results demonstrated that icariin
treatment could notably sensitize ovarian cancer cells to cisplatin
and inhibit autophagy. As autophagy is negatively correlated with
the efficacy of chemotherapy (29,30),
it was hypothesized that enhanced autophagy may affect the
sensitivity of OC cells to icariin and cisplatin. As presented in
Fig. 3A and B, icariin markedly
suppressed cisplatin-induced autophagy, while rapamycin, an
autophagy activator, notably alleviated the suppressive effects of
icariin on SKVCR cells, as determined by TEM and immunofluorescence
analysis, respectively. In addition, flow cytometry was used to
analyze cell cycle distribution and apoptosis rates. As presented
in Fig. 3C-F, the enhanced
autophagy induced by rapamycin significantly reversed the
inhibitory effects of icariin on cell cycle progression and
apoptosis (P<0.05). The molecular mechanisms underlying the
effects of rapamycin on autophagy were investigated via western
blotting. As presented in Fig. 3G,
the levels of LC3B, Beclin-1 and ATG5 expression were
downregulated, while that of p62 was upregulated in the cisplatin +
icariin group, when compared with the groups treated with cisplatin
or rapamycin alone, suggesting that icariin treatment could
suppress autophagy induced by cisplatin or rapamycin. Furthermore,
icariin activated the AKT/mTOR pathway, as demonstrated by
upregulation of p-AKT and p-mTOR; however, rapamycin treatment
reversed the effects of icariin on the expression of
autophagy-associated proteins. Collectively, these data suggested
that the increase in chemosensitivity induced by icariin could be
reversed by the enhanced autophagy triggered by rapamycin (Fig. 3G).
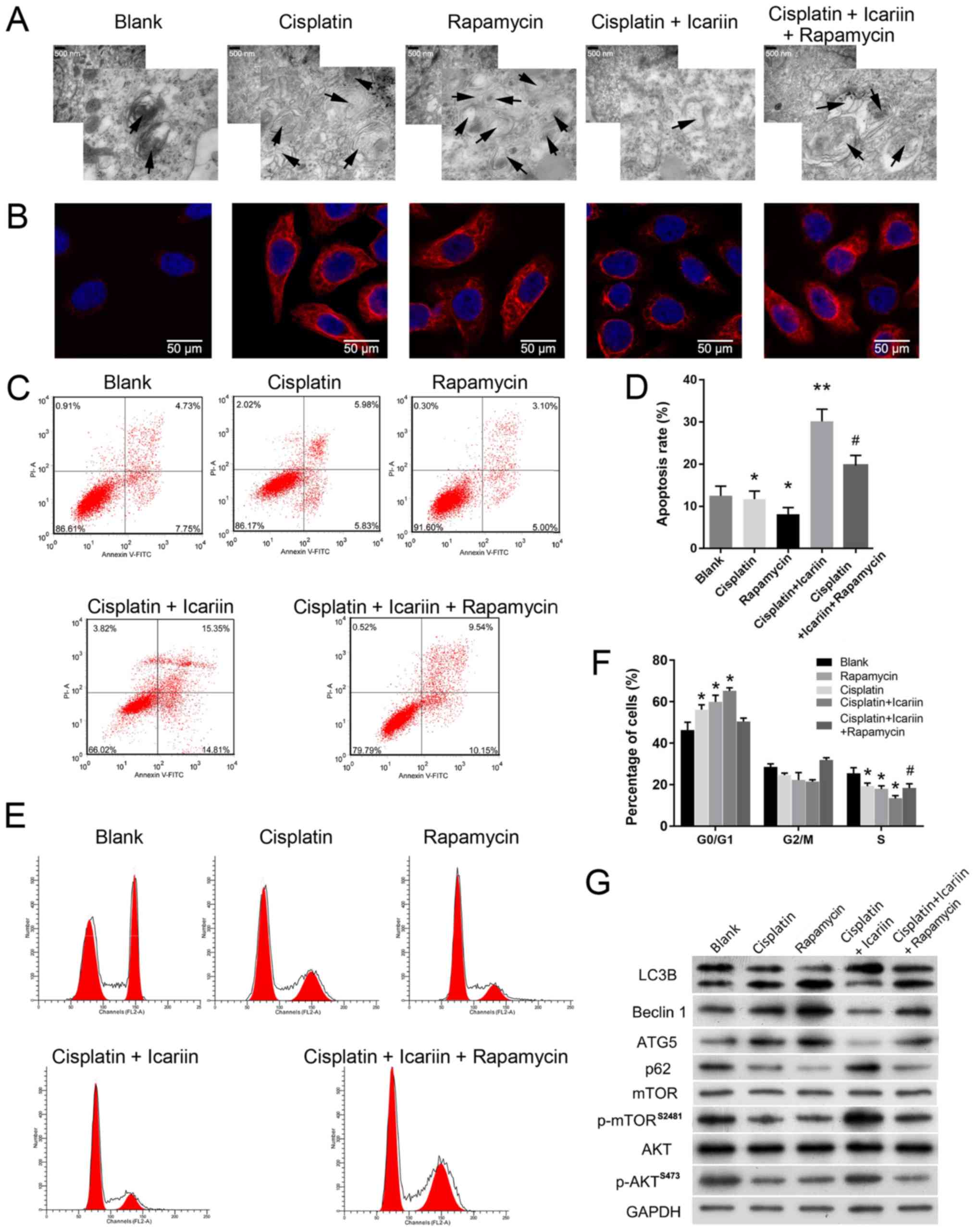 | Figure 3Enhanced autophagy reduces the
sensitivity of ovarian cancer cells to icariin. (A) Transmission
electron microscope images of autophagosomes in SKVCR cells. Scale
bar, 500 nm. (B) LC3B expression in SKVCR cells was examined using
an immunofluorescence assay. Scale bar, 50 μm. Flow
cytometry was used to analyze (C and D) the cell apoptosis and (E
and F) the cell cycle distribution in SKVCR cells. (G) Expression
levels of LC3B, Beclin-1, ATG5, p62, AKT, p-AKT, mTOR and p-mTOR
proteins in SKVCR cells were determined by western blot analysis.
GAPDH was used an internal control. Icariin, 20 μg/ml,
cisplatin, 5 μg/ml. *P<0.05 vs. Blank group;
**P<0.01 vs. cisplatin group; #P<0.05
vs. cisplatin + icariin group; AKT, protein kinase B; ATG5,
autophagy-related 5; LC3B, microtubule-associated protein 1 light
chain 3β; p, phosphorylated; mTOR, mammalian target of
rapamycin. |
Upregulation of ATG5 reduces the
sensitivity of OC cells to icariin
ATG5 is one of several well-established
autophagy-associated proteins, and is critical for the biogenesis
of autophagosomes (31). It was
reported that knockdown of ATG5 promoted cisplatin-induced
apoptotic death of human lung cancer cells (32). To further confirm the association
between autophagy and chemotherapy, overexpression of ATG5 was
conducted in SKVCR cells following treatment with icariin +
cisplatin. As expected, the results of TEM (Fig. 4A) and immunofluorescence revealed
that cisplatin or overexpression of ATG5 enhanced autophagy
compared with the blank group; however, overexpression of ATG5
reversed the decreased autophagy in the icariin + cisplatin group
(Fig. 4A and B). Consistent with
this finding, the promoting effects of icariin on cell apoptosis
(P<0.05; Fig. 4C and D) and the
cell cycle (P<0.01; Fig. 4E and
F) in cisplatin-resistant SKVCR cells was significantly
alleviated by ATG5 overexpression. Western blot analysis was also
conducted to examine the expression of autophagy- and
apoptosis-associated proteins. As presented in Fig. 4G, compared with blank group, the
levels of LC3B, Beclin-1 and ATG5 were markedly increased in
cisplatin-treated cells or ATG5-overexpressing cells, which was
accompanied with p62 downregulation. While, cells treated with
cisplatin and icariin exhibited weaker levels of
autophagy-associated protein compared with ATG5-overexpressing
cells. The activity of the AKT/mTOR signaling cascade in
icariin-treated cells was notably enhanced as demonstrated by
increased expression levels of activated p-AKT and p-mTOR, but was
impaired by ATG5 overexpression. Importantly, ATG5, a promoter of
autophagy, was downregulated by icariin treatment. Collectively,
these results further demonstrated that icariin had enhanced the
sensitivity of SKVCR cells to cisplatin, partially by inhibiting
autophagy.
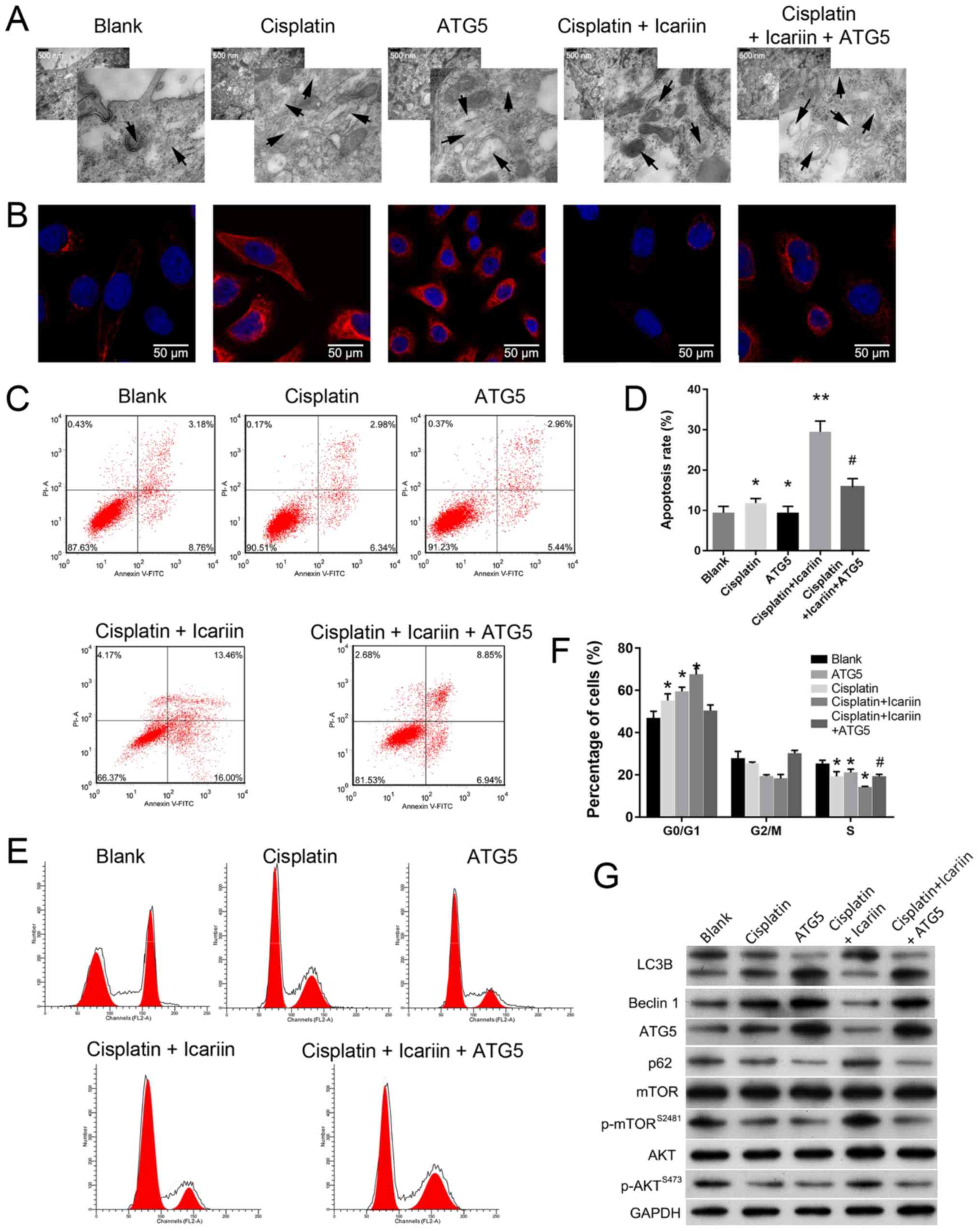 | Figure 4Upregulation of ATG5 reduces the
sensitivity of ovarian cancer cells to icariin. (A) Transmission
electron microscopy analysis of autophagosomes in SKVCR cells.
Scale bar, 500 nm. (B) LC3B expression in SKVCR cells was examined
using via immunofluorescence analysis. Scale bar, 50 μm.
Flow cytometry was used to analyze (C and D) the cell apoptosis and
(E and F) the cell cycle distribution in SKVCR cells. (G) The
levels of LC3B, Beclin-1, ATG5, p62, AKT, p-AKT, mTOR, p-mTOR and
caspase-3 proteins in SKVCR cells were determined by western blot
analysis. Icariin, 20 μg/ml, cisplatin, 5 μg/ml.
*P<0.05 vs. Blank group; **P<0.01 vs.
cisplatin group; #P<0.05 vs. cisplatin + icariin
group. AKT, protein kinase B; ATG5, autophagy-related 5; LC3B,
microtubule-associated protein 1 light chain 3β; p, phosphorylated;
mTOR, mammalian target of rapamycin. |
Discussion
In the present study, the underlying mechanism as to
how icariin modulates the viability, apoptosis and autophagy of
SKVCR cells was investigated, and provide a possible mechanism for
how icariin increases the sensitivity of SKVCR cells to cisplatin
was proposed. In our previous study, it was suggested that icariin
can inhibit malignant phenotypes in ovarian cancer cell lines by
inhibiting autophagy (26). To the
best of our knowledge, the present study is the first to
demonstrate that icariin can enhance the sensitivity of SKVCR cells
to cisplatin via inhibiting autophagy by inducing cell apoptosis
and inhibiting the cell cycle.
Cisplatin is a platinum-based drug that forms inter-
and intra-strand DNA crosslinks, and is used as a first-line
chemotherapeutic agent in treating epithelial ovarian carcinoma
(22). Compared with cisplatin
treatment, the results indicated that treatment with cisplatin +
icariin induced apoptosis in SKVCR cells and inhibited the
viability of those cells. Furthermore, we observed a corresponding
decrease in autophagy, which suggests a role for icariin in
regulating autophagy in cisplatin-treated SKVCR cells.
Autophagy is a lysosomal pathway that delivers
cellular components, including proteins, organelles and cytosol to
lysosomes, where they are degraded to maintain cellular homeostasis
(33). It has been reported that
that autophagy is modulated by extracellular and intracellular
stress, as well as signaling pathways (34). A variety of proteins are involved
in the progression of autophagy. ATG5 is essential for
autophagosome formation and autophagy promotion (35). Beclin-1 is a central component of
the PI3K-III complex, which recruits several autophagy proteins
during the formation of autophagosomes (36). An efficient autophagy recycling
process relies on numerous proteins, including LC3B, which is an
autophagy indicator that is cleaved into LC3B I and LC3B II during
autophagy (37). LC3B serves an
essential role in the biogenesis of autophagosomes and recruitment
of autophagosome cargo (37). A
previous study demonstrated that p62 can bind to ubiquitin and
LC3B, and a lack of autophagy is usually accompanied with the
downregulation of p62 (38). In
the present study, when compared with OC cells treated with
cisplatin alone, treatment with cisplatin + icariin exhibited
downregulated levels of LC, Beclin-1 and ATG5 expression that were
accompanied by upregulated p62 expression, indicating inactivation
of the autophagic pathway. These results are consistent with the
autophagy phenomenon that was observed by electron microscopy.
Interestingly, increased levels of p-AKT and p-mTOR protein were
evident in cells treated with cisplatin + icariin when compared
with cisplatin alone. The phosphorylation of AKT and mTOR is
considered as a biomarker for the activation of AKT/mTOR signaling,
as well as for AKT and mTOR activity (39,40).
The AKT/mTOR pathway serves a critical role in cancer development
and functions as a major regulator of autophagy progression
(12,41). Evidence has indicated that AKT can
be inhibited by rapamycin, an inhibitor of mTOR (18); thus autophagy may be induced via
inhibition of AKT/mTOR pathway. In the present study inhibition of
the PI3K/AKT/mTOR pathway was proposed to activate autophagy,
whereas induction of the pathway suppressed autophagy (42).
In the present study, activation of the AKT/mTOR
pathway may have been responsible for the decreased autophagy in
cells treated with cisplatin + icariin when compared with cisplatin
alone. Therefore, icariin may inhibit autophagy via the AKT/mTOR
pathway to re-sensitize SKVCR to cisplatin. A recent study
demonstrated that Tanshinone IIA mediated autophagy via the
PI3K/AKT/mTOR pathway in oral squamous cancer (43). Similarly, the PI3K/AKT/mTOR pathway
has been proposed to be involved in the autophagic process mediated
by a neuroactive compound (44).
Furthermore, it has been demonstrated that the prototype mTOR
inhibitor, rapamycin, can initiate cellular autophagy (45). The rapamycin-mediated attenuation
of mTOR leads to overexpression of LC3B II and Beclin-1 in the
infant brain (42). In the present
study, rapamycin treatment notably reversed the effects of icariin
on p-AKT, p-mTOR, and autophagy-associated protein expression. This
further confirmed that icariin had activated the AKT/mTOR pathway,
induced the downregulation of LC3B, Beclin-1 and ATG5, while
upregulating p62. Therefore, icariin-mediated inhibition of
autophagy may occur via activation of the AKT/mTOR pathway.
Additionally, overexpression of ATG5 was observed to
impair the phosphorylation of AKT and mTOR, upregulate LC3B and
Beclin-1, and downregulate p62. ATG5 is anchored to the phagophore
membrane in a complex with ATG12 and ATG16 (46). Hu et al (47) reported that ATG5 upregulation
affects pseudotube formation by enhancing the activation of AKT in
endothelial progenitor cells. In bovine aortic endothelial cells,
AKT activation and ROS production are stimulated by elevated ATG5
levels (48). In the present
study, we also found that ATG5 overexpression decreased the
activation of AKT. Furthermore, ATG5 accumulation may lead to a
negative feedback to the upstream signal involving AKT and mTOR.
Therefore, the phosphorylation of downstream mTOR was reduced,
which led to activation of the autophagic pathway via inhibition of
AKT/mTOR signaling in SKVCR cells with overexpression of ATG5.
Activation of the apoptosis process has been
reported to be responsible for the cytotoxic effects of
chemotherapy on tumor cells; however, alterations in the apoptotic
components are usually associated with the sensitivity of tumor
cells to chemotherapy (49). It
has been revealed that apoptosis is negatively correlated with the
AKT/mTOR pathway in numerous types of cancer (50). For example, cell proliferation was
stimulated and apoptosis was suppressed by leptin via its ability
to activate the PI3K/AKT/mTOR pathway (50). Thioridazine prevented the growth of
cervical and endometrial cancer cells via its ability to induce
apoptosis mediated by the PI3K/AKT/mTOR/p70S6K pathway (51). We observed that, compared with
cisplatin alone, treatment with cisplatin + icariin inhibited cell
viability, and also activated apoptosis and the AKT/mTOR pathway.
The present study proposed that the inhibition of viability and
induction of apoptosis were not directly associated with the
AKT/mTOR pathway. Crosstalk between autophagy and apoptosis has
been demonstrated (10). Under
certain circumstances, such as nutrient deficiency, abrogation of
autophagy can accelerate cell death and activate certain
apoptosis-associated enzymes, including caspases (52). Tumor cells can enhance their basal
levels of autophagy for the purpose of maintaining their
mitochondrial function and energy homeostasis to meet the elevated
metabolic demands of growth and viability (53,54).
Conversely, autophagy-induced apoptosis was proposed as a method
for treating cancer. Autophagic cell death is another type of cell
death, which is morphologically different from apoptosis and was
reported to be induced by high levels of autophagy (55). Caspase-3 is a key catalyst of
apoptosis in mammalian cells (56). Our results suggested that tumor
cells may induce autophagy for the purpose of surviving when
treated with cisplatin, whereas icariin treatment decreased
autophagy, thereby increasing the sensitivity of tumor cells to
cisplatin rather than their propensity to autophagic cell death,
which is characterized by the dysregulation of apoptosis-associated
proteins. Icariin was proposed to enhance the susceptibility of
SKVCR cells to the chemotherapeutic agent cisplatin by regulating
autophagy induced by activation of the AKT/mTOR pathway.
In conclusion, our results are the first to
demonstrate that icariin enhanced ovarian cell sensitivity to
cisplatin by reducing autophagy in SKVCR cells by mediating the
AKT/mTOR/ATG5 signaling pathway, to the best of our knowledge.
Autophagy may serve a major role as a chemotherapy sensitization
mechanism in SKVCR cells treated with icariin. Thus, effective
suppression of autophagy may provide a prospective method for
enhancing the cisplatin-induced inhibition of SKVCR cell growth and
be used to improve the clinical outcomes of chemotherapy.
Funding
The present study was supported by the Shenzhen
Basic Research Program (grant no. 20160427191320225).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SJ made substantial contributions to the design of
the present study. SJ and HC performed all the experiments and
collected all the data; SJ, DF, HC and SD analyzed the data. SJ and
DF drafted the manuscript, which was revised by HC. All authors
approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
CCK-8
|
Cell Counting Kit-8
|
|
ESCC
|
esophageal squamous cell carcinoma
|
|
IC50
|
half-maximal inhibitory
concentration
|
|
mTOR
|
mammalian target of rapamycin
|
|
OC
|
ovarian cancer
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
PI
|
propidium iodide
|
References
|
1
|
Jelovac D and Armstrong DK: Recent
progress in the diagnosis and treatment of ovarian cancer. CA
Cancer J Clin. 61:183–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Agarwal R and Kaye SB: Ovarian cancer:
Strategies for overcoming resistance to chemotherapy. Nat Rev
Cancer. 3:502–516. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang ZJ, Chee CE, Huang S and Sinicrope
FA: The role of autophagy in cancer: Therapeutic implications. Mol
Cancer Ther. 10:1533–1541. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rubinsztein DC: The roles of intracellular
protein-degradation pathways in neurodegeneration. Nature.
443:780–786. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lum JJ, Bauer DE, Kong M, Harris MH, Li C,
Lindsten T and Thompson CB: Growth factor regulation of autophagy
and cell survival in the absence of apoptosis. Cell. 120:237–248.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Onodera J and Ohsumi Y: Autophagy is
required for maintenance of amino acid levels and protein synthesis
under nitrogen starvation. J Biol Chem. 280:31582–31586. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim KW, Myers CJ, Jung DK and Lu B:
NVP-BEZ-235 enhances radiosensitization via blockade of the
PI3K/mTOR pathway in cisplatin-resistant non-small cell lung
carcinoma. Genes Cancer. 5:293–302. 2014.PubMed/NCBI
|
|
12
|
Shinojima N, Yokoyama T, Kondo Y and Kondo
S: Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in
curcumin-induced autophagy. Autophagy. 3:635–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Engelman JA, Luo J and Cantley LC: The
evolution of phosphatidylinositol 3-kinases as regulators of growth
and metabolism. Nat Rev Genet. 7:606–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li X, Hu X, Wang J, Xu W, Yi C, Ma R and
Jiang H: Inhibition of autophagy via activation of PI3K/Akt/mTOR
pathway contributes to the protection of hesperidin against
myocardial ischemia/reperfusion injury. Int J Mol Med.
42:1917–1924. 2018.PubMed/NCBI
|
|
15
|
Wu YT, Tan HL, Shui G, Bauvy C, Huang Q,
Wenk MR, Ong CN, Codogno P and Shen HM: Dual role of
3-methyladenine in modulation of autophagy via different temporal
patterns of inhibition on class I and III phosphoinositide
3-kinase. J Biol Chem. 285:10850–10861. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marone R, Cmiljanovic V, Giese B and
Wymann MP: Targeting phosphoinositide 3-kinase: Moving towards
therapy. Biochim Biophys Acta. 1784:159–185. 2008. View Article : Google Scholar
|
|
17
|
Li J, Hou N, Faried A, Tsutsumi S,
Takeuchi T and Kuwano H: Inhibition of autophagy by 3-MA enhances
the effect of 5-FU-induced apoptosis in colon cancer cells. Ann
Surg Oncol. 16:761–771. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu D, Yang Y, Liu Q and Wang J:
Inhibition of autophagy by 3-MA potentiates cisplatin-induced
apoptosis in esophageal squamous cell carcinoma cells. Med Oncol.
28:105–111. 2011. View Article : Google Scholar
|
|
19
|
Singh BN, Kumar D, Shankar S and
Srivastava RK: Rottlerin induces autophagy which leads to apoptotic
cell death through inhibition of PI3K/Akt/mTOR pathway in human
pancreatic cancer stem cells. Biochem Pharmacol. 84:1154–1163.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu B, Xu C, Wu X, Liu F, Du Y, Sun J, Tao
J and Dong J: Icariin exerts an antidepressant effect in an
unpredictable chronic mild stress model of depression in rats and
is associated with the regulation of hippocampal neuroinflammation.
Neuroscience. 294:1932052015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang Y, Jacobi A, Vater C, Zou L, Zou X
and Stiehler M: Icariin promotes angiogenic differentiation and
prevents oxidative stress-induced autophagy in endothelial
progenitor cells. Stem Cells. 33:1863–1877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mo ZT, Li WN, Zhai YR and Gong QH: Icariin
attenuates OGD/R-induced autophagy via Bcl-2-dependent cross talk
between apoptosis and autophagy in PC12 cells. Evid Based
Complement Alternat Med. 2016:43430842016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan C, Yang Y, Liu Y, Jiang S, Di S, Hu W,
Ma Z, Li T, Zhu Y, Xin Z, et al: Icariin displays anticancer
activity against human esophageal cancer cells via regulating
endoplasmic reticulum stress-mediated apoptotic signaling. Sci Rep.
6:211452016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu X, Qiao B, Liu Q and Zhang W:
Upregulation of extracellular matrix metalloproteinase inducer
promotes hypoxia-induced epithelial-mesenchymal transition in
esophageal cancer. Mol Med Rep. 12:7419–7424. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harhaji-Trajkovic L, Vilimanovich U,
Kravic-Stevovic T, Bumbasirevic V and Trajkovic V: AMPK-mediated
autophagy inhibits apoptosis in cisplatin-treated tumour cells. J
Cell Mol Med. 13:3644–3654. 2009. View Article : Google Scholar
|
|
26
|
Jiang S, Chang H, Deng S and Fan D:
Icariin inhibits autophagy and promotes apoptosis in SKVCR cells
through mTOR signal pathway. Cell Mol Biol. 64:4–10. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang SS, Chen YH, Chen N, Wang LJ, Chen
DX, Weng HL, Dooley S and Ding HG: Hydrogen sulfide promotes
autophagy of hepatocellular carcinoma cells through the
PI3K/Akt/mTOR signaling pathway. Cell Death Dis. 8:e26882017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo L, Zhao J, Qu Y, Yin R, Gao Q, Ding S,
Zhang Y, Wei J and Xu G: microRNA-20a inhibits autophagic process
by targeting ATG7 and ATG16L1 and favors mycobacterial survival in
macrophage cells. Front Cell Infect Microbiol. 6:1342016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Y, Zhou X, Qiao J and Bao A:
MiR-142-3p overexpression increases chemo-sensitivity of NSCLC by
inhibiting HMGB1-mediated autophagy. Cell Physiol Biochem.
41:1370–1382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang Z, Huo L, Li K, Wu Y and Hu Z:
Blocked autophagy by miR-101 enhances osteosarcoma cell
chemosensitivity in vitro. Scientific World Journal.
2014:7947562014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim C, Kim W, Lee H, Ji E, Choe YJ,
Martindale JL, Akamatsu W, Okano H, Kim HS, Nam SW, et al: The
RNA-binding protein HuD regulates autophagosome formation in
pancreatic β cells by promoting autophagy-related gene 5
expression. J Biol Chem. 289:112–121. 2014. View Article : Google Scholar
|
|
32
|
Jiang K, Li Y, Zhu Q, Xu J, Wang Y, Deng
W, Liu Q, Zhang G and Meng S: Pharmacological modulation of
autophagy enhances Newcastle disease virus-mediated oncolysis in
drug-resistant lung cancer cells. BMC Cancer. 14:5512014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Eskelinen EL and Saftig P: Autophagy: A
lysosomal degradation pathway with a central role in health and
disease. Biochim Biophys Acta. 1793:664–673. 2009. View Article : Google Scholar
|
|
34
|
He C and Klionsky DJ: Regulation
mechanisms and signaling pathways of autophagy. Annu Rev Genet.
43:67–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yousefi S, Perozzo R, Schmid I, Ziemiecki
A, Schaffner T, Scapozza L, Brunner T and Simon HU:
Calpain-mediated cleavage of Atg5 switches autophagy to apoptosis.
Nat Cell Biol. 8:1124–1132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McKnight NC and Zhenyu Y: Beclin 1, an
essential component and master regulator of PI3K-III in health and
disease. Curr Pathobiol Rep. 1:231–238. 2013. View Article : Google Scholar
|
|
37
|
Wilkinson DS, Jariwala JS, Anderson E,
Mitra K, Meisenhelder J, Chang JT, Ideker T, Hunter T, Nizet V,
Dillin A, et al: Phosphorylation of LC3 by the Hippo kinases
STK3/STK4 is essential for autophagy. Mol Cell. 57:55–68. 2015.
View Article : Google Scholar :
|
|
38
|
Sheng R, Zhang LS, Han R, Liu XQ, Gao B
and Qin ZH: Autophagy activation is associated with neuroprotection
in a rat model of focal cerebral ischemic preconditioning.
Autophagy. 6:482–494. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akcakanat A, Sahin A, Shaye AN, Velasco MA
and Meric-Bernstam F: Comparison of Akt/mTOR signaling in primary
breast tumors and matched distant metastases. Cancer.
112:2352–2358. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chiang GG and Abraham RT: Phosphorylation
of mammalian target of rapamycin (mTOR) at Ser-2448 is mediated by
p70S6 kinase. J Biol Chem. 280:25485–25490. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chang L, Graham PH, Hao J, Ni J, Bucci J,
Cozzi PJ, Kearsley JH and Li Y: PI3K/Akt/mTOR pathway inhibitors
enhance radiosen-sitivity in radioresistant prostate cancer cells
through inducing apoptosis, reducing autophagy, suppressing NHEJ
and HR repair pathways. Cell Death Dis. 5:e14372014. View Article : Google Scholar
|
|
42
|
Heras-Sandoval D, Pérez-Rojas JM,
Hernández-Damián J and Pedraza-Chaverri J: The role of
PI3K/AKT/mTOR pathway in the modulation of autophagy and the
clearance of protein aggregates in neurodegeneration. Cell Signal.
26:2694–2701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qiu Y, Li C, Wang Q, Zeng X and Ji P:
Tanshinone IIA induces cell death via Beclin-1-dependent autophagy
in oral squamous cell carcinoma SCC-9 cell line. Cancer Med.
7:397–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Saiki S, Sasazawa Y, Imamichi Y, Kawajiri
S, Fujimaki T, Tanida I, Kobayashi H, Sato F, Sato S, Ishikawa K,
et al: Caffeine induces apoptosis by enhancement of autophagy via
PI3K/Akt/mTOR/p70S6K inhibition. Autophagy. 7:176–187. 2011.
View Article : Google Scholar :
|
|
45
|
Nazio F, Strappazzon F, Antonioli M,
Bielli P, Cianfanelli V, Bordi M, Gretzmeier C, Dengjel J,
Piacentini M, Fimia GM, et al: mTOR inhibits autophagy by
controlling ULK1 ubiquitylation, self-association and function
through AMBRA1 and TRAF6. Nat Cell Biol. 15:406–416. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Klionsky DJ, Cuervo AM and Seglen PO:
Methods for monitoring autophagy from yeast to human. Autophagy.
3:181–206. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu N, Kong LS, Chen H, Li WD, Qian AM,
Wang XY, Du XL, Li CL, Yu XB and Li XQ: Autophagy protein 5
enhances the function of rat EPCs and promotes EPCs homing and
thrombus recanalization via activating AKT. Thromb Res.
136:642–651. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Du J, Teng RJ, Guan T, Eis A, Kaul S,
Konduri GG and Shi Y: Role of autophagy in angiogenesis in aortic
endothelial cells. Am J Physiol Cell Physiol. 302:C383–C391. 2012.
View Article : Google Scholar :
|
|
49
|
Eramo A, Ricci-Vitiani L, Zeuner A,
Pallini R, Lotti F, Sette G, Pilozzi E, Larocca LM, Peschle C and
De Maria R: Chemotherapy resistance of glioblastoma stem cells.
Cell Death Differ. 13:1238–1241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang D, Chen J, Chen H, Duan Z, Xu Q, Wei
M, Wang L and Zhong M: Leptin regulates proliferation and apoptosis
of colorectal carcinoma through PI3K/Akt/mTOR signalling pathway. J
Biosci. 37:91–101. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kang S, Dong SM, Kim BR, Park MS, Trink B,
Byun HJ and Rho SB: Thioridazine induces apoptosis by targeting the
PI3K/Akt/mTOR pathway in cervical and endometrial cancer cells.
Apoptosis. 17:989–997. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jiang W and Ogretmen B: Autophagy paradox
and ceramide. Biochim Biophys Acta. 1841:783–792. 2014. View Article : Google Scholar :
|
|
53
|
Lin YC, Lin JF, Wen SI, Yang SC, Tsai TF,
Chen HE, Chou KY and Hwang TI: Inhibition of high basal level of
autophagy induces apoptosis in human bladder cancer cells. J Urol.
195:1126–1135. 2016. View Article : Google Scholar
|
|
54
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Schweichel JU and Merker HJ: The
morphology of various types of cell death in prenatal tissues.
Teratology. 7:253–266. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
He Z, Ma WY, Hashimoto T, Bode AM, Yang CS
and Dong Z: Induction of apoptosis by caffeine is mediated by the
p53, Bax, and caspase 3 pathways. Cancer Res. 63:4396–4401.
2003.PubMed/NCBI
|