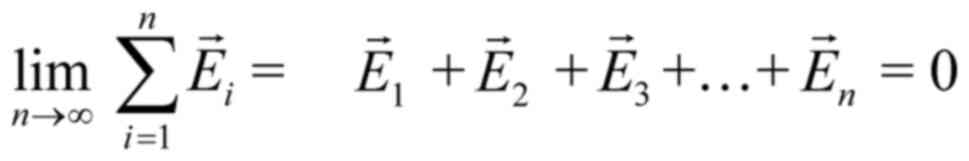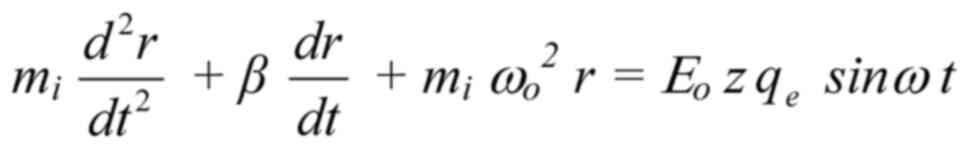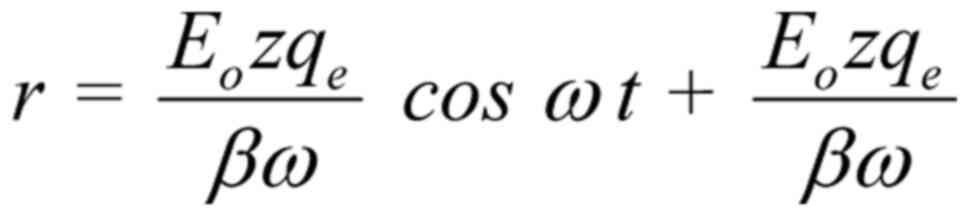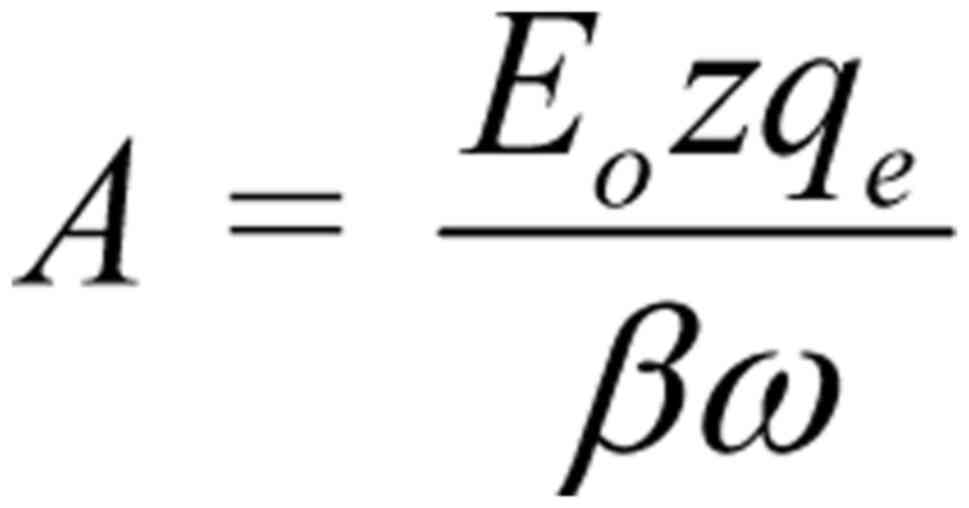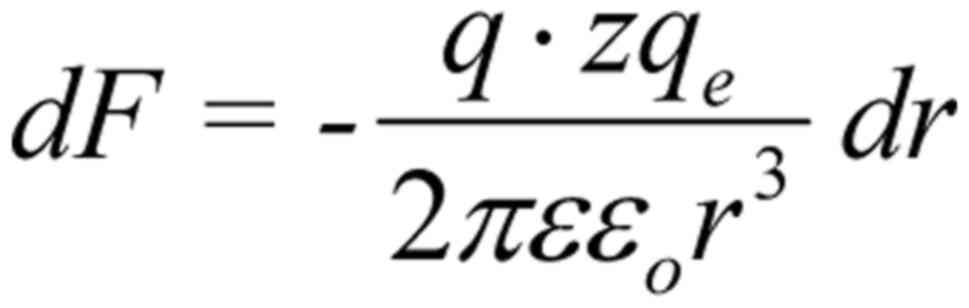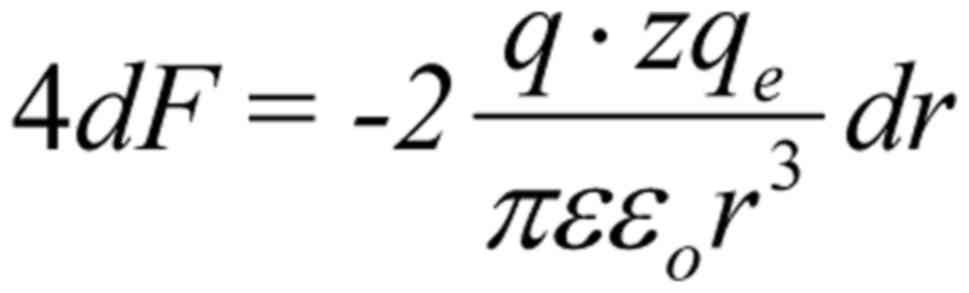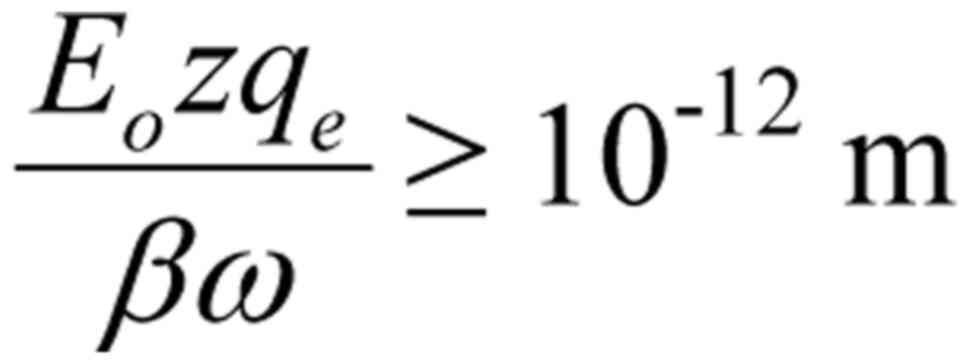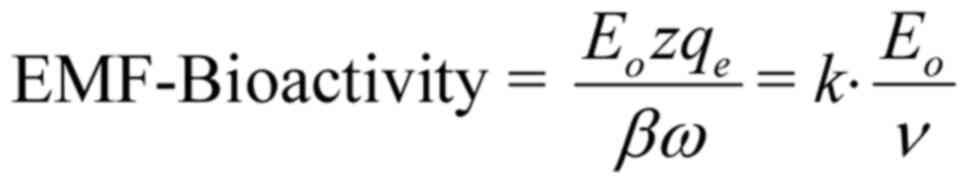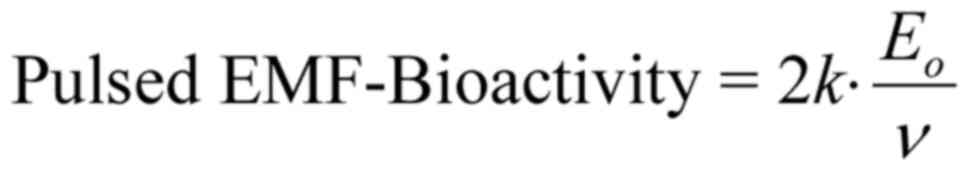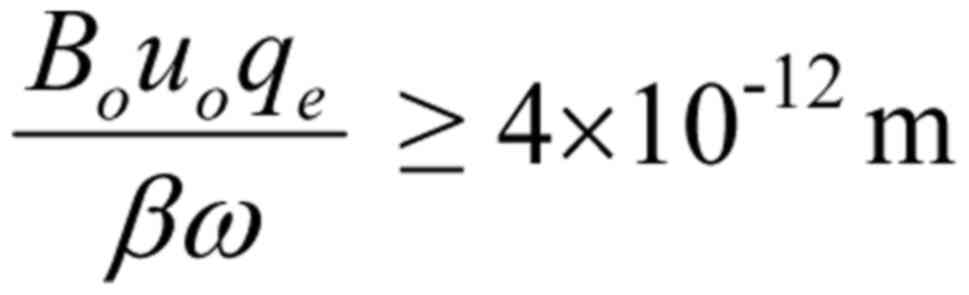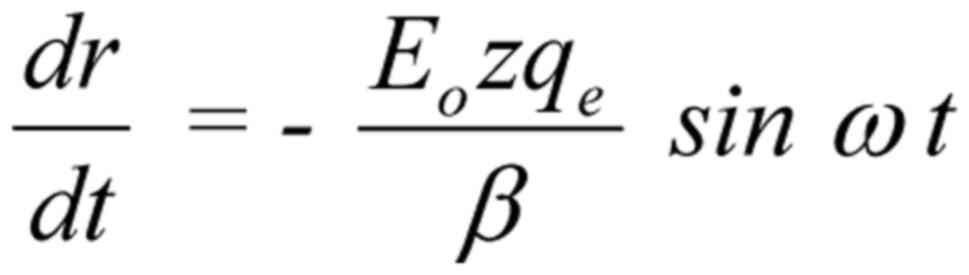1. Introduction
Experimental and epidemiological findings
connecting exposure of living organisms to ELF and complex RF
human-made EMFs with genetic damage, infertility and cancer
There is a plethora of experimental findings
connecting the in vivo or in vitro exposure of
experimental animals or cells to extremely low frequency (ELF)
(3-3000 Hz) or radio-frequency (RF)/microwave (300 kHz-300 GHz)
electromagnetic fields (EMFs), with genetic damage/alterations (DNA
damage, chromosome damage and mutations, among others), cell death
and related effects (1-4). Most findings concern exposure to
wireless communication (WC) EMFs [from mobile phones/antennas,
cordless domestic phones (DECT: digitally enhanced cordless
telecommunications), internet (Wi-Fi: wireless fidelity) or
'Bluetooth' wireless connections, among others], which necessarily
combine RF/microwave carrier frequencies with ELF pulsing and
modulation, and ultra low frequency (ULF) (0-3 Hz) random
variability of the signal. Today, almost all technical RF EMFs (not
only of WC, but also from radars, radio and television antennas,
among others) contain ELF/ULF components in the form of on/off
pulsations, modulation, and signal variability. These are usually
called simply 'RF', but actually they are a combination of RF and
ELF/ULF (4).
The number of experimental-laboratory studies
showing genetic damage and related effects induced by human-made
ELF or RF (combined with ELF) EMFs on a variety of organisms/cell
types under different experimental conditions has rapidly
increased, especially in recent years (5-55).
Several of the aforementioned findings involve DNA
damage and consequent cell death in reproductive cells of different
animals, resulting in decreased reproduction. In particular, the
effects of pulsing WC EMFs on the DNA of reproductive cells, as
reported by different studies on a variety of animals (25,30,31,36,40,41,46), display a marked similarity and
explain other findings that connect WC EMF exposure with insect,
bird and mammalian (including human) infertility (56-64), or declines in bird and insect
populations (especially bees) during the past 15 years (65-69). A significant decrease in
reproduction (decrease in egg laying or embryonic death) after
exposure to mobile telephony (MT) radiation was identically
observed in fruit flies (30,40,57,58), chicken eggs (61), birds (65-67), and bees (63). Similar effects are reported for
amphibians (70,71), rats (31,62), and human sperm (decreased number
and motility of spermatozoa) (59,60). These markedly similar findings in
different organisms by different research groups can be explained
by the observed cell death in reproductive cells after DNA damage,
as seen in fruit fly ovarian cells (30,40,41,46), human sperm cells (36), mouse and rat sperm cells (25,31). Decreased reproduction after DNA
damage and cell death in reproductive cells or embryonic death
induced by purely ELF EMF-exposure is also reported (4,9,14,22,47).
At the same time, epidemiological/statistical
studies increasingly link man-made EMF exposure with health
problems, genetic damage and cancer in human populations. More
specifically, ELF EMFs from power lines and high-voltage
transformers (mainly 50-60 Hz plus additional frequencies due to
harmonics, noise and discharges, among others) are linked with
childhood leukemia (72-82) for magnetic field intensities down
to 2 mG (0.2 µT) (76,82), or distances from power lines up to
600 m (81), and electric field
intensities down to 10 V/m (78).
RF exposure from various antennas always containing ELF components,
especially MT antennas, is linked to various forms of cancer.
Hallberg and Johansson (83)
found a connection between skin cancer (melanoma) incidence in
humans and residential exposure to radio broadcasting antennas,
while two recent studies found significantly increased genetic
damage in the peripheral blood lymphocytes of people residing in
the vicinity of MT base antennas (84,85). During the past 15 years,
epidemiological studies have found an increasing association
between mobile or cordless phone use and brain tumors in humans
(86-98). Moreover, during the past 20 years,
statistical studies have found associations between exposure to MT
base station antennas and devices, and reported symptoms of
un-wellness referred to as 'microwave syndrome' or
'electro-hypersensitivity' (EHS). The symptoms include headaches,
fatigue, sleep disorders, etc. (99-107). A high percentage (~80%) of EHS
self-reporting patients were recently found with increased
oxidative stress (OS) [intracellular increase in free
radicals/reactive oxygen species (ROS)] in their peripheral blood
(108).
A review of studies involving exposure to complex RF
EMFs with ELF pulsation/modulation revealed that 93% of them
reported induction of OS/ROS overproduction in biological systems
(109).
Induction of cancer in experimental animals by
long-term MT exposure, including ELF pulsations, has also been
reported (110,111). A recent study of the USA
National Toxicology Program (NTP) found that rats exposed for 2
years, 9 h per day, in the near-field of simulated 2nd generation
(2G) or 3rd generation (3G) MT emissions, developed brain cancer
(glioma) and heart cancer (malignant schwannoma), with both lower
and higher radiation levels than the officially accepted limits
(112). Moreover the study found
significantly increased DNA damage (strand breaks) in the brains of
exposed animals (113),
confirming that DNA damage is closely related to carcinogenesis. An
Italian life-span exposure study of rats in a simulated 2G MT
far-field also found induction of heart schwannomas and brain glial
tumors, confirming the results of the NTP study (114).
These findings on animal carcinogenicity along with
the epidemiological cancer findings on humans, the DNA damage and
OS findings, and the adverse effects on reproduction due to DNA
damage in the gametes or embryonic death, point towards the same
direction, i.e., that human-made EMF exposure causes OS and DNA
damage that may lead to cancer, reproductive declines and related
diseases. It is important to note that the exposure levels in the
vast majority of all the aforementioned studies (1-114) were significantly below the
officially accepted exposure limits for ELF and RF EMFs, which have
been set to prevent discharges on humans in the case of ELF and
heating of living tissues in the case of RF (115,116).
At the same time, several other studies have
reported no effects of ELF or RF EMFs in all the aforementioned
end-points (1-4,47,57,115-124), especially studies that employed
simulated MT/WC exposure from generators with invariable parameters
(intensity, frequency and pulsations, among others) and no
modulation or random variability. By contrast, more than 95% of the
studies that employed real-life MT/WC exposure from commercially
available devices (mobile/cordless phones and Wi-Fi, among others)
with high signal variability found effects (4,121,122). Regardless of real-life or
simulated exposure, the majority of experimental studies (more than
70%) both in the RF (combined with ELF) and purely ELF bands do
find effects (4,109,123,124). In a recent review of 138 RF
studies with frequencies >6 GHz evaluating potential effects of
the under deployment 5th generation (5G) MT/WC system, it was not
specifically examined whether there were ELF components in the
exposure and what type, or whether there was any similarity between
the signals produced by generators in the studies, and those of the
5G, apart from the carrier frequency. While most of the reviewed
studies reported effects, they were criticised in this review for
not being 'independently replicated' and for employing 'low quality
methods of exposure assessment and control' (125). Thus, despite the incomplete
review methodology, the authors of the review attempted to
downgrade any reported effects.
Under the increasing weight of scientific evidence,
the International Agency for Research on Cancer (IARC) has for a
long time now classified both ELF and RF EMFs as possibly
carcinogenic to humans (group 2B) (117-119). Based on additional scientific
evidence after the 2011 IARC classification for RF EMFs, several
studies have suggested that RF/WC EMFs should be re-evaluated and
classified as probably carcinogenic (group 2A) or carcinogenic
(group 1) to humans (92,97,126,127). As already emphasized, in the
vast majority of studies characterized as 'RF', the ELF/ULF
components were present.
While the reported effects in the vast majority of
the above studies (1-124) induced by ELF or complex RF
(containing ELF) EMFs were not accompanied by any significant
heating of the exposed living tissues, it is well established that
purely RF/microwave EMFs cause heating of exposed materials (e.g.
microwave ovens). The heating becomes significant for high
power/intensity (≥0.1 mW/cm2) and high frequency (at GHz
range) microwaves (128). In
addition, purely RF EMFs, which are of very limited technological
use, are scarcely reported to induce non-thermal effects, and it is
questionable in such cases, whether the presence of any ELFs was
carefully excluded (129).
DNA damage and related pathologies
It is well documented that DNA damage is connected
with cell senescence (cell aging and loss of replicative capacity),
cell death, neurodegenerative diseases and aging of an organism,
and is the main cause of carcinogenesis induced by environmental
stressors (3,130-138). DNA damaging events take place at
any time in the cells of any living organism due to a variety of
events (such as exposure to ultraviolet radiation, natural
radioactivity or cytotoxic chemicals), but efficient DNA repair
mechanisms have evolved to provide protection. Damage in the DNA is
any modification in a nucleotide base, deoxyribose, a break in a
covalent bond between deoxyribose and nucleotide base, or a break
in a phosphodiester bond in one or both strands (3,130-139).
Replication of damaged (or inaccurately repaired)
DNA that may occur before repair or blocking can lead to gene
mutations, which will then give rise to altered proteins. Mutations
in oncogenes, tumor-suppressor genes, DNA repair genes or genes
that control the cell cycle can generate a clonal cell population
with a distinct ability to proliferate. DNA methylation that may
prohibit the expression of DNA repair genes and synthesis of
related proteins can result in inaccurate ('error-prone') DNA
repair. Many such events, which may accumulate over a long period
of time in cases of chronic exposure to carcinogens, can lead to
genomic instability and cancer (133,134,136,139).
When the genomic DNA of a cell is damaged by an
external stressor and the damage is either not reparable or
inaccurately repaired, the following outcomes are possible: i) The
cell dies (necrosis) or is led to suicide (induced apoptosis). In
the case of cell types with the ability to proliferate, the
organism compensates for their loss by creating new cells,
practically with no adverse consequences apart from energy
consumption, which may lead to accelerated aging when such events
occur at a high rate. In the case of cell types that do not have
ability to proliferate, such as neural cells or chondrocytes, the
loss of a significant number of cells will probably result in the
inability of certain tissues/organs to operate normally. In the
case of neural cells, this may lead to neurodegenerative diseases
such as Alzheimer and Parkinson, and autoimmune disorders, among
others. ii) The cell does not die but survives with modified DNA.
In the case of somatic cells that proliferate, the modified genome
will reproduce itself. Even though the organism may recognize such
mutant cells as foreign and try to isolate them and remove them,
they strive to survive and may start proliferating uncontrollably,
initiating cancer. In the case of reproductive cells (oocytes and
spermatocytes), this may lead to mutated new organisms that may be
problematic in many ways or cancer-prone. In both cases (somatic or
reproductive cells) cell senescence is an alternative pathway for
eliminating surviving genetically defective cells. Thus, cells with
irreparably damaged genomic DNA will result in cell senescence,
cell death, cancer or mutated offspring, depending on cell type and
specific biological/environmental conditions (3,4,122,130-132,135-137).
The duration of cancer development (latency period)
after irreparable DNA damage may be a number of years, depending on
the organism and the type of cancer. The latency period for gliomas
(a type of brain cancer) is usually >20 years in humans
(140). This probably explains
why only during the past ~15 years epidemiological studies have
started showing an association between mobile phone use and cancer
(86), whereas cancer from power
lines, which are several decades older than MT/WC, has been
indicated long before (72).
Purpose of the present study
As aforementioned, a growing number of experimental
and epidemiological/statistical findings connect man-made EMF
exposure with genetic damage and cancer, and this involves the
breakage of chemical/electronic bonds in molecules/atoms, in other
words ionization. The human-made EMFs with frequencies up to the
lower limit of infrared (0-3×1011 Hz) discussed in the
present study cannot directly cause ionization, except for very
strong field intensities (≥106 V/m) (141,142). Such field intensities rarely
exist environmentally, apart from atmospheric discharges
(lightning) or in very close proximity to high-voltage power lines
and transformers. The question therefore is how human-made EMFs at
environmental intensities are capable of damaging DNA and other
biological molecules. Obviously they have the ability of breaking
chemical bonds indirectly through the action of some primary
biophysical mechanism(s) and subsequent initiation of intracellular
biochemical processes.
Visible and infrared natural light cannot break
chemical bonds, even though they expose us at higher frequencies
and radiation intensities than human-made EMFs in daily life
(143). There must be a unique
property of the human-made EMFs that makes them capable of inducing
adverse biological/health effects and ionization, in contrast to
natural infrared and visible light. This unique property is that
human-made EMFs/radiation are totally polarized and coherent,
meaning that they possess net electric and magnetic fields, apart
from radiation intensity, which exert forces on any electrically
charged (or polar) particle/molecule such as mobile/dissolved ions
and charged macromolecules in any biological system (143).
The purpose of the present study is to suggest a
realistic primary biophysical mechanism for polarized and coherent
EMFs at environmentally relevant intensities, to impair cellular
function and initiate plausible intracellular biochemical processes
resulting in genetic damage and carcinogenesis, as reported in the
aforementioned studies.
2. Biophysical action of polarized/coherent
EMFs resulting in voltage-gated ion channel (VGIC) dysfunction and
disruption of cell electrochemical balance
It has been shown that polarized/coherent EMFs, even
at very low field intensities in the ULF and ELF bands, can cause
irregular gating of electro-sensitive ion channels or VGICs on the
cell membranes through the 'ion forced-oscillation mechanism'
(143-146), with consequent disruption of the
cell's electrochemical balance (the electrical and osmotic
equilibrium maintained by specific concentrations of all
dissolved/mobile ions across all cell membranes according to the
Nernst equation) (144,147,148). Since, as explained, ELF/ULF
components exist also in the complex WC/RF EMFs, this mechanism,
which will be thoroughly reviewed next, accounts for the biological
effects of the vast majority of human-made (polarized and coherent)
EMFs.
The mechanism is based on molecular/physical data,
and the forces on mobile ions, in the vicinity of the
voltage-sensors of VGICs, exerted by an applied polarized
oscillating EMF. The oscillating field will force mobile ions to
oscillate on parallel planes and in phase with the field. This
coordinated motion of electrically charged particles exerts
electric forces on the voltage-sensors, similar to the forces
exerted on them by changes in the transmembrane electric field
known to physiologically gate these channels, and thus the channels
are gated irregularly by the applied EMF. The forces are
proportional to the amplitude of the forced-oscillation, and thus,
the amplitude is a direct measure of the bioactivity of the applied
EMF. It has been shown that the amplitude (bioactivity) is
proportional to EMF intensity, inversely proportional to EMF
frequency and doubles for pulsed EMFs. The validity of the proposed
mechanism has been verified by numerical testing, while other
previously suggested mechanisms have failed to pass the same test
(149,150). Repeated irregular gating of
electro-sensitive ion channels disrupts cellular electrochemical
balance and homeostasis (147,148), leading to overproduction of
ROS/free radicals as described next.
It is known from a plethora of experimental data
that the most bioactive EMFs are the lower frequency ones
(ELF/ULF). In numerous cases of induced biological effects by
complex RF EMFs modulated by ELFs, it has been found that the
modulation (ELF) and not the carrier (RF) is responsible for the
recorded effects. In addition, it has been repeatedly found that
pulsing RF EMFs with ELF pulse-repetition rates are more active
biologically than continuous (non-pulsed) fields of identical other
parameters (1-5,44,45,47,151-159). These findings are in direct
agreement with the described mechanism.
Biological molecules of critical importance such as
ions, water molecules, proteins, nucleic acids and lipids, among
others, are either polar or carry a net electric charge (147,148). The net electric field from an
infinite number of individual electric pulses of random
polarization and/or random phase (as e.g. photons of natural light)
tends to zero at any moment (and similarly the net magnetic
field).
Thus, non-polarised/incoherent EMFs (as e.g. light
and cosmic microwaves) at any radiation intensity cannot cause any
parallel/coherent oscillation of charged/polar molecules (143). On the contrary, polarized and
coherent (human-made) oscillating EMFs force all charged/polar
molecules in biological tissue to oscillate on planes parallel to
their polarization and in phase with them. This is crucially
important for understanding the mechanism described. The
forced-oscillation will be most intense on the mobile ions, the
smallest charged particles dissolved in large concentrations in the
cytosolic and extracellular aqueous solutions in all living
cells/tissues controlling practically all cellular/biological
functions (147,148).
Even though all molecules move randomly with much
greater velocities/displacements due to thermal energy, this has no
biological effect other than increasing tissue temperature. By
contrast, a polarized and coherent oscillation of much lower energy
than average thermal molecular energy can initiate biological
effects (143-145).
The majority of cation channels (Ca2+,
K+, Na+ and H+, among others) on
the membranes of all animal cells are voltage-gated (147,148). These ion channels convert
between open and closed states when the electrostatic force on
their voltage sensors, due to transmembrane voltage changes,
exceeds some critical value. The voltage sensors are four
symmetrically arranged, transmembrane, positively charged
α-helices, each one named S4. The S4 helices occupy the 4 th
position in a group of 6 parallel α-helices (S1-S6). The channel
consists of four identical such groups in symmetrical positions
around the pore of the channel. The S5-S6 helices of the four
groups form the pore walls (147,148). More specifically, the sensors
are positive Lys and Arg amino acids in the S4 helices. Changes in
the transmembrane voltage of the order of ~30 mV are normally
required to gate electrosensitive channels (change their status
from opened to closed and vice-versa) (160,161). Among the S1-S4 α-helices, the S4
helices are the closest to the pore-forming S5-S6 helices, being
<1 nm in distance from the pore (162,163). Several ions may interact
simultaneously at any instant with an S4 sensor from a distance of
the order of 1 nm, as, except for the ion(s) that may be passing
through the pore any moment or are just outside the gate ready to
pass, a few more ions are bound close to the pore at specific
ion-binding sites (e.g. three in potassium channels) (164,165). Proton voltage-gated channels
studied more recently also contain S4 transmembrane helices with
charged Arg residues as voltage-sensors, similar to the metallic
cation channels (166,167).
Let us consider four identical mobile ions at
distances of the order of 1 nm from the channel-sensors (S4) and an
externally applied oscillating EMF. The average electric (and
magnetic) force on each ion due to any non-polarized EMF is zero
(Eq. 1). By contrast, the force
due to a polarized field with an electrical component E, is
F=Ezqe, (with zqe the
electric charge of the ion).
In the most usual and simplest case of a sinusoidal
alternating electric field, E=Eo sinωt,
the motion (forcedoscillation) equation of a mobile ion is as
follows (143-146):
where mi is the mass of the ion, r is the
displacement of the ion due to the forced-oscillation, z is
the valence of the ion (z=1 for K+,
Na+ or z=2 for Ca2+ ions),
qe =1.6×10−19C is the elementary
charge, β is the damping coefficient (being within channels
kg/s, with Em (~107 V/m) the
transmembrane electric field, and uo=0.25 m/s the
velocity of the ion through an open channel calculated from
patch-clamp measurements of channel ion-currents).
ωo=2πνo (νo
the ion's oscillation self-frequency accepted to be equal to the
recorded spontaneous intracellular ionic oscillation frequencies on
the order of 0.1 Hz), ω=2πν (ν the frequency
of the applied field) and Eo is the intensity
amplitude of the applied oscillating field. Detailed calculations
of the parameters are provided in Panagopoulos et al 2000
(
144).
The right part of Eq.
2 is the force on the ion due to the applied E-field. The first
term of the left part is the resultant force on the ion, the
second term is a damping force and the third term
(mi ωo2 r) a
restoration force exerted by the medium (144,145). While an oscillating ion close to
the S4 sensors exerts gating forces on them, it receives zero
opposite force, as the S4 charges are paired with opposite charges
from adjacent helices of the channel (148). Eq. 2 is a second-order linear
differential equation with constant coefficients, which is solvable
once we know the values of the different parameters.
The general solution of Equation 2 (144) is:
The constant term in the solution represents a
constant displacement of the ion and has no effect on the
oscillating term . This constant displacement represents
a jump of the whole oscillation at a distance equal to the
amplitude, in other words it doubles the amplitude of the
oscillation at the moment when the field is applied or interrupted.
For pulsed fields (such as the vast majority of human-made complex
RF/microwave EMFs, especially those employed in modern WC), this
interruption/repetition occurs constantly with every repeated
pulse. Therefore, pulsed fields are predicted to be twice as
bioactive as continuous/non-pulsed fields of the same other
parameters, and this explains a plethora of experimental findings
showing increased bioactivity of pulsed compared with non-pulsed RF
EMFs, which were previously unexplained (44,45,154, 155,157-159).
Ignoring the constant term in Eq. 3, the amplitude of the
forced-oscillation is:
An oscillating ion of charge zqe
(whose motion is described by Eq.
3) close to the S4 helices of a voltage-gated channel exerts a
force F on the effective charge q of each S4, as
described by Coulomb's law: , (r here is the distance of the
oscillating ion from the S4). The ion displaced by dr during
its oscillation, induces an additional force dF on each S4
sensor:
While in the case of a random/chaotic movement of
the ion due to e.g. thermal motion , and
, in the case of a coordinated
polarized and coherent forcedoscillation, the sum force on each S4
from all four ions, is:
The effective charge of each S4 domain is found to
be: q=1.7qe (161). The force on this charge exerted
by a change of 30 mV in the transmembrane voltage required normally
to gate the channel, is calculated to be (144): dF=8.16×10−13
N.
The displacement of one single-valence ion within
the channel corresponding to this minimum force, according to
Eq. 5 (for z=1, ε
≅ 4, and r ~1 nm), is: dr=4×10−12 m.
The dielectric constant within proteins is
significantly lower than in the aqueous solutions (4/80), and ion
concentration in cells is of the order of 1 ion per nm3
(144,147,148).
For 4 single-valence ions oscillating on parallel
planes and in phase with an applied polarized (and coherent)
oscillating field, the minimum displacement is (according to
Eq. 6) reduced to:
dr=10−12 m. The corresponding necessary
displacement for ions outside the channel would be about 20-fold
higher due to the higher dielectric constant of the aqueous
solutions.
Thus, a crucial finding has been reached: Any
external polarized and coherent oscillating EMF (like all
technical/human-made EMFs) able to force mobile ions to oscillate
with amplitude
is able to irregularly gate VGICs on cell membranes.
For z=1 (e.g. K+ ions), and
replacing qe, β by their values in
Condition 7, we get:
(ν in Hz, Eo in V/m)
For double-valence cations (z=2) (e.g.
Ca2+) the condition becomes:
(ν in Hz, Eo in V/m)
For pulsed fields (such as all MT/WC fields) the
right part of Condition 9 is further divided by 2, becoming:
(ν in Hz, Eo in V/m)
It is clear that the amplitude of the
forced-oscillation given by Eq.
4 is the critical parameter to determine the ability of a
polarized/coherent EMF to induce biological/health effects. We
shall name it 'Bioactivity of the EMF' or 'EMF-Bioactivity'.
Thus:
where is a constant quantity (depending upon
the membrane electric field Em and the velocity
of the ion through an open channel uo),
Eo is the intensity amplitude and ν is the
frequency of the applied electric field. We shall name k the
'bioactivity constant'.
Thus, a most reasonable and elegant result is
reached, that the bioactivity of a polarized oscillating EMF is
proportional to its maximum intensity (Eo) and
inversely proportional to its frequency (ν), meaning that
lower frequency fields are predicted to be more bioactive than
higher frequency ones of the same intensity and waveform. Although
this result was obtained considering the most usual/simple case of
harmonically oscillating polarized EMFs, it is evident that
non-harmonically oscillating polarized fields can also be
approximately described in terms of their bioactivity by Eq. 11.
For pulsed EMFs with harmonically oscillating
carriers, the amplitude doubles and so does the bioactivity:
The same mechanism explains the biological action
of polarized oscillating magnetic fields as well, if we replace in
Eq. 2 the electric force
FE=Ezqe, by a magnetic
force:
exerted on an ion with charge zqe, moving with
velocity u, vertically to the direction of a magnetic field
of intensity B (in which case the magnetic force is
maximum). In the simplest (and most usual) case of an alternating
magnetic field B=Bosinωt with intensity
amplitude Bo and based on the same reasoning as
aforementioned, corresponding bioactivity conditions are obtained
for an oscillating magnetic field.
For one single-valence ion moving through an open
channel vertically to the direction of the applied magnetic field
with u=uo=0.25 m/s (the velocity
calculated for ions moving through an open channel) (144) and for the case of a continuous
oscillating magnetic field, the corresponding bioactivity condition
is:
from which is obtained:
(ν in Hz, Bo in T), or
(ν in Hz, Bo in ¼T), or
For double-valence ions the right part of Condition
16 is divided by 2:
(ν in Hz, Bo in µT)
For double-valence ions and pulsing magnetic field
the right part of Condition 17 is further divided by 2, and the
bioactivity condition becomes:
(ν in Hz, Bo in µT)
It should be noted that apart from the drift
velocity of the ion through the channel (uo=0.25
m/s) that is accepted as initial velocity, the ion will acquire an
additional velocity dr/dt due to the forced-oscillation.
From Eq. 3, the following is
obtained:
(or respectively: for a
sinusoidal magnetic field)
The corresponding magnetic force due to this
additional velocity, Bzqe(dr/dt), is
negligible (more than 108 times smaller) compared with
the damping force β(dr/dt), and thus, it is not taken
into account in Eq. 2.
The maximum of this additional velocity is
independent of the frequency of the field (ω), and is much
smaller for usual field intensities than the ion velocity through
an open channel (uo=0.25 m/s), which in turn is
more than 103 times smaller than its corresponding
average thermal velocity ukT (168). Thus, the described ion
forced-oscillation does not add to tissue temperature and this
mechanism is 'non-thermal', in contrast to the known heating
ability of the high intensity microwaves (128). The non-thermal nature of
human-made EMF-bioeffects, including those of low power
modulated/pulsing RF/microwaves, in contrast to high power
microwaves, has also been discussed in previous studies (169,170).
This theory allows certain predictions for the
bioactivity of some human-made EMFs widely present in the modern
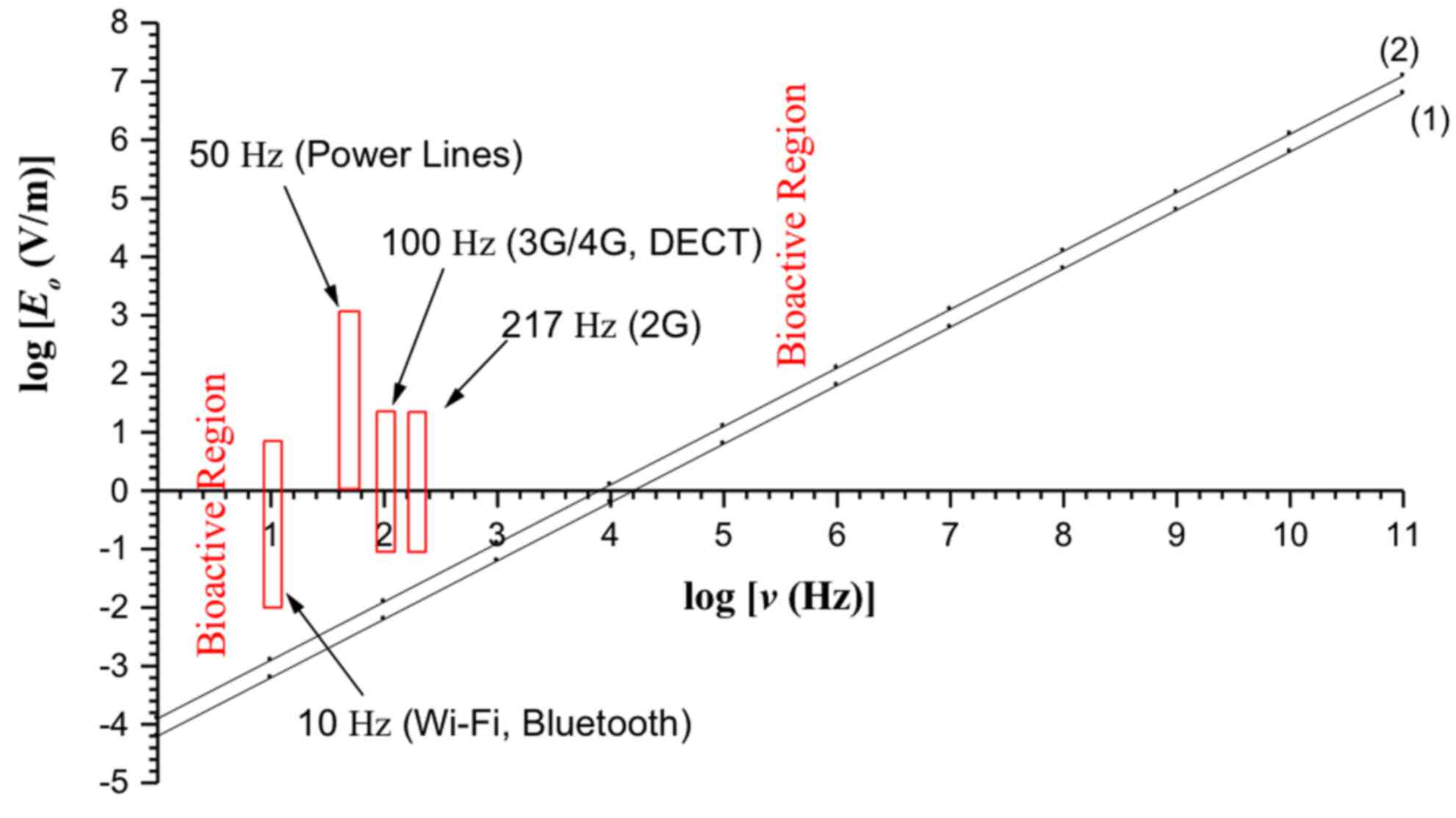
environment: For the sinusoidal alternating (continuous) 50-Hz
E and B fields of high-voltage power lines with
intensities of the order of E ~10 kV/m and B ~0.1-1 G
(or ~10-100 µT) at close distances (10-20 m) from such lines
the conditions 9 and 17 for double valence cations (e.g.
Ca2+) give: Eo≥6×10−3 V/m
or Eo≥6 mV/m (which is satisfied by more than
106 times), and Bo ≥105
µT, which is not satisfied, showing that the recorded
effects from high-voltage power lines are due to the electric
rather than the magnetic component of the resultant EMF, in
contrast to what is usually considered. Thus, the electric
component of power line EMFs is certainly capable of inducing
biological effects in living organisms according to the mechanism
presented, even for intensities down to 1-10 V/m, which exist in
most homes and work places.
For the pulsing ELF E and B fields of
MT/WC EMFs with a pulsing repetition frequency of ~100 Hz (3G/4G
MT, DECT), E ~10 V/m and B ~1 mG (or ~0.1 µT)
(30,40,54,55), the bioactivity conditions 10 and
18 respectively give: Eo≥6×10−3 V/m or
Eo≥6 mV/m, which is satisfied by more than
103 times, and Bo
≥105µT, which is not satisfied for direct action,
but it may be satisfied by the magnetically induced electric field,
which is significant in this case due to the short rise/fall times
of the pulses (143). Similar
results are obtained for the 217-Hz pulsing E/B fields of 2G MT
(30,40).
For Wi-Fi and Bluetooth wireless connections with a
pulsing frequency of ~10 Hz, E ~1 V/m and B ~0.1 mG
(or ~0.01 µT) (171), the
bioactivity conditions 10 and 18 respectively give:
Eo ≥0.6×10−3 V/m or
Eo≥0.6 mV/m, which is satisfied by more than
103 times, and Bo≥104
µT, which is not satisfied for direct action.
The aforementioned numerical examples show that it
is the electric field that seems to be the bioactive component of
an EMF and not the magnetic field, in contrast to what has been
considered before by health agencies (117). The magnetically induced electric
field can also be bioactive in the case of ELF pulses of WC signals
with short rise/fall times (143).
The bioactivity conditions 9 and 10 for continuous
and pulsed electric fields respectively are depicted in Fig. 1. The region above line 1
(including the line) represents the bioactive combinations of
intensity amplitude (Eo) and frequency (ν)
for pulsed fields, and above line 2 (including the line) for
continuous fields. The ELF electric field of power lines, 2G/3G/4G
MT, DECT, WiFi and 'Bluetooth', lie within the bioactive region
predicted by the presented theory.
3. Biochemical processes activated by
irregular gating of VGICs, leading to DNA damage
Irregular gating of ion channels and
ROS
Irregular gating of VGICs by oscillating polarized
and coherent ELF EMFs as described [and originally in (143-146)] has been verified experimentally
for calcium (Ca2+), potassium (K+) and sodium
(Na+) VGICs (172-174). This can alter intracellular
ionic concentrations, disrupting the electrochemical balance of the
cell and leading to DNA damage by OS/ROS overproduction (175-179).
Most ROS are free radicals. Free radicals are
highly unstable molecules containing an unpaired electron, which is
denoted by a dot (•), and have a tremendous tendency to
chemically react with surrounding molecules and/or with each other
in order to couple the unpaired electron and become stable. This is
the reason why they have extremely short lifetimes. Most ROS react
rapidly with surrounding biomolecules inducing chemical alterations
(180). Overproduction of ROS in
living cells due to EMF exposure has been reliably documented, with
two important ROS found after EMF exposure being superoxide anion
(O2•−) and nitric oxide (NO•)
(109). These may result in
hydroxyl radical (OH•) and peroxynitrite
(ONOO−) correspondingly, both of which ROS are very
reactive with biological molecules and specifically DNA, as
discussed next. ONOO− may interact directly with DNA,
as, similarly with NO•, it can be diffused everywhere in
the cell (181). Superoxide
anion radical (O2•−) is catalyzed by
superoxide dismutase enzymes in the cytosol or the mitochondria and
is converted to hydrogen peroxide (H2O2)
(109,182):
H2O2 is a critical molecule
in oxidative damage since it can move to any intracellular site
(including the nucleus), where it can be converted to the most
potent OH•, which can damage any biological molecule,
including DNA (183-187).
DNA damage by ROS leading to mutations and disease
has been well studied (188,189). Pall (190), in a review of EMF-bioeffects
studies with calcium channel blockers, noted a connection between
voltage-gated calcium channels (VGCCs) and
NO•/ONOO− overproduction. This verified
earlier observations of EMF-induced effects on intracellular
calcium concentrations, and the unique role of VGCCs (1,151-153,191,192).
It is known that the intracellular redox status can
activate Ca2+, Na+ and K+ channels
in order to reinstate homeostasis (178), and inversely, activation of
these channels determines the redox status and the electrochemical
balance of the cell (179).
Multiple studies have found connections between the impaired
function of calcium, potassium, sodium and chloride channels with
the induction of OS and related pathologies (175-177). These studies provide additional
evidence for the validity of the presented biophysical mechanism
(143-146).
Calcium signaling and mitochondrial ROS
production
Alteration of intracellular ionic concentrations
will affect key cellular signaling pathways, including the
Ca2+ signaling system, which regulates a variety of
cellular functions including cell proliferation, differentiation,
the ROS regulatory system and apoptosis (192-196). Impaired function of VGCCs in the
plasma or in the mitochondrial membranes leading to critical
changes in cytosolic or mitochondrial concentrations of
Ca2+ ions, such as those following EMF exposure, is
connected with pathogenesis and cytotoxicity (195,196).
Voltage-gated anion channels in the outer membrane
of the mitochondria regulate Ca2+ entry into the
inter-membrane space and in the matrix, which is crucial for
mitochondrial ROS production. Increased level of Ca2+
stimulates O2•− production by the electron
transport chain in the mitochondria and/or activation of nitric
oxide synthase (NOS), to generate more NO•.
NO• inhibits complex IV of the electron transport chain,
triggering production of even more ROS (109,193). ROS overproduction in the
mitochondria can damage DNA both in the mitochondria and the
nucleus, and initiate a signaling cascade leading to apoptosis, as
found in human spermatozoa after MT EMF exposure (36). Moreover, increased concentrations
of NO• in living cells due to activation of NOS at
different locations of the cell may lead to formation of
ONOO− (181,182).
Regulation of apoptosis is crucial for anticancer
control (197). However,
excessive apoptosis, induced by increased ROS levels, is connected
with inflammatory diseases and cancer (198). When overproduction of ROS in a
cell overloads the capacity of the antioxidant system of the cell,
the cell/organism is under OS. This condition may lead to
significant DNA damage with consequent genomic instability and
carcinogenesis (182,183,194-198).
K+ channels have also been shown to be
involved in the activation of apoptosis (194), and voltage-gated Ca2+
and K+ channels have been shown to be connected with
cell proliferation and carcinogenesis (199). Thus, cytosolic concentrations of
Ca2+ and K+ ions play major roles in cellular
function and metabolism. In addition, voltage-gated calcium and
potassium channels play important roles in iron entry into the
cells. Iron catalyzes the production of OH• via the
Fenton reaction and thus, impaired function of these channels can
promote cellular toxicity (200-202).
NADPH oxidase and ROS production
Apart from the effect of EMFs on metallic cation
voltage-gated channels (such as Ca2+, Na+ and
K+), proton (H+) voltage-gated channels will
be affected as well, as they operate in a very similar way
(166,167). This in turn would affect the
function of NADPH oxidase, a plasma membrane enzyme found in
abundance in all cells, which normally generates ROS for the
elimination of invading microorganisms (203,204). The activity of NADPH oxidase is
strongly associated with H+ channels and it may even act
directly as a H+ voltage-gated channel due to its
gp91phox transmembrane subunit (205,206). NADPH oxidase generates an
electron flux for the reduction of extracellular O2 to
O2•− (203,207).
NADPH oxidase is activated by cytosolic
Ca2+ and possesses a Ca2+-binding site in
addition to its H+ voltage-gated channel
(gp91phox transmembrane region) (204). Thus, perturbation of
intracellular concentrations of either H+ or
Ca2+, after irregular gating of their voltage-gated
channels, will affect the function of NADPH oxidase and trigger
irregular ROS production.
NADPH oxidase has been reasonably suggested as a
primary target of EMF exposure in living cells. In 2007, Friedman
et al (208) found rapid
ROS production in cultured cells after a few min of exposure to RF
EMF emitted by a generator.
Na+/K+-ATPase and
ROS production
Impaired function of Na+, K+,
Mg2+ and Ca2+ voltage-gated channels may also
affect the function of the Na+/K+ pump
(ATPase) and Ca2+ pumps in the plasma membranes of all
cells. The ion pumps (active ion transporters) across all cell
membranes in coordination with the ion channels (passive ion
transporters) determine the membrane voltage, the volume of the
cell and the electrochemical balance (147,148). A positive-feedback amplification
loop between Na+/K+-ATPase signaling and ROS
production by the mitochondria was experimentally demonstrated in
primary cultures of cardiac myocytes (209).
Na+/K+-ATPase became a target for
ROS-initiated signaling, and in turn, stimulation of
Na+/K+-ATPase signaling function led to
increased ROS production. This model can definitely be associated
with dysfunction in living cells under EMF-exposure.
Therefore, it is clearly indicated that irregular
gating of VGICs on plasma and intracellular membranes due to
EMF-exposure will most likely trigger ROS overproduction and
consequent cellular damage. Although plenty of data connecting ion
channel dysfunction and the induction of cell death or cancer have
been available for a long time (194,199), the connection between the
dysfunction of VGICs and ROS overproduction (175-179,190-192) leading to DNA damage has not
perhaps gained the attention it deserves.
Apart from action via ROS/free radicals, DNA damage
may be brought about by irregular activation of DNases after
alteration of intracellular ionic concentrations. Of the two forms
of endonucleases implicated in the initiation of apoptosis, one of
them is Ca2+-dependent (DNase I). An increased level of
intracellular Ca2+ in some cases is associated with
increased apoptosis, possibly due to the activation of DNase I
(210). Thus, the possible
activation of DNase I by increased levels of intracellular
Ca2+ may be an alternative way for DNA damage and
related pathologies.
ROS and DNA damage
OH• is considered the most potent
oxidant of DNA. The main mechanism for OH• production
involves the iron-catalyzed conversion of
H2O2 via the Fenton reaction (211): Fe2+ is oxidized by
H2O2 to Fe3+, producing an
OH• radical and a hydroxide ion (OH−)
(Eq. 21). Fe3+ is
then reduced back to Fe2+ by another molecule of
H2O2, producing a hydroperoxyl radical and a
proton (Eq. 22).
The net effect is the conversion of two hydrogen
peroxide molecules to produce two different oxygen-radical species,
with water (H+ + OH−) as a byproduct.
The OH• radical reacts with any
biological molecule in its immediate environment, including DNA.
For example, it can break macromolecules (R-R or R-H) or abstract
atoms from them (such as the various hydrogen atoms of the
deoxyribose) by breakage of covalent bonds. This results in
chemical alterations of the macromolecules and production of new
free radicals (R• or RO•):
The new free radicals will further react with other
molecules resulting in additional chemical alterations.
Corresponding evidence for DNA damage by ONOO− is
available as well (181).
In conclusion, there is a clear sequence of events
starting from the irregular gating of VGICs by EMFs up to DNA
damage and related pathologies, including carcinogenesis.
4. Discussion
The present study reviewed experimental and
epidemiological findings connecting exposure to purely ELF, and RF
(containing ELF) human-made EMFs, with DNA damage and related
pathologies, including cancer. It is documented that both such
types of human-made EMF-exposure can induce OS (3,34,36-39,43,45,109), DNA damage (1-55,84,85) and infertility (56-71). It is also documented that the same
types of EMF-exposure are linked with increased cancer risk both in
humans and experimental animals (72-83,86-98,110-114).
We attempted to provide a complete, plausible
explanation of these DNA damage-related findings on a biophysical
and biochemical basis. According to the ion forced-oscillation
mechanism for dysfunction of VGICs (143-146), human-made (polarized and
coherent) ELF/ULF EMFs or the ELF/ULF
modulation/pulsing/variability components of modern RF/WC EMFs can
alter intracellular ionic concentrations by irregular gating of
VGICs on cell membranes. This leads to immediate OS by ROS
(over)production in the cytosol and/or the mitochondria, which can
damage DNA when cells are unable to reinstate electrochemical
balance (normal intracellular ionic concentrations). Consequently,
DNA damage can lead to reproductive disabilities, neurodegenerative
diseases, aging, genetic alterations and cancer.
According to the presented biophysical mechanism,
the bioactivity of a polarized/coherent EMF is proportional to its
intensity, inversely proportional to its frequency and doubles for
pulsed fields, meaning that the ELF/ULF EMFs and even more the
pulsing RF EMFs with ELF pulsations such as all WC EMFs, are
predicted to be the most bioactive. This explains the recorded
effects of purely ELF EMFs (1-5,9,13-18,22,47, 50,72-82,117,212) and those of
modulated/pulsing/variable RF EMFs (1,3,4,6-8,19-21,23-46,48,49,51-55,57-71,84-107, 109-114,118,121-126). As emphasized, all types of RF
exposure from all types of antennas and WC devices (WC EMFs)
necessarily combine RF carrier signals with ELF/ULF components in
the form of pulsing, modulation and random variability. The RF
carrier signal alone does not contain information. The information
is always contained in the ELF signals that modulate the RF
(4). Significant experimental
evidence shows that the bioactive parameters in a complex signal
are its ELF components, and that non-modulated and non-pulsed RF
signals alone do not usually induce biological effects (4,44,45,151-159), apart from heating when they
possess high enough frequency and intensity (128,168-170). Therefore, the present study
suggests that the vast majority of non-thermal effects attributed
till now to various types of RF EMF-exposure, are actually due to
their ELF/ULF components.
The presented biophysical mechanism and the
provided numerical examples show that it is the direct ELF electric
fields (and the magnetically induced electric fields in the case of
sudden pulses), not the magnetic, that are the bioactive
components, in contrast to what has been considered before by
health agencies (117), and in
agreement with previous experimental findings (191). Although electric fields are less
penetrating in living tissue than magnetic fields, penetration
depends upon the inverse square root of frequency, and thus ELF
electric fields are significantly penetrating. Penetration depends
also upon the inverse square root of the medium conductivity
(213). Even though seawater is
much more conductive than living tissue, ELF electromagnetic waves
(thus both the electric and the magnetic parts of the waves) are
penetrating several meters into seawater, accommodating
communications with submarines (214). Moreover, it is known that
isolated tissues respond to externally applied pulsed or sinusoidal
ELF electric fields at very low thresholds (~10−3 V/m)
similar to those predicted by this theory (143,215-217). This evidence shows that ELF
electric fields penetrate enough to induce effects into living
tissue, even at very low field intensities. Finally, skin cells,
nerve terminals, eyes and organs close to the surface, such as the
brain and heart, are directly exposed to externally applied EMFs.
For all these reasons, no distinction is made between externally
applied ELF electric fields and internally induced ones.
The ion forced-oscillation mechanism/theory was
described in the present study by realistic equations based on the
forces exerted on mobile ions in the vicinity of the
voltage-sensors of VGICs on cell membranes by externally applied
human-made (polarized) EMFs. The solution of the basic Eq. 2 resulted in bioactivity conditions
connecting the intensity of an applied polarized EMF with its
frequency. The bioactivity conditions 8-10, and 16-18, provided the
bioactive intensity-frequency combinations for continuous and
pulsed electric and magnetic fields. The final numbers explain
almost all the experimental and epidemiological findings connecting
biological/health effects with human-made EMF-exposure.
Although the mechanism was first published in 2000
(144) based on the available
data on the structure and function of the VGICs, newer details on
the roles of S1-S6 helices, channel structure, relaxation,
hysteresis and gating, have not refuted but verified and extended
that knowledge (162,163,165,218-221).
What is more difficult to explain is the existence
of non-linear phenomena such as the increased bioactivity 'windows'
reported occasionally in the EMF-bioeffects literature, where
certain effects are intensified within certain values of an
EMF-exposure parameter (intensity in most cases, or frequency)
(1,40,151-153,222). The existence of 'windows' shows
that the response of living cells/organisms to EMFs is not
generally proportional to the aforementioned EMF-parameters.
Non-linear responses of living cells have not been explored in
depth and it will take a number of years until they are. A possible
explanation of observed intensity 'windows' according to the
described mechanism has been suggested as being due to an existing
upper limit in the membrane gating voltage change (222). Indeed, such an upper limit seems
to exist. The VGICs respond to membrane voltage changes from ~30 mV
(minimum) to ~100 mV (maximum) where the conductivity of the
channel saturates (218,221). Apart from this possible
explanation, no other explanation for the observed 'window' effects
has been provided so far.
An effect not included in the bioactivity Eqs. 11 and 12 is the increased bioactivity of
highly and unpredictably varying exposure such as those from WC
devices (including mobile phones and Wi-Fi) and corresponding
antennas (4,121,122). The described mechanism results
in accurate predictions when the applied EMFs have constant
parameters (intensity and frequency, among others). When the
parameters are highly and unpredictably variable, the mechanism,
and any possible mechanism, can only estimate effects according to
the average and maximum exposure values of the varying EMFs.
Finally, the bioactivity equations include field (and tissue)
parameters and not exposure variables such as exposure duration or
intermittence, which are also very important (16,17,19,41,55,122). One way to include such
parameters is to multiply the right parts of Eqs. 11 and 12 by certain coefficient(s), which
would be estimated experimentally. This could be a subject for
future development of the theory.
This theory has successfully explained for the
first time the sensing of upcoming earthquakes by animals, and the
sensing of upcoming thunderstorms by sensitive individuals through
the action of the partially polarized natural EMFs associated with
these phenomena (146,223).
Any 'mechanism' in science (particularly in
physics) must be based on simple and reasonable postulates, and
must necessarily be expressed quantitatively (by solvable equations
and numbers). The values of the different parameters in the
equations must be based on physical/molecular data. Qualitative
descriptions alone or incomplete quantitative descriptions based on
incomplete or unsolvable equations do not constitute a 'mechanism'.
The presented biophysical mechanism (143-146) is the only one that fulfills the
afore-mentioned criteria in the case of EMF-induced bioeffects.
Previous important attempts on mechanisms focusing on ions moving
inside membrane channels or other proteins (224-227) were not successful, mainly for
the following reasons: i) They had not taken into account damping
and restoration forces (224,226), or did not calculate them
(225,227). The difficulty was not related
with considering such forces, as this is standard in oscillation
mechanics, but with calculating their parameters such as β
and ωo, or the maximum velocity of the ion
(uo) within a channel. ii) They did not consider
coordinated motion of several ions oscillating in parallel and in
phase due to polarization and coherence, exerting additive forces
on channel sensors, which prevail against the greater but chaotic
forces due to the random thermal motion of the ions. iii) They
focused on magnetic fields and magnetically induced electric ones,
and ignored externally applied electric fields, which eventually
seem to be more bioactive (191). iv) They did not result in
numbers for field intensity versus frequency necessary to affect
cells, although some experimental reports have indicated bioactive
frequencies close to those predicted by Liboff's ion cyclotron
resonance (ICR) model (224,228), possibly indicating some
additional/secondary resonance mechanism involving ICR phenomenon
(169). v) Apart from the study
by Balcavage et al (226), there was no focus on the gating
of VGICs, which is by far a more probable event to initiate
biological effects, but simply on the motion of ions within
channels/proteins.
Several other suggestions on possible mechanisms
also face problems on fundamental issues (229-231). What is termed by Pall 'VGCC
activation mechanism' and presented as his own discovery is none
other than the mechanism presented here. A commentary paper/letter
to the editor was published on this major ethical issue (129). An extended review of suggested
mechanisms has been written by Creasey and Goldberg (169).
It has been claimed that the ELF components of
complex RF-ELF EMFs of WC need to be 'demodulated' in order to be
sensed by living organisms (232). 'Demodulated' or not, the fact is
that the ELF components of modulated/pulsed WC signals can be
directly sensed by both ELF meters/spectrum analyzers and living
organisms (40,55).
Although there have been successive publications of
this mechanism since 2000 (144), the subject is of great
importance and in each consecutive publication additional important
aspects are elucidated and/or refined. In our previous study in
2002 (145), the mechanism was
extended to include oscillating magnetic fields and the thermal
noise problem was discussed in more depth, while in 2015 (143) the mechanism was applied to
reveal the importance of polarization/coherence in the bioactivity
of man-made EMFs. In 2017 (223)
and 2020 (146), it was applied
to explain the sensing of upcoming thunderstorms and earthquakes,
respectively, by sensitive humans/animals. In the present study,
several aspects are further refined, including: i) The distance of
S4 sensors from the channel pore; ii) more details on damping
coefficient β and bioactivity constant k (Eq. 11); iii) further explanation of
the role of the constant term in the solution (Eq. 3); iv) the similarity of proton
voltage-gated channels with the other VGICs; v) numerical examples
demonstrating the ability of the pulsing ELF electric and magnetic
fields of 2G/3G/4G MT, DECT, Wi-Fi, Bluetooth, and the power line
ELF fields to induce biological/health effects; vi) the velocity of
oscillating ions; vii) bioactivity diagram extended to intensities
down to 10−5 V/m; and viii) discussion on other
suggested mechanisms.
Moreover, the present study documented how the
impaired function of VGICs on the membranes of living cells
triggers (over)production of free radicals/ROS, such as the most
potent OH• produced by H2O2 via
the Fenton reaction, and ONOO− produced by
NO•. These are considered the main damaging species for
DNA and other critical biological molecules. It is estimated that
approximately two-thirds of the DNA damage caused by ionizing
radiation is due to OH• (233,234). Although OH• can only
diffuse at distances comparable to the length of a macromolecule,
H2O2 can move to any intracellular site.
Thus, even though the most potent OH• due to its high
reactivity has an extremely short lifetime (of the order of
10−9-10−4 s depending on the presence of
other molecules) it can be formed by H2O2 at
any location within the cell (including the nucleus) and act
instantly upon DNA or other macromolecules (233,234). As for
NO•/ONOO−, they can be diffused anywhere in
the cell and thus directly affect any molecule, including DNA
(181). Even though the present
study identified specific pathways of ROS over-production or the
release of DNases connected with disrupted ionic concentrations in
EMF-exposed cells, the exact molecular mechanisms need to be
further explored and elucidated.
Finally, the present study discussed how
unrepaired/misrepaired DNA lesions/damage such as strand breaks,
covalent bond breakage or nucleotide base damages, lead to cell
senescence, cell death or mutations, and related pathologies,
including cancer. Even though effective mechanisms have evolved in
all animals/cells for repairing DNA damage induced by environmental
stressors, it is very different when the damaging events are
isolated or random (e.g. radioactive particles or γ-photons of
cosmic/natural radioactivity, or sporadic x-ray diagnostic
exposure), compared with persisting/repeated exposure to cytotoxic
agents, even when these agents are relatively weaker. Exposure to
human-made EMFs and especially to the most detrimental ones from WC
antennas/devices and high-voltage transmission lines (4) has become a new reality in modern
life. Billions of people are exposed to such EMFs on a daily basis.
Although they are less cytotoxic than radioactivity or certain
cytotoxic chemicals, they represent the most persistent daily
cytotoxic stressors against which any repair mechanisms cannot be
efficient enough. By contrast, previously existing cytotoxic agents
expose us randomly as isolated events. When an organism is
constantly under OS due to a totally new cytotoxic agent such as
human-made EMFs, no protective mechanism, evolved in the billions
of years of biological evolution to protect from natural
(non-polarized) EMFs/radiation or isolated hazardous events, can be
effective enough.
The repair capability of cells in response to DNA
damage is crucial for the final outcome. The threshold of damage
above which it becomes irreparable depends on cell type and the
health and status of the organism. An organism with poor health
and/or under stress and inflammation due to OS is expected to have
decreased repair capability and increased cancer risk. Epigenetic
effects such as altered gene expression may also lead to cellular
dysfunction and carcinogenesis (133,235,236).
Both DNA damage and alterations in protein
synthesis, especially increased levels of stress proteins, are
reported to be induced similarly by both ELF and pulsing RF EMFs
(237,238). However, the effects of pulsing
RF were attributed to the carrier frequency, and it was not
considered that perhaps in both cases (ELF and pulsing RF) the ELF
components might be responsible for the effects, as suggested now
by the present study.
To the best of our knowledge, the present study
provides for the first time a complete and precise
biophysical/biochemical picture to explain the great number of
experimental and epidemiological findings connecting human-made EMF
exposure with DNA damage and related pathologies such as cancer,
infertility and neurodegenerative diseases.
The long-existing experimental and epidemiological
findings connecting exposure to human-made EMFs and DNA damage,
infertility and cancer, are now explained by the presented complete
mechanism. The present study should provide a basis for further
research and encourage health authorities to take measures for the
protection of life on Earth against unrestricted use of human-made
EMFs.
Availability of data and materials
Not applicable.
Authors' contributions
DJP designed the study and wrote the main
manuscript. AK verified all equations and calculations. IY
coauthored section 3 on biochemical processes. GPC reviewed and
evaluated all data. All authors have read and approved the
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
DECT
|
digitally enhanced cordless
telecommunications
|
|
ELF
|
extremely low frequency
|
|
EMF
|
electromagnetic field
|
|
MT
|
mobile telephony
|
|
OS
|
oxidative stress
|
|
RF
|
radio frequency
|
|
ROS
|
reactive oxygen species
|
|
ULF
|
ultra low frequency
|
|
VGICs
|
voltage-gated ion channels
|
|
VGCCs
|
voltage-gated calcium channels
|
|
WC
|
wireless communications
|
|
Wi-Fi
|
wireless fidelity
|
|
2G/3G/4G/5G
|
second/third/fourth/fifth-generation
of mobile telephony
|
References
|
1
|
Goodman EM, Greenebaum B and Marron MT:
Effects of electromagnetic fields on molecules and cells. Int Rev
Cytol. 158:279–338. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Santini MT, Ferrante A, Rainaldi G,
Indovina P and Indovina PL: Extremely low frequency (ELF) magnetic
fields and apoptosis: A review. Int J Radiat Biol. 81:1–11. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Phillips JL, Singh NP and Lai H:
Electromagnetic fields and DNA damage. Pathophysiology. 16:79–88.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Panagopoulos DJ: Comparing DNA damage
induced by mobile telephony and other types of man-made
electromagnetic fields. Mutat Res Rev Mutat Res. 781:53–62. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Delgado JMR: Biological effects of
extremely low frequency electromagnetic fields. J Bioelectricity.
4:75–92. 1985. View Article : Google Scholar
|
|
6
|
Garaj-Vrhovac V, Horvat D and Koren Z: The
effect of microwave radiation on the cell genome. Mutat Res.
243:87–93. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Garaj-Vrhovac V, Horvat D and Koren Z: The
relationship between colony-forming ability, chromosome aberrations
and incidence of micronuclei in V79 Chinese hamster cells exposed
to microwave radiation. Mutat Res. 263:143–149. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garaj-Vrhovac V, Fucić A and Horvat D: The
correlation between the frequency of micronuclei and specific
chromosome aberrations in human lymphocytes exposed to microwave
radiation in vitro. Mutat Res. 281:181–186. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma TH and Chu KC: Effect of the extremely
low frequency (ELF) electromagnetic field (EMF) on developing
embryos of the fruit fly (Drosophila melanogaster L.). Mutat Res.
303:35–39. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sarkar S, Ali S and Behari J: Effect of
low power microwave on the mouse genome: A direct DNA analysis.
Mutat Res. 320:141–147. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lai H and Singh NP: Acute low-intensity
microwave exposure increases DNA single-strand breaks in rat brain
cells. Bioelectromagnetics. 16:207–210. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lai H and Singh NP: Single- and
double-strand DNA breaks in rat brain cells after acute exposure to
radiofrequency electro-magnetic radiation. Int J Radiat Biol.
69:513–521. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lai H and Singh NP: Acute exposure to a 60
Hz magnetic field increases DNA strand breaks in rat brain cells.
Bioelectromagnetics. 18:156–165. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Svedenstål BM, Johanson KJ and Mild KH:
DNA damage induced in brain cells of CBA mice exposed to magnetic
fields. In Vivo. 13:551–552. 1999.
|
|
15
|
Koana T, Okada MO, Takashima Y, Ikehata M
and Miyakoshi J: Involvement of eddy currents in the mutagenicity
of ELF magnetic fields. Mutat Res. 476:55–62. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ivancsits S, Diem E, Pilger A, Rüdiger HW
and Jahn O: Induction of DNA strand breaks by intermittent exposure
to extremely-low-frequency electromagnetic fields in human diploid
fibroblasts. Mutat Res. 519:1–13. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ivancsits S, Diem E, Jahn O and Rüdiger
HW: Intermittent extremely low frequency electromagnetic fields
cause DNA damage in a dose-dependent way. Int Arch Occup Environ
Health. 76:431–436. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Winker R, Ivancsits S, Pilger A, Adlkofer
F and Rüdiger HW: Chromosomal damage in human diploid fibroblasts
by intermittent exposure to extremely low-frequency electromagnetic
fields. Mutat Res. 585:43–49. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Diem E, Schwarz C, Adlkofer F, Jahn O and
Rüdiger H: Non-thermal DNA breakage by mobile-phone radiation (1800
MHz) in human fibroblasts and in transformed GFSH-R17 rat granulosa
cells in vitro. Mutat Res. 583:178–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mausset-Bonnefont AL, Hirbec H, Bonnefont
X, Privat A, Vignon J and de Sèze R: Acute exposure to GSM 900-MHz
electromagnetic fields induces glial reactivity and biochemical
modifications in the rat brain. Neurobiol Dis. 17:445–454. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ji S, Oh E, Sul D, Choi JW, Park H and Lee
E: DNA damage of lymphocytes in volunteers after 4 h use of mobile
phone. J Prev Med Public Health. 37:373–380. 2004.PubMed/NCBI
|
|
22
|
Hong R, Zhang Y, Liu Y and Weng EQ:
Effects of extremely low frequency electromagnetic fields on DNA of
testicular cells and sperm chromatin structure in mice. Zhonghua
Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. 23:414–417. 2005.In
Chinese.
|
|
23
|
Belyaev IY, Hillert L, Protopopova M, Tamm
C, Malmgren LO, Persson BR, Selivanova G and Harms-Ringdahl M: 915
MHz microwaves and 50 Hz magnetic field affect chromatin
conformation and 53BP1 foci in human lymphocytes from
hypersensitive and healthy persons. Bioelectromagnetics.
26:173–184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Markovà E, Hillert L, Malmgren L, Persson
BR and Belyaev IY: Microwaves from GSM mobile telephones affect
53BP1 and gamma-H2AX foci in human lymphocytes from hypersensitive
and healthy persons. Environ Health Perspect. 113:1172–1177. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aitken RJ, Bennetts LE, Sawyer D, Wiklendt
AM and King BV: Impact of radio frequency electromagnetic radiation
on DNA integrity in the male germline. Int J Androl. 28:171–179.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nikolova T, Czyz J, Rolletschek A,
Blyszczuk P, Fuchs J, Jovtchev G, Schuderer J, Kuster N and Wobus
AM: Electromagnetic fields affect transcript levels of
apoptosis-related genes in embryonic stem cell-derived neural
progenitor cells. FASEB J. 19:1686–1688. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang DY, Xu ZP, Chiang H, Lu DQ and Zeng
QL: Effects of GSM 1800 MHz radiofrequency electromagnetic fields
on DNA damage in Chinese hamster lung cells. Zhonghua Yu Fang Yi
Xue Za Zhi. 40:149–152. 2006.In Chinese. PubMed/NCBI
|
|
28
|
Lixia S, Yao K, Kaijun W, Deqiang L,
Huajun H, Xiangwei G, Baohong W, Wei Z, Jianling L and Wei W:
Effects of 1.8 GHz radiofrequency field on DNA damage and
expression of heat shock protein 70 in human lens epithelial cells.
Mutat Res. 602:135–142. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ferreira AR, Knakievicz T, Pasquali MA,
Gelain DP, Dal-Pizzol F, Fernández CE, de Salles AA, Ferreira HB
and Moreira JC: Ultra high frequency-electromagnetic field
irradiation during pregnancy leads to an increase in erythrocytes
micronuclei incidence in rat offspring. Life Sci. 80:43–50. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Panagopoulos DJ, Chavdoula ED, Nezis IP
and Margaritis LH: Cell death induced by GSM 900-MHz and DCS
1800-MHz mobile telephony radiation. Mutat Res. 626:69–78. 2007.
View Article : Google Scholar
|
|
31
|
Yan JG, Agresti M, Bruce T, Yan YH,
Granlund A and Matloub HS: Effects of cellular phone emissions on
sperm motility in rats. Fertil Steril. 88:957–964. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yao K, Wu W, Wang K, Ni S, Ye P, Yu Y, Ye
J and Sun L: Electromagnetic noise inhibits radiofrequency
radiation-induced DNA damage and reactive oxygen species increase
in human lens epithelial cells. Mol Vis. 14:964–969.
2008.PubMed/NCBI
|
|
33
|
Yadav AS and Sharma MK: Increased
frequency of micronucleated exfoliated cells among humans exposed
in vivo to mobile telephone radiations. Mutat Res. 650:175–180.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sokolovic D, Djindjic B, Nikolic J,
Bjelakovic G, Pavlovic D, Kocic G, Krstic D, Cvetkovic T and
Pavlovic V: Melatonin reduces oxidative stress induced by chronic
exposure of microwave radiation from mobile phones in rat brain. J
Radiat Res. 49:579–586. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee KS, Choi JS, Hong SY, Son TH and Yu K:
Mobile phone electromagnetic radiation activates MAPK signaling and
regulates viability in Drosophila. Bioelectromagnetics. 29:371–379.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
De Iuliis GN, Newey RJ, King BV and Aitken
RJ: Mobile phone radiation induces reactive oxygen species
production and DNA damage in human spermatozoa in vitro. PLoS One.
4:e64462009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Agarwal A, Desai NR, Makker K, Varghese A,
Mouradi R, Sabanegh E and Sharma R: Effects of radiofrequency
electromagnetic waves (RF-EMW) from cellular phones on human
ejaculated semen: An in vitro pilot study. Fertil Steril.
92:1318–1325. 2009. View Article : Google Scholar
|
|
38
|
Mailankot M, Kunnath AP, Jayalekshmi H,
Koduru B and Valsalan R: Radio frequency electromagnetic radiation
(RF-EMR) from GSM (0.9/1.8 GHz) mobile phones induces oxidative
stress and reduces sperm motility in rats. Clinics (Sao Paulo).
64:561–565. 2009. View Article : Google Scholar
|
|
39
|
Luukkonen J, Hakulinen P, Mäki-Paakkanen
J, Juutilainen J and Naarala J: Enhancement of chemically induced
reactive oxygen species production and DNA damage in human SH-SY5Y
neuroblastoma cells by 872 MHz radiofrequency radiation. Mutat Res.
662:54–58. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Panagopoulos DJ, Chavdoula ED and
Margaritis LH: Bioeffects of mobile telephony radiation in relation
to its intensity or distance from the antenna. Int J Radiat Biol.
86:345–357. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chavdoula ED, Panagopoulos DJ and
Margaritis LH: Comparison of biological effects between continuous
and intermittent exposure to GSM-900-MHz mobile phone radiation.
Detection of apoptotic cell death features Mutat Res. 700:51–61.
2010.
|
|
42
|
Guler G, Tomruk A, Ozgur E and Seyhan N:
The effect of radio-frequency radiation on DNA and lipid damage in
non-pregnant and pregnant rabbits and their newborns. Gen Physiol
Biophys. 29:59–66. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tomruk A, Guler G and Dincel AS: The
influence of 1800 MHz GSM-like signals on hepatic oxidative DNA and
lipid damage in nonpregnant, pregnant, and newly born rabbits. Cell
Biochem Biophys. 56:39–47. 2010. View Article : Google Scholar
|
|
44
|
Franzellitti S, Valbonesi P, Ciancaglini
N, Biondi C, Contin A, Bersani F and Fabbri E: Transient DNA damage
induced by high-frequency electromagnetic fields (GSM 1.8 GHz) in
the human trophoblast HTR-8/SVneo cell line evaluated with the
alkaline comet assay. Mutat Res. 683:35–42. 2010. View Article : Google Scholar
|
|
45
|
Campisi A, Gulino M, Acquaviva R, Bellia
P, Raciti G, Grasso R, Musumeci F, Vanella A and Triglia A:
Reactive oxygen species levels and DNA fragmentation on astrocytes
in primary culture after acute exposure to low intensity microwave
electromagnetic field. Neurosci Lett. 473:52–55. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Panagopoulos DJ: Effect of microwave
exposure on the ovarian development of Drosophila melanogaster.
Cell Biochem Biophys. 63:121–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Panagopoulos DJ, Karabarbounis A and
Lioliousis C: ELF alternating magnetic field decreases reproduction
by DNA damage induction. Cell Biochem Biophys. 67:703–716. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu C, Gao P, Xu SC, Wang Y, Chen CH, He
MD, Yu ZP, Zhang L and Zhou Z: Mobile phone radiation induces
mode-dependent DNA damage in a mouse spermatocyte-derived cell
line: A protective role of melatonin. Int J Radiat Biol.
89:993–1001. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Pesnya DS and Romanovsky AV: Comparison of
cytotoxic and genotoxic effects of plutonium-239 alpha particles
and mobile phone GSM 900 radiation in the Allium cepa test. Mutat
Res. 750:27–33. 2013. View Article : Google Scholar
|
|
50
|
Mihai CT, Rotinberg P, Brinza F and
Vochita G: Extremely low-frequency electromagnetic fields cause DNA
strand breaks in normal cells. J Environ Health Sci Eng. 12:152014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Daroit NB, Visioli F, Magnusson AS, Vieira
GR and Rados PV: Cell phone radiation effects on cytogenetic
abnormalities of oral mucosal cells. Braz Oral Res. 29:1–8. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Banerjee S, Singh NN, Sreedhar G and
Mukherjee S: Analysis of the genotoxic effects of mobile phone
radiation using buccal micronucleus assay: A comparative
evaluation. J Clin Diagn Res. 10:ZC82–ZC85. 2016.PubMed/NCBI
|
|
53
|
D'Silva MH, Swer RT, Anbalagan J and
Rajesh B: Effect of radiofrequency radiation emitted from 2G and 3G
cell phone on developing liver of chick embryo-a comparative study.
J Clin Diagn Res. 11:AC05–AC09. 2017.
|
|
54
|
Panagopoulos DJ: Chromosome damage in
human cells induced by UMTS mobile telephony radiation. Gen Physiol
Biophys. 38:445–454. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Panagopoulos DJ: Comparing chromosome
damage induced by mobile telephony radiation and a high caffeine
dose: Effect of combination and exposure duration. Gen Physiol
Biophys. 39:531–544. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Magras IN and Xenos TD: RF
radiation-induced changes in the prenatal development of mice.
Bioelectromagnetics. 18:455–461. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Panagopoulos DJ, Karabarbounis A and
Margaritis LH: Effect of GSM 900-MHz mobile phone radiation on the
reproductive capacity of Drosophila melanogaster. Electromagn Biol
Med. 23:29–43. 2004. View Article : Google Scholar
|
|
58
|
Panagopoulos DJ, Chavdoula ED,
Karabarbounis A and Margaritis LH: Comparison of bioactivity
between GSM 900 MHz and DCS 1800 MHz mobile telephony radiation.
Electromagn Biol Med. 26:33–44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wdowiak A, Wdowiak L and Wiktor H:
Evaluation of the effect of using mobile phones on male fertility.
Ann Agric Environ Med. 14:169–172. 2007.PubMed/NCBI
|
|
60
|
Agarwal A, Deepinder F, Sharma RK, Ranga G
and Li J: Effect of cell phone usage on semen analysis in men
attending infertility clinic: An observational study. Fertil
Steril. 89:124–128. 2008. View Article : Google Scholar
|
|
61
|
Batellier F, Couty I, Picard D and
Brillard JP: Effects of exposing chicken eggs to a cell phone in
'call' position over the entire incubation period. Theriogenology.
69:737–745. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gul A, Celebi H and Uğraş S: The effects
of microwave emitted by cellular phones on ovarian follicles in
rats. Arch Gynecol Obstet. 280:729–733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sharma VP and Kumar NR: Changes in honey
bee behaviour and biology under the influence of cell phone
radiations. Curr Sci. 98:1376–1378. 2010.
|
|
64
|
La Vignera S, Condorelli RA, Vicardi E,
D'Agata R and Calogero AE: Effects of the exposure to mobile phones
on male reproduction: A review of the literature. J Androl.
33:350–356. 2012. View Article : Google Scholar
|
|
65
|
Balmori A: Possible effects of
electromagnetic fields from phone masts on a population of white
stork (Ciconia ciconia). Electromagn Biol Med. 24:109–119. 2005.
View Article : Google Scholar
|
|
66
|
Balmori A and Hallberg O: The urban
decline of the house sparrow (Passer domesticus): A possible link
with electromagnetic radiation. Electromagn Biol Med. 26:141–151.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Everaert J and Bauwens D: A possible
effect of electromagnetic radiation from mobile phone base stations
on the number of breeding house sparrows (Passer domesticus).
Electromagn Biol Med. 26:63–72. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bacandritsos N, Granato A, Budge G,
Papanastasiou I, Roinioti E, Caldon M, Falcaro C, Gallina A and
Mutinelli F: Sudden deaths and colony population decline in Greek
honey bee colonies. J Invertebr Pathol. 105:335–340. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cucurachi S, Tamis WL, Vijver MG,
Peijnenburg WJ, Bolte JF and de Snoo GR: A review of the ecological
effects of radio-frequency electromagnetic fields (RF-EMF). Environ
Int. 51:116–140. 2013. View Article : Google Scholar
|
|
70
|
Balmori A: The incidence of
electromagnetic pollution on the amphibian decline: Is this an
important piece of the puzzle? Toxicol Environ Chem. 88:287–299.
2006. View Article : Google Scholar
|
|
71
|
Balmori A: Mobile phone mast effects on
common frog (Rana temporaria) tadpoles: The city turned into a
laboratory. Electromagn Biol Med. 29:31–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wertheimer N and Leeper E: Electrical
wiring configurations and childhood cancer. Am J Epidemiol.
109:273–284. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Savitz DA, Wachtel H, Barnes F, John EM
and Tvrdik JG: Case-control study of childhood cancer and exposure
to 60-Hz magnetic fields. Am J Epidemiol. 128:21–38. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Coleman MP, Bell CM, Taylor HL and
Primic-Zakelj M: Leukaemia and residence near electricity
transmission equipment: A case-control study. Br J Cancer.
60:793–798. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Feychting M and Ahlbom A: Magnetic fields
and cancer in children residing near Swedish high-voltage power
lines. Am J Epidemiol. 138:467–481. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Feychting M and Ahlbom A: Magnetic fields,
leukemia, and central nervous system tumors in Swedish adults
residing near high-voltage power lines. Epidemiology. 5:501–509.
1994.PubMed/NCBI
|
|
77
|
Feychting M and Ahlbom A: Childhood
leukemia and residential exposure to weak extremely low frequency
magnetic fields. Environ Health Perspect. 103(Suppl 2): S59–S62.
1995.
|
|
78
|
Coghill RW, Steward J and Philips A: Extra
low frequency electric and magnetic fields in the bed place of
children diagnosed with leukaemia: A case-control study. Eur J
Cancer Prev. 5:153–158. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Ahlbom A, Day N, Feychting M, Roman E,
Skinner J, Dockerty J, Linet M, McBride M, Michaelis J, Olsen JH,
et al: A pooled analysis of magnetic fields and childhood
leukaemia. Br J Cancer. 83:692–698. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Greenland S, Sheppard AR, Kaune WT, Poole
C and Kelsh MA: A pooled analysis of magnetic fields, wire codes,
and childhood leukemia. Childhood leukemia-EMF study group
Epidemiology. 11:624–634. 2000.
|
|
81
|
Draper G, Vincent T, Kroll ME and Swanson
J: Childhood cancer in relation to distance from high voltage power
lines in England and Wales: A case-control study. BMJ.
330:12902005. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Kheifets L, Ahlbom A, Crespi CM, Draper G,
Hagihara J, Lowenthal RM, Mezei G, Oksuzyan S, Schüz J, Swanson J,
et al: Pooled analysis of recent studies on magnetic fields and
childhood leukaemia. Br J Cancer. 103:1128–1135. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hallberg O and Johansson O: Melanoma
incidence and frequency modulation (FM) broadcasting. Arch Environ
Health. 57:32–40. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Gulati S, Yadav A, Kumar N, Kanupriya,
Aggarwal NK, Kumar R and Gupta R: Effect of GSTM1 and GSTT1
polymorphisms on genetic damage in humans populations exposed to
radiation from mobile towers. Arch Environ Contam Toxicol.
70:615–625. 2016. View Article : Google Scholar
|
|
85
|
Zothansiama, Zosangzuali M, Lalramdinpuii
M and Jagetia GC: Impact of radiofrequency radiation on DNA damage
and anti-oxidants in peripheral blood lymphocytes of humans
residing in the vicinity of mobile phone base stations. Electromagn
Biol Med. 36:295–305. 2017. View Article : Google Scholar
|
|
86
|
Hardell L, Carlberg M, Söderqvist F, Mild
KH and Morgan LL: Long-term use of cellular phones and brain
tumours: Increased risk associated with use for > or =10 years.
Occup Environ Med. 64:626–632. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hardell L, Carlberg M and Hansson Mild K:
Epidemiological evidence for an association between use of wireless
phones and tumor diseases. Pathophysiology. 16:113–122. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Khurana VG, Teo C, Kundi M, Hardell L and
Carlberg M: Cell phones and brain tumors: A review including the
long-term epidemiologic data. Surg Neurol. 72:205–214. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hardell L and Carlberg M: Mobile phones,
cordless phones and the risk for brain tumours. Int J Oncol.
35:5–17. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Hardell L, Carlberg M, Söderqvist F and
Mild KH: Pooled analysis of case-control studies on acoustic
neuroma diagnosed 1997-2003 and 2007-2009 and use of mobile and
cordless phones. Int J Oncol. 43:1036–1044. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Hardell L, Carlberg M, Söderqvist F and
Mild KH: Case-control study of the association between malignant
brain tumours diagnosed between 2007 and 2009 and mobile and
cordless phone use. Int J Oncol. 43:1833–1845. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hardell L, Carlberg M and Hansson Mild K:
Use of mobile phones and cordless phones is associated with
increased risk for glioma and acoustic neuroma. Pathophysiology.
20:85–110. 2013. View Article : Google Scholar
|
|
93
|
Wang Y and Guo X: Meta-analysis of
association between mobile phone use and glioma risk. J Cancer Res
Ther. 12(Suppl): C298–C300. 2016. View Article : Google Scholar
|
|
94
|
Carlberg M and Hardell L: Evaluation of
mobile phone and cordless phone use and glioma risk using the
bradford hill viewpoints from 1965 on association or causation.
Biomed Res Int. 2017:92184862017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hardell L: Effects of mobile phones on
children's and adolescents' health: A commentary. Child Dev.
89:137–140. 2018. View Article : Google Scholar
|
|
96
|
Momoli F, Siemiatycki J, McBride ML,
Parent MÉ, Richardson L, Bedard D, Platt R, Vrijheid M, Cardis E
and Krewski D: Probabilistic multiple-bias modelling applied to the
Canadian data from the INTERPHONE study of mobile phone use and
risk of glioma, meningioma, acoustic neuroma, and parotid gland
tumors. Am J Epidemiol. 186:885–893. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Miller AB, Morgan LL, Udasin I and Davis
DL: Cancer epidemiology update following the 2011 IARC evaluation
of radiofrequency electromagnetic fields (Monograph 102). Environ
Res. 167:673–683. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Miller AB, Sears ME, Morgan LL, Davis DL,
Hardell L, Oremus M and Soskolne CL: Risks to health and well-being
from radio-frequency radiation emitted by cell phones and other
wireless devices. Front Public Health. 7:2232019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Santini R, Santini P, Danze JM, Le Ruz P
and Seigne M: Study of the health of people living in the vicinity
of mobile phone base stations: I. Influences of distance and sex
Pathol Biol. 50:369–373. 2002. View Article : Google Scholar
|
|
100
|
Navarro A, Garcia JS, Portoles M and
Gómez-Perretta G: The microwave syndrome: A preliminary study in
Spain. Electromagn Biol Med. 22:161–169. 2003. View Article : Google Scholar
|
|
101
|
Salama OE and Abou El Naga RM: Cellular
phones: Are they detrimental? J Egypt Public Health Assoc.
79:197–223. 2004.
|
|
102
|
Hutter HP, Moshammer H, Wallner P and
Kundi M: Subjective symptoms, sleeping problems, and cognitive
performance in subjects living near mobile phone base stations.
Occup Environ Med. 63:307–313. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Abdel-Rassoul G, El-Fateh OA, Salem MA,
Michael A, Farahat F, El-Batanouny M and Salem E: Neurobehavioral
effects among inhabitants around mobile phone base stations.
Neurotoxicology. 28:434–440. 2007. View Article : Google Scholar
|
|
104
|
Blettner M, Schlehofer B, Breckenkamp J,
Kowall B, Schmiedel S, Reis U, Potthoff P, Schüz J and
Berg-Beckhoff G: Mobile phone base stations and adverse health
effects: Phase 1 of a population-based, cross-sectional study in
Germany. Occup Environ Med. 66:118–123. 2009. View Article : Google Scholar
|
|
105
|
Viel JF, Clerc S, Barrera C, Rymzhanova R,
Moissonnier M, Hours M and Cardis E: Residential exposure to
radiofrequency fields from mobile phone base stations, and
broadcast transmitters: A population-based survey with personal
meter. Occup Environ Med. 66:550–556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Kundi M and Hutter HP: Mobile phone base
stations-effects on wellbeing and health. Pathophysiology.
16:123–135. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Shahbazi-Gahrouei D, Karbalae M, Moradi HA
and Baradaran-Ghahfarokhi M: Health effects of living near mobile
phone base transceiver station (BTS) antennae: A report from
Isfahan, Iran. Electromagn Biol Med. 33:206–210. 2014. View Article : Google Scholar
|
|
108
|
Irigaray P, Caccamo D and Belpomme D:
Oxidative stress in electrohypersensitivity self-reporting
patients: Results of a prospective in vivo investigation with
comprehensive molecular analysis. Int J Mol Med. 42:1885–1898.
2018.PubMed/NCBI
|
|
109
|
Yakymenko I, Tsybulin O, Sidorik E,
Henshel D, Kyrylenko O and Kyrylenko S: Oxidative mechanisms of
biological activity of low-intensity radiofrequency radiation.
Electromagn Biol Med. 35:186–202. 2016. View Article : Google Scholar
|
|
110
|
Tillmann T, Ernst H, Streckert J, Zhou Y,
Taugner F, Hansen V and Dasenbrock C: Indication of cocarcinogenic
potential of chronic UMTS-modulated radiofrequency exposure in an
ethylnitrosourea mouse model. Int J Radiat Biol. 86:529–541. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lerchl A, Klose M, Grote K, Wilhelm AF,
Spathmann O, Fiedler T, Streckert J, Hansen V and Clemens M: Tumor
promotion by exposure to radiofrequency electromagnetic fields
below exposure limits for humans. Biochem Biophys Res Commun.
459:585–590. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
National Toxicology Program: Toxicology
and carcinogenesis studies in Sprague Dawley (Hsd:Sprague Dawley
SD) rats exposed to whole-body radio frequency radiation at a
frequency (900 MHz) and modulations (GSM and CDMA) used by cell
phones. Natl Toxicol Program Tech Rep Ser: NTP-TR-595. 2018.
|
|
113
|
Smith-Roe SL, Wyde ME, Stout MD, Winters
JW, Hobbs CA, Shepard KG, Green AS, Kissling GE, Shockley KR, Tice
RR, et al: Evaluation of the genotoxicity of cell phone
radiofrequency radiation in male and female rats and mice following
subchronic exposure. Environ Mol Mutagen. 61:276–290. 2020.
View Article : Google Scholar :
|
|
114
|
Falcioni L, Bua L, Tibaldi E, Lauriola M,
De Angelis L, Gnudi F, Mandrioli D, Manservigi M, Manservisi F,
Manzoli I, et al: Report of final results regarding brain and heart
tumors in Sprague-Dawley rats exposed from prenatal life until
natural death to mobile phone radiofrequency field representative
of a 1.8 GHz GSM base station environmental emission. Environ Res.
165:496–503. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
International Commission on Non-Ionizing
Radiation Protection: Guidelines for limiting exposure to
time-varying electric and magnetic fields (1 Hz to 100 kHz). Health
Phys. 99:818–836. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
International Commission on Non-Ionizing
Radiation Protection: Guidelines for limiting exposure to
electromagnetic fields (100 kHz to 300 GHz). Health Phys.
118:483–524. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
IARC: Non-ionizing radiation, part 1:
Static and extremely low-frequency (ELF) electric and magnetic
fields. 80. World Health Organization. IARC Press; Lyon: 2002
|
|
118
|
IARC: Non-Ionizing Radiation, Part 2:
Radiofrequency Electromagnetic Fields. 102. IARC Press; Lyon:
2013
|
|
119
|
Baan R, Grosse Y, Lauby-Secretan B, El
Ghissassi F, Bouvard V, Benbrahim-Tallaa L, Guha N, Islami F and
Galichet L: Carcinogenicity of radiofrequency electromagnetic
fields. Lancet Oncol. 12:624–626. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Verschaeve L: Misleading scientific papers
on health effects from wireless communication devices. Microwave
Effects on DNA and Proteins. Geddes CD: Springer; Cham: pp.
159–233. 2017, View Article : Google Scholar
|
|
121
|
Panagopoulos DJ, Johansson O and Carlo GL:
Real versus simulated mobile phone exposures in experimental
studies. Biomed Res Int. 2015:6070532015. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Panagopoulos DJ: Mobile telephony
radiation effects on insect ovarian cells. The necessity for real
exposures bioactivity assessment The key role of polarization, and
the 'ion forced-oscillation mechanism'. Microwave Effects on DNA
and Proteins. Geddes CD: Springer; Cham: pp. 1–48. 2017
|
|
123
|
Manna D and Ghosh R: Effect of
radiofrequency radiation in cultured mammalian cells: A review.
Electromagn Biol Med. 35:265–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Leach V, Weller S and Redmayne M: A novel
database of bio-effects from non-ionizing radiation. Rev Environ
Health. 33:273–280. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Karipidis K, Mate R, Urban D, Tinker R and
Wood A: 5G mobile networks and health-a state-of-the-science review
of the research into low-level RF fields above 6 GHz. J Expo Sci
Environ Epidemiol. 31:585–605. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hardell L and Nyberg R: Appeals that
matter or not on a moratorium on the deployment of the fifth
generation, 5G, for microwave radiation. Mol Clin Oncol.
12:247–257. 2020.PubMed/NCBI
|
|
127
|
Hardell L and Carlberg M: Health risks
from radiofrequency radiation, including 5G, should be assessed by
experts with no conflicts of interest. Oncol Lett.
20:152020.PubMed/NCBI
|
|
128
|
Metaxas AC: Microwave heating. Power Eng.
5:237–247. 1991. View Article : Google Scholar
|
|
129
|
Panagopoulos DJ: Comments on Pall's
'Millimeter (MM) wave and microwave frequency radiation produce
deeply penetrating effects: The biology and the physics'. Rev
Environ Health. Jul 12–2021, https://doi.org/10.1515/reveh-2021-0090. View Article : Google Scholar
|
|
130
|
Ames BN: Endogenous DNA damage as related
to cancer and aging. Mutat Res. 214:41–46. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Lieber MR: Pathological and physiological
double-strand breaks: Roles in cancer, aging, and the immune
system. Am J Pathol. 153:1323–1332. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Helleday T, Loc J, van Gentd DC and
Engelward BP: DNA double-strand break repair: From mechanistic
understanding to cancer treatment. DNA Repair (Amst). 6:923–935.
2007. View Article : Google Scholar
|
|
133
|
Lahtz C and Pfeifer GP: Epigenetic changes
of DNA repair genes in cancer. J Mol Cell Biol. 3:51–58. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Yao Y and Dai W: Genomic instability and
cancer. J Carcinog Mutagen. 5:10001652014.PubMed/NCBI
|
|
135
|
Bernstein C, Prasad AR, Nfonsam V and
Bernstein H: DNA damage, DNA repair and cancer. New Research
Directions in DNA Repair. Clarc C: InTech; Rijeka: pp. 413–465.
2013
|
|
136
|
Basu AK: DNA damage, mutagenesis and
cancer. Int J Mol Sci. 19:9702018. View Article : Google Scholar :
|
|
137
|
von Zglinicki T, Saretzki G, Ladhoff J,
d'Adda di Fagagna F and Jackson SP: Human cell senescence as a DNA
damage response. Mech Ageing Dev. 126:111–117. 2005. View Article : Google Scholar
|
|
138
|
Shah DJ, Sachs RK and Wilson DJ:
Radiation-induced cancer: A modern view. Br J Radiol.
85:e1166–e1173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Rodgers K and McVey M: Error-prone repair
of DNA double-strand breaks. J Cell Physiol. 231:15–24. 2016.
View Article : Google Scholar
|
|
140
|
Nadler DL and Zurbenko IG: Estimating
cancer latency times using a weibull model. Adv Epidemiol.
2014:7467692014. View Article : Google Scholar
|
|
141
|
Francis G: Ionization Phenomena in Gases.
Butterworths Scientific Publications; London: 1960
|
|
142
|
Gomer R: Field Emission and Field
Ionization. Harvard University Press; Cambridge: 1961
|
|
143
|
Panagopoulos DJ, Johansson O and Carlo GL:
Polarization: A key difference between man-made and natural
electromagnetic fields, in regard to biological activity. Sci Rep.
5:149142015. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Panagopoulos DJ, Messini N, Karabarbounis
A, Filippetis AL and Margaritis LH: A mechanism for action of
oscillating electric fields on cells. Biochem Biophys Res Commun.
272:634–640. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Panagopoulos DJ, Karabarbounis A and
Margaritis LH: Mechanism for action of electromagnetic fields on
cells. Biochem Biophys Res Commun. 298:95–102. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Panagopoulos DJ, Balmori A and Chrousos
GP: On the biophysical mechanism of sensing upcoming earthquakes by
animals. Sci Total Environ. 717:1369892020. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Alberts B, Johnson A, Lewis J, Raff M,
Roberts K and Walter P: Molecular Biology of the Cell. Garland
Publishing, Inc; New York: 1994
|
|
148
|
Stryer L: Biochemistry. 4th edition.
Freeman WH: Freeman and Company; New York, NY: 1995
|
|
149
|
Halgamuge MN and Abeyrathne CD: Behavior
of charged particles in a biological cell exposed to AC-DC
electromagnetic fields. Environ Eng Sci. 28:1–10. 2001. View Article : Google Scholar
|
|
150
|
Panagopoulos DJ and Karabarbounis A:
Comments on study of charged particle's behavior in a biological
cell exposed to AC-DC electromagnetic fields, and on comparison
between two models of interaction between electric and magnetic
fields and proteins in cell membranes. Environ Eng Sci. 28:749–751.
2011. View Article : Google Scholar
|
|
151
|
Bawin SM, Kaczmarek LK and Adey WR:
Effects of modulated VHF fields, on the central nervous system. Ann
N Y Acad Sci. 247:74–81. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Bawin SM, Adey WR and Sabbot IM: Ionic
factors in release of 45Ca2+ from chicken
cerebral tissue by electromagnetic fields. Proc Natl Acad Sci USA.
75:6314–6318. 1978. View Article : Google Scholar
|
|
153
|
Blackman CF, Benane SG, Elder JA, House
DE, Lampe JA and Faulk JM: Induction of calcium-ion efflux from
brain tissue by radiofrequency radiation: Effect of sample number
and modulation frequency on the power density window.
Bioelectromagnetics. 1:35–43. 1980. View Article : Google Scholar
|
|
154
|
Frei M, Jauchem J and Heinmets F:
Physiological effects of 2.8 GHz radio-frequency radiation: A
comparison of pulsed and continuous-wave radiation. J Microw Power
Electromagn Energy. 23:85–93. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Bolshakov MA and Alekseev SI: Bursting
responses of Lymnea neurons to microwave radiation.
Bioelectromagnetics. 13:119–129. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Penafiel LM, Litovitz T, Krause D, Desta A
and Mullins JM: Role of modulation on the effect of microwaves on
ornithine decarboxylase activity in L929 cells.
Bioelectromagnetics. 18:132–141. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Huber R, Treyer V, Borbély AA, Schuderer
J, Gottselig JM, Landolt HP, Werth E, Berthold T, Kuster N, Buck A
and Achermann P: Electromagnetic fields, such as those from mobile
phones, alter regional cerebral blood flow and sleep and waking
EEG. J Sleep Res. 11:289–295. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Höytö A, Luukkonen J, Juutilainen J and
Naarala J: Proliferation, oxidative stress and cell death in cells
exposed to 872 MHz radio-frequency radiation and oxidants. Radiat
Res. 170:235–243. 2008. View Article : Google Scholar
|
|
159
|
Mohammed HS, Fahmy HM, Radwan NM and
Elsayed AA: Non-thermal continuous and modulated electromagnetic
radiation fields effects on sleep EEG of rats. J Adv Res.
4:181–187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Noda M, Ikeda T, Kayano T, Suzuki H,
Takeshima H, Kurasaki M, Takahashi H and Numa S: Existence of
distinct sodium channel messenger RNAs in rat brain. Nature.
320:188–192. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Liman ER, Hess P, Weaver F and Koren G:
Voltage-sensing residues in the S4 region of a mammalian
K+ channel. Nature. 353:752–756. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Tombola F, Pathak MM and Isacoff EY: How
does voltage open an ion channel? Annu Rev Cell Dev Biol. 22:23–52.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Schmidt WF and Thomas CG: More precise
model of α-helix and transmembrane α-helical peptide backbone
structure. J Biophy Chem. 3:295–303. 2012. View Article : Google Scholar
|
|
164
|
Miller C: An overview of the potassium
channel family. Genome Biol. 1:REVIEWS00042000. View Article : Google Scholar
|
|
165
|
Zhang XC, Yang H, Liu Z and Sun F:
Thermodynamics of voltage-gated ion channels. Biophys Rep.
4:300–319. 2018. View Article : Google Scholar :
|
|
166
|
DeCoursey TE: Interactions between NADPH
oxidase and voltage-gated proton channels: Why electron transport
depends on proton transport. FEBS Lett. 555:57–61. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Seredenina T, Demaurex N and Krause KH:
Voltage-gated proton channels as novel drug targets: From NADPH
oxidase regulation to sperm biology. Antioxid Redox Sign.
23:490–513. 2015. View Article : Google Scholar
|
|
168
|
Panagopoulos DJ, Johansson O and Carlo GL:
Evaluation of specific absorption rate as a dosimetric quantity for
electromagnetic fields bioeffects. PLoS One. 8:e626632013.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Creasey WA and Goldberg RB: A new twist on
an old mechanism for EMF bioeffects? EMF Health Rep. 9:1–11.
2001.
|
|
170
|
Belyaev IY: Non-thermal biological effects
of microwaves. Microw Rev. 11:13–29. 2005.
|
|
171
|
Zhou R, Xiong Y, Xing G, Sun L and Ma J:
ZiFi: Wireless LAN discovery via ZigBee interference signatures.
MobiCom'10. In: September 20-24; Chicago, Illinois, USA. 2010
Proceedings of the 16th Annual International Conference on Mobile
Computing and Networking; View Article : Google Scholar
|
|
172
|
Piacentini R, Ripoli C, Mezzogori D,
Azzena GB and Grassi C: Extremely low-frequency electromagnetic
fields promote in vitro neurogenesis via upregulation of
Ca(v)1-channel activity. J Cell Physiol. 215:129–139. 2008.
View Article : Google Scholar
|
|
173
|
Cecchetto C, Maschietto M, Boccaccio P and
Vassanelli S: Electromagnetic field affects the voltage-dependent
potassium channel Kv1.3. Electromagn Biol Med. 39:316–322. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Zheng Y, Xia P, Dong L, Tian L and Xiong
C: Effects of modulation on sodium and potassium channel currents
by extremely low frequency electromagnetic fields stimulation on
hippocampal CA1 pyramidal cells. Electromagn Biol Med. 40:274–285.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Batcioglu K, Uyumlu AB, Satilmis B,
Yildirim B, Yucel N, Demirtas H, Onkal R, Guzel RM and Djamgoz MB:
Oxidative stress in the in vivo DMBA rat model of breast cancer:
Suppression by a voltage-gated sodium channel inhibitor (RS100642).
Basic Clin Pharmacol Toxicol. 111:137–141. 2012.PubMed/NCBI
|
|
176
|
Ramírez A, Vázquez-Sánchez AY,
Carrión-Robalino N and Camacho J: Ion channels and oxidative stress
as a potential link for the diagnosis or treatment of liver
diseases. Oxid Med Cell Longev. 2016:39287142016. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
O'Hare Doig RL, Chiha W, Giacci MK, Yates
NJ, Bartlett CA, Smith NM, Hodgetts SI, Harvey AR and Fitzgerald M:
Specific ion channels contribute to key elements of pathology
during secondary degeneration following neurotrauma. BMC Neurosci.
18:622017. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Akbarali HI: Oxidative stress and ion
channels. Systems Biology of Free Radicals and Antioxidants. Lahers
I: Springer; Berlin, Heidelberg: pp. 355–373. 2014, View Article : Google Scholar
|
|
179
|
Kourie JI: Interaction of reactive oxygen
species with ion transport mechanisms. Am J Physiol. 275:C1–C24.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Lushchak VI: Free radicals, reactive
oxygen species, oxidative stress and its classification. Chem Biol
Interact. 224:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Pacher P, Beckman JS and Liaudet L: Nitric
oxide and peroxynitrite in health and disease. Physiol Rev.
87:315–424. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar
|
|
183
|
Halliwell B: Biochemistry of oxidative
stress. Biochem Soc Trans. 35:1147–1150. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Balasubramanian B, Pogozelski WK and
Tullius TD: DNA strand breaking by the hydroxyl radical is governed
by the accessible surface areas of the hydrogen atoms of the DNA
backbone. Proc Natl Acad Sci USA. 95:9738–9743. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Cadet J, Delatour T, Douki T, Gasparutto
D, Pouget JP, Ravanat JL and Sauvaigo S: Hydroxyl radicals and DNA
base damage. Mutat Res. 1424:9–21. 1999. View Article : Google Scholar
|
|
186
|
Cadet J and Wagner JR: DNA base damage by
reactive oxygen species, oxidizing agents, and UV radiation. Cold
Spring Harb Perspect Biol. 5:a0125592013. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Tsunoda M, Sakaue T, Naito S, Sunami T,
Abe N, Ueno Y, Matsuda A and Takénaka A: Insights into the
structures of DNA damaged by hydroxyl radical: Crystal structures
of DNA duplexes containing 5-formyluracil. J Nucleic Acids.
2010:1072892010. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Cooke MS, Evans MD, Dizdaroglu M and Lunec
J: Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB
J. 17:1195–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Barzilai A and Yamamoto K: DNA damage
responses to oxidative stress. DNA Repair (Amst). 3:1109–1115.
2004. View Article : Google Scholar
|
|
190
|
Pall ML: Electromagnetic fields act via
activation of voltage-gated calcium channels to produce beneficial
or adverse effects. J Cell Mol Med. 17:958–965. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Liburdy RP: Calcium signaling in
lymphocytes and ELF fields. Evidence for an electric field metric
and a site of interaction involving the calcium ion channel. FEBS
Lett. 301:53–59. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Walleczek J: Electromagnetic field effects
on cells of the immune system: The role of calcium signaling. FASEB
J. 6:3177–3185. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Brookes PS, Yoon Y, Robotham JL, Anders MW
and Sheu SS: Calcium, ATP, and ROS: A mitochondrial love-hate
triangle. Am J Physiol Cell Physiol. 287:C817–C833. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Lang F, Föller M, Lang KS, Lang PA, Ritter
M, Gulbins E, Vereninov A and Huber SM: Ion channels in cell
proliferation and apoptotic cell death. J Membr Biol. 205:147–157.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Görlach A, Bertram K, Hudecova S and
Krizanova O: Calcium and ROS: A mutual interplay. Redox Biol.
6:260–271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Lombardi AA, Gibb AA, Arif E, Kolmetzky
DW, Tomar D, Luongo TS, Jadiya P, Murray EK, Lorkiewicz PK,
Hajnóczky G, et al: Mitochondrial calcium exchange links metabolism
with the epigenome to control cellular differentiation. Nat Commun.
10:45092019. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Ikwegbue PC, Masamba P, Oyinloye BE and
Kappo AP: Roles of heat shock proteins in apoptosis, oxidative
stress, human inflammatory diseases, and cancer. Pharmaceuticals
(Basel). 11:22017. View Article : Google Scholar
|
|
199
|
Becchetti A: Ion channels and transporters
in cancer. 1. Ion channels and cell proliferation in cancer. Am J
Physiol Cell Physiol. 301:C255–C265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Gaasch JA, Geldenhuys WJ, Lockman PR,
Allen DD and Van der Schyf CJ: Voltage-gated calcium channels
provide an alternate route for iron uptake in neuronal cell
cultures. Neurochem Res. 32:1686–1693. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Chattipakorn N, Kumfu S, Fucharoen S and
Chattipakorn S: Calcium channels and iron uptake into the heart.
World J Cardiol. 3:215–218. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Salsbury G, Cambridge EL, McIntyre Z,
Arends MJ, Karp NA, Isherwood C, Shannon C, Hooks Y; Sanger Mouse
Genetics Project; Ramirez-Solis R, et al: Disruption of the
potassium channel regulatory subunit KCNE2 causes iron-deficient
anemia. Exp Hematol. 42:1053–1058.e1. 2014. View Article : Google Scholar
|
|
203
|
Gamaley I, Augsten K and Berg H:
Electrostimulation of macrophage NADPH oxidase by modulated
high-frequency electromagnetic fields. Bioelectrochem Bioenerg.
38:415–418. 1995. View Article : Google Scholar
|
|
204
|
Panday A, Sahoo MK, Osorio D and Batra S:
NADPH oxidases: An overview from structure to innate
immunity-associated pathologies. Cell Mol Immunol. 12:5–23. 2015.
View Article : Google Scholar :
|
|
205
|
Henderson LM: NADPH oxidase subunit
gp91phox: A proton pathway. Protoplasma. 217:37–42. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Musset B, Cherny VV, Morgan D and
DeCoursey TE: The intimate and mysterious relationship between
proton channels and NADPH oxidase. FEBS Lett. 583:7–12. 2009.
View Article : Google Scholar :
|
|
207
|
DeCoursey T, Morgan D and Cherny V: The
voltage dependence of NADPH oxidase reveals why phagocytes need
proton channels. Nature. 422:531–534. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Friedman J, Kraus S, Hauptman Y, Schiff Y
and Seger R: Mechanism of short-term ERK activation by
electromagnetic fields at mobile phone frequencies. Biochem J.
405:559–568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Pratt RD, Brickman CR, Cottrill CL,
Shapiro JI and Liu J: The Na/K-ATPase signaling: from specific
ligands to general reactive oxygen species. Int J Mol Sci.
19:26002018. View Article : Google Scholar :
|
|
210
|
Nitahara JA, Cheng W, Liu Y, Li B, Leri A,
Li P, Mogul D, Gambert SR, Kajstura J and Anversa P: Intracellular
calcium, DNase activity and myocyte apoptosis in aging Fischer 344
rats. J Mol Cell Cardiol. 30:519–535. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Fenton HJH: Oxidation of tartaric acid in
presence of iron. J Chem Soc Trans. 65:899–911. 1894. View Article : Google Scholar
|
|
212
|
Bawin SM and Adey WR: Sensitivity of
calcium binding in cerebral tissue to weak environmental electric
fields oscillating at low frequency. Proc Natl Acad Sci USA.
73:1999–2003. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Jackson JD: Classical Electrodynamics. 2nd
edition. John Wiley & Sons, Inc; New York, NY: 1975
|
|
214
|
Barr R, Llanwyn Jones D and Rodger CJ: ELF
and VLF radio waves. J Atmos Sol-Terr Phys. 62:1689–1718. 2000.
View Article : Google Scholar
|
|
215
|
McLeod KJ, Lee RC and Ehrlich HP:
Frequency dependence of electric field modulation of fibroblast
protein synthesis. Science. 236:1465–1469. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Cleary SF, Liu LM, Graham R and Diegelmann
RF: Modulation of tendon fibroplasia by exogenous electric
currents. Bioelectromagnetics. 9:183–194. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Lee RC, Canaday DJ and Doong H: A review
of the biophysical basis for the clinical application of electric
fields in soft-tissue repair. J Burn Care Rehabil. 14:319–335.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Sandipan C and Baron C: Basic mechanisms
of voltage sensing. Handbook of Ion Channels. Zheng J and Trudeau
MC: CRC Press; London: pp. 25–39. 2015
|
|
219
|
Groome JR and Bayless-Edwards L: Roles for
countercharge in the voltage sensor domain of ion channels. Front
Pharmacol. 11:1602020. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Shi YP, Thouta S and Claydon TW:
Modulation of hERG K+ channel deactivation by voltage
sensor relaxation. Front Pharmacol. 11:1392020. View Article : Google Scholar
|
|
221
|
Villalba-Galea CA and Chiem AT: Hysteretic
behavior in voltage-gated channels. Front Pharmacol. 11:5795962020.
View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Panagopoulos DJ and Margaritis LH: The
identification of an intensity 'window' on the bioeffects of mobile
telephony radiation. Int J Radiat Biol. 86:358–366. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Panagopoulos DJ and Balmori A: On the
biophysical mechanism of sensing atmospheric discharges by living
organisms. Sci Total Environ. 599-600:2026–2034. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Liboff AR: Cyclotron resonance in membrane
transport. Interactions between Electromagnetic Fields and Cells.
Chiabrera A, Nicolini C and Schwan HP: Plenum Press; London: pp.
281–296. 1985
|
|
225
|
Bianco B, Chiabrera A, Morro A and Parodi
M: Effects of magnetic exposure on ions in electric fields.
Ferroelectrics. 86:159–168. 1988. View Article : Google Scholar
|
|
226
|
Balcavage WX, Alvager T, Swez J, Goff CW,
Fox MT, Abdullyava S and King MW: A mechanism for action of
extremely low frequency electromagnetic fields on biological
systems. Biochem Biophys Res Commun. 222:374–378. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Zhadin MN: Combined action of static and
alternating magnetic fields on ion motion in a macromolecule:
Theoretical aspects. Bioelectromagnetics. 19:279–292. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Liboff AR: Ion cyclotron resonance in
biological systems: Experimental evidence. Biological Effects of
Electromagnetic Fields. Stavroulakis P: Springer; Berlin: pp.
76–113. 2003
|
|
229
|
Lednev VV: Possible mechanism for the
influence of weak magnetic fields on biological systems.
Bioelectromagnetics. 12:71–75. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Kirschvink JL: Magnetite biomineralization
and geomagnetic sensitivity in higher animals: An update and
recommendations for future study. Bioelectromagnetics. 10:239–259.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Brocklehurst B and McLauchlan KA: Free
radical mechanism for the effects of environmental electromagnetic
fields on biological systems. Int J Radiat Biol. 69:3–24. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Sheppard AR, Swicord ML and Balzano Q:
Quantitative evaluations of mechanisms of radiofrequency
interactions with biological molecules and processes. Health Phys.
93:365–396. 2008. View Article : Google Scholar
|
|
233
|
Hall EJ and Giaccia AJ: Radiobiology for
the Radiologist. 6th edition. Lippincott Williams & Wilkins;
Philadelphia, PA: 2006
|
|
234
|
Coggle JE: Biological Effects of
Radiation. Taylor & Francis; 1983
|
|
235
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Blank M and Goodman R: Electromagnetic
fields stress living cells. Pathophysiology. 16:71–78. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Blank M and Goodman R: DNA is a fractal
antenna in electromagnetic fields. Int J Radiat Biol. 87:409–415.
2011. View Article : Google Scholar : PubMed/NCBI
|