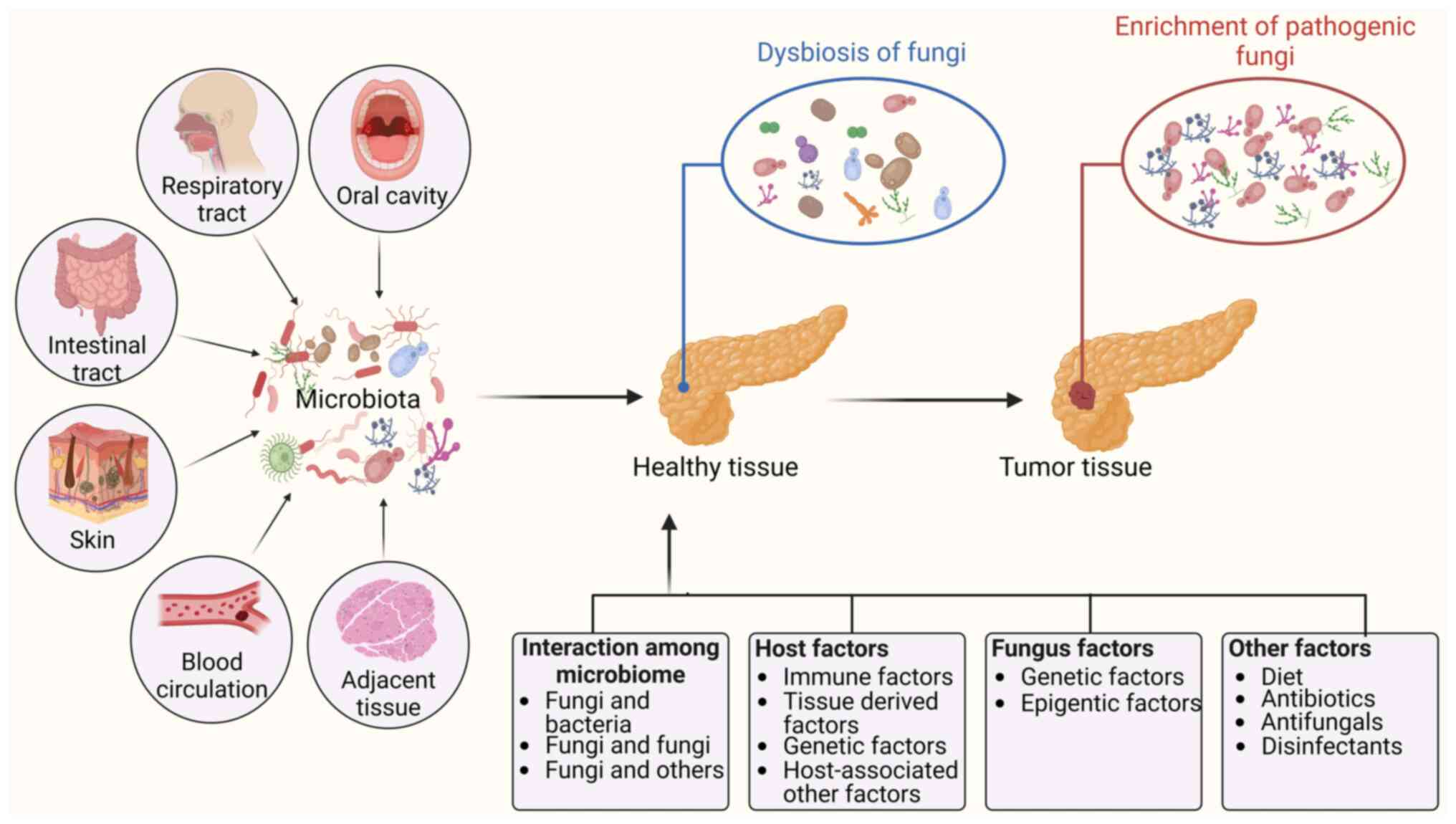
1. Introduction
Fungi are microeukaryotes that inhabit different
anatomic sites in the human body. More than 400 fungal species,
mainly including three phyla, Ascomycota,
Basidiomycota and Chytridiomycota, are associated
with the human body (1). Over 100
fungal species, including 50 genera, are also found in mice
(2). These fungi are less
abundant in the human microbiome than other organisms, such as
bacteria (3). However, emerging
evidence has shown that fungi significantly influence host health
and disease (3); for example,
fungi are involved in the occurrence and development of tumors
(4-7).
Tumors have complex ecosystems. They have their own
unique microbiome, which includes bacteria, viruses and fungi.
These intratumoral organisms participate in tumorigenesis and tumor
development (8,9). Most studies of microbial dysbiosis
in tumors, especially colorectal cancer (CRC), have focused on
bacteria (10). However,
sequencing technologies have also detected viruses, fungi and
archaea in tumor tissues and revealed cancer type-specific
microbial signatures (11). In
2022, Narunsky-Haziza et al (12) uncovered the fungal microbiome
atlas of 35 types of cancer and demonstrated that fungi were also
detected in all studied tumor types. Dohlman et al (13) also found tumor-related fungi in
cancers of the gastrointestinal (GI) tract, lung, breast and head
and neck by analyzing cancer genome data. Interestingly, different
cancers exhibit cancer type-specific fungal profiles, such as
Candida species, which are involved in the pathogenesis of
CRC (3,13). Notably, multi-kingdom microbiota
analyses have also provided biomarkers of CRC and bacterial-fungal
interactions (14). These
intratumoral fungi can be classified into six categories based on
different anatomic sites, including the oral cavity, gut, adjacent
tissue, lung, skin and blood circulation (15). Due to the significant enrichment
of specific fungi in malignant tumors, the associations between
fungi and human cancer have attracted increasing attention in
recent years (3).
Multiple factors, such as interactions between
bacteria and fungi, interactions between different fungi, and
interactions between fungi and host factors, fungal genetic
factors, and epigenetic factors, might be involved in the
enrichment of fungi in tumor tissues and/or the conversion of
commensal fungi to pathogenic fungi. Intratumoral fungi are
potential therapeutic target(s) and/or diagnostic and prognostic
indicators for tumors. These fungi are regulated by factors such as
diet, fecal microbiota transplantation (FMT), probiotics,
prebiotics and genetically engineered probiotics. In the present
review, the associations between fungi and human cancer, cancer
type-specific fungal profiles and the mechanisms by which fungi
induce tumorigenesis were discussed. Furthermore, the factors that
cause fungal enrichment in tumor tissues and/or the conversion of
commensal fungi to pathogenic fungi, as well as potential
therapeutic and preventive strategies based on intratumoral fungi
were summarized.
2. Signatures of fungal species in
tumors
Tumors are complicated ecosystems that are composed
of cancer cells, immune cells, fibroblasts, endothelial cells and
microbiota. The intratumoral microbiota is a novel and integral
tumor component, which includes bacteria, that was recently
identified in various cancer types. Poore et al (11) revealed cancer type-specific
microbial signatures in tumor tissue. Indeed, each type of tumor
has a distinct microbiota composition; for example, there is a
particularly rich and diverse microbiome in breast cancer (16). Recent findings have further
revealed the spatial and population heterogeneity of the
intratumoral microbiome (17).
These intratumoral microbiota can be used for multiple purposes,
such as distinguishing normal tissue from cancer tissue,
distinguishing metastatic cancers from non-metastatic cancers,
distinguishing patients with cancer that respond to drugs from
those that do not respond to drugs and distinguishing patients with
a favorable prognosis from those with a bad prognosis (18).
Interestingly, human tumor tissues also harbor
tumor-associated fungi (12,13). For example, Narunsky-Haziza et
al (12) reported that 31
fungi, such as Saccharomyces cerevisiae (99.7% coverage),
were present in analyzed tumor tissues. In support of this finding,
another study also revealed a high abundance and prevalence of
Saccharomycetales in different tumors (13). Other fungi, including Candida
albicans, Malassezia globosa, Malassezia
restricta and Blastomyces gilchristii, could also be
present in different types of human cancer (12,13,19). Indeed, fungi have been found in
multiple types of tumors (12),
such as those associated with CRC (13,20-22), pancreatic (23), breast (24), prostate (25), ovarian (26) and esophageal cancer (27). The signatures of the main specific
fungi in different tumors are illustrated in Fig. 1.
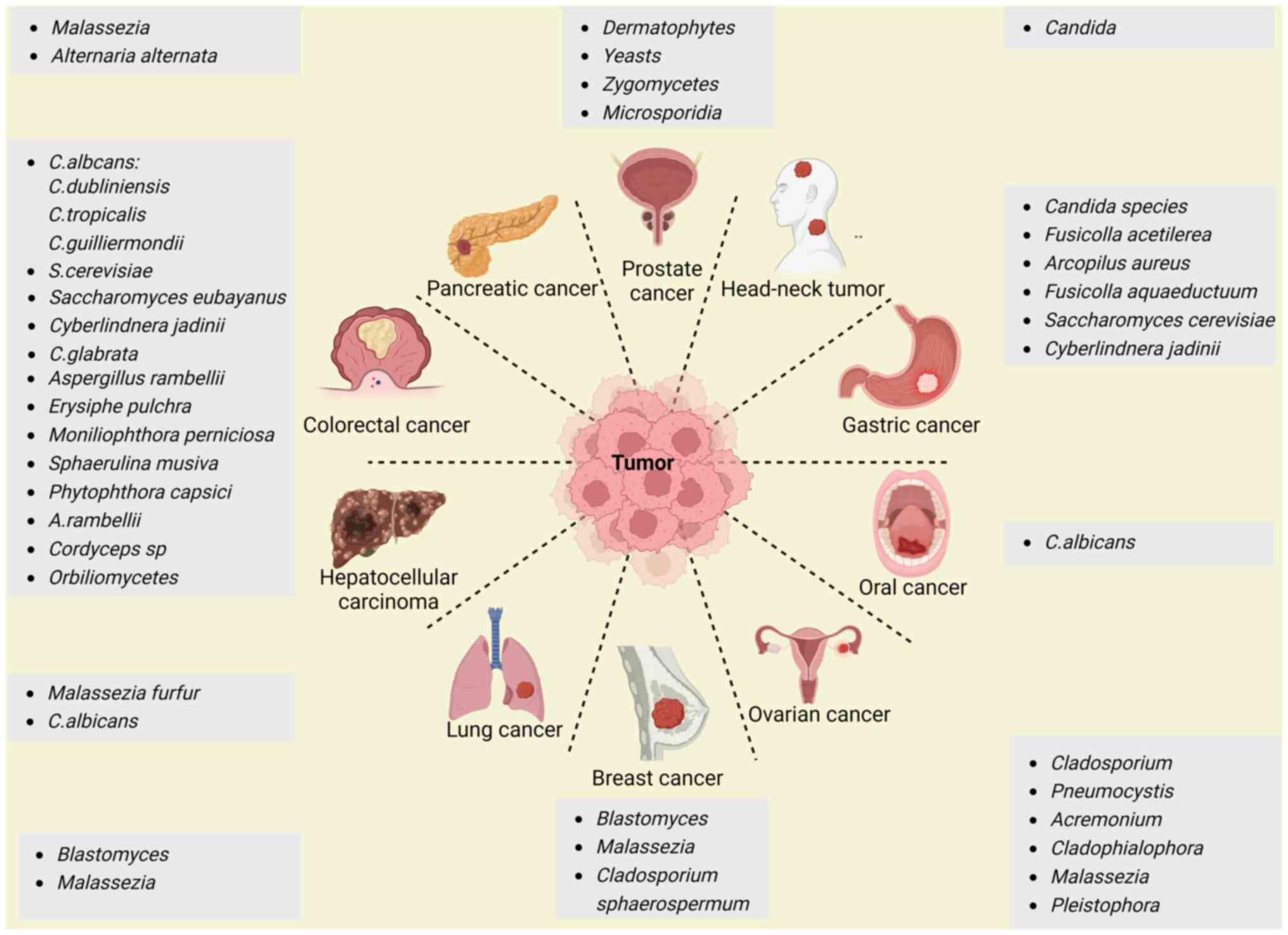 | Figure 1Signatures of specific fungi in
different tumors. Fungi can be detected in different tumor tissues,
including colorectal, hepatocellular, pancreatic, prostate, head
and neck, gastric, lung, oral, breast, lung and ovarian cancer
tissues. The figure was created with BioRender (https://servicecenter.oit.ucdenver.edu/CherwellPortal/IT). |
CRC
CRC is the fourth most common cancer worldwide and
is responsible for the deaths of >500,000 individuals every year
(28). Interestingly, CRC is
associated with changes in the fungal community of the colon in
patients (14,21,22). Fungal dysbiosis was detected in
patients with colorectal polyps (29) and adenomas (20), suggesting the involvement of fungi
in early-stage CRC. Indeed, there was a co-abundance group
associated with Candida albicans that included Candida
dubliniensis, Candida guilliermondii and Candida
tropicalis, and a group associated with Saccharomyces
cerevisiae, which included Saccharomyces eubayanus,
Cyberlindnera jadinii and Candida glabrata (13). These findings also indicated that
GI tract cancers may be separated into Candida- and
Saccharomyces-associated tumors (13). Notably, the abundance and
prevalence of the species Candida dubliniensis, Candida
glabrata, Candida guilliermondii, Candida lusitaniae, Candida
parapsilosis, Candida tropicalis and Pichia
membranifaciens were also lower in CRC according to a
metagenomic analysis of whole-genome sequencing (WGS) data from
multiple tumor samples from patients with different cancers in The
Cancer Genome Atlas (13).
Several other studies also indicated the existence of
Candida species, Cyberlindnera jadinii and
Saccharomyces cerevisiae in CRC tissues (30-32). However, a previous study also
revealed that Aspergillus species were highly enriched in
the CRC tissues of patients from both Asia and Europe through fecal
shotgun metagenomic sequencing (22). In addition, other fungi, such as
Cordyceps sp. RAO-2017, were also detected in CRC
tissues (21). The abundance of
Orbiliomycetes was different in the CRC and polyp groups
(29).
Gastric cancer (GC)
GC, which is one of the most common malignancies and
one of the main causes of tumor-associated deaths worldwide, is
also related to fungi (33,34). A metagenomic analysis of WGS data
revealed that several fungi, such as Candida species,
Saccharomyces cerevisiae and Cyberlindnera jadinii,
were highly abundant in the mycobiome communities of patients with
GI tract cancer (13). A
different study by internal transcribed spacer 2 (ITS2) analysis of
GC tissues revealed significant increases in the abundance of
Candida albicans, Fusicolla acetilerea, Arcopilus
aureus and Fusicolla aqueductuum in cancer lesions and
adjacent non-cancerous tissues of 45 patients with GC from
Shenyang, China (35). Notably,
the abundances of other fungi, such as Aspergillus
montevidensis and Candida glabrata, were markedly
reduced (35,36). Increased Candida abundance was
also linked to the expression of proinflammatory factors, which
could lead to the occurrence and development of tumors (13). Notably, Candida albicans
might also cause GC by decreasing the diversity and richness of
fungi in the stomach, which contributes to the pathogenesis of GC
(35).
Hepatocellular carcinoma (HCC)
Using ITS2 rDNA sequencing, alpha diversity analyses
revealed that patients with HCC had reduced fungal diversity when
compared with controls (37).
Aberrant colonization of the gut by Candida albicans and
Malassezia furfur promoted the occurrence and development of
HCC (37). HCC tumor weight and
volume significantly increased in the Candida albicans and
Malassezia furfur groups compared with the control group
(37).
Pancreatic cancer
Pancreatic cancer, which is one of the leading
causes of cancer-related deaths, is also associated with fungi. A
recent preclinical and clinical study demonstrated that pancreatic
ductal adenocarcinomas (PDACs) harbored significant enrichment of a
specific fungus in mouse models and human specimens. Indeed,
enriched fungi were observed in the pancreas of patients with PDAC
and in mouse models of pancreatic cancer by principal coordinate
analysis (23). Malassezia
species were more prevalent in PDAC tissues in both mice and humans
(23). Another analysis also
demonstrated that Malassezia and Alternaria were the
most abundant fungi in PDAC tumors using 18S rRNA sequencing
(5). Significantly high levels of
fungal and bacterial alpha diversity in the gut were also observed
in patients with PDAC by 16S rRNA gene sequencing (38). Bacteria and fungi can be
translocated to the pancreas and induce local and systemic changes
to promote the development of PDAC (39). GFP-labeled Saccharomyces
cerevisiae was detected in the pancreas of mice within 30 min
of consumption (23).
Ovarian cancer
Significant differences in the abundances of
Cladosporium, Pneumocystis, Acremonium,
Cladophialophora, Malassezia and Pleistophora
were detected in all the ovarian cancer samples. Rhizomucor,
Rhodotorula, Alternaria and Geotrichum were
also associated with >95% of the ovarian cancer samples
according to a pan-pathogen array (PathoChip) combined with
capture-next generation sequencing (26).
Prostate cancer
A fungal signature was observed in prostate cancer
samples when compared with benign prostate hyperplasia samples
(25). Dermatophytes
(31%), yeasts (15%), Zygomycetes (15%) and
Microsporidia (12%) were detected in the analyzed samples
(25). The majority of fungal
signatures were from the Ascomycota phylum (61%), but 50% of
the fungi belonged to the class Eurotiomycetes according to
hierarchical clustering analysis (25).
Breast cancer
A study revealed that Blastomyces and
Malassezia species were abundant in breast tumors (13). ITS2 amplicon sequencing revealed
that Cladosporium was enriched in patients with breast
cancer who were ≥50 years old (12). Cladosporium was also
enriched in human epidermal growth factor receptor 2-negative
tumors (12). Malassezia
restricta, another skin fungus, was also present in breast
cancer samples (12). In
addition, 7, 8 and 14% of the total hybridization signals for
Ajellomyces were endocrine receptor-positive, endocrine
receptor triple-positive and endocrine receptor 2-positive breast
cancer, respectively, whereas Rhizomucor accounted for 19%
of the hybridization signals for endocrine receptor triple-negative
breast cancer (24).
Lung cancer
Blastomyces and Malassezia are
associated with lung cancer (13); for instance, Blastomyces
DNA was detected in 6 out of 50 patients with squamous cell lung
carcinomas via metagenomic analysis of WGS data (13). Greater fungal diversity and a more
complex network was also found in patients with non-small cell lung
cancer (12,40).
Other tumors
A metagenomic analysis of WGS data revealed that
Candida is related to head and neck tumors (13). Using Illumina™ 2×300 bp chemistry,
Candida albicans was revealed to play a role in the
occurrence and development of oral cancer (OC) based on the fungal
ITS2 region (41).
3. Fungal-associated factors that lead to
cancer
Numerous studies have shown that some specific fungi
play important roles in the promotion, progression and recurrence
of cancers. These fungi modulate the immune system (42), stimulate the production of
specific metabolites (43,44)
and potentially reconstruct different microenvironments such as
biofilms. All of these factors affect not only immunity against
tumors but also the genome, transcriptome, epigenome,
epi-transcriptome, proteome and metabolome of tumor cells.
Immune factors
Cancers are related to fungus-mediated immune
responses. Different intratumoral microbiome interactions may cause
different immune responses in host tumor tissues. One study
revealed three distinct clusters in tumors, termed mycotypes F1
(Malassezia-Ramularia-Trichosporon), F2
(Aspergillus-Candida) and F3 (multiple genera, including
Yarrowia), which could discriminate the types of immune
response, suggesting that these intratumoral mycobiomes could
elicit different host responses (12). Tumors enriched with the F1 and F2
mycotypes were enriched in tumor suppressing inflammatory responses
across 20 types of cancer (12).
A previous study has also shown that the cell wall components of
Candida guilliermondii, Candida krusei, Candida
tropicalis, Candida auris and Candida albicans
can trigger different types of recognition by innate immune cells
in humans (42). A different
study revealed the multiple mechanisms by which fungal-mediated
immune factors can lead to the occurrence and development of
cancers (Fig. 2). However, the
aforementioned study did not examine inflammatory markers, such as
C-reactive protein and albumin levels; neutrophil, lymphocyte and
white blood cell counts; or the neutrophil/lymphocyte ratio, which
are associated with tumor size and tissue grade in fungi-mediated
tumors (45).
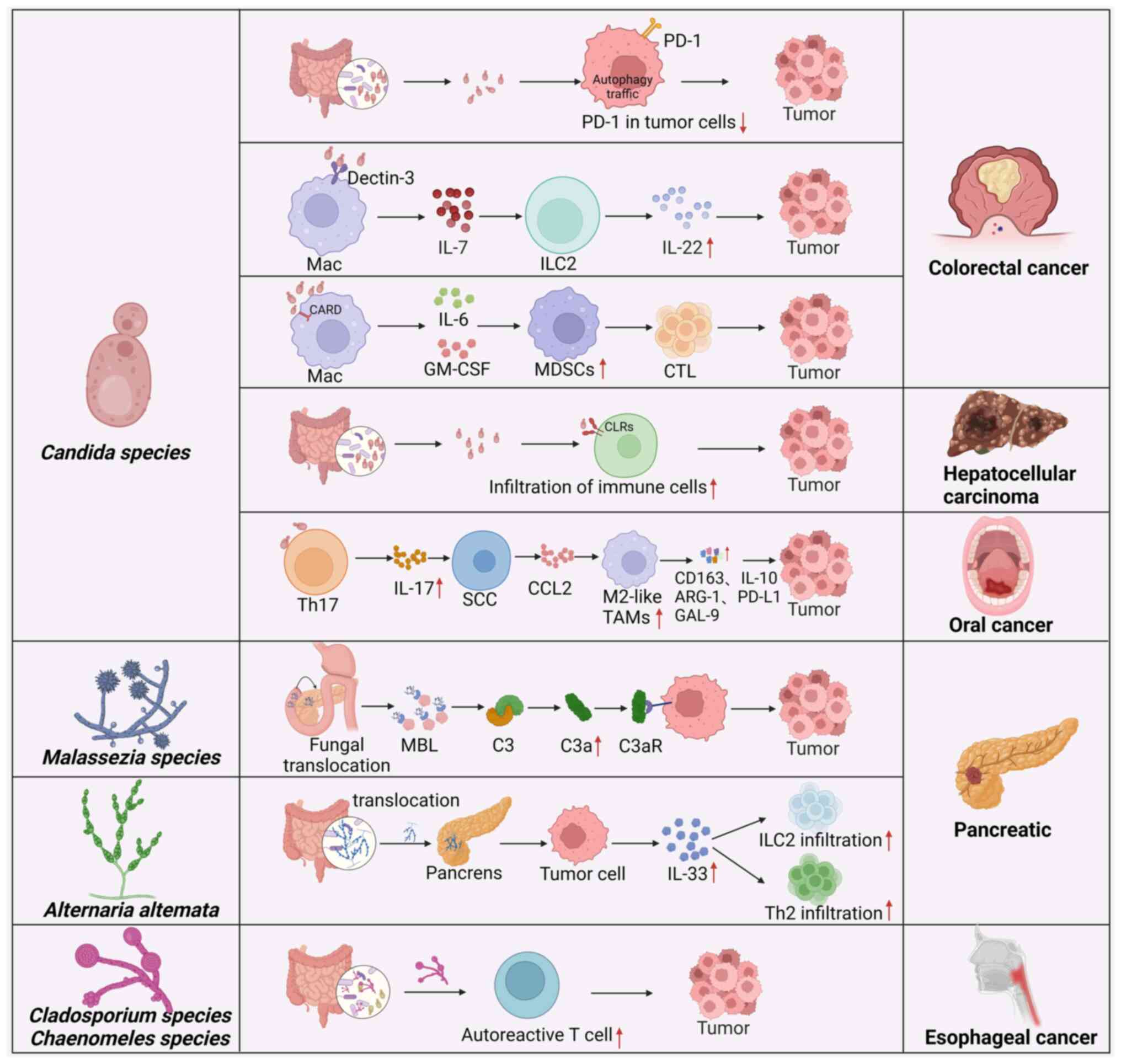 | Figure 2Fungal-associated immune factors that
lead to carcinogenesis. The interactions of fungi with immune cells
or tumor cells can increase or reduce the number of different
immune cell populations and the level of immune cytokines, which
can directly and indirectly affect carcinogenesis. MDSCs,
myeloid-derived suppressor cells; PD-1, programmed cell death
protein 1; Mac, macrophage; IL, interleukin; ILC2, innate
lymphocyte 2; CARD, caspase recruitment domain; GM-CSF,
granulocyte-macrophage colony-stimulating factor; CTL, cytotoxic T
lymphocyte; CLRs, C-type lectins; Th, T helper; SCC, squamous cell
carcinoma; CCL2, C-C motif chemokine ligand 2; TAM,
tumor-associated macrophage; ARG-1, arginase-1; GAL-9, galentin-9;
MBL, mannose-binding lectin; C3aR, C3a receptor. The figure was
created with BioRender. |
CRC
Myeloid-derived suppressor cells (MDSCs) are
immunosuppressive cells that promote the occurrence and development
of tumors. Fungal dysbiosis can increase the abundance of MDSCs,
which contribute to the development of CRC. Fungal overgrowth led
to the accumulation of MDSCs in the colon and worsened CRC in
caspase recruitment domain 9 (CARD9)−/−
mice. Treatment with the antifungal drug fluconazole suppressed CRC
in CARD9−/− mice, which was associated with
reduced MDSC accumulation (4).
CARD9 expressed in immune cells participates in innate and adaptive
immune responses via interactions between CARD9 and other molecules
(46). A previous study has
reported that CARD9 promotes colitis-associated cancer (47). Mutations in CARD9 are strongly
associated with increased susceptibility to both fungal infections
and inflammatory bowel disease in humans (48). Interestingly, when bone marrow
cells were cocultured with Candida tropicalis, Candida
tropicalis promoted the differentiation and function of MDSCs.
In germ-free mice mono-colonized with Candida tropicalis,
there was also an abundance of MDSCs in the colon (4). Further studies demonstrated that gut
fungi promoted the immunosuppressive function of MDSCs by pyruvate
kinase M1/2-dependent glycolysis, which promoted colorectal
tumorigenesis (32). Multiple
studies have reported that aerobic glycolysis is essential for
MDSCs in tumors (49,50). To maintain immunosuppressive
activities, MDSCs in tumors increase the level of glycolysis.
Notably, MDSCs are able to absorb intratumoral glucose in the tumor
microenvironment (TME) (51).
However, Malik et al (6)
reported that the fungal-mediated signaling axis, which is mediated
by CARD9 and its upstream activator spleen tyrosine kinase (SYK),
could also hinder CRC development by inducing inflammasome
activation. Deletion of CARD9 or SYK in MDSCs inhibited
inflammasome activation and interleukin (IL)-18 maturation and
enhanced susceptibility to CRC after fungal exposure (6). Supplementation with MDSCs or IL-18
decreased the tumor burden in azoxymethane/dextran sulfate sodium
(AOM/DSS)-treated CARD9−/− and
SYKfl/flLysMCre/+ mice,
whereas antifungal agents promoted colitis and CRC development
(6).
In addition, Candida albicans can trigger
glycolysis in macrophages and induce the production of IL-7, which
causes the secretion of IL-22 in RAR-related orphan receptor gamma
t innate lymphoid cells (ILCs) via the aryl hydrocarbon receptor
and signal transducer and activator of transcription 3 to promote
the progression of CRC (52). A
previous study also demonstrated that the development of Candida
tropicalis-mediated CRC involved reducing tumor cell-intrinsic
programmed cell death protein 1 (PD-1) levels through autophagy
(7). Autophagy inhibitors and
Candida tropicalis treatment can limit CRC tumor growth and
reverse downregulation of PD-1 expression. This finding suggested
that Candida tropicalis can promote CRC progression by
controlling the expression of PD-1 on tumor cells (7).
Pancreatic cancer
Analysis of PDAC revealed that Alternaria
alternata, but not Candida or Aspergillus, led to
the secretion of IL-33 in tumors, thereby promoting the recruitment
of type 2 immune cells to promote tumor development (5). Indeed, single-cell analyses of
CD45+ cells from a mouse model of pancreatic cancer
revealed the presence of T helper 2 cells (TH2) and ILC2 cells,
which were hallmarks of type II immune responses (5). Genetic deletion of IL-33 or
antifungal treatment decreased TH2 and ILC2 infiltration and
increased survival in mice. IL-33 knockdown in tumor cells in an
orthotopic model demonstrated that reduced IL-33 levels decreased
the infiltration of type 2 immune cells and tumor growth. Treatment
with the antifungal drug amphotericin B or IL-33 depletion caused a
significant decrease in tumor burden, increased survival and
reduced the number of tumor-infiltrating ILC2 and TH2 cells. TH2
cells, which infiltrate the pancreas in the early stages of
tumorigenesis, can produce type 2 cytokines such as IL-4 and IL-13,
which promote the metabolic reprogramming of cancer cells in murine
KrasG12D-driven PDAC. Consistent with type 2
immune responses that induce PDAC progression in mouse models,
patients with PDAC with predominant TH2-polarized cell infiltration
also exhibited reduced survival compared with patients with more
TH1 cells (53). Notably, ILC2s
are also present in tumors from patients with pancreatic cancer
(5), and high IL-33 expression is
observed in ~20% of human patients with PDAC (5).
However, the fungal community in PDAC was markedly
enriched in Malassezia species in both mice and humans
(23). The ligated product of
mannose-binding lectin (MBL) can bind to glycans in the fungal wall
to activate the complement cascade, thus causing an increase in
C3a. Subsequently, C3a can bind to C3a receptor (C3aR) on the
surface of tumor cells to promote tumor proliferation, motility and
invasion (23). Indeed, MBL or C3
deletion in the extratumoral compartment or knockdown of the C3aR
in tumor cells protected against tumor growth (23). Notably, Malassezia-mediated
oncogenic progression was delayed in mice lacking MBL (23). Mice that were treated with
antifungal drugs and colonized with Malassezia globosa had
larger tumors. Increased levels of Malassezia were observed
in human pancreatic cancer samples (23).
Esophageal cancer
Autoreactive T cells and chronic fungal infection
cause esophageal carcinogenesis (27). Ikkα knock-in
(IkkαKA/KA) mice develop impaired central
tolerance, autoinflammation, chronic fungal infection and
esophageal squamous cell carcinoma (ESCC) (27). Interestingly, during this process,
autoreactive CD4+ T cells are generated, which permit
fungal infection and cause tissue injury and inflammation.
Antifungal treatment or the depletion of autoreactive
CD4+ T cells could rescue ESCC development, whereas oral
fungal administration promoted ESCC development. Thus, autoreactive
T cells and chronic fungal infection promote ESCC development
(27). Cladosporium
cladosporioides and Chaenomeles lagenaria, which are two
major fungal species that colonize the oral cavities and esophagi
of IkkαKA/KA mice, might spread from the oral
cavity to the esophagus. Notably, fungal infection is highly
related to ESCC in non-autoimmune patients (27).
OC
Candida albicans promoted OC via
IL-17A/IL-17RA and macrophages (54). IL-17A neutralization and
macrophage depletion reduced the number of tumor-associated
macrophages and tumor size in mice with Candida albicans
infection (54). Mechanistically,
Candida albicans infection promoted IL-17A production by
Th17 cells. Following activation of the IL-17RA signal, tumor cells
can release C-C motif chemokine ligand 2 to attract macrophages to
the TME, and these macrophages exhibit an immunosuppressive
phenotype with upregulated expression of IL-10, arginase-1, PD-L1
and galectin-9.
HCC
The expression of fungal recognition receptors
C-type lectins (CLRs), such as dectin-1, dectin-2 and dectin-3, is
downregulated in HCC. The expression of these genes is related to
the clinical prognosis of patients with HCC (55). CLR-triggered immune responses
might enhance the effects of immunotherapy against HCC (55). The expression of CLRs was
significantly related to immune infiltration and immunotherapy
efficacy in HCC.
Notably, there is still absence of evidence on the
role of fungi in renal cell carcinoma (RCC). Since RCC is heavily
infiltrated by T cells and myeloid cells (56), future studies should first solve
whether fungi infection is related to the infiltration of T cells
and myeloid cells in the occurrence and development of RCC.
Metabolites and toxins
Toxins and bioactivated amines from fungi have been
linked to carcinogenesis (43,44). These factors may cause genetic,
epigenetic and metabolic changes. For example, Candida
albicans generates nitrosamine and metabolizes ethanol to
acetaldehyde (57), which is an
electrophilic and genotoxic substance that affects DNA repair,
oxidative stress, DNA damage and gene mutations (58). The fungus-associated metabolite
aflatoxin B1 that is produced by the Aspergillus species can
induce the development of HCC via highly mutagenic DNA (59). Additionally, interactions between
bacteria and fungi can also induce colorectal carcinogenesis by
activating butanoate metabolism (14). Two marker genes, oraS and
oraE, in the D-arginine metabolism pathway were
significantly enhanced in CRC samples (14). Differential abundance analyses of
the mycobiome also suggested that increased Candida
abundance could promote metastasis, cellar adhesion, extracellular
matrix-receptor interactions and focal adhesion (19).
Biofilms
Another possible mechanism by which the microbiota
affects tumorigenesis is the formation of biofilms (60). Candida albicans can
cooperate with bacteria such as E. faecalis to produce
biofilms. Biofilms are closely related to CRC based on the
enhancement of precancerous inflammation and escaping the host
immune response (61).
Interestingly, biofilm homogenates from patients with CRC can cause
colon tumorigenesis in mice (62).
Fungal extracellular vesicles (EVs)
Fungal EVs can be isolated from yeast and
filamentous fungi. The pathogenic role of fungal EVs has been
widely reviewed (63-65). They carry pigments, carbohydrates,
proteins, nucleic acids, lipids and prions, which modulate the
immune responses of host cells and are tightly related to virulence
(63). Furthermore, EVs play
pivotal roles in orchestrating fungal communities, bolstering
pathogenicity and mediating interactions with the environment
(64,66). EVs from Candida albicans
and Saccharomyces brasiliensis activate dendritic cells to
produce cytokines such as IL-12p40, IFN-γ, TNF-α, IL-10 and TGF-β
(67). EVs from pathogenic fungi
also promote the production of TNF-α, TGF-β and nitric oxide by
macrophages (66,68). Exophiala dermatitidis EVs
could induce cell death. Understanding the function of fungal EVs
can provide new and specific targets for antifungal drugs. However,
there is lack of studies on the effects of fungal EVs on
tumorigenesis.
4. Factors related to the enrichment and
carcinogenicity of fungal species
Intratumoral fungi can come from different anatomic
sites, including the oral cavity, the gut, adjacent normal tissue,
the lung, skin and blood circulation (15). Multiple factors are potentially
related to the enrichment and carcinogenicity of fungal species,
including interactions between microbes such as fungi and bacteria,
host factors including immune factors, tissue-derived factors, and
fungal genetic and epigenetic factors (Fig. 3). Notably, fungi not only are the
causative agents of diseases but are also isolated from mammals
without diseases (69-71), suggesting that there are two
fungal types, namely, commensal and pathogenic fungi (72). Indeed, the pathogenicity of some
fungi depends on their ability to change from a commensal to a
pathogenic fungus (73). Li et
al (74) reported that
Candida albicans can aggravate intestinal inflammation by
inducing proinflammatory phenotypes in vivo.
Interactions among the microbiome
Multiple different kinds of microbiota exist in the
organs and tissues of humans, such as the gut. These organisms live
together and form complex and dynamic ecosystems to impact host
health (75). Multiple kingdom
analyses of fecal samples from patients with CRC revealed strong
interkingdom interactions between bacteria and fungi (14,22). A different study also revealed
four kingdom microbiota alterations using metagenomic datasets from
1,368 CRC samples from 8 distinct geographic cohorts. The
researchers found not only significant fungal-bacterial
interactions between Aspergillus rambellii and
Fusobacterium nucleatum but also significant interactions
between Aspergillus rambellii and Parvimonas micra in
both patients with CRC and patients with adenoma (14,22). The signature of CRC-associated
fungi included 6 different enriched fungi, namely, Aspergillus
rambellii, Cordyceps sp. RAO-2017, Erysiphe pulchra, Moniliophthora
perniciosa, Sphaerulina musiva and Phytophthora capsici.
Aspergillus rambellii is closely related to the CRC-enriched
bacterium Fusobacterium nucleatum (21). Notably, experimental studies have
demonstrated interactions between fungi and bacteria. For example,
Lactobacillus can produce metabolites to antagonize
Candida albicans growth and filamentation (76,77). Reductions in short-chain fatty
acid (SCFA) levels in the murine gut were associated with an
increase in Candida albicans (78). The SCFAs butyrate and propionate
also inhibited the growth of the yeast Pichia kudriavzevii
(79). Negative correlations
between Penicillium and Faecalibacterium were found
in the human gut (80). In
addition, bacterium-induced immunity could also limit Candida
albicans colonization of the gut lumen. Anaerobic bacteria
promoted the expression of cathelicidin-related antimicrobial
peptide, which can eliminate Candida albicans (78). Lactobacillus exhibits an
enhanced probiotic potential following coculture with
Kluyveromyces marxianus (81). Notably, bacteria-fungi
interactions have revealed that bacteria can shape the immune
environment that controls fungi (12). Lactobacillus
kefiranofaciens and Saccharomyces cerevisiae isolated
from Tibetan kefir grain alleviated AOM/DSS mediated inflammation
and colorectal carcinogenesis (82). Interestingly, the presence of
Candida and Saccharomyces was associated with
different Fusobacterium spp. in colon cancer (13). In stomach cancer, Candida
was positively associated with Dialister abundance and
negatively associated with Akkermansia municiphila,
Ruminococcus and Barnesiella intestinihominis abundance
(13).
In addition, the fungal community also affects
bacteria. Candida albicans has been shown to antagonize
colonization by Escherichia and Klebsiella species.
Cocolonization experiments in mice confirmed that Candida
albicans could limit Klebsiella colonization in the gut
(83). Lactobacillus spp.,
especially Lactobacillus gasseri, are frequently found in
the presence of Candida and Saccharomyces (13). This observation was consistent
with studies reporting that the interaction between
Lactobacillus spp. and Candida influences
pathogenicity (76).
Candida was strongly associated with Lactobacillus in
GC (13). In head and neck
tumors, Candida and Saccharomyces are related to
similar bacteria, such as Bifidobacterium, which support
intestinal barrier function in head and neck cancers (13). Fungal dysbiosis with an increased
Basidiomycota: Ascomycota ratio was observed in the
feces of patients with CRC (22),
implying that interactions between bacteria and fungi could
contribute to colorectal carcinogenesis (22,84).
Host factors
Host factors including tissue-derived, genetic,
immune and other factors can affect the enrichment and/or
conversion of fungi from a commensal state to a pathogenic state.
There have been several reviews on fungal immunity (85,86) and the correlations between immune
responses and genetics (86).
Notably, tissue-derived factors were found to affect fungi such as
Candida auris, which led to the observation of
subpopulations of aggregative and filamentous isolates in some
clinical studies (72). Host
genetic factors are also related to the transition of fungi from a
commensal to a pathogenic fungus. Typically,
Dectin-3−/− mice exhibited an increase in
pathogenic Candida albicans (52).
Fungal genetic and epigenetic
factors
Multiple fungal genetic and epigenetic factors,
which are related to the enrichment and carcinogenicity of fungal
species, such as ume6, which is a master regulator from yeast to
hyphae Candida albicans, can suppress gut colonization by
promoting the expression of the hypha-specific proinflammatory
protease secreted aspartic protease 6 and the hyphal cell surface
adhesion protein glutathione peroxidase-like peroxiredoxin HYR1
(87). Candida albicans in
the gut causes a developmental switch of the white-opaque regulator
1 transcription factor, which leads to a commensal cell type
(88). Fungi can also regulate
iron uptake genes via Sef1/Sfu1, which play a role in fungal
virulence and colonization (89).
Candida auris also activates a stress response program via
mitogen-activated protein kinase HOG1, which is necessary for
virulence (90). Notably,
Candida species can generate numerous more phospholipases
than other fungal strains (91).
In intestinal inflammation, Candida can produce
candidalysin, which induces damage to cause hyphal invasion across
mucosal barriers (92).
Additionally, set1-mediated H3K4 methylation was required for
Candida albicans virulence based on controlling reactive
oxygen species levels. Candida auris also modulates genome
integrity, stress responses, cell filamentation and virulence via
the lncRNA DINOR (93).
Other factors
Other host factors, such as diet and age, can
affect the variability of the gut mycobiota (94-96). Antibiotics, antifungals and
disinfectants also affect the enrichment of fungi and/or the
conversion of fungi from a commensal state to a pathogenic state.
For example, antibiotics can lead to an increase in Candida
in the gut, oral cavity and vagina (97,98), which facilitates invasive fungal
infection through bloodstream translocation from the gut (99).
5. Application of intratumoral fungi in the
diagnosis and treatment of cancers
Potential therapeutic targets
Fungi can be engineered to enhance their effects on
the occurrence and development of tumors. Furthermore, intratumoral
fungi can also induce innate and adaptive immune responses to
prevent tumor progression (6,100). Fungi, such as Capnodiales
and its genus Cladosporium, which are significantly enriched
in non-responders, are also associated with immunotherapy response
in patients with metastatic melanoma (12). Thus, fungi in tumor tissues might
be a new potential therapeutic target in cancer therapy. At
present, other microbiota, such as bacteria, have been approved by
the Food and Drug Administration for the treatment of cancer
(101,102).
Diagnosis and prognosis evaluation
Several studies have also reported the role of
intratumoral microorganisms in diagnosis (103,104). Due to the presence of tumor
type- and subtype-specific fungal profiles, intratumoral fungi have
the potential to be used as diagnostic tools. However, whether
fungi can be used for diagnosis has not been determined. In
addition, the tumor microbiome is related to the survival rates of
different patients. The presence of some intratumoral fungi may be
closely related to the poor prognosis of patients with tumors. For
example, in GI tumors, the presence of Candida DNA is
predictive of decreased survival. Narunsky-Haziza et al
(12) also suggested that fungi
have prognostic and diagnostic roles in tumor tissues by comparing
intratumoral fungal communities with matched bacteriomes and
immunomes. The associations of fungi with clinical parameters such
as the detection of early-stage cancers, overall survival in breast
cancer patients and immunotherapy response in melanoma patients
supported the clinical application of fungi as potential biomarkers
and therapeutic targets (12).
Strategies to modulate the fungal
community
Multiple therapeutic strategies targeting the
microbiota (105,106), such as antifungal drugs, have
been used to inhibit the oncogenic progression of PDAC. As some
specific fungal species are related to the occurrence and
development of tumors, antifungal chemical compounds such as
terbinafine, fluconazole and itraconazole could be used for tumor
therapy. The combination of an antifungal drug and chemotherapy
exhibited a synergistic anticancer effect against PDAC in animal
models (3). Notably, broad
antibiotic application also increased the risk of cancer incidence
and impaired the response to immunotherapy (107).
Specific modulation of intratumoral fungi in the
clinical practice is challenging. However, the factors that
regulate the gut fungal community are also potential tools for
therapy against tumors.
Diet
Diet and nutrition can affect the composition of
the gut microbiota and are involved in CRC onset (108). Diet-induced changes in the gut
microbiome depend on whether volunteers consume a plant- or
animal-based diet (109).
FMT
FMT can regulate the composition of fungi to affect
tumor therapy efficacy. A high abundance of Saccharomyces
and Aspergillus in donor stool was associated with effective
FMT, whereas reduced FMT efficacy was related to an increase in
Candida albicans in donor stool. Further study revealed that
Candida was negatively correlated with total saturated fatty
acids and positively correlated with carbohydrates, whereas
Aspergillus was negatively correlated with the recent
ingestion of SCFAs. These metabolites could directly and indirectly
affect the therapeutic effectiveness of FMT against tumors.
Probiotics and prebiotics
Several functions of probiotics, such as the
suppression of pathogen growth by the production of certain
antimicrobial mediators (110),
have been reported. Prebiotics can prevent CRC development by
modifying the composition of the gut microbiota (111) and exert strong preventive
effects against CRC. Notably, Saccharomyces cerevisiae plays
a probiotic role in CRC by promoting cancer cell apoptosis.
Saccharomyces cerevisiae reduces CRC progression by
modulating the microbial structure in the mucus (31). In addition, genetically engineered
microbiota could also be used as a vehicle to provide metabolic
support for intratumoral T cells (112), which is essential for the proper
functioning of cytotoxic T cells (113).
6. Conclusion and perspective
Omics analyses of host-microbiome interactions in
human health and diseases have revealed associations between fungi
and human cancer. Several cancer type-specific fungi have been
identified, such as Candida species in CRC,
Malassezia species in pancreatic cancer and
Blastomyces species in lung and breast cancer. Importantly,
some specific fungal species that lead to the occurrence and
development of tumors, such as Candida species, induce CRC
through the accumulation of MDSCs, and Malassezia species
promote pancreatic oncogenesis by activating the complement
cascade. In addition, multiple factors, such as interactions among
the microbiome, are related to the enrichment of type-specific
fungi in tumor tissues and/or conversion from a commensal to a
pathogenic fungus. A growing body of evidence has revealed the
diagnostic, prognostic and therapeutic potential of intratumoral
fungi in cancer. Fungal dysbiosis in the gut can be regulated by
multiple factors, such as diet, FMT, probiotics and prebiotics,
which potentially affect tumor development.
However, these studies are just a start for
studying intratumoral fungi, and numerous questions remain to be
answered: i) What determines the abundance and composition of
intratumoral fungi? Studies have shown that there is abundance of
fungi in tumor tissues. Furthermore, the composition of fungi in
different tumor is also different. At present, it is unclear what
determines the abundance and composition of intratumoral fungi. ii)
What are the origins of the intratumoral fungi? Fungi can be found
not only in colorectal carcinoma but also in other tumors, such as
those associated with prostate, ovarian and breast cancer. But, the
origins of these intratumoral fungi are incompletely clear. iii)
How do intratumoral fungi bridge cancer cells and the immune
system? Fungi-mediated immune factors play important roles in
tumorigenesis. It is also incompletely clear how these intratumoral
fungi bridge cancer and immune cells. iv) What exact mechanism(s)
do specific fungi use to induce tumorigenesis? There are different
mechanisms involved in fungus-mediated tumorigenesis. An exact
mechanism to induce any specific tumor needs to be investigated. v)
What is the difference between commensal fungi and fungi isolated
from tumor tissues, such as commensal fungi in the gut and fungi in
CRC and what kind of factor(s) cause the conversion of commensal
fungi to pathogenic fungi? There are two kinds of fungi, commensal
fungi and fungi in the tumor tissues. At present, the difference
between commensal and pathogenic fungi remains unclear. In
addition, it also is unclear what kinds of factor can cause the
conversion of commensal fungi to pathogenic fungi. vi) What are the
functional differences between intracellular tumor-resident and
extracellular tumor-resident fungi? Intracellular and extracellular
fungi can be found in tumor tissues. The existence and type of
differences between intracellular and extracellular fungi are
unclear. In addition, the functional and physiological significance
of these fungi in the TME is also unclear.
The investigation of the aforementioned questions
will be decisive not only for understanding the mechanism of
fungi-mediated tumor development but also for new opportunities for
cancer therapy and diagnosis.
Availability of data and materials
Not applicable.
Authors' contributions
WC, FL and YG wrote the original draft and created
the figures. RY conceptualized the study and contributed to the
writing of the final version of the manuscript. All authors read
and approved the final manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 82271779; 91842302; 81970457;
81901677 and 91629102), the Tianjin Science and Technology
Commission (grant no. 18JCZDJC35300), the Ministry of Science and
Technology (grant no. 2016YFC1303604), the State Key Laboratory of
Medicinal Chemical Biology and the Fundamental Research Funds for
the Central University, Nankai University (grant no. 63191724) and
the Nankai University Tianjin Application and Basis Research (grant
no. 22JCQNJC00520).
References
|
1
|
Wheeler ML, Limon JJ and Underhill DM:
Immunity to commensal fungi: Detente and disease. Annu Rev Pathol.
12:359–385. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iliev ID, Funari VA, Taylor KD, Nguyen Q,
Reyes CN, Strom SP, Brown J, Becker CA, Fleshner PR, Dubinsky M, et
al: Interactions between commensal fungi and the C-type lectin
receptor Dectin-1 influence colitis. Science. 336:1314–1317. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Okuno K, Tokunaga M, Von Hoff D, Kinugasa
Y and Goel A; PDAC Biomarker Working Group: Intratumoral
malasseziaglobosa levels predict survival and therapeutic response
to adjuvant chemotherapy in patients with pancreatic ductal
adenocarcinoma. Gastroenterology. 165:502–504 e2. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang T, Fan C, Yao A, Xu X, Zheng G, You
Y, Jiang C, Zhao X, Hou Y, Hung MC and Lin X: The Adaptor Protein
CARD9 protects against colon cancer by restricting
mycobiota-mediated expansion of myeloid-derived suppressor cells.
Immunity. 49:504–514 e4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alam A, Levanduski E, Denz P,
Villavicencio HS, Bhatta M, Alhorebi L, Zhang Y, Gomez EC, Morreale
B, Senchanthisai S, et al: Fungal mycobiome drives IL-33 secretion
and type 2 immunity in pancreatic cancer. Cancer Cell. 40:153–167
e11. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Malik A, Sharma D, Malireddi RKS, Guy CS,
Chang TC, Olsen SR, Neale G, Vogel P and Kanneganti TD: SYK-CARD9
Signaling axis promotes gut fungi-mediated inflammasome activation
to restrict colitis and colon cancer. Immunity. 49:515–530 e5.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qu J, Chen Q, Bing Z, Shen S, Hou Y, Lv M
and Wang T: C. tropicalis promotes CRC by down-regulating tumor
cell-intrinsic PD-1 receptor via autophagy. J Cancer. 14:1794–1808.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang L, Li A, Wang Y and Zhang Y:
Intratumoral microbiota: Roles in cancer initiation, development
and therapeutic efficacy. Signal Transduct Target Ther. 8:352023.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Azevedo MM, Pina-Vaz C and Baltazar F:
Microbes and Cancer: Friends or Faux? Int J Mol Sci. 21:31152020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shkoporov AN and Hill C: Bacteriophages of
the Human Gut: The 'Known Unknown' of the Microbiome. Cell Host
Microbe. 25:195–209. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Poore GD, Kopylova E, Zhu Q, Carpenter C,
Fraraccio S, Wandro S, Kosciolek T, Janssen S, Metcalf J, Song SJ,
et al: Microbiome analyses of blood and tissues suggest cancer
diagnostic approach. Nature. 579:567–574. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Narunsky-Haziza L, Sepich-Poore GD,
Livyatan I, Asraf O, Martino C, Nejman D, Gavert N, Stajich JE,
Amit G, González A, et al: Pan-cancer analyses reveal
cancer-type-specific fungal ecologies and bacteriome interactions.
Cell. 185:3789–3806 e17. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dohlman AB, Klug J, Mesko M, Gao IH,
Lipkin SM, Shen X and Iliev ID: A pan-cancer mycobiome analysis
reveals fungal involvement in gastrointestinal and lung tumors.
Cell. 185:3807–3822 e12. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu NN, Jiao N, Tan JC, Wang Z, Wu D, Wang
AJ, Chen J, Tao L, Zhou C, Fang W, et al: Multi-kingdom microbiota
analyses identify bacterial-fungal interactions and biomarkers of
colorectal cancer across cohorts. Nat Microbiol. 7:238–250. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang M, Yu F and Li P: Intratumor
microbiota in cancer pathogenesis and immunity: From mechanisms of
action to therapeutic opportunities. Front Immunol. 14:12690542023.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nejman D, Livyatan I, Fuks G, Gavert N,
Zwang Y, Geller LT, Rotter-Maskowitz A, Weiser R, Mallel G, Gigi E,
et al: The human tumor microbiome is composed of tumor
type-specific intracellular bacteria. Science. 368:973–980. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Galeano Nino JL, Wu H, LaCourse KD,
Kempchinsky AG, Baryiames A, Barber B, Futran N and Houlton J:
Effect of the intratumoral microbiota on spatial and cellular
heterogeneity in cancer. Nature. 611:810–817. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fu A, Yao B, Dong T and Cai S: Emerging
roles of intratumor microbiota in cancer metastasis. Trends Cell
Biol. 33:583–593. 2023. View Article : Google Scholar
|
|
19
|
Zong Z, Zhou F and Zhang L: The fungal
mycobiome: a new hallmark of cancer revealed by pan-cancer
analyses. Signal Transduct Target Ther. 8:502023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luan C, Xie L, Yang X, Miao H, Lv N, Zhang
R, Xiao X, Hu Y, Liu Y, Wu N, et al: Dysbiosis of fungal microbiota
in the intestinal mucosa of patients with colorectal adenomas. Sci
Rep. 5:79802015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin Y, Lau HC, Liu Y, Kang X, Wang Y, Ting
NL, Kwong TN, Han J, Liu W, Liu C, et al: Altered mycobiota
signatures and enriched pathogenic aspergillus rambellii are
associated with colorectal cancer based on multicohort fecal
metagenomic analyses. Gastroenterology. 163:908–921. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coker OO, Nakatsu G, Dai RZ, Wu WKK, Wong
SH, Ng SC, Chan FKL, Sung JJY and Yu J: Enteric fungal microbiota
dysbiosis and ecological alterations in colorectal cancer. Gut.
68:654–662. 2019. View Article : Google Scholar
|
|
23
|
Aykut B, Pushalkar S, Chen R, Li Q,
Abengozar R, Kim JI, Shadaloey SA, Wu D, Preiss P, Verma N, et al:
The fungal mycobiome promotes pancreatic oncogenesis via activation
of MBL. Nature. 574:264–267. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Banerjee S, Tian T, Wei Z, Shih N, Feldman
MD, Peck KN, DeMichele AM, Alwine JC and Robertson ES: Distinct
microbial signatures associated with different breast cancer types.
Front Microbiol. 9:9512018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Banerjee S, Alwine JC, Wei Z, Tian T, Shih
N, Sperling C, Guzzo T, Feldman MD and Robertson ES: Microbiome
signatures in prostate cancer. Carcinogenesis. 40:749–764. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Banerjee S, Tian T, Wei Z, Shih N, Feldman
MD, Alwine JC, Coukos G and Robertson ES: The ovarian cancer
oncobiome. Oncotarget. 8:36225–36245. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu F, Willette-Brown J, Song NY, Lomada
D, Song Y, Xue L, Gray Z, Zhao Z, Davis SR, Sun Z, et al:
Autoreactive T cells and chronic fungal infection drive esophageal
carcinogenesis. Cell Host Microbe. 21:478–493 e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gao R, Kong C, Li H, Huang L, Qu X, Qin N
and Qin H: Dysbiosis signature of mycobiota in colon polyp and
colorectal cancer. Eur J Clin Microbiol Infect Dis. 36:2457–2468.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alnuaimi AD, Wiesenfeld D, O'Brien-Simpson
NM, Reynolds EC, Peng B and McCullough MJ: The development and
validation of a rapid genetic method for species identification and
genotyping of medically important fungal pathogens using
high-resolution melting curve analysis. Mol Oral Microbiol.
29:117–130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li JQ, Li JL, Xie YH, Wang Y, Shen XN,
Qian Y, Han JX, Chen YX and Fang JY: Saccharomyces cerevisiae may
serve as a probiotic in colorectal cancer by promoting cancer cell
apoptosis. J Dig Dis. 21:571–582. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Z, Zheng Y, Chen Y, Yin Y, Chen Y,
Chen Q, Hou Y, Shen S, Lv M and Wang T: Gut fungi enhances
immunosuppressive function of myeloid-derived suppressor cells by
activating PKM2-dependent glycolysis to promote colorectal
tumorigenesis. Exp Hematol Oncol. 11:882022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Machlowska J, Baj J, Sitarz M, Maciejewski
R and Sitarz R: Gastric cancer: Epidemiology, risk factors,
classification, genomic characteristics and treatment strategies.
Int J Mol Sci. 21:41022020. View Article : Google Scholar
|
|
34
|
Thrift AP and El-Serag HB: Burden of
gastric cancer. Clin Gastroenterol Hepatol. 18:534–542. 2020.
View Article : Google Scholar
|
|
35
|
Zhong M, Xiong Y, Zhao J, Gao Z, Ma J, Wu
Z, Song Y and Hong X: Candida albicans disorder is associated with
gastric carcinogenesis. Theranostics. 11:4945–4956. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vallianou N, Kounatidis D, Christodoulatos
GS, Panagopoulos F, Karampela I and Dalamaga M: Mycobiome and
Cancer: What is the evidence? Cancers (Basel). 13:31492021.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang L, Chen C, Chai D, Li C, Qiu Z,
Kuang T, Liu L, Deng W and Wang W: Characterization of the
intestinal fungal microbiome in patients with hepatocellular
carcinoma. J Transl Med. 21:1262023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Del Castillo E, Meier R, Chung M, Koestler
DC, Chen T, Paster BJ, Charpentier KP, Kelsey KT, Izard J and
Michaud DS: The microbiomes of pancreatic and duodenum tissue
overlap and are highly subject specific but differ between
pancreatic cancer and noncancer subjects. Cancer Epidemiol
Biomarkers Prev. 28:370–383. 2019. View Article : Google Scholar
|
|
39
|
Vitiello GA, Cohen DJ and Miller G:
Harnessing the microbiome for pancreatic cancer immunotherapy.
Trends Cancer. 5:670–676. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao Y, Yi J, Xiang J, Jia W, Chen A, Chen
L, Zheng L, Zhou W, Wu M, Yu Z and Tang J: Exploration of lung
mycobiome in the patients with non-small-cell lung cancer. BMC
Microbiol. 23:812023. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Perera M, Al-Hebshi NN, Perera I, Ipe D,
Ulett GC, Speicher DJ, Chen T and Johnson NW: A dysbiotic mycobiome
dominated by Candida albicans is identified within oral
squamous-cell carcinomas. J Oral Microbiol. 9:13853692017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Navarro-Arias MJ, Hernández-Chávez MJ,
García-Carnero LC, Amezcua-Hernández DG, Lozoya-Pérez NE,
Estrada-Mata E, Martínez-Duncker I, Franco B and Mora-Montes HM:
Differential recognition of Candida tropicalis, Candida
guilliermondii, Candida krusei, and Candida auris by human innate
immune cells. Infect Drug Resist. 12:783–794. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chang F, Syrjänen S, Wang L and Syrjänen
K: Infectious agents in the etiology of esophageal cancer.
Gastroenterology. 103:1336–1348. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang CS: Research on esophageal cancer in
China: A review. Cancer Res. 40(8 Pt 1): 2633–2644. 1980.PubMed/NCBI
|
|
45
|
Hashimoto K, Nishimura S, Shinyashiki Y,
Ito T and Akagi M: Characterizing inflammatory markers in highly
aggressive soft tissue sarcomas. Medicine (Baltimore).
101:e306882022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu X, Jiang B, Hao H and Liu Z: CARD9
Signaling, inflammation, and diseases. Front Immunol.
13:8808792022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bergmann H, Roth S, Pechloff K, Kiss EA,
Kuhn S, Heikenwälder M, Diefenbach A, Greten FR and Ruland J:
Card9-dependent IL-1β regulates IL-22 production from group 3
innate lymphoid cells and promotes colitis-associated cancer. Eur J
Immunol. 47:1342–1353. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Glocker EO, Hennigs A, Nabavi M, Schäffer
AA, Woellner C, Salzer U, Pfeifer D, Veelken H, Warnatz K, Tahami
F, et al: A homozygous CARD9 mutation in a family with
susceptibility to fungal infections. N Engl J Med. 361:1727–1735.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Leone RD and Powell JD: Metabolism of
immune cells in cancer. Nat Rev Cancer. 20:516–531. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Deng Y, Yang J, Luo F, Qian J, Liu R,
Zhang D, Yu H and Chu Y: mTOR-mediated glycolysis contributes to
the enhanced suppressive function of murine tumor-infiltrating
monocytic myeloid-derived suppressor cells. Cancer Immunol
Immunother. 67:1355–1364. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Reinfeld BI, Madden MZ, Wolf MM, Chytil A,
Bader JE, Patterson AR, Sugiura A, Cohen AS, Ali A, Do BT, et al:
Cell-programmed nutrient partitioning in the tumour
microenvironment. Nature. 593:282–288. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhu Y, Shi T, Lu X, Xu Z, Qu J, Zhang Z,
Shi G, Shen S, Hou Y, Chen Y and Wang T: Fungal-induced glycolysis
in macrophages promotes colon cancer by enhancing innate lymphoid
cell secretion of IL-22. EMBO J. 40:e1053202021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
De Monte L, Reni M, Tassi E, Clavenna D,
Papa I, Recalde H, Braga M, Di Carlo V, Doglioni C and Protti MP:
Intratumor T helper type 2 cell infiltrate correlates with
cancer-associated fibroblast thymic stromal lymphopoietin
production and reduced survival in pancreatic cancer. J Exp Med.
208:469–478. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang X, Wu S, Wu W, Zhang W, Li L, Liu Q
and Yan Z: Candida albicans promotes oral cancer via
IL-17A/IL-17RA-Macrophage axis. mBio. 14:e00447232023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xia J, Ding H, Liu S, An R, Shi X, Chen M
and Ren H: C-Type lectin receptors-triggered antifungal immunity
may synergize with and optimize the effects of immunotherapy in
hepatocellular carcinoma. J Inflamm Res. 16:19–33. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Erendor F, Sahin EO, Sanlioglu AD, Balci
MK, Griffith TS and Sanlioglu S: Lentiviral gene therapy vectors
encoding VIP suppressed diabetes-related inflammation and augmented
pancreatic beta-cell proliferation. Gene Ther. 28:130–141. 2021.
View Article : Google Scholar
|
|
57
|
Gainza-Cirauqui ML, Nieminen MT, Novak
Frazer L, Aguirre-Urizar JM, Moragues MD and Rautemaa R: Production
of carcinogenic acetaldehyde by Candida albicans from patients with
potentially malignant oral mucosal disorders. J Oral Pathol Med.
42:243–249. 2013. View Article : Google Scholar
|
|
58
|
Smith MT, Guyton KZ, Gibbons CF, Fritz JM,
Portier CJ, Rusyn I, DeMarini DM, Caldwell JC, Kavlock RJ, Lambert
PF, et al: Key characteristics of carcinogens as a basis for
organizing data on mechanisms of carcinogenesis. Environ Health
Perspect. 124:713–721. 2016. View Article : Google Scholar :
|
|
59
|
Rushing BR and Selim MI: Aflatoxin B1: A
review on metabolism, toxicity, occurrence in food, occupational
exposure, and detoxification methods. Food Chem Toxicol.
124:81–100. 2019. View Article : Google Scholar
|
|
60
|
Johnson CH, Dejea CM, Edler D, Hoang LT,
Santidrian AF, Felding BH, Ivanisevic J, Cho K, Wick EC,
Hechenbleikner EM, et al: Metabolism links bacterial biofilms and
colon carcinogenesis. Cell Metab. 21:891–897. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hold GL and Allen-Vercoe E: Gut microbial
biofilm composition and organisation holds the key to CRC. Nat Rev
Gastroenterol Hepatol. 16:329–330. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tomkovich S, Dejea CM, Winglee K, Drewes
JL, Chung L, Housseau F, Pope JL, Gauthier J, Sun X, Mühlbauer M,
et al: Human colon mucosal biofilms from healthy or colon cancer
hosts are carcinogenic. J Clin Invest. 129:1699–1712. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Garcia-Ceron D, Bleackley MR and Anderson
MA: Fungal extracellular vesicles in pathophysiology. Subcell
Biochem. 97:151–177. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Freitas MS, Bonato VLD, Pessoni AM,
Rodrigues ML, Casadevall A and Almeida F: Fungal extracellular
vesicles as potential targets for immune interventions. mSphere.
4:e00747–19. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Rodrigues ML and Casadevall A: A two-way
road: Novel roles for fungal extracellular vesicles. Mol Microbiol.
110:11–15. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Freitas MS, Bitencourt TA, Rezende CP,
Martins NS, Dourado TMH, Tirapelli CR and Almeida F: Aspergillus
fumigatus extracellular vesicles display increased galleria
mellonella survival but partial pro-inflammatory response by
macrophages. J Fungi (Basel). 9:5412023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Vargas G, Rocha JD, Oliveira DL,
Albuquerque PC, Frases S, Santos SS, Nosanchuk JD, Gomes AM,
Medeiros LC, Miranda K, et al: Compositional and immunobiological
analyses of extracellular vesicles released by Candida albicans.
Cell Microbiol. 17:389–407. 2015. View Article : Google Scholar
|
|
68
|
Bielska E, Sisquella MA, Aldeieg M, Birch
C, O'Donoghue EJ and May RC: Pathogen-derived extracellular
vesicles mediate virulence in the fatal human pathogen Cryptococcus
gattii. Nat Commun. 9:15562018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Hamad I, Ranque S, Azhar EI, Yasir M,
Jiman-Fatani AA, Tissot-Dupont H, Raoult D and Bittar F:
Culturomics and amplicon-based metagenomic approaches for the study
of fungal population in human gut microbiota. Sci Rep. 7:167882017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Leong C, Schmid B, Toi MJ, Wang J,
Irudayaswamy AS, Goh JPZ, Bosshard PP, Glatz M and Dawson TL Jr:
Geographical and ethnic differences influence culturable commensal
yeast diversity on healthy skin. Front Microbiol. 10:18912019.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chen Y, Chen Z, Guo R, Chen N, Lu H, Huang
S, Wang J and Li L: Correlation between gastrointestinal fungi and
varying degrees of chronic hepatitis B virus infection. Diagn
Microbiol Infect Dis. 70:492–498. 2011. View Article : Google Scholar
|
|
72
|
Proctor DM, Drummond RA, Lionakis MS and
Segre JA: One population, multiple lifestyles: Commensalism and
pathogenesis in the human mycobiome. Cell Host Microbe. 31:539–553.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Tsui C, Kong EF and Jabra-Rizk MA:
Pathogenesis of Candida albicans biofilm. Pathog Dis.
74:ftw0182016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Li XV, Leonardi I, Putzel GG, Semon A,
Fiers WD, Kusakabe T, Lin WY, Gao IH, Doron I, Gutierrez-Guerrero
A, et al: Immune regulation by fungal strain diversity in
inflammatory bowel disease. Nature. 603:672–678. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Seelbinder B, Lohinai Z, Vazquez-Uribe R,
Brunke S, Chen X, Mirhakkak M, Lopez-Escalera S, Dome B,
Megyesfalvi Z, Berta J, et al: Candida expansion in the gut of lung
cancer patients associates with an ecological signature that
supports growth under dysbiotic conditions. Nat Commun.
14:26732023. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zeise KD, Woods RJ and Huffnagle GB:
Interplay between Candida albicans and lactic acid bacteria in the
gastrointestinal tract: Impact on colonization resistance,
microbial carriage, opportunistic infection, and host immunity.
Clin Microbiol Rev. 34:e00323202021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
MacAlpine J, Daniel-Ivad M, Liu Z, Yano J,
Revie NM, Todd RT, Stogios PJ, Sanchez H, O'Meara TR, Tompkins TA,
et al: A small molecule produced by Lactobacillus species blocks
Candida albicans filamentation by inhibiting a DYRK1-family kinase.
Nat Commun. 12:61512021. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fan D, Coughlin LA, Neubauer MM, Kim J,
Kim MS, Zhan X, Simms-Waldrip TR, Xie Y, Hoope LV and Koh AY:
Activation of HIF-1α and LL-37 by commensal bacteria inhibits
Candida albicans colonization. Nat Med. 21:808–814. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Boutin RC, Petersen C, Woodward SE,
Serapio-Palacios A, Bozorgmehr T, Loo R, Chalanuchpong A, Cirstea
M, Lo B, Huus KE, et al: Bacterial-fungal interactions in the
neonatal gut influence asthma outcomes later in life. Elife.
10:e677402021. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Nash AK, Auchtung TA, Wong MC, Smith DP,
Gesell JR, Ross MC, Stewart CJ, Metcalf GA, Muzny DM, Gibbs RA, et
al: The gut mycobiome of the Human Microbiome Project healthy
cohort. Microbiome. 5:1532017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Gonzalez-Orozco BD, Kosmerl E,
Jiménez-Flores R and Alvarez VB: Enhanced probiotic potential of
Lactobacillus kefiranofaciens OSU-BDGOA1 through co-culture with
Kluyveromyces marxianus bdgo-ym6. Front Microbiol. 14:12366342023.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zeng X, Jia H, Shi Y, Chen K, Wang Z, Gao
Z, Yuan Y and Yue T: Lactobacillus kefiranofaciens JKSP109 and
Saccharomyces cerevisiae JKSP39 isolated from Tibetan kefir grain
co-alleviated AOM/DSS induced inflammation and colorectal
carcinogenesis. Food Funct. 13:6947–6961. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Rao C, Coyte KZ, Bainter W, Geha RS,
Martin CR and Rakoff-Nahoum S: Multi-kingdom ecological drivers of
microbiota assembly in preterm infants. Nature. 591:633–638. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hanahan D: Hallmarks of cancer: New
dimensions. Cancer Discov. 12:31–46. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Hoft MA, Hoving JC and Brown GD: Signaling
C-Type lectin receptors in antifungal immunity. Curr Top Microbiol
Immunol. 429:63–101. 2020.PubMed/NCBI
|
|
86
|
Hatinguais R, Willment JA and Brown GD:
PAMPs of the fungal cell wall and mammalian PRRs. Curr Top
Microbiol Immunol. 425:187–223. 2020.PubMed/NCBI
|
|
87
|
Witchley JN, Penumetcha P, Abon NV,
Woolford CA, Mitchell AP and Noble SM: Candida albicans
Morphogenesis Programs Control the Balance between Gut Commensalism
and Invasive Infection. Cell Host Microbe. 25:432–443 e6. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pande K, Chen C and Noble SM: Passage
through the mammalian gut triggers a phenotypic switch that
promotes Candida albicans commensalism. Nat Genet. 45:1088–1091.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Chen C, Pande K, French SD, Tuch BB and
Noble SM: An iron homeostasis regulatory circuit with reciprocal
roles in Candida albicans commensalism and pathogenesis. Cell Host
Microbe. 10:118–135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Day AM, McNiff MM, da Silva Dantas A, Gow
NAR and Quinn J: Hog1 regulates stress tolerance and virulence in
the emerging fungal pathogen Candida auris. mSphere. 3:e00506–18.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Deorukhkar SC, Saini S and Mathew S:
Non-albicans Candida Infection: An emerging threat. Interdiscip
Perspect Infect Dis. 2014:6159582014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Moyes DL, Wilson D, Richardson JP,
Mogavero S, Tang SX, Wernecke J, Höfs S, Gratacap RL, Robbins J,
Runglall M, et al: Candidalysin is a fungal peptide toxin critical
for mucosal infection. Nature. 532:64–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Gao J, Chow EWL, Wang H, Xu X, Cai C, Song
Y, Wang J and Wang Y: LncRNA DINOR is a virulence factor and global
regulator of stress responses in Candida auris. Nat Microbiol.
6:842–851. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Boutin RCT, Sbihi H, McLaughlin RJ, Hahn
AS, Konwar KM, Loo RS, Dai D, Petersen C, Brinkman FSL, Winsor GL,
et al: Composition and associations of the infant gut fungal
microbiota with environmental factors and childhood allergic
outcomes. mBio. 12:e03396202021. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Yamaguchi N, Sonoyama K, Kikuchi H, Nagura
T, Aritsuka T and Kawabata J: Gastric colonization of Candida
albicans differs in mice fed commercial and purified diets. J Nutr.
135:109–115. 2005. View Article : Google Scholar
|
|
96
|
Robbins J, Passmore GM, Abogadie FC,
Reilly JM and Brown DA: Effects of KCNQ2 gene truncation on M-type
Kv7 potassium currents. PLoS One. 8:e718092013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Goncalves B, Ferreira C, Alves CT,
Henriques M, Azeredo J and Silva S: Vulvovaginal candidiasis:
Epidemiology, microbiology and risk factors. Crit Rev Microbiol.
42:905–927. 2016. View Article : Google Scholar
|
|
98
|
Seelbinder B, Chen J, Brunke S,
Vazquez-Uribe R, Santhaman R, Meyer AC, de Oliveira Lino FS, Chan
KF, Loos D, Imamovic L, et al: Antibiotics create a shift from
mutualism to competition in human gut communities with a
longer-lasting impact on fungi than bacteria. Microbiome.
8:1332020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhai B, Ola M, Rolling T, Tosini NL,
Joshowitz S, Littmann ER, Amoretti LA, Fontana E, Wright RJ,
Miranda E, et al: High-resolution mycobiota analysis reveals
dynamic intestinal translocation preceding invasive candidiasis.
Nat Med. 26:59–64. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Chandra D, Selvanesan BC, Yuan Z, Libutti
SK, Koba W, Beck A, Zhu K, Casadevall A, Dadachova E and Gravekamp
C: 32-Phosphorus selectively delivered by listeria to pancreatic
cancer demonstrates a strong therapeutic effect. Oncotarget.
8:20729–20740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Sepich-Poore GD, Zitvogel L, Straussman R,
Hasty J, Wargo JA and Knight R: The microbiome and human cancer.
Science. 371:eabc45522021. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Dhankhar R, Gupta V, Kumar S, Kapoor RK
and Gulati P: Microbial enzymes for deprivation of amino acid
metabolism in malignant cells: Biological strategy for cancer
treatment. Appl Microbiol Biotechnol. 104:2857–2869. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Halley A, Leonetti A, Gregori A, Tiseo M,
Deng DM, Giovannetti E and Peters GJ: The role of the microbiome in
cancer and therapy efficacy: Focus on lung cancer. Anticancer Res.
40:4807–4818. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Brandi G, Turroni S, McAllister F and
Frega G: The human microbiomes in pancreatic cancer: Towards
evidence-based manipulation strategies? Int J Mol Sci. 22:99142021.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Fazzino L, Anisman J, Chacón JM and
Harcombe WR: Phage cocktail strategies for the suppression of a
pathogen in a cross-feeding coculture. Microb Biotechnol.
13:1997–2007. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wong CC and Yu J: Gut microbiota in
colorectal cancer development and therapy. Nat Rev Clin Oncol.
20:429–452. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Elkrief A, Derosa L, Kroemer G, Zitvogel L
and Routy B: The negative impact of antibiotics on outcomes in
cancer patients treated with immunotherapy: A new independent
prognostic factor? Ann Oncol. 30:1572–1579. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Mayne ST, Playdon MC and Rock CL: Diet
nutrition, and cancer: Past present and future. Nat Rev Clin Oncol.
13:504–515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
David LA, Maurice CF, Carmody RN,
Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y,
Fischbach MA, et al: Diet rapidly and reproducibly alters the human
gut microbiome. Nature. 505:559–563. 2014. View Article : Google Scholar :
|
|
110
|
Roy S and Dhaneshwar S: Role of
prebiotics, probiotics, and synbiotics in management of
inflammatory bowel disease: Current perspectives. World J
Gastroenterol. 29:2078–2100. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Clark MJ, Robien K and Slavin JL: Effect
of prebiotics on biomarkers of colorectal cancer in humans: A
systematic review. Nutr Rev. 70:436–443. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Canale FP, Basso C, Antonini G, Perotti M,
Li N, Sokolovska A, Neumann J, James MJ, Geiger S, Jin W, et al:
Metabolic modulation of tumours with engineered bacteria for
immunotherapy. Nature. 598:662–666. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Geiger R, Rieckmann JC, Wolf T, Basso C,
Feng Y, Fuhrer T, Kogadeeva M, Picotti P, Meissner F, Mann M, et
al: L-Arginine Modulates T cell metabolism and enhances survival
and anti-tumor activity. Cell. 167:829–842 e13. 2016. View Article : Google Scholar : PubMed/NCBI
|