Introduction
Gastric cancer is one of the most common types of
malignant tumors worldwide, and is possibly the main cause of
cancer-associated mortality (1). The
incidence of gastric cancer varies across populations, with almost
two-thirds of gastric cancer cases and mortalities occurring in
less developed regions (2). Despite a
decline in the incidence of gastric cancer in the West, it remains
one of the most frequent types of cancer diagnosed in China. There
has been no significant change in the number of cases of gastric
cancer-associated mortality, and the treatment methods remain
challenging (3). Although a previous
study has indicated that several mechanisms are involved in the
development of gastric cancer, its pathogenesis remains unclear
(4).
Src-associated in mitosis of 68 kDa (Sam68), which
was originally identified as a substrate for Src kinase, is a
member of the signal transduction and activation of splicing RNA
family of K homology (KH) domain-containing RNA-binding proteins
(5,6).
Sam68 is ubiquitously expressed in numerous tissues and cell lines,
and it performs important roles in gene transcription, signaling
transduction and alternative splicing via the phosphorylation
modification (7). Sam68 has
deregulated expression, and it is involved in the promotion of cell
cycle progression, cell proliferation, transformation,
tumorigenesis and metastasis in numerous types of cancer (8). However, at present, it is uncertain
whether Sam68 has clinical significance in gastric cancer.
Therefore, the present study investigated the expression of Sam68
in gastric cancer samples to identify its potential prognostic
role.
Materials and methods
Patients and tissue samples
In this retrospective study, a total of 161
surgically-resected gastric cancer tissue specimens (53 male and
108 female; 24–72 year of age), obtained from patients who were
enrolled according to the 7th edition of the International System
of Staging for Gastric Cancer (4),
were collected at the Affiliated Nantong Cancer Hospital of Nantong
University (Nantong, China) between February 2004 and February
2007. Patient baseline demographics, clinicopathological
characteristics and surgical approach information was collected
following a review of the clinical notes and histopathology
reports. Outcome data, including long-term survival, were recorded.
None of the patients received radiotherapy or chemotherapy prior to
tumor resection. Patients with disease recurrence or metastasis
were treated with platinum-based systemic chemotherapy. The present
study was approved by the Ethics Committee of the Affiliated
Nantong Cancer Hospital of Nantong University, and written informed
consent was obtained from all patients.
Western blot analysis
Frozen gastric cancer samples were homogenized in
radioimmunoprecipitation assay buffer (25 mM Tris, 150 mM NaCl,
0.1% SDS, 0.5% sodium deoxycholate and 1% Triton X-100). Following
centrifugation at 12,000 × g for 20 min at 4°C, 60 µg total protein
from each sample was resolved with 10% SDS-PAGE and transferred to
a polyvinylidene fluoride membrane (EMD Millipore, Billerica, MA,
USA). Subsequent to blocking with 5% non-fat milk at room
temperature for 60 min, the membranes were incubated with rabbit
monoclonal antibodies against Sam68 (dilution, 1:1,000; cat. no.
sc-4249; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and GAPDH
(dilution, 1:1,000; cat. no. sc-47724; Santa Cruz Biotechnology,
Inc.) at 4°C overnight. Membranes were then washed with
TBS-Tween-20 (TBST) and incubated with a horseradish peroxidase
(HRP)-conjugated anti-rabbit secondary antibody (dilution,
1:10,000; cat. no. sc-2030; Santa Cruz Biotechnology, Inc.) for 60
min at room temperature. After washing with TBST, the membrane was
developed using an enhanced chemiluminescence system (EMD
Millipore). The intensities of the protein bands were determined by
densitometry using ImageJ software (version 2.1; National
Institutes of Health, Bethesda, MD, USA).
Immunohistochemistry
The gastric cancer tissues were fixed with 10%
formalin for 24 h at room temperature and then embedded with
paraffin at 60°C for 5 min. The 8-µm thick slides were immersed in
EDTA (pH 8.0) and incubated for 20 min in a microwave oven for
antigen retrieval. Subsequent to rinsing with PBS, endogenous
peroxidase was blocked with 0.3% hydrogen peroxide in PBS at room
temperature for 15 min. The slides were incubated with anti-Sam68
antibody (dilution, 1:50; cat. no. sc-4249; Santa Cruz
Biotechnology, Inc.) in a humidified chamber at 4°C overnight.
Following additional washing with PBS three times, the sections
were sequentially incubated with HRP-conjugated secondary antibody
(dilution, 1:100; cat. no. sc-6772; Santa Cruz Biotechnology, Inc.)
at 37°C for 30 min and then washed three times with PBS. Finally,
diaminobenzidine tetrahydrochloride was used for the signal
development. Subsequently, 10% hematoxylin staining buffer (cat.
no. H9627; Sigma-Aldrich; Merck KGaA) was added to stain the
nucleus at room temperature for 30 sec. PBS was used as a negative
control. Immunoreactivity was detected using light microscopy
(DM2000, Leica Microsystems GmbH, Wetzlar, Germany), and evaluated
independently by two experienced gynecopathologists blinded to the
clinical data. The detection of Sam68 expression was performed as
previously reported (9). To analyze
the staining of Sam68 in these tissue, the 100 hematoxylin-positive
cells in which Sam68 was also positive were identified. The
staining proportion was scored as follows: 0, no positive cells; 1,
<10% positive cells; 2, 10–35% positive cells; 3, 35–70%
positive cells; 4, >70% positive cells. Staining intensity was
graded according to the method of mean optical density: 0, no
staining; 1, weak staining (light yellow); 2, moderate staining
(yellow brown); 3, strong staining (brown). The immunoreactivity
score (IRS) was calculated as the product of the staining intensity
score and the percentage of positive cells that ranged between 0
and 12. Optimized cutoff points for each categorical score were
determined using log-rank statistics. This scoring was based on the
assumption that a staining index score ≥6 indicated high Sam68
expression, whereas a staining index score ≤6 indicated low Sam68
expression as previous reported (10).
Cell culture and small interfering RNA
(siRNA) transfection
AGS cells were obtained from the American Type
Culture Collection (Manassas, VA, USA) and cultured in Dulbecco's
modified Eagle's medium (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 µg/ml streptomycin in 5% CO2 at 37°C. The
Sam68-specific siRNA, which targets the sequences
5′-TGGGATGGAGTGATAGTA-3′, 5′-AACGAAACTGGCTTTGAAA-3′,
5′-TTTGTACCACATATCCCAT-3′ and 5′-CATTTGTGACCTATGCCAT-3′, and
non-targeting control siRNA (5′-TCGTCGTTACCTCTTTCC-3′) were
designed and synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China), and transfected with Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) in compliance with the
manufacturer's protocol. Transfections were performed for 48 h at
37°C, and the efficacy of gene silencing was assessed by western
blot analysis as aforementioned (9).
Cell proliferation, migration and
invasion analysis
Cell proliferation was assessed with the Cell
Counting Kit-8 assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan), according to the kit's protocol, and the
absorbance of the samples was measured with a plate reader at 370
nm. The cell migration ability was assessed via a wound healing
assay. Equal numbers (1×105) of transfected AGS cells
were seeded onto 6-well tissue culture plates. When the cells
reached 90% confluence, a scratch wound was created in the center
of the cell monolayer by gently removing the attached cells with a
sterile plastic pipette tip. The debris was removed by washing the
cells in serum-free culture medium. Cells bordering the wound were
visualized and images were captured under an inverted microscope at
magnification, ×400 (Leica Microsystems GmbH) 24 h after the wound
was created. After wound healing from the initial distance, the
migrated distances of the cells into the wounded areas were
calculated by subtracting the distance at 24 h before. A total of
nine areas were selected randomly from each well by light
microscopy at magnification, ×400 and the cells in the triplicate
wells of each group were quantified using ImageJ (version, 2.1;
National Institutes of Health) in each experiment. Cells were also
used for invasion assays performed as previously described
(9). Transwell filters were coated on
the upper side with 30 mg Matrigel (100 µg/µl; EMD Millipore) for 2
h at 37°C, and then 1×106 cells were subcultured on the
Transwell filter. Following incubation for 24 h at 37°C, cells on
the lower surface of the chamber were fixed with PBS containing 4%
paraformaldehyde at room temperature for 24 h and stained with PBS
which contained 1% toluidine blue at room temperature for 30 min,
prior to being counted under at magnification, ×400 by light
microscopy. In all experiments, data were collected from triplicate
chambers.
Flow cytometry
To investigate the cell cycle distribution,
106 AGS cells were fixed with 70% ethanol at room
temperature for 2 h, and then washed with PBS which contained 1%
bovine serum albumin (BSA; Fluka; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) as the blocking step three times for 5 min each
at room temperature. Subsequently, cells were incubated with 40
µg/ml of RNase and 50 µg/ml of propidium iodide (PI) dissolved in
1% BSA/PBS at room temperature for 30 min. The PI content per cell
was measured with a flow cytometer. The data were analyzed using
ModFit 4.1 software (BD Biosciences, Franklin Lakes, NJ, USA) for
cell-cycle analysis.
Fluorescence microscopy
A total of 103 ASG cells were fixed with
cold PBS containing 4% paraformaldehyde at 4°C for 20 min,
permeabolized with 0.1% Triton X-100 for 10 min and then blocked
with 1% BSA/PBS for 2 h. Following washing in PBS, the cells were
incubated with TRITC-conjugated phalloidin (dilution, 1:100;
Sigma-Aldrich; Merck KGaA) and Hoechst (dilution, 1:100;
Sigma-Aldrich; Merck KGaA) for 30 min at room temperature, then
examined under a Leica confocal fluorescence microscope (DL3500;
Leica Microsystems GmbH) at magnification, ×400.
Statistical analysis
Levels of Sam68 are expressed as the median and
standard deviation. Due to the non-normal distribution of these
parameters in all groups, the non-parametric Kruskal-Wallis test
was used to analyze the association between Sam68 levels and the
clinicopathological characteristics. Spearman's correlation
analysis was used to examine the correlations between continuous
variables. Univariate survival analysis was performed using the
Kaplan-Meier method and the log-rank test. Multivariate analysis
was conducted to determine an independent effect on survival using
the Cox proportional hazards method. P<0.05 was considered to
indicate a statistically significant difference. Statistical
analyses were conducted using SPSS 16.0 (SPSS, Inc., Chicago, IL,
USA).
Results
Expression of Sam68 in gastric cancer
and adjacent normal tissue
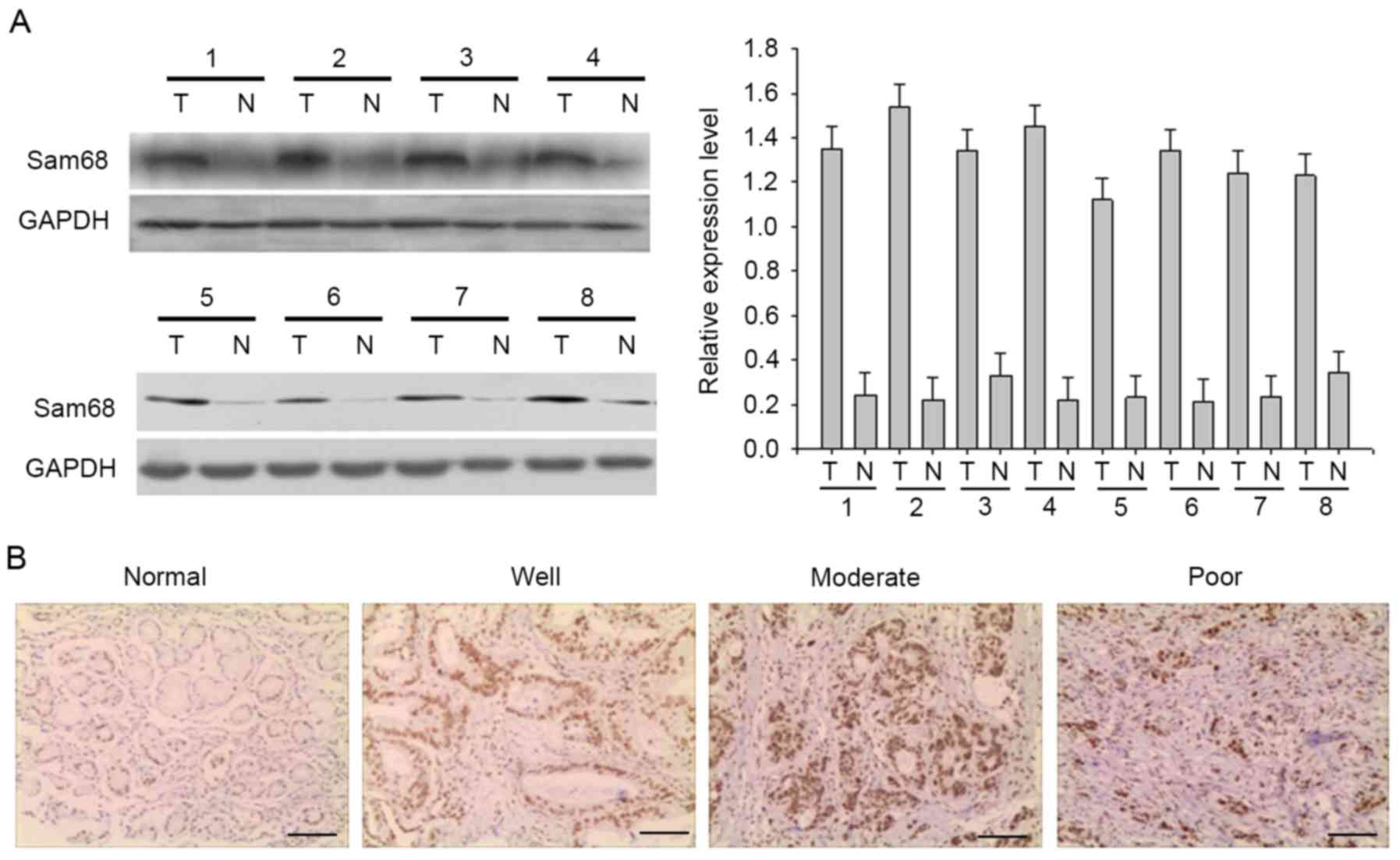
To determine the expression of Sam68, total proteins
were extracted from eight frozen matched gastric cancer and
adjacent normal tissues, and Sam68 expression was detected by
western blot analysis (Fig. 1A).
Sam68 protein was highly expressed in gastric cancer samples
compared with in the adjacent normal tissue. In addition,
immunochemistry was used to investigate Sam68 expression in the
tissue samples. In gastric cancer and adjacent normal tissue, Sam68
was predominantly found in the nucleus, although weak cytoplasmic
immunoreaction was also observed. Representative examples of
reactivity for Sam68 are presented in Fig. 1B. Absent or low Sam68 expression was
observed in adjacent normal tissue, while Sam68 expression was
upregulated in gastric cancer samples. A total of 161 cases of
gastric cancer were evaluated, and Sam68 expression was
negative/low in 78 cases (48.4%) and high in 83 cases (51.6%).
Association of Sam68 expression with
clinicopathological parameters in gastric cancer
The clinicopathological data of the patients are
summarized in Table I. As listed in
Table I, the associations of Sam68
expression with clinical variables were evaluated. Sam68 expression
was significantly associated with tumor grade (P=0.003),
infiltration depth (P=0.001), tumor-node-metastasis (TNM) stage
(P=0.012) and lymph node metastasis (P=0.001), whereas no
association was observed between Sam68 and age (P=0.224), sex
(P=0.125) and nerve invasion (P=0.987) (Table I). Furthermore, the association
between Sam68 and cell proliferation marker Ki-67 was investigated;
in the majority of specimens, the proportion of Sam68-positive
tumor cells was similar to the proportion of Ki-67-positive tumor
cells (P<0.001; Table I).
 | Table I.Sam68 expression and
clinicopathological characteristics on 161 gastric specimens. |
Table I.
Sam68 expression and
clinicopathological characteristics on 161 gastric specimens.
|
|
| Sam68 expression,
n |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Total, n | Low (78) | High (83) | P-value |
|---|
| Age |
|
|
| 0.224 |
| ≤60
years | 68 | 25 | 43 |
|
| >60
years | 93 | 53 | 40 |
|
| Sex |
|
|
| 0.125 |
|
Female | 53 | 14 | 39 |
|
| Male | 108 | 64 | 44 |
|
| Tumor grade |
|
|
| 0.003a |
| Well | 16 | 11 | 5 |
|
|
Moderate | 82 | 49 | 33 |
|
| Poor and
others | 63 | 18 | 45 |
|
| Infiltration
depth |
|
|
| 0.001a |
| Inferior
mucous membrane layer | 18 | 14 | 4 |
|
| Muscular
layer | 69 | 31 | 38 |
|
| Serous
layer | 74 | 33 | 41 |
|
| TNM stage |
|
|
| 0.012a |
| I–II | 75 | 42 | 33 |
|
|
III–IV | 86 | 36 | 50 |
|
| Lymph node |
|
|
| 0.001a |
|
Negative | 16 | 13 | 2 |
|
|
Positive | 145 | 65 | 81 |
|
| Nerve invasion |
|
|
| 0.987 |
|
Negative | 96 | 46 | 50 |
|
|
Positive | 63 | 30 | 33 |
|
| Survival
status |
|
|
| 0.008a |
|
Alive | 95 | 61 | 34 |
|
|
Dead | 66 | 17 | 49 |
|
| Ki-67
expression |
|
|
| 0.001 |
|
Low | 63 | 47 | 16 |
|
|
High | 98 | 31 | 67 |
|
Association between Sam68 and patient
survival
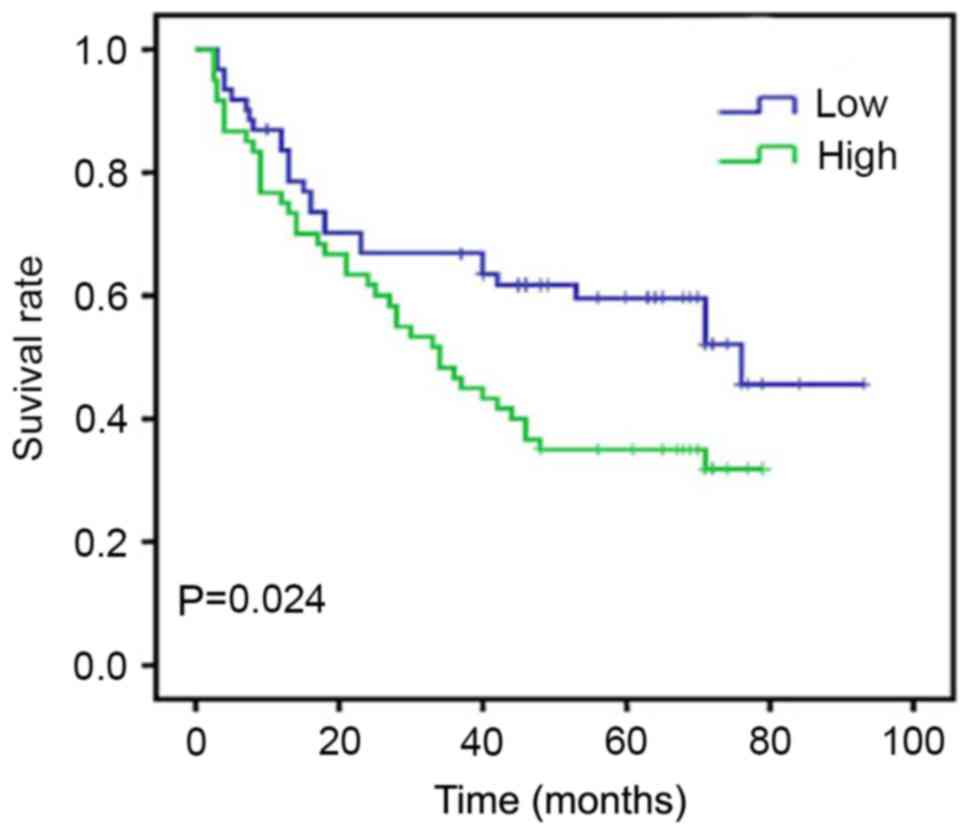
At the end of clinical follow-up of 60 months,
survival information was available in 161 cases of 161 patients
(100%). Of these 161 patients, only 34 of 83 (40.9%) patients in
the Sam68 high expression group were alive vs. 61 of 78 (78.2%) in
the Sam68 low expression group (Table
I). When all variables were compared separately with survival
status, only tumor grade (P=0.047), infiltration depth (P=0.030),
lymph node metastasis (P=0.015), Sam68 (P=0.008) and Ki-67
(P=0.025) significantly affected survival (Table II). In survival rate analysis, the
Kaplan-Meier survival curves revealed that high Sam68 expression
was associated with a poor survival, with statistical significance
(Fig. 2). Cox's proportional hazards
regression model revealed that Sam68 expression serves as an
independent marker in patients with gastric cancer (Table III).
| Table II.Contribution of various potential
prognostic factors to survival by univariate analysis in 161
gastric specimens. |
Table II.
Contribution of various potential
prognostic factors to survival by univariate analysis in 161
gastric specimens.
|
|
| Survival status,
n |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total, n | Died | Alive | P-value |
|---|
| Age |
|
|
| 0.209 |
| ≤60
years | 68 | 24 | 44 |
|
| >60
years | 93 | 42 | 51 |
|
| Sex |
|
|
| 0.664 |
|
Female | 53 | 23 | 30 |
|
|
Male | 108 | 43 | 65 |
|
| Tumor grade |
|
|
| 0.047a |
|
Well | 16 | 3 | 13 |
|
|
Moderate | 82 | 31 | 51 |
|
| Poor
and others | 63 | 32 | 31 |
|
| Infiltration
depth |
|
|
| 0.030a |
|
Inferior mucous membrane
layer | 18 | 4 | 14 |
|
|
Muscular layer | 69 | 24 | 45 |
|
| Serous
layer | 74 | 38 | 36 |
|
| TNM stage |
|
|
| 0.127 |
|
I–II | 75 | 26 | 49 |
|
|
III–IV | 86 | 40 | 46 |
|
| Lymph node |
|
|
| 0.015a |
|
Negative | 16 | 2 | 14 |
|
|
Positive | 145 | 64 | 81 |
|
| Nerve invasion |
|
|
| 0.587 |
|
Negative | 96 | 37 | 59 |
|
|
Positive | 63 | 27 | 36 |
|
| Sam68
expression |
|
|
| 0.008a |
|
Low | 78 | 61 | 17 |
|
|
High | 83 | 34 | 49 |
|
| Ki-67
expression |
|
|
| 0.025a |
|
Low | 63 | 19 | 44 |
|
|
High | 98 | 47 | 51 |
|
| Table III.Contribution of various potential
prognostic factors to survival by Cox regression analysis in 161
gastric specimens. |
Table III.
Contribution of various potential
prognostic factors to survival by Cox regression analysis in 161
gastric specimens.
| Category | 95% confidence
interval | P-value |
|---|
| Tumor grade | 0.875–2.053 | 0.177 |
| TNM stage | 1.038–2.838 | 0.035a |
| Lymph node | 1.362–28.857 | 0.018a |
| Sam68
expression | 1.687–2.121 | 0.012a |
| Ki-67
expression | 1.287–4.433 | 0.006a |
Effect of Sam68 expression on cell
proliferation in gastric cancer cells
Based on the present study, the role of Sam68 in the
proliferation of gastric cancer cells was elucidated. As previously
reported, cells were arrested in the G1 phase through
serum deprivation for 72 h. Upon serum addition, the cells
reentered the S phase (Fig. 3A).
Western blot analysis revealed that the expression of Sam68 was
increased as early as 12 h after serum stimulation (Fig. 3B). To detect the role of Sam68 on cell
proliferation, siRNAs were used to suppress Sam68 expression. The
third siRNA resulted in an ~60% decrease in Sam68 protein
expression compared with the control siRNA (Fig. 3C). In addition, the suppression of
endogenous Sam68 effectively inhibited cell viability, the cell
cycle and colony formation ability of gastric cancer cells
(Fig. 3D-F). Cell cycle-associated
protein expression levels were detected in Sam68-knockdown gastric
cells. As presented in Fig. 3G, the
expression of cyclin-dependent kinase (CDK)4, CDK6, proliferating
cell nuclear antigen and cyclin D1 was markedly decreased in
Sam68-siRNA-transfected cells, and p16 was increased. Combined,
these results confirmed that Sam68 promotes cell proliferation in
gastric cancer.
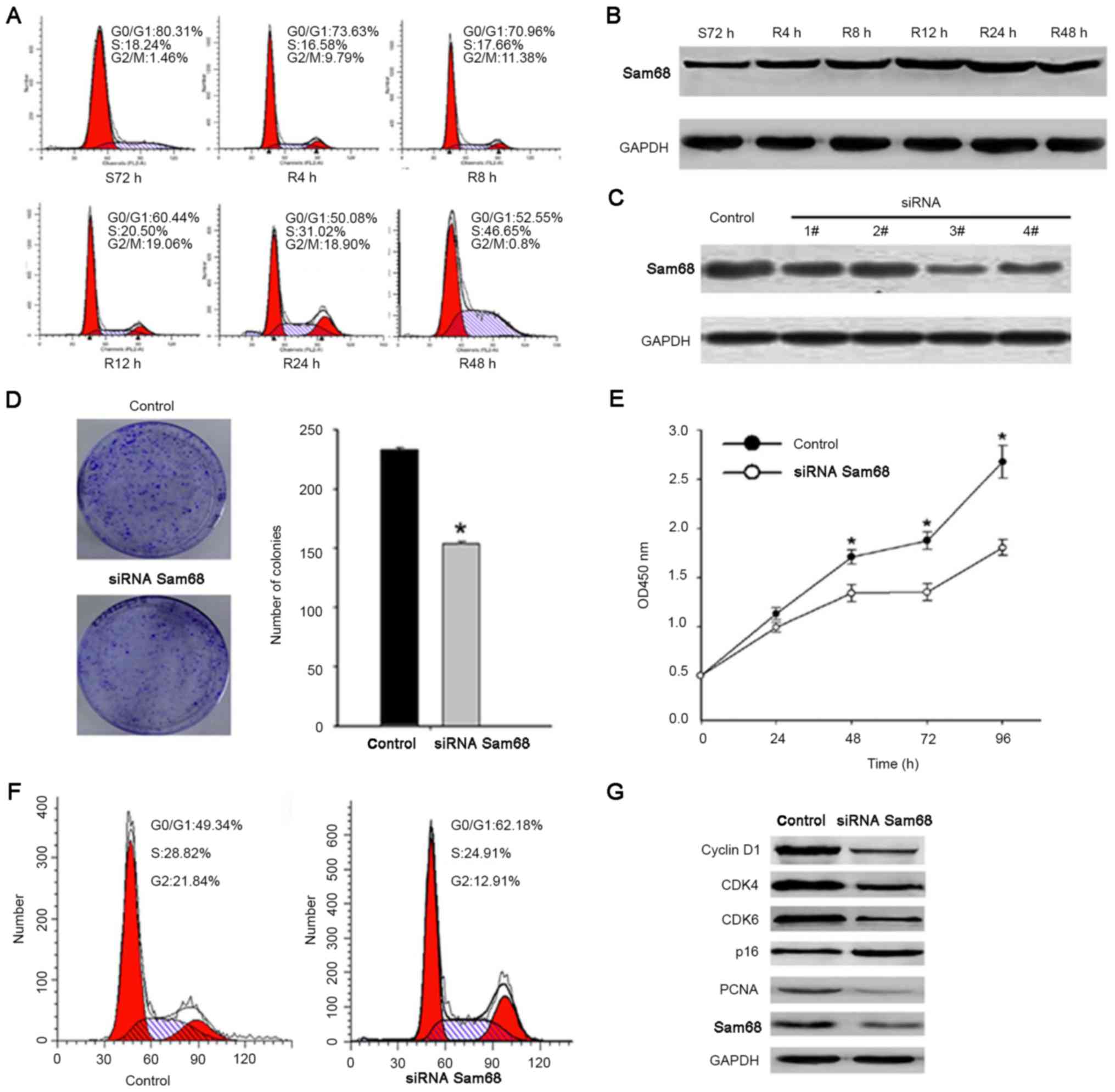 | Figure 3.Silencing of Sam68 decreases gastric
cancer cell growth. (A) Cells synchronized at
G0/G1 progressed in the cell cycle when the
S72 h cells were released by reseeding with serum for 0 (0 h=S72
h), 4, 8, 12, 24 or 48 h. (B) Cell lysates of the corresponding
time point were prepared and analyzed by western blot analysis
using antibodies against Sam68 and GAPDH. (C) The expression of
Sam68 was detected by western blot analysis following transfection
with Sam68 siRNA, while Sam68 siRNA3 achieved the best
downregulation effect. (D) Equal numbers of control-siRNA
transfected and Sam68-siRNA transfected cells were seeded onto
60-mm plates. Cells were fixed and stained with giemsa after 14
days. The number of Sam68-siRNA cell colonies was significantly
less than that of the control-siRNA cells. *P<0.05, compared
with the control cells. (E) Growth curves of cells transfected with
the Sam68 siRNA or control siRNA were drawn. *P<0.05, compared
with the control cells. (F) Flow cytometry analysis of the cell
cycle distribution of cells transfected with the Sam68 siRNA or
control siRNA. (G) Western blot analysis indicated the effect of
Sam68-knockdown on the protein expression of CDK4, CDK6, cyclin D1,
PCNA and p16. Sam68, Src-associated in mitosis of 68 kDa; CDK,
cyclin-dependent kinase; PCNA, proliferating cell nuclear antigen;
siRNA, small interfering RNA. |
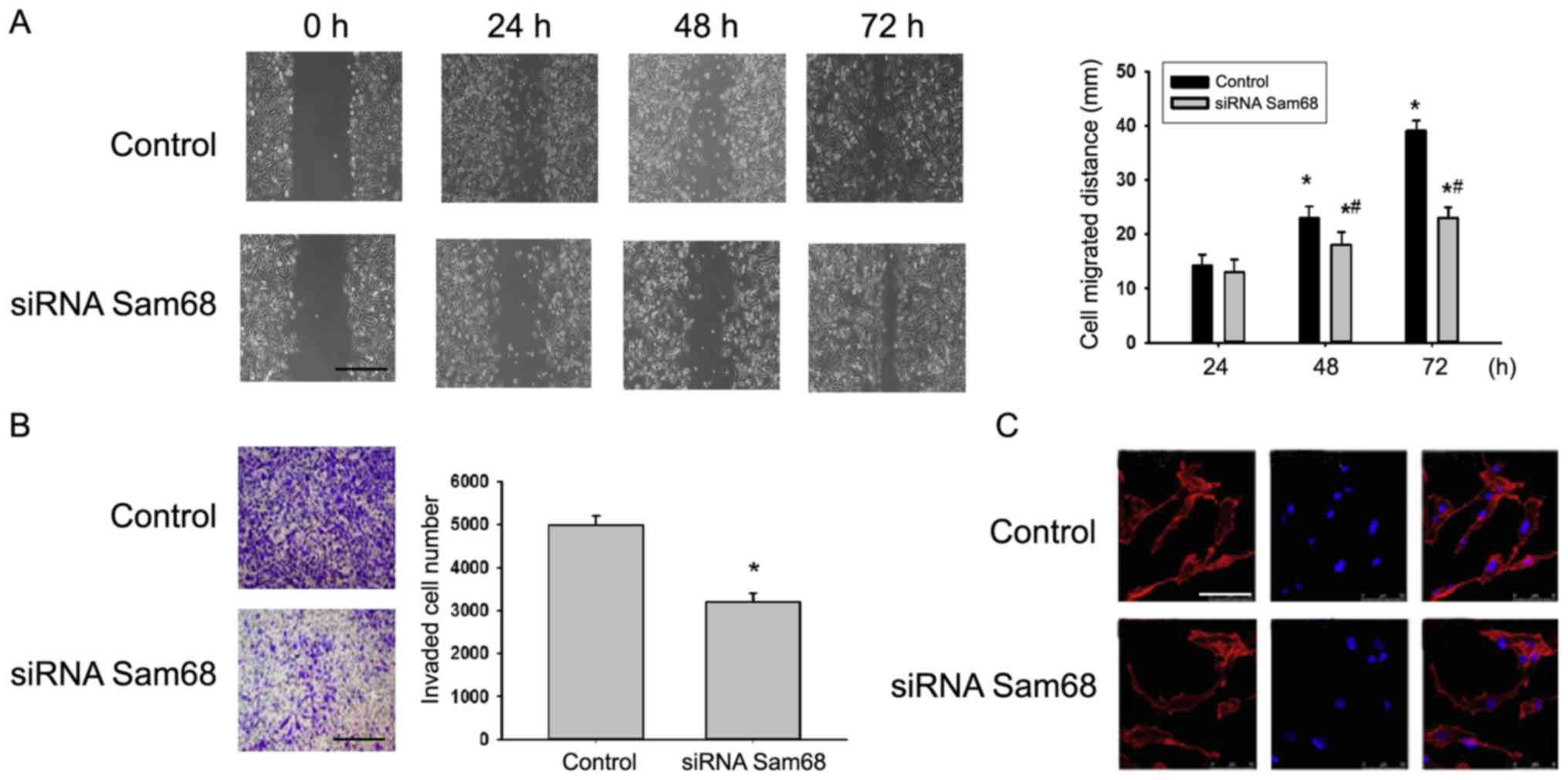
Effect of Sam68 expression on gastric
cancer cell migration and invasion
In the present study, the effect of Sam68 on cancer
metastasis was detected. Fig. 4A
depicts representative photo-micrographs captured at 0, 24, 48 and
72 h after the cell monolayer wounding. The migratory capacity of
cells transfected with siRNA targeting Sam68 was significantly
decreased compared with cells transfected with non-specific siRNA
(Fig. 4A). The effect of Sam68 on the
invasion of cell lines was then detected via a Transwell assay.
Depletion of Sam68 by siRNA decreased the number of invading cells
compared with the control (Fig. 4B).
Furthermore, it was revealed that the F-actin distribution in
Sam68-knockdown cells was abnormal when compared with the control
group (Fig. 4C). Combined, the
present data indicated that Sam68 may perform a vital role in cell
migration and invasion.
Discussion
In recent years, mounting evidence has demonstrated
that Sam68 is involved in regulating the expression of genes
relevant to multiple human diseases (11,12).
However, the deregulation of Sam68 has only been reported in
certain human cancer types, and it is unclear whether the
deregulation of Sam68 is also an event in human gastric cancer. The
current study indicated that Sam68 expression was significantly
elevated in gastric cancer tissues, as compared with in adjacent
non-cancerous tissues.
Sam68 is a substrate of the oncogenic Src kinase,
which is frequently activated in cancer (13). Current data regarding the role of
Sam68 are context-dependent and contrary (14). A previous study indicated that Sam68
functions as a tumor suppressor. Sam68 deficiency resulted in the
neoplastic transformation of murine NIH-3T3 fibroblasts, and Sam68
downregulation was associated with the capacity to form metastatic
tumors in nude mice, whereas the overexpression of Sam68 in NIH-3T3
fibroblasts induced cell cycle arrest and apoptosis (8). By contrast, studies have demonstrated
that Sam68 acts as an oncogene (15).
Sam68 haploid sufficiency delays the onset of mammary tumorigenesis
and metastasis in nude mice (16).
Busà et al (17) established
that Sam68 is upregulated in prostate cancer, and that Sam68
downregulation delayed the cell cycle progression and reduced
prostate cancer cell proliferation. The upregulation of Sam68 is
also associated with shorter survival time in cervical, gastric and
renal cell carcinoma (18). The
current study revealed that Sam68 is elevated in gastric cancer
tissues, and that high Sam68 expression levels are significantly
associated with the characteristics of aggressive gastric cancer,
including advanced TNM stage and lymph node metastasis. In
addition, the results demonstrated that Sam68 can promote the
development and progression of gastric cancer, supporting the
oncogenic role of Sam68 in cancer. Furthermore, high Sam68
expression levels may function as an independent biomarker for
gastric cancer prognosis.
In the majority of cells, Sam68 predominantly
resides within the nucleus and is involved in gene transcription,
alternative splicing and nuclear export (19). In the present study, Sam68 was
determined to localize in cancer cell nuclei (17,20).
However, contrary to the present results, the cytoplasmic
localization of Sam68 was significantly associated with poor
prognosis and progression in renal cell carcinoma and gastric
cancer. This may be due to the varying functions of Sam68 in
various signaling pathways, and the cytoplasmic and nuclear
localization of Sam68 could contribute to neoplastic transformation
or tumor progression through different molecular mechanisms in
varying cancer types or cellular contexts (21–23).
In conclusion, the present study evaluated the
potential of Sam68 as a clinically relevant indicator for patients
with gastric cancer. Additional studies of the mechanisms
underlying the involvement of Sam68 in the development and
progression of gastric cancer are required.
References
|
1
|
Li Y, Gao H, Wang Y and Dai C:
Investigation the mechanism of the apoptosis induced by lactacystin
in gastric cancer cells. Tumour Biol. 36:3465–3470. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mimica M, Tomić M, Babić E, Karin M,
Bevanda M, Alfirević D, Godler Ğ and Karan D: Gastric cancer with
bone marrow invasion presenting as severe thrombocytopenia. Turk J
Gastroenterol. 25 Suppl 1:S229–S230. 2014. View Article : Google Scholar
|
|
3
|
Cho JM, Jang YJ, Kim JH, Park SS, Park SH
and Mok YJ: Pattern, timing and survival in patients with recurrent
gastric cancer. Hepatogastroenterology. 61:1148–1153.
2014.PubMed/NCBI
|
|
4
|
Sobin LH, Gospodarowicz MK and Wittekind
C: TNM Classification of Malignant Tumors. 7th. Wiley-Blackwell;
Oxford: 2010
|
|
5
|
Quintana-Portillo R, Canfrán-Duque A,
Issad T, Sánchez-Margalet V and González-Yanes C: Sam68 interacts
with IRS1. Biochem Pharmacol. 83:78–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rajan P, Gaughan L, Dalgliesh C, El-Sherif
A, Robson CN, Leung HY and Elliott DJ: The RNA-binding and adaptor
protein Sam68 modulates signal-dependent splicing and
transcriptional activity of the androgen receptor. J Pathol.
215:67–77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reddy TR, Suhasini M, Xu W, Yeh LY, Yang
JP, Wu J, Artzt K and Wong-Staal F: A role for KH domain proteins
(Sam68-like mammalian proteins and quaking proteins) in the
post-transcriptional regulation of HIV replication. J Biol Chem.
277:5778–5784. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pedrotti S, Bielli P, Paronetto MP,
Ciccosanti F, Fimia GM, Stamm S, Manley JL and Sette C: The
splicing regulator Sam68 binds to a novel exonic splicing silencer
and functions in SMN2 alternative splicing in spinal muscular
atrophy. EMBO J. 29:1235–1247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Q, Li M, Zhang X, Huang H, Huang J,
Ke J, Ding H, Xiao J, Shan X, Liu Q, et al: Upregulation of CDK7 in
gastric cancer cell promotes tumor cell proliferation and predicts
poor prognosis. Exp Mol Pathol. 100:514–521. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang T, Wan C, Shi W, Xu J, Fan H, Zhang
S, Lin Z, Ni R and Zhang X: The RNA-binding protein Sam68 regulates
tumor cell viability and hepatic carcinogenesis by inhibiting the
transcriptional activity of FOXOs. J Mol Histol. 46:485–497. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huot ME, Brown CM, Lamarche-Vane N and
Richard S: An adaptor role for cytoplasmic Sam68 in modulating Src
activity during cell polarization. Mol Cell Biol. 29:1933–1943.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huot MÉ, Vogel G, Zabarauskas A, Ngo CT,
Coulombe-Huntington J, Majewski J and Richard S: The Sam68 STAR
RNA-binding protein regulates mTOR alternative splicing during
adipogenesis. Mol Cell. 46:187–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sánchez-Jiménez F and Sánchez-Margalet V:
Role of Sam68 in post-transcriptional gene regulation. Int J Mol
Sci. 14:23402–23419. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stockley J, Markert E, Zhou Y, Robson CN,
Elliott DJ, Lindberg J, Leung HY and Rajan P: The RNA-binding
protein Sam68 regulates expression and transcription function of
the androgen receptor splice variant AR-V7. Sci Rep. 5:134262015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fu K, Sun X, Zheng W, Wier EM, Hodgson A,
Tran DQ, Richard S and Wan F: Sam68 modulates the promoter
specificity of NF-κB and mediates expression of CD25 in activated T
cells. Nat Commun. 4:19092013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Richard S, Vogel G, Huot ME, Guo T, Muller
WJ and Lukong KE: Sam68 haploinsufficiency delays onset of mammary
tumorigenesis and metastasis. Oncogene. 27:548–556. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Busà R, Geremia R and Sette C: Genotoxic
stress causes the accumulation of the splicing regulator Sam68 in
nuclear foci of transcriptionally active chromatin. Nucleic Acids
Res. 38:3005–3018. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Busà R, Paronetto MP, Farini D,
Pierantozzi E, Botti F, Angelini DF, Attisani F, Vespasiani G and
Sette C: The RNA-binding protein Sam68 contributes to proliferation
and survival of human prostate cancer cells. Oncogene.
26:4372–4382. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lock P, Fumagalli S, Polakis P, McCormick
F and Courtneidge SA: The human p62 cDNA encodes Sam68 and not the
RasGAP-associated p62 protein. Cell. 84:23–24. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song J and Richard S: Sam68 regulates S6K1
alternative splicing during adipogenesis. Mol Cell Biol.
35:1926–1939. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao X, Li Z, He B, Liu J, Li S, Zhou L,
Pan C, Yu Z and Xu Z: Sam68 is a novel marker for aggressive
neuroblastoma. Onco Targets Ther. 6:1751–1760. 2013.PubMed/NCBI
|
|
22
|
Modem S, Badri KR, Holland TC and Reddy
TR: Sam68 is absolutely required for Rev function and HIV-1
production. Nucleic Acids Res. 33:873–879. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Paronetto MP, Messina V, Barchi M, Geremia
R, Richard S and Sette C: Sam68 marks the transcriptionally active
stages of spermatogenesis and modulates alternative splicing in
male germ cells. Nucleic Acids Res. 39:4961–4974. 2011. View Article : Google Scholar : PubMed/NCBI
|