Introduction
Liver cancer, one of the most malignant cancer
types, is a leading cause of cancer-associated cases of mortality.
It was responsible for 782,500 and 745,500 cases if mortality
worldwide in 2012 (1). The majority
(85–90%) of primary liver cancer cases are hepatocellular carcinoma
(HCC) (2). Interleukin 6 (IL-6) is
one of the best-characterized tumorigenic cytokines, particularly
in promoting HCC progression (3–6).
Expression levels of IL-6 were previously identified to be
increased in liver cirrhosis and HCC (7,8). An
increasing number of studies have demonstrated that following
chronic liver damage or viral hepatitis, elevated IL-6 activates
compensatory proliferation of quiescent hepatocytes, which
eventually results in HCC (5,9,10).
Accumulation of unfolded or misfolded proteins in
the endoplasmic reticulum (ER) lumen causes ER stress and initiates
the activation of the unfolded protein response (UPR). In mammals,
UPR pathways are comprised of three branches, which are initiated
by three ER-localized transmembrane signal transducers. These
include activating transcription factor 6 (ATF6),
inositol-requiring enzyme 1 (IRE1) and double-stranded
RNA-activated protein kinase-like ER kinase (PERK). Among the UPR
pathways, the IRE1α-XBP1 branch is the most conserved, indicating
its essential role in cells (11–13). Upon
activation, IRE1α catalyzes the non-conventional splicing of the
mRNA encoding X-box-binding protein 1 (XBP1) by removing a
26-nucleotide intron, and thereby produces an active spliced form
(XBP1s) to initiate a key UPR program (14).
Persistent activation of UPR is reported in various
solid tumor types, including liver cancer tissue sections (15). An increasing number of reports have
identified somatic IRE1α mutations in various types of cancer,
including glioblastoma, adenocarcinoma in lung and stomach, renal
clear cell carcinoma and serous ovarian cancer (16,17).
While signal transducer and activator of
transcription 3 (STAT3) is transiently activated in normal cells,
it is frequently reported to maintain a constitutively activated
state and promote tumorigenesis by enhancing angiogenesis and cell
proliferation and survival in different types of cancer, including
colon cancer, melanoma and myeloma (18–20).
Notably, IL-6 was revealed to act in an autocrine/paracrine manner
to provide a pivotal survival signal via activation of STAT3
signaling in lymphoid malignancies (20) and melanoma (21). A previous study demonstrated that the
spliced form of XBP1 may drive the transcription of IL-6 in
macrophages upon lipopolysaccharide (LPS) stimulation (22). Notably, IL-6 was recently identified
to induce the expression of XBP1 during liver regeneration
(23). These results suggest a
complex relationship between IL-6 and XBP1. However, the molecular
mechanisms underlying the regulation of hepatic expression of IL-6
and XBP1 during the pathogenesis of HCC remain unclear.
The current study reports the critical role of the
IRE1α-XBP1 branch of UPR in promoting the proliferation of HCC
cells. Elevated expression of IL-6 driven by XBP1s led to HCC cell
proliferation via activation of STAT3 signaling. This effect of
IRE1α-XBP1 was abolished when IL-6-STAT3 signaling was blocked.
Patients and methods
Patient characteristics
Paired human non-cancerous liver tissues and HCC
tissues were collected from 17 patients and analyzed in the current
study. The patients were diagnosed with HCC from 2013 to 2016 in
the Department of Pathology, The Second Affiliated Hospital and
Yuying Children's Hospital, Wenzhou Medical University (Wenzhou,
China). The information of each patient was recorded and could be
accessed during and after the data collection in this study. The
clinical characteristics of the patients are presented in Table I. The tissue sample collection was
approved by the Ethics Committee of The Second Affiliated Hospital
and Yuying Children's Hospital, Wenzhou Medical University.
Informed consent was obtained from all subjects.
 | Table I.Clinical characteristics of patients
with hepatocellular carcinoma. |
Table I.
Clinical characteristics of patients
with hepatocellular carcinoma.
| Characteristic | n (%) |
|---|
| Age, years |
|
|
≤45 | 6 (35.2) |
|
45–65 | 8 (47.1) |
|
≥65 | 3 (17.6) |
| Sex |
|
|
Male | 11 (64.7) |
|
Female | 6 (35.3) |
| Risk factor |
|
|
HBV | 8 (47.1) |
|
HCV | 4 (23.5) |
| Alcohol | 3 (17.6) |
| Other | 2 (11.8) |
Cell culture and ELISA
Normal hepatocyte cell lines (LO2 and THLE-2) and
HCC cell lines (Hep3B, Huh7, SKHep-1, MHCC97L and MHCC97H) were
obtained from Cell Bank of Shanghai, Chinese Academy of Sciences
(Shanghai, China). All cell lines were cultured in high-glucose
Dulbecco's modified Eagle's medium supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA).
To overexpress IRE1α or XBP1s, indicated plasmids
were transfected into cells using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. For inhibition of IRE1α activity, 4µ8C (Selleck
Chemicals, Shanghai, China) was dissolved in DMSO and added to the
medium of indicated cells at a final concentration of 10 µM for 24
h. To block IL-6 receptors, cells were incubated with tocilizumab
(Genentech; Roche Diagnostics, Basel, Switzerland) for 8 h prior to
further assays and analysis.
To knockdown endogenous XBP1, 50 nM shXBP1
(Genepharma; Shanghai, China) were transfected into cells using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. 48 h after transfection,
cells were collected for further analysis. The sequences are as
following:
shXBP1: Sense, 5′-CCAGUCAUGUUCUUCAAAUTT-3′ and
antisense, 5′-AUUUGAAGAACAUGACUGGTT-3′; Negative control shRNA for
shXBP1: Sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′.
Cell culture medium of Hep3B cells was collected and
used for the determination of IL-6 content using a human IL-6 ELISA
kit (eBioscience; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol.
CCK8 and BrdU assay
To determine the effects of IRE1α and XBP1s on cell
proliferation, CCK8 and BrdU assays were performed as previously
described. Briefly, 1×103 cells were seeded onto 96-well
culture plates at day 0. Then, cells were cultured for different
time periods (1–5 days) and incubated with CCK8 reagent (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) for 2 h at 37°C on
the indicated day. The staining intensity in the medium was
measured by reading the absorbance at 450 nm. BrdU assays were
performed using a BrdU Cell Proliferation assay kit (Cell Signaling
Technology, Inc., Danvers, MA, USA) according to the manufacturer's
protocol.
Luciferase reporter assay
The pGL3 basic plasmid was constructed with the
insertion of the promoter of the human IL-6 gene,
corresponding to the region of −2000 to +100 bp with respect to the
putative transcription start site (denoted nucleotide +1). The ACGT
core from the IL-6 promoter was deleted under a PCR-based strategy.
The designed plasmids were transfected into 293T cells and
luciferase activities were measured using a Dual-Luciferase assay
kit (Promega Corporation, Madison, WI, USA) according to the
manufacturer's protocol. Renilla luciferase activity was used as an
internal control for normalization.
Chromatin immunoprecipitation
(ChIP)
ChIP assays were conducted using an Agarose ChIP kit
(Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Firstly, indicated cells were subjected to
cross-linking with 1% formaldehyde. Glycine solution was added to
stop the cross-linking process then the cells were lysed for the
preparation of nuclear extracts. Subsequently, chromatin-XBP1s
complexes were immunoprecipitated with anti-Flag (diluted 1:500;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) or anti-XBP1s
(diluted 1:100; BioLegend, Inc., San Diego, CA, USA) antibodies at
4°C overnight, followed by incubation with beads from the kit at
4°C for 1 h with gentle agitating. The complexes were eluted from
the beads using several washes with the elution buffer, prior to
being subjected to further PCR analysis.
Statistical analysis
All experiments in the current study were repeated
more than three times. Data are presented as the mean ± standard
error of the mean. Statistical analysis was performed using
GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Data were analyzed using two-tailed unpaired Student's t-tests
after a demonstration of homogeneity of variance with the F test,
or one-way or two-way analysis of variance (ANOVA) for comparisons
of more than two groups. Turkey post hoc test was used after
one-way ANOVA and Bonferroni post hoc tests were used after two-way
ANOVA. P<0.05 was considered to indicate a statistically
significant difference. For correlation analysis, linear regression
analysis is applied and the coefficient of determination
(r2) and P-value are indicated.
Results
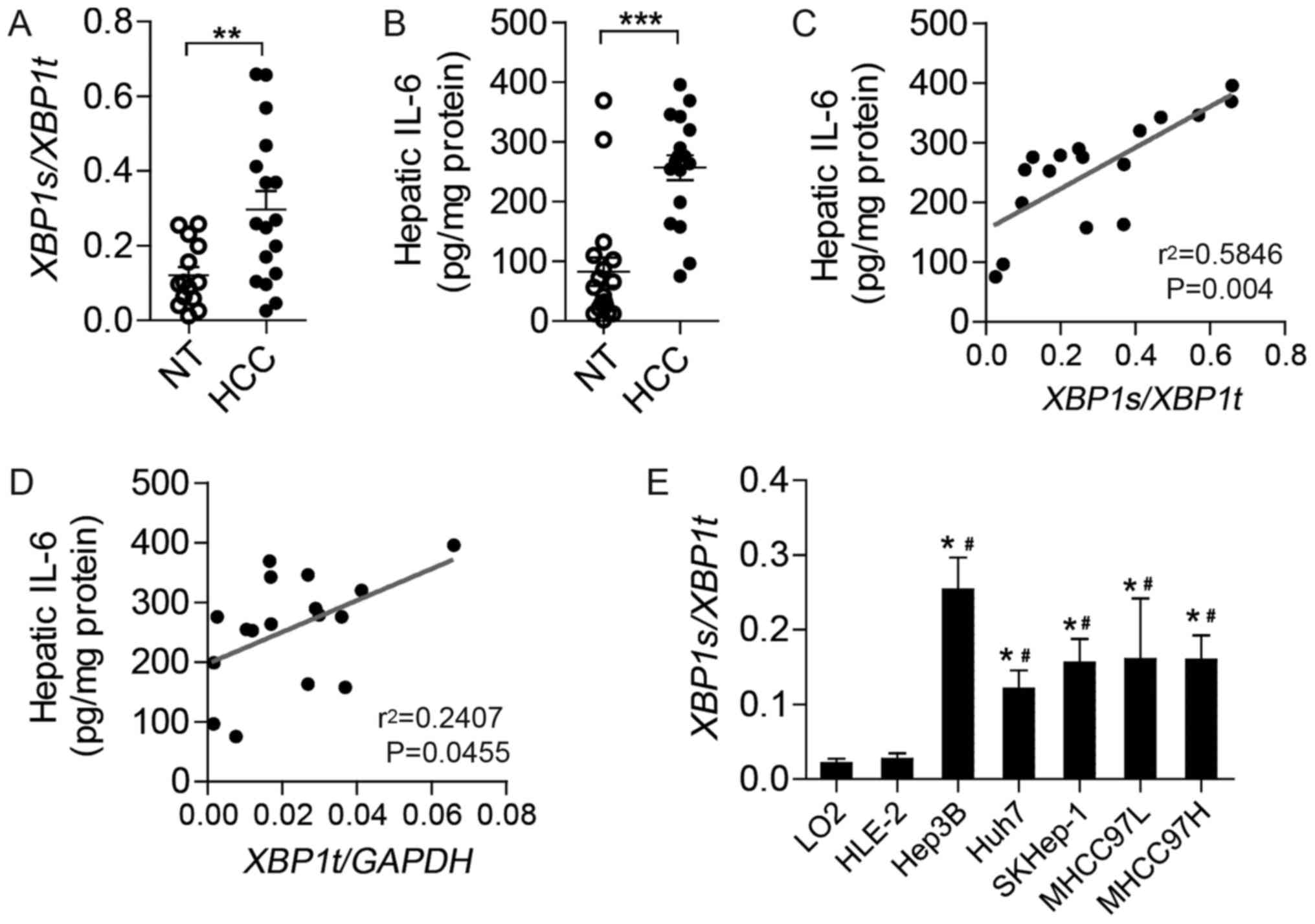
Elevated XBP1 splicing in tumor
tissues of patients with HCC and HCC cell lines
To investigate the expression of XBP1s and
IL-6 in human HCC tissues, splicing levels of XBP1 mRNA and
IL-6 content were analyzed in normal liver tissues and tumor
tissues of patients with HCC. Compared with normal liver tissues,
HCC tumors exhibited markedly increased XBP1 splicing
(Fig. 1A) and IL-6 protein (Fig. 1B). Notably, further analysis revealed
a positive correlation between hepatic IL-6 content and the level
of XBP1 splicing (Fig. 1C) as
well as XBP1t mRNA levels (Fig.
1D), indicating a close association between IL-6 and XBP1 in
HCC.
To explore the extent of XBP1 splicing in
HCC, XBP1 splicing was evaluated in a series of cell lines,
including human normal hepatocyte cell lines (LO2 and THLE-2) and
HCC cell lines (Hep3B, Huh7, SKHep-1, MHCC97L and MHCC97H).
Relative to LO2 and THLE-2 cells, almost all the HCC cell lines
exhibited notably higher levels of XBP1s generated from the
alternative splicing of XBP1u (unspliced XBP1) mRNA,
indicating enhanced activation of the IRE1α-XBP1 branch of UPR in
HCC cells (Fig. 1E).
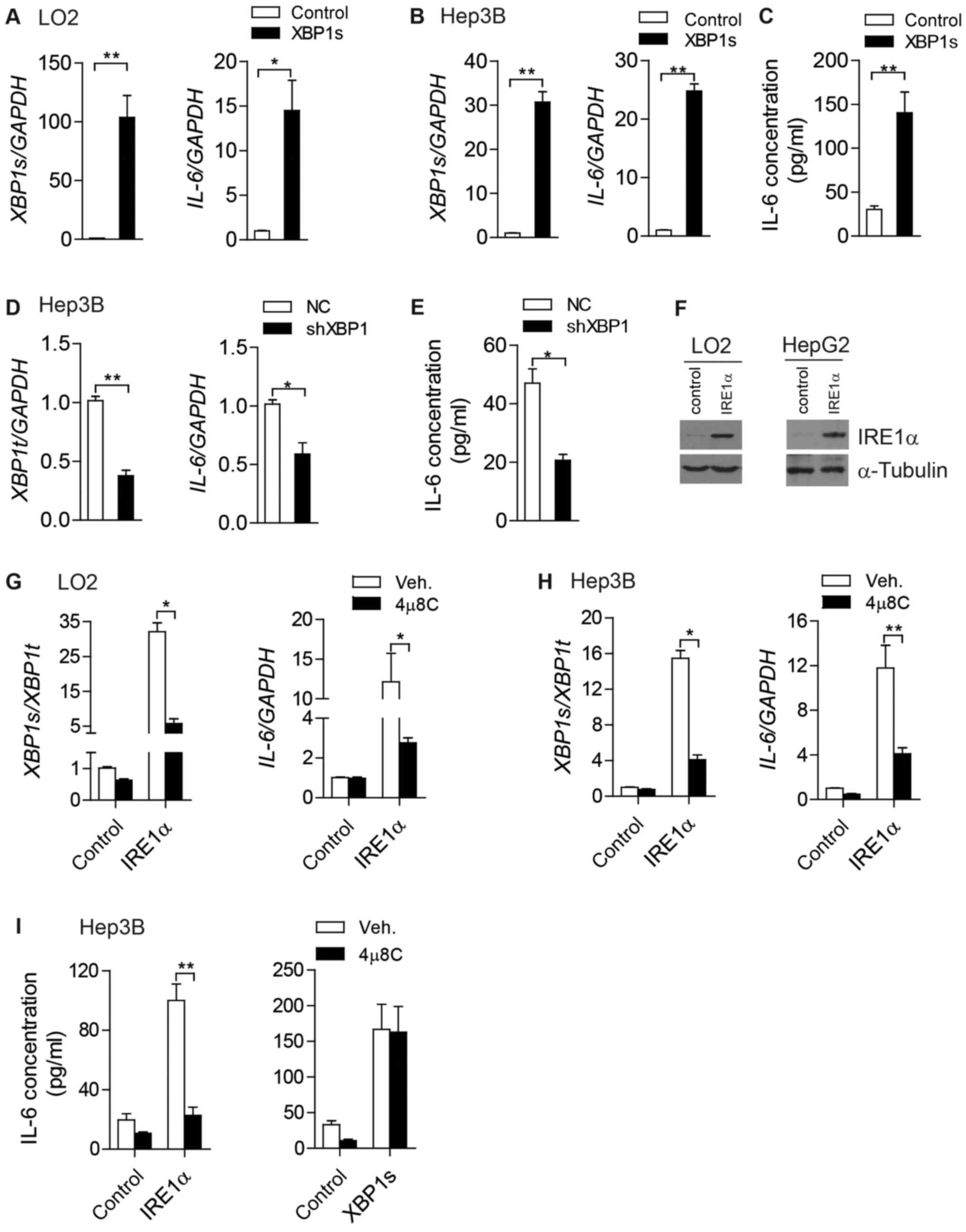
IRE1α-XBP1 pathway regulates IL-6
expression in LO2 and Hep3B cells
To investigate the physiological functions of
increased XBP1s in HCC cells, XBP1s was overexpressed in LO2 and
Hep3B cells (Fig. 2A and B). Notably,
mRNA levels of Il-6 were markedly increased in
XBP1s-overexpressing LO2 cells (Fig.
2A) and Hep3B cells (Fig. 2B). To
explore the secretion of IL-6 by HCC cells, the cell culture medium
of Hep3B cells was collected and subjected to ELISA in order to
determine extracellular levels of IL-6. Extracellular IL-6 content
also exhibited a marked increase following the overexpression of
XBP1s in Hep3B cells (Fig. 2C).
Consistent with these results, inhibition of XBP1 in Hep3B cells
resulted in a significant reduction of both Il-6 mRNA levels
(Fig. 2D) and extracellular IL-6
content (Fig. 2E).
Next, IRE1α was overexpressed in LO2 and Hep3B cells
to determine the role of IRE1α in regulating IL-6 expression
(Fig. 2F). Trans-autophosphorylation
and subsequent activation of RNase activity of IRE1α may occur
following excess accumulation of the protein, which would catalyze
the alternative splicing process of XBP1 mRNA (24). A significant increase in XBP1s
mRNA was observed in IRE1α-overexpressing LO2 cells (Fig. 2G) and Hep3B cells (Fig. 2H). This effect was abolished when the
RNase activity of IRE1α was inhibited by the addition of 4µ8C
(Fig. 2G and H) (24). Consistent with this, ectopic
expression of IRE1α increased the mRNA levels of IL-6 in LO2
(Fig. 2G) and Hep3B cells (Fig. 2H). With decreased levels of XBP1
splicing, 4µ8C-treated LO2 cells and Hep3B cells exhibited
attenuated IL-6 expression even when IRE1α was overexpressed
(Fig. 2G and H). Consistent with the
intracellular changes of IL-6 mRNA, extracellular secretion
of IL-6 by Hep3B cells was markedly upregulated when IRE1α or XBP1s
were overexpressed (Fig. 2I).
Furthermore, 4µ8C treatment blocked the effects of IRE1α
overexpression on IL-6 expression, but did not have an impact on
the effects of XBP1s overexpression (Fig.
2I).
XBP1s binds to the IL-6 promoter and
drives its expression in Hep3B cells
To explore the underlying mechanisms by which XBP1s
promotes IL-6 expression, potential promoter sequences of mouse and
human IL-6 were analyzed. Notably, a putative UPR element
for XBP1 binding was highly conserved in both mouse and human
IL-6, and contained the ‘ACGT’ core sequence as reported
previously (Fig. 3A) (25).
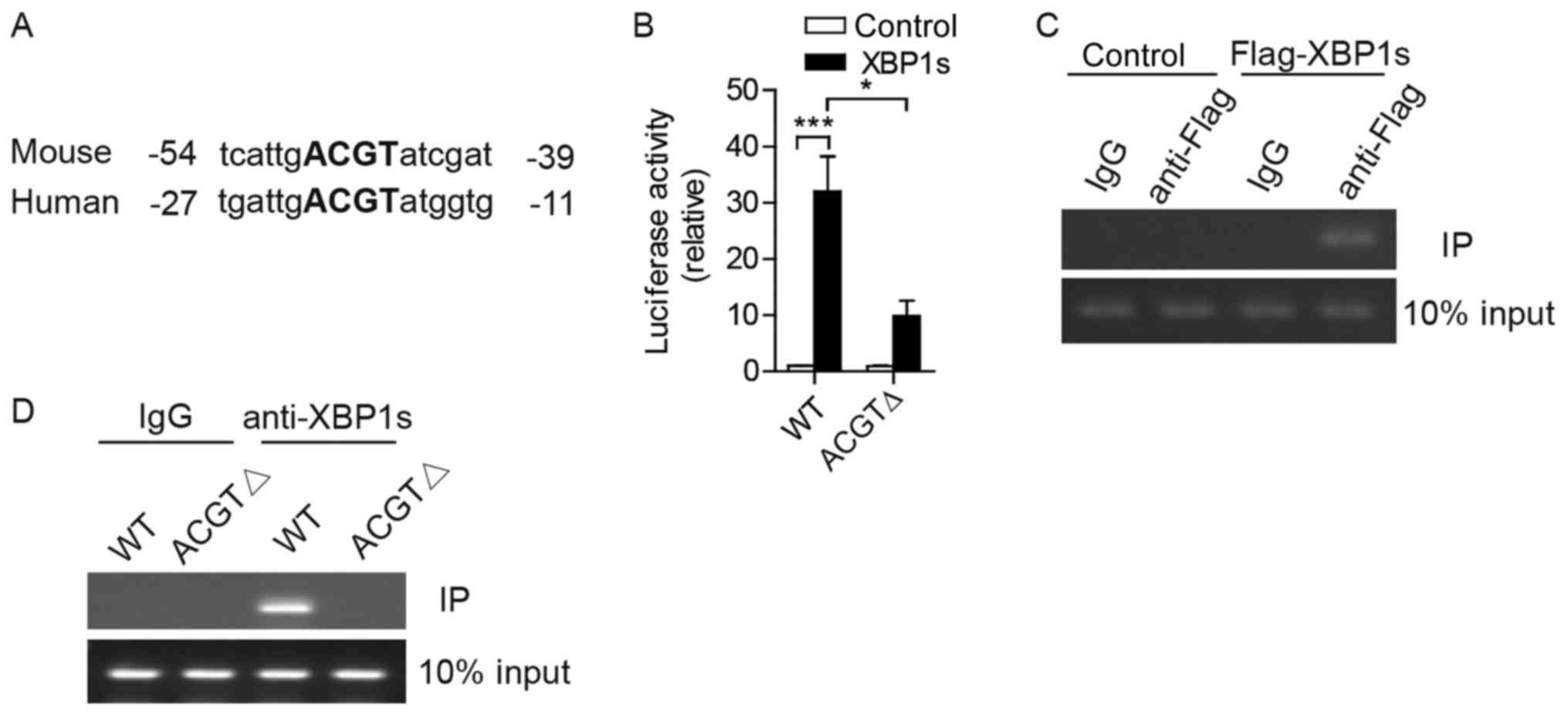 | Figure 3.XBP1s binds to IL-6 promoter
to activate its expression in Hep3B cells. (A) Alignment of the
sequences of IL-6 promoters to find the putative UPR element
from the mouse and human. The ‘ACGT’ core is indicated in bold. (B)
Luciferase reporter assays were performed in 293T cells
co-transfected with plasmids of pCMV-XBP1s together with luciferase
reporter constructs controlled by the human IL-6 promoter
(WT) or IL-6 promoter without ACGT core (ΔACGT). (C) ChIP
assays were conducted using IgG as the control or anti-Flag
antibody in extracts from Hep3B cells transfected with plasmids of
Flag-tagged XBP1s or vector control. The figure indicates
representative results of PCR, which was performed to amplify the
indicated region of the IL-6 promoter. (D) ChIP assays were
performed in extracts from 293T cells co-transfected with plasmids
of pCMV-XBP1s together with the plasmids of human IL-6
promoter (WT) or IL-6 promoter without ACGT core using IgG
or anti-XBP1s antibodies. The figure indicates representative
results of PCR, which was performed to amplify the indicated region
of the IL-6 promoter. Results are from more than three
independent experiments. Data are presented as the mean ± standard
error of the mean. *P<0.05, ***P<0.001 by two-way analysis of
variance. UPR, unfolded protein response; XBP1, X-box-binding
protein 1; IL-6, interleukin-6; PCR, polymerase chain reaction; WT,
wild type; IgG, immunoglobulin G; ChIP, chromatin
immunoprecipitation |
To investigate if this core sequence was important
in XBP1s-activated IL-6 expression, luciferase reporter
plasmids were constructed containing human IL-6 promoter of
full length (WT) or with ‘ACGT’ deletion (ΔACGT). A reporter assay
was performed in 293T cells. Transcriptional activity of the
IL-6 promoter was markedly enhanced in cells with ectopic
expression of XBP1s, and this effect was diminished when the ‘ACGT’
core sequence was deleted (Fig.
3B).
To determine whether XBP1s directly binds to the
IL-6 promoter, a ChIP assay was subsequently conducted. Notably,
exogenous Flag-tagged XBP1s proteins were co-immunoprecipitated
with chromatin, including a putative IL-6 promoter, using
anti-Flag antibodies in Hep3B cells (Fig.
3C). In 293T cells, exogenous XBP1s interacted with the DNA of
the IL-6 promoter (Fig. 3C).
Following the deletion of the ‘ACGT’ core sequence (ΔACGT) from the
IL-6 promoter, XBP1s proteins lost the ability to bind to the
IL-6 promoter (Fig. 3D). In
summary, these results demonstrate that XBP1s binds directly to the
IL-6 promoter and activates its transcription in human HCC
cells.
Effect of the IRE1α-XBP1 branch of UPR
on Hep3B cell proliferation is dependent on IL-6 signaling
To investigate the function of upregulated
extracellular IL-6, CCK8 assays and BrdU assays were performed to
assess the proliferation of Hep3B cells (Fig. 4). Notably, overexpression of XBP1s
markedly elevated the proliferation of Hep3B cells, but this effect
was diminished when tocilizumab, a humanized monoclonal antibody
against the IL-6 receptor, was added (Fig. 4A and B). Consistent with this, a
similar phenomenon of cell proliferation was observed in
IRE1α-overexpressing Hep3B cells (Fig. 4C
and D). These results suggest a critical role of IL-6 signaling
in HCC cell proliferation, which may be promoted by the IRE1α-XBP1
branch of UPR.
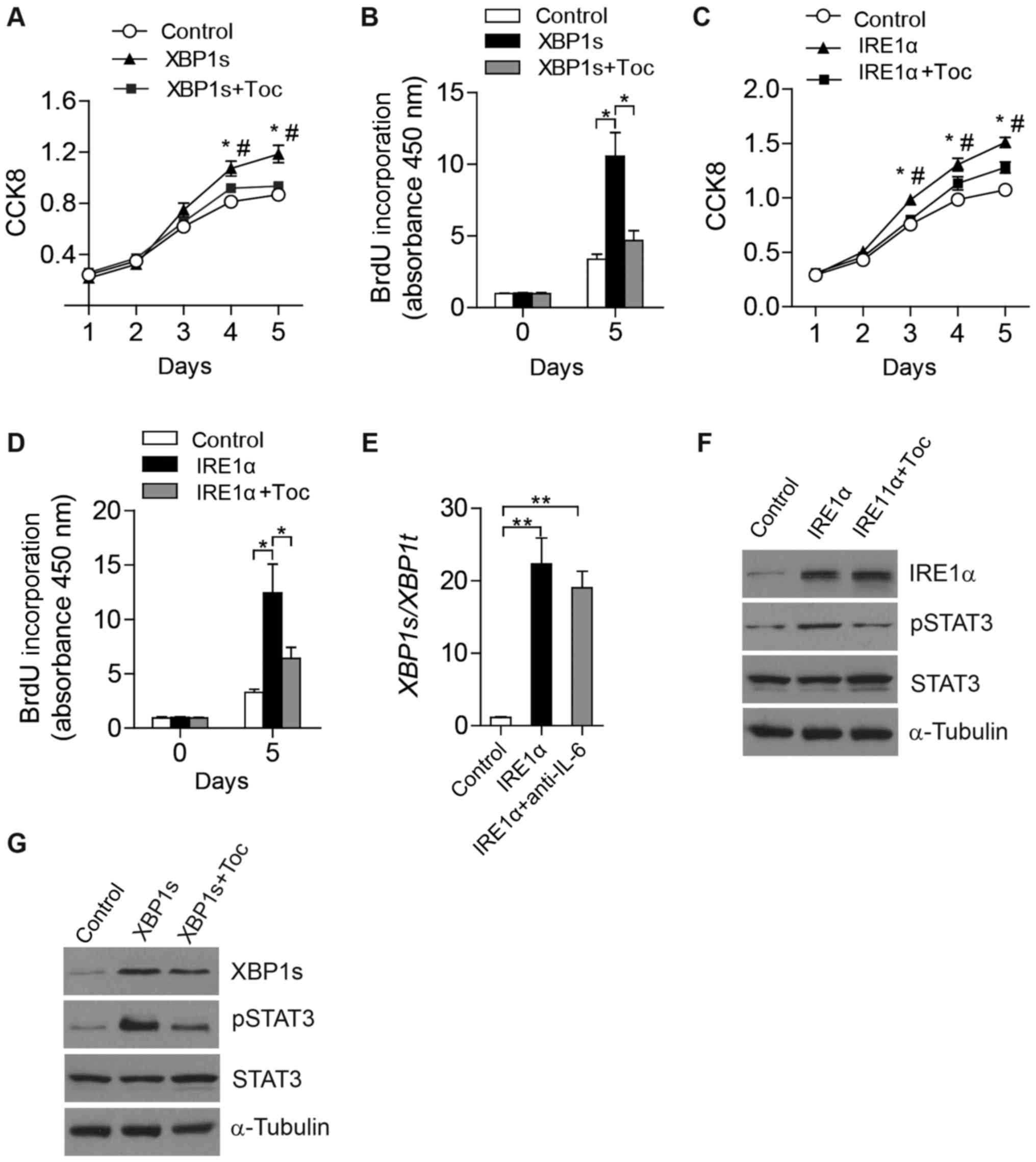 | Figure 4.IRE1α-XBP1 pathway promotes Hep3B
cell proliferation via regulating IL-6-STAT3 signaling. Cell
proliferation was analyzed by (A) CCK8 assays and (B) BrdU assays
in Hep3B cells that were transfected with plasmids of pCMV-XBP1s or
vector control and then treated with Toc. Cell proliferation was
analyzed by (C) CCK8 assays and (D) BrdU assays in Hep3B cells that
were transfected with plasmids of pCMV-IRE1α or vector control and
then treated with Toc. XBP1 splicing levels were determined
by (E) quantitative PCR and (F) levels of indicated proteins were
analyzed by western blotting in Hep3B cells that were transfected
with plasmids of pCMV-IRE1α or vector control and then treated with
Toc. (G) Indicated protein levels were analyzed by western blotting
in Hep3B cells transfected with pCMV-XBP1s or vector control and
then treated with Toc. Results are from more than three independent
experiments. Data are presented as the mean ± standard error of the
mean. (A and C) *P<0.05, **P<0.01 vs. ctrl;
#P<0.05 vs. XBP1s + Toc or IRE1α + Toc by one-way
ANOVA with Tukey's post hoc tests. (B and D) *P<0.05, as
indicated, by two-way ANOVA. (E) **P<0.01 vs. control, by
one-way ANOVA relative to ctrl. IRE1α, inositol-requiring enzyme 1α
XBP1, X-box-binding protein 1; PCR, polymerase chain reaction; Toc,
tocilizumab; ANOVA, analysis of variance; IL-6, interleukin-6;
STAT3, signal transducer and activator of transcription 3. |
Tocilizumab attenuates the effect of
the IRE1α-XBP1 branch on activating STAT3 signaling in Hep3B
cells
To determine whether the IRE1α-XBP1 branch of UPR
regulates the activation of IL-6 signaling, levels of STAT3
phosphorylation were evaluated in Hep3B cells that were
overexpressing IRE1α or XBP1s. Notably, markedly increased STAT3
phosphorylation was detected following ectopic expression of IRE1α
(Fig. 4F) or XBP1s (Fig. 4G). To determine if upregulated
extracellular IL-6 activated STAT3 signaling, tocilizumab was added
to block the interaction between IL-6 and its receptor. The
addition of tocilizumab did not exhibit any effects on the
intracellular levels of IRE1α protein (Fig. 4F), or XBP1s mRNA (Fig. 4E) or protein (Fig. 4G). As expected, increased STAT3
phosphorylation induced by overexpressed IRE1α or XBP1s was
diminished following treatment of Hep3B cells with tocilizumab
(Fig. 4F and G). In summary, these
data demonstrate that the IRE1α-XBP1 pathway regulates the
activation of STAT3 signaling by increasing IL-6 expression and
secretion in HCC cells.
Discussion
ER stress and UPR pathways are implicated to be
essential in the development of HCC (4), but the exact mechanisms of this have not
yet been elucidated. The current study reveals a novel and critical
function of IRE1α-XBP1 signaling in HCC progression. Liu et
al (26) recently reported that
IRE1α is essential in controlling hepatocyte proliferation and
liver regeneration via regulation of the STAT3 pathway.
Furthermore, IRE1α has also been reported to be implicated in
promoting cell proliferation of obesityinduced pancreatic islet
cells (27) and certain cancer cell
lines (28). However, whether the
IRE1α-XBP1 branch of UPR is linked to HCC cell proliferation
remains unclear. To the best of our knowledge, the current study
was the first to demonstrate a critical role of the IRE1α-XBP1
pathway in promoting the proliferation of HCC cells and the
underlying molecular mechanism of this.
In the current study, increased splicing levels of
XBP1 were detected in human HCC tissues and HCC cell lines compared
with normal liver tissues or hepatocyte cell lines. Furthermore,
hepatic IL-6 content exhibited a positive correlation with XBP1
splicing. Although IL-6 was mainly from resident immune cells,
hepatocytes also contribute to the total IL-6 in the local
microenvironment of the liver, which promotes the compensatory
proliferation of hepatocytes, particularly during the progression
of tumors (5).
In the current study, it was demonstrated that XBP1s
could bind to the IL-6 promoter and activate its
transcription in human HCC cells, indicating a highly conserved
role of XBP1 in controlling IL-6 expression. As a key component
downstream of IRE1α signaling, XBP1 usually acts as a potent
transcription activator and mediates the transcription of numerous
genes to relieve ER stress and restore ER homeostasis (11–13,29). An
increasing number of studies has revealed that the IRE1α-XBP1
pathway is also involved in the regulation of various physiological
processes, in addition to ER stress, via activation of gene
expression, including fatty acid synthase (30), peroxisome proliferator-activated
receptor α (31), protein disulphide
isomerase (32) and
UDP-galactose-4-epimerase (33), or
via non-transcriptional activity, such as promoting degradation of
the forkhead box O1 (34).
Furthermore, in a study of murine macrophages in innate immunology,
XBP1s was demonstrated to bind to the Il-6 promoter and
activate its transcription upon LPS stimulation (22). Consistent with these findings, it was
also identified in the current study that XBP1s worked as a
transcriptional activator in regulating IL-6 expression during the
development of HCC. Additionally, 4µ8C blocked the generation of
XBP1s and thus attenuated the IL-6 expression and secretion that
was induced by IRE1α overexpression, indicating the importance of
IRE1α RNase activity in controlling IL-6 expression. Argemí et
al (23) demonstrated that IL-6
could induce the expression of XBP1 during liver regeneration.
Combined with the current data, this indicates a positive feedback
loop; XBP1s activates IL-6 expression and IL-6 induces more XBP1 to
be spliced into XBP1s. Further studies are required to elucidate
the complex interactions between IL-6 and XBP1 in the liver.
IL-6 mRNA transcription and secretion were
increased in LO2 and Hep3B cells following ectopic expression of
IRE1α and XBP1s. This induced the activation of intracellular STAT3
signaling in an autocrine/paracrine manner, which could be
abolished by blocking the IL-6 receptor. The activation of
IL-6-STAT3 signaling by the IRE1α-XBP1 pathway was also
demonstrated to promote Hep3B cell proliferation. These results
were consistent with the critical role of IL-6-STAT3 signaling in
regulating cell proliferation and tissue regeneration (18–20),
particularly in HCC (3,5,35). It is
worth noting that XBP1s was recently identified to upregulate the
expression of STAT3 during liver regeneration (23), suggesting that XBP1s could also
amplify the activation of STAT3 signaling, as well as driving IL-6
expression in HCC. Furthermore, the addition of IL-6 receptor
antibodies (tocilizumab) diminished the effect of IRE1α-XBP1
signaling in activating STAT3 phosphorylation and promoting the
proliferation of Hep3B cells in the current in vitro
results. An in vivo study is required to explore whether
tocilizumab, an immunosuppressive drug for the treatment of
rheumatoid arthritis, has a potential function in inhibiting the
progression of liver cancer.
In summary, the present study reveals that the
IRE1α-XBP1 branch of UPR promotes cell proliferation and
progression of HCC via upregulation of IL-6 expression and
activation of IL-6-STAT3 signaling. Although further research is
required to verify the role of the IRE1α-XBP1 pathway in HCC
development in vivo, the current study provides a novel
promising therapeutic target for drug discovery and suggests that
tocilizumab may have an application in the clinical treatment of
patients with HCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by Zhejiang
Provincial Natural Science Foundation of China (grant no.
LY17H160054).
Availability of data and materials
The datasets used and/or analyzed during the current
study available from the corresponding author on reasonable
request.
Authors' contributions
PF, LX, SH, CP and YZ conceived and designed the
study. PF, LX, SH and CP conducted the majority of the experiments
and analyzed the data. LJ, GZ and LZhu performed some of the
cellular experiments. HF and LZho analyzed the data from the human
tissues. PF, CP and YZ wrote the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Second Affiliated Hospital and Yuying Children's
Hospital, Wenzhou Medical University and written informed consent
was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taniguchi K and Karin M: IL-6 and related
cytokines as the critical lynchpins between inflammation and
cancer. Semin Immunol. 26:54–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakagawa H, Umemura A, Taniguchi K,
Font-Burgada J, Dhar D, Ogata H, Zhong Z, Valasek MA, Seki E,
Hidalgo J, et al: ER stress cooperates with hypernutrition to
trigger TNF-dependent spontaneous HCC development. Cancer Cell.
26:331–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park EJ, Lee JH, Yu GY, He G, Ali SR,
Holzer RG, Osterreicher CH, Takahashi H and Karin M: Dietary and
genetic obesity promote liver inflammation and tumorigenesis by
enhancing IL-6 and TNF expression. Cell. 140:197–208. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sakurai T, He G, Matsuzawa A, Yu GY, Maeda
S, Hardiman G and Karin M: Hepatocyte necrosis induced by oxidative
stress and IL-1 alpha release mediate carcinogen-induced
compensatory proliferation and liver tumorigenesis. Cancer Cell.
14:156–165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Trikha M, Corringham R, Klein B and Rossi
JF: Targeted anti-interleukin-6 monoclonal antibody therapy for
cancer: A review of the rationale and clinical evidence. Clin
Cancer Res. 9:4653–4665. 2003.PubMed/NCBI
|
|
8
|
Tilg H, Wilmer A, Vogel W, Herold M,
Nölchen B, Judmaier G and Huber C: Serum levels of cytokines in
chronic liver diseases. Gastroenterology. 103:264–274. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wong VW, Yu J, Cheng AS, Wong GL, Chan HY,
Chu ES, Ng EK, Chan FK, Sung JJ and Chan HL: High serum
interleukin-6 level predicts future hepatocellular carcinoma
development in patients with chronic hepatitis B. Int J Cancer.
124:2766–2770. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakagawa H, Maeda S, Yoshida H, Tateishi
R, Masuzaki R, Ohki T, Hayakawa Y, Kinoshita H, Yamakado M, Kato N,
et al: Serum IL-6 levels and the risk for hepatocarcinogenesis in
chronic hepatitis C patients: An analysis based on gender
differences. Int J Cancer. 125:2264–2269. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schröder M and Kaufman RJ: The mammalian
unfolded protein response. Annu Rev Biochem. 74:739–789. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Walter P and Ron D: The unfolded protein
response: From stress pathway to homeostatic regulation. Science.
334:1081–1086. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoshida H, Matsui T, Yamamoto A, Okada T
and Mori K: XBP1 mRNA is induced by ATF6 and spliced by IRE1 in
response to ER stress to produce a highly active transcription
factor. Cell. 107:881–891. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang CC, Yang F, Thorne RF, Zhu BK,
Hersey P and Zhang XD: Human melanoma cells under endoplasmic
reticulum stress acquire resistance to microtubule-targeting drugs
through XBP-1-mediated activation of Akt. Neoplasia. 11:436–447.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ghosh R, Wang L, Wang ES, Perera BG,
Igbaria A, Morita S, Prado K, Thamsen M, Caswell D, Macias H, et
al: Allosteric inhibition of the IRE1α RNase preserves cell
viability and function during endoplasmic reticulum stress. Cell.
158:534–548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xue Z, He Y, Ye K, Gu Z, Mao Y and Qi L: A
conserved structural determinant located at the interdomain region
of mammalian inositol-requiring enzyme 1alpha. J Biol Chem.
286:30859–30866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding BB, Yu JJ, Yu RY, Mendez LM,
Shaknovich R, Zhang Y, Cattoretti G and Ye BH: Constitutively
activated STAT3 promotes cell proliferation and survival in the
activated B-cell subtype of diffuse large B-cell lymphomas. Blood.
111:1515–1523. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Niu G, Bowman T, Huang M, Shivers S,
Reintgen D, Daud A, Chang A, Kraker A, Jove R and Yu H: Roles of
activated Src and Stat3 signaling in melanoma tumor cell growth.
Oncogene. 21:7001–7010. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin L, Liu A, Peng Z, Lin HJ, Li PK, Li C
and Lin J: STAT3 is necessary for proliferation and survival in
colon cancer-initiating cells. Cancer Res. 71:7226–7237. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen C and Zhang X: IRE1α-XBP1 pathway
promotes melanoma progression by regulating IL-6/STAT3 signaling. J
Transl Med. 15:422017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martinon F, Chen X, Lee AH and Glimcher
LH: TLR activation of the transcription factor XBP1 regulates
innate immune responses in macrophages. Nat Immunol. 11:411–418.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Argemí J, Kress TR, Chang HCY, Ferrero R,
Bértolo C, Moreno H, González-Aparicio M, Uriarte I, Guembe L,
Segura V, et al: X-box binding protein 1 regulates unfolded
protein, acute-phase, and DNA damage responses during regeneration
of mouse liver. Gastroenterology. 152:1203–1216.e15. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shan B, Wang X, Wu Y, Xu C, Xia Z, Dai J,
Shao M, Zhao F, He S, Yang L, et al: The metabolic ER stress sensor
IRE1α suppresses alternative activation of macrophages and impairs
energy expenditure in obesity. Nat Immunol. 18:519–529. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kanemoto S, Kondo S, Ogata M, Murakami T,
Urano F and Imaizumi K: XBP1 activates the transcription of its
target genes via an ACGT core sequence under ER stress. Biochem
Biophys Res Commun. 331:1146–1153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Shao M, Wu Y, Yan C, Jiang S, Liu
J, Dai J, Yang L, Li J, Jia W, et al: Role for the endoplasmic
reticulum stress sensor IRE1α in liver regenerative responses. J
Hepatol. 62:590–598. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu T, Yang L, Yan C, Wang X, Huang P, Zhao
F, Zhao L, Zhang M, Jia W, Wang X and Liu Y: The IRE1α-XBP1 pathway
regulates metabolic stress-induced compensatory proliferation of
pancreatic β-cells. Cell Res. 24:1137–1140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thorpe JA and Schwarze SR: IRE1αlpha
controls cyclin A1 expression and promotes cell proliferation
through XBP-1. Cell Stress Chaperones. 15:497–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin JH, Li H, Yasumura D, Cohen HR, Zhang
C, Panning B, Shokat KM, Lavail MM and Walter P: IRE1 signaling
affects cell fate during the unfolded protein response. Science.
318:944–949. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee AH, Scapa EF, Cohen DE and Glimcher
LH: Regulation of hepatic lipogenesis by the transcription factor
XBP1. Science. 320:1492–1496. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shao M, Shan B, Liu Y, Deng Y, Yan C, Wu
Y, Mao T, Qiu Y, Zhou Y, Jiang S, et al: Hepatic IRE1α regulates
fasting-induced metabolic adaptive programs through the XBP1s-PPARα
axis signalling. Nat Commun. 5:35282014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang S, Chen Z, Lam V, Han J, Hassler J,
Finck BN, Davidson NO and Kaufman RJ: IRE1α-XBP1s induces PDI
expression to increase MTP activity for hepatic VLDL assembly and
lipid homeostasis. Cell Metab. 16:473–486. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Deng Y, Wang ZV, Tao C, Gao N, Holland WL,
Ferdous A, Repa JJ, Liang G, Ye J, Lehrman MA, et al: The
Xbp1s/GalE axis links ER stress to postprandial hepatic metabolism.
J Clin Invest. 123:455–468. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou Y, Lee J, Reno CM, Sun C, Park SW,
Chung J, Lee J, Fisher SJ, White MF, Biddinger SB and Ozcan U:
Regulation of glucose homeostasis through a XBP-1-FoxO1
interaction. Nat Med. 17:356–365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He G and Karin M: NF-κB and STAT3 - key
players in liver inflammation and cancer. Cell Res. 21:159–168.
2011. View Article : Google Scholar : PubMed/NCBI
|