Introduction
Approximately 15–20% of breast cancer cells are
triple-negative (TNBC cells) (1,2),
lacking estrogen receptors (ERs), progesterone receptors (PRs) and
epidermal growth factor receptor 2 (EGFR2). Expression of these
receptors allows for treatment with endocrine or targeted therapies
in clinical cases (3–5), which are not useful for clinical TNBC
cell treatment (6–8). Therefore, it is important to develop
new methods for suppressing TNBC cell growth and survival.
Methotrexate (MTX) is a well-known antagonist of folic acid
(9,10) and has been used widely for
rheumatoid arthritis treatment (11,12).
In addition, MTX has been applied for clinical cancer treatment
(13,14). Previous studies demonstrated that
MTX can inhibit the growth of various cancer cells, including
hepatoma, leukemia, lymphoma and gastric cancer cells (15–17).
Nevertheless, MTX alone is not effective for breast cancer
treatment. In order to enhance the anticancer activities of MTX on
breast cancer cells, combining MTX with other agents has been
considered. Currently, combined chemotherapy with MTX and other
anticancer drugs, such as mitomycin C, cyclophosphamide and
5-fluorouracil, is used to treat breast cancer (18–20).
However, serious side-effects of these chemicals have been reported
(21–25). Therefore, drugs that can promote the
anticancer activities of MTX with reduced side-effects are urgently
needed.
Vitamin C, a common nutrient, has anti-oxidative
(26,27) and anticancer activities (28,29).
Previous studies have also demonstrated that combined treatment
with vitamin C and conventional anticancer agents can enhance
anticancer activities (15,30,31).
Currently, vitamin C supplements are being applied for clinical
cancer therapy (32–34). However, vitamin C actually inhibits
tamoxifen-induced cell death in ER-positive breast cancer (35). Alternatively, high-dose vitamin C
alone can inhibit cancer cell growth, though the mechanisms remain
elusive (36–38). One study also showed that vitamin C
can attenuate the incidence of ER-positive breast cancer cells
(39). However, there is no
evidence demonstrating that vitamin C alone is useful for TNBC
treatment.
A recent study reported that vitamin C (30 µM to 4
mM) plus MTX can inhibit the growth of MCF-7 cells (an ER-positive
breast cancer cell line) and MDA-MB-231 cells (a type of TNBC)
through G2/M elongation and PI3K activation (30). However, the mechanisms of vitamin
C/MTX-induced cytotoxicity on breast cancer cells are still
unclear. Therefore, whether combined treatment with low-dose
vitamin C (5 µM) and MTX can inhibit TNBC cell growth and the
mechanisms of vitamin C/MTX-induced cytotoxicity were examined in
the present study.
Materials and methods
Materials
Vitamin C, Luminol, Lucigenin and Hoechst 33342 were
purchased from Sigma-Aldrich (St. Louis, MO, USA). Anti-tubulin
(1:1,000; cat. no. BS1699), anti-p38 (1:400; cat. no. BS3567) and
anti-p-p38 (1:400; cat. no. BS4766) primary rabbit polyclonal
antibodies were acquired from Bioworld Technology, Inc., (Louis
Park, MN, USA). Anti-cleaved PARP (1:2000; cat. no. 9544) and
anti-caspase-3 (1:1000; cat. no. 9965) primary rabbit polyclonal
antibodies and horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG secondary antibody (1:2,000, cat. no. 7074) were
from Cell Signaling Technology (Danvers, MA, USA). Tarceva
(Erlotinib) was purchased from Roche Ltd. (Kaiseraugst,
Switzerland). An MTT assay kit was obtained from Bio Basic Inc.
(Markham, ON, Canada). Fetal bovine serum (FBS), Dulbeccos modified
Eagles medium (DMEM), non-essential amino acids, L-glutamine and
penicillin/streptomycin were obtained from Gibco-BRL (Invitrogen
Life Technologies, Carlsbad, CA, USA).
Cell lines and culture
Triple-negative breast cancer cell lines (MDA-MB-231
and MDA-MB-468) were purchased from the Bioresource Collection and
Research Center (Hsin-chu, Taiwan). Tarceva-resistant MDA-MB-231
cells (MDA-MB-231 TR) were kindly provided by Dr Yung-Luen Yu
(Graduate Institute of Biomedical Sciences, China Medical
University, Taichung, Taiwan). These cells were maintained in a
humidified atmosphere containing 5% CO2 at 37°C and
cultured with DMEM supplemented with 10% FBS, 0.1 mM non-essential
amino acids, 2 mM L-glutamine and 100 IU/ml
penicillin/streptomycin. In addition, 100 µM tarceva was added to
the media for MDA-MB-231 TR culture.
Determination of cell viability
Cell viability was measured by the MTT assay
described in previous studies (40,41).
Briefly, cells were cultured into 96-well plates (5×103
cells/well). Every 24 h, the control and experimental groups were
treated with MTT. After incubation for 3 h at 37°C, the formazan
product was dissolved and absorbance measured at 570 nm (A570)
using a Multiskan™ FC microplate photometer (Molecular Devices,
Sunnyvale, CA, USA). The viable cell count (%) was calculated as
(A570 experimental group)/(A570 control group) × 100%.
Measurements of intracellular
H2O2 and O2−
Intracellular H2O2 and
O2− were measured using the
lucigenin-amplified chemiluminescence method (40,42).
The samples (200 µl) were added to 0.2 mmol/ml of luminol solution
(100 µl) for H2O2 measurement or to 0.1
mmol/ml lucigenin solution (500 µl) for O2−
measurement. Next, all samples were analyzed using a
chemiluminescence analyzing system (CLA-FSI; Tohoku Electronic
Industrial, Co., Ltd., Sendai, Japan). The
H2O2 and O2− were
observed and incubated for 5 min.
Observation of DNA fragmentation and
nuclear condensation
Nuclear condensation and DNA fragmentation, cardinal
characteristics of apoptotic cells, were observed using Hoechst
33342 nuclear staining (40,41).
Control and experimental (MTX and/or vitamin C-treated) cells were
incubated in Hoechst 33342 (10 µg/ml) for 5 min. DNA fragmentation
and nuclear condensation were observed under an Olympus DP71
fluorescence microscope (excitation, 352 nm; emission, 450 nm;
Olympus Corp., Tokyo, Japan).
SDS electrophoresis and western
blotting
Cells were lysed in radio-immunoprecipitation assay
(RIPA) buffer (cat. no. 20-188; EMD Millipore, Billerica, MA, USA).
Proteins were collected from the supernatant layer after
centrifugation (16,000 × g; 4°C) for 20 min. The protein
concentration was measured using a protein assay kit (cat. no.
23200; Thermo Fischer Scientific, Inc., Waltham, MA, USA). Equal
quantities (40 µg) of protein were separated by SDS-PAGE using
13.3% gels (80 volts) and transferred onto polyvinylidene
difluoride membranes (EMD Millipore). The membranes were blocked
with 5% non-fat milk at room temperature for 2 h then washed with
phosphate-buffered saline (PBS). After the incubation with primary
antibodies for 4 h, the membranes were washed with PBS and treated
with anti-rabbit HRP-conjugated secondary antibodies at room
temperature for 1 h. Finally, the immunolabeled proteins were
treated with Western Lightning® chemiluminescence Plus
reagent (Perkin-Elmer, Inc., Waltham, MA, USA) and observed with a
Luminescence Image Analysis system (LAS-4000; FujiFilm Electronic
Materials Taiwan, Co., Ltd., Tainan, Taiwan).
Statistical analysis
All data were obtained from four independent
experiments and presented as the mean ± SE. Means were compared by
Students t-test using Microsoft Excel (http://microsoft-excel-2010.updatestar.com/zh-tw).
A P<0.05 was considered statistically significant.
Results
Combined treatment with low-dose
vitamin C and MTX effectively inhibits TNBC cell proliferation and
viability
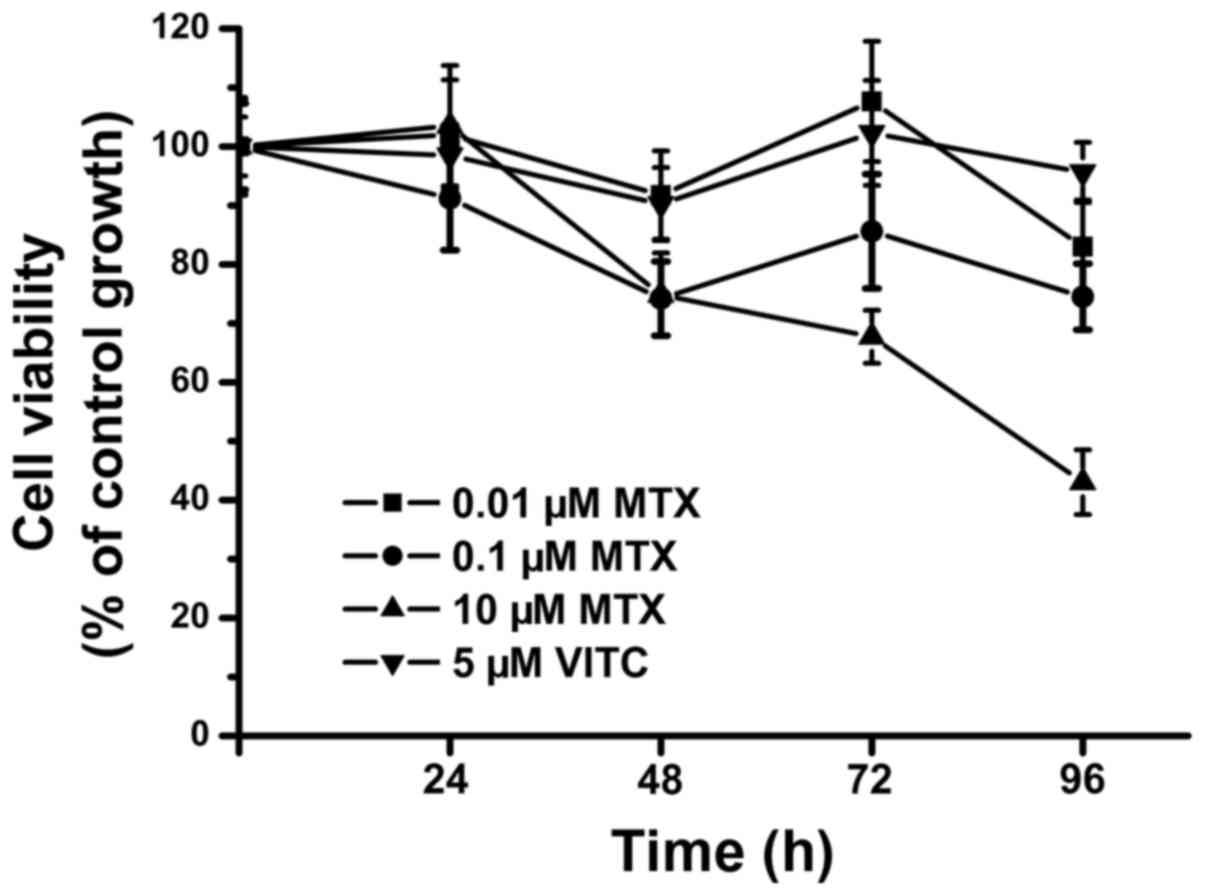
We first examined the effects of various
concentrations of MTX on TNBC cell (MDA-MB-231) growth and
survival. Low-dose MTX alone (0.1 and 0.01 µM) did not inhibit TNBC
cell growth after 96-h treatment, and cell viability as measured by
MIT assay was maintained at ~75–100% of control from 24 to 96 h
(Fig. 1). Only 10 µM MTX reliably
inhibited TNBC cell growth at 96 h, with cell viability <50%
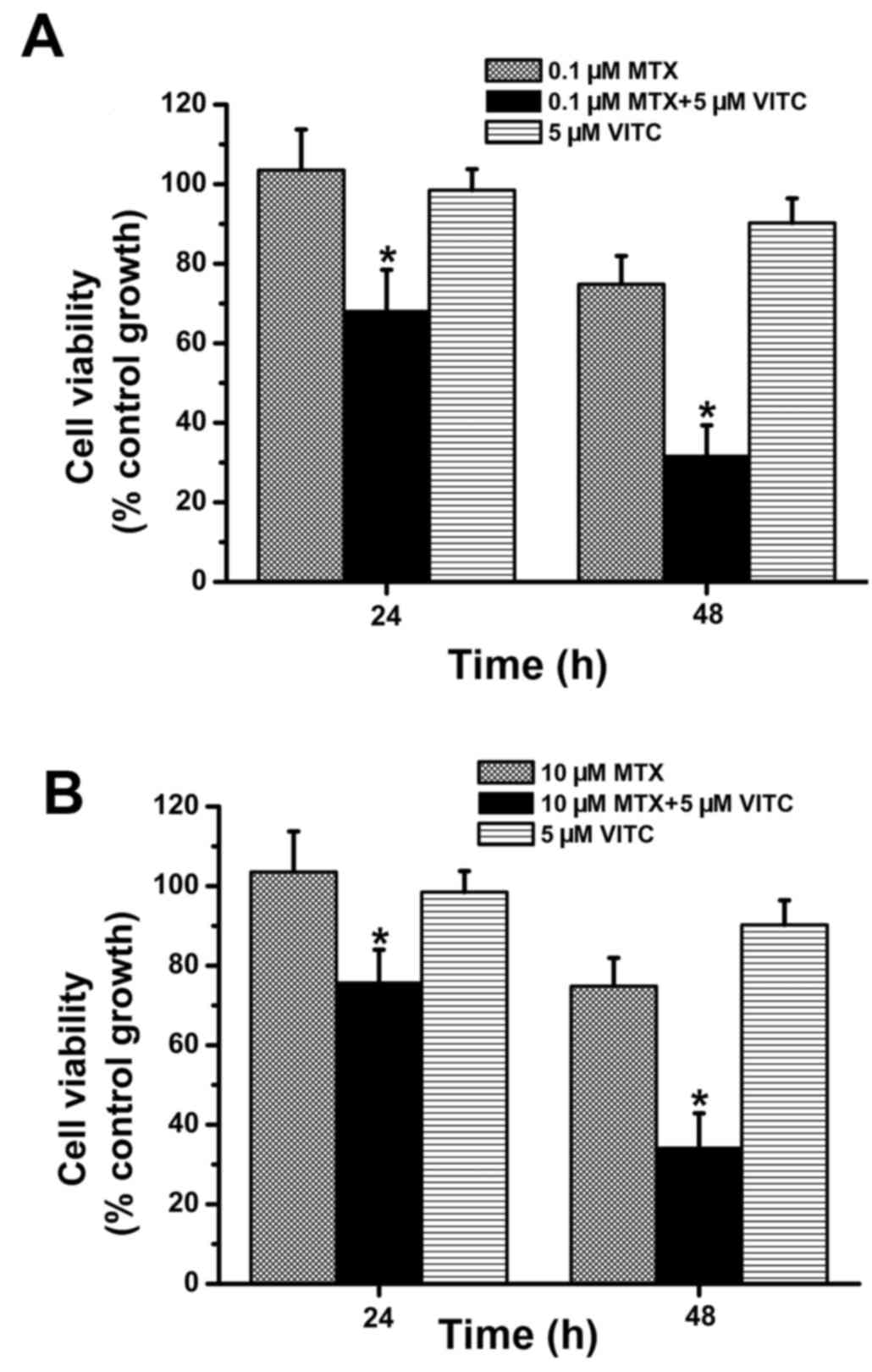
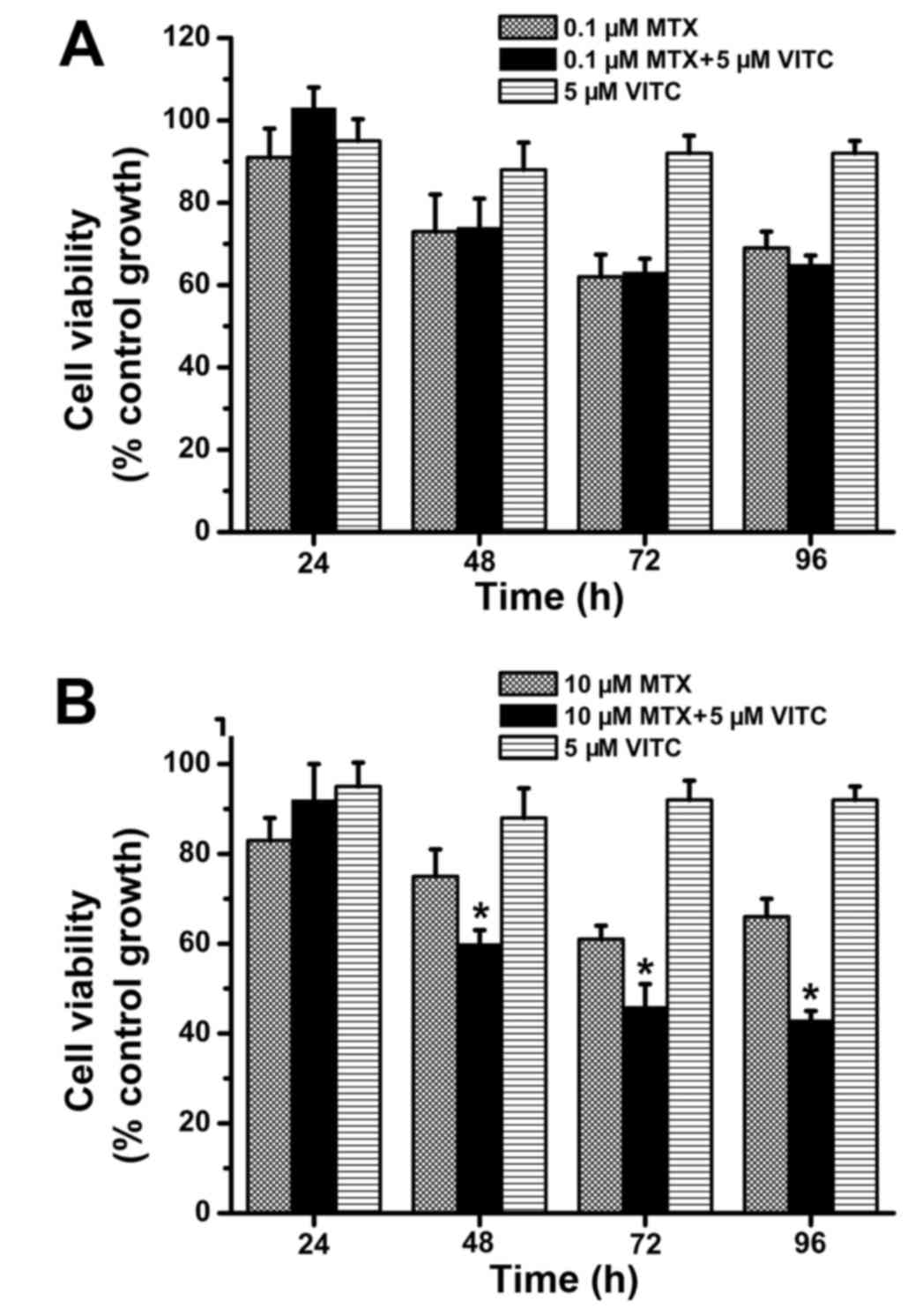
(Fig. 1). Next, combined treatment
with low-dose vitamin C (5 µM) and MTX was examined. As shown in
Fig. 2, compared to MTX-treated and
vitamin C-treated groups, the MTX plus vitamin C-treated group
exhibited significantly lower cell viability at 24 and 48 h. Cell
viability was >70% in all MTX-treated and vitamin C-treated
groups (Fig. 2), but <40% at 48
h in the 0.1 µM MTX plus vitamin C-treated group and 10 µM MTX plus
vitamin C-treated group (Fig. 2).
Overall, these data demonstrate that combined treatment with
low-dose vitamin C and MTX effectively inhibits TNBC cell
proliferation and survival.
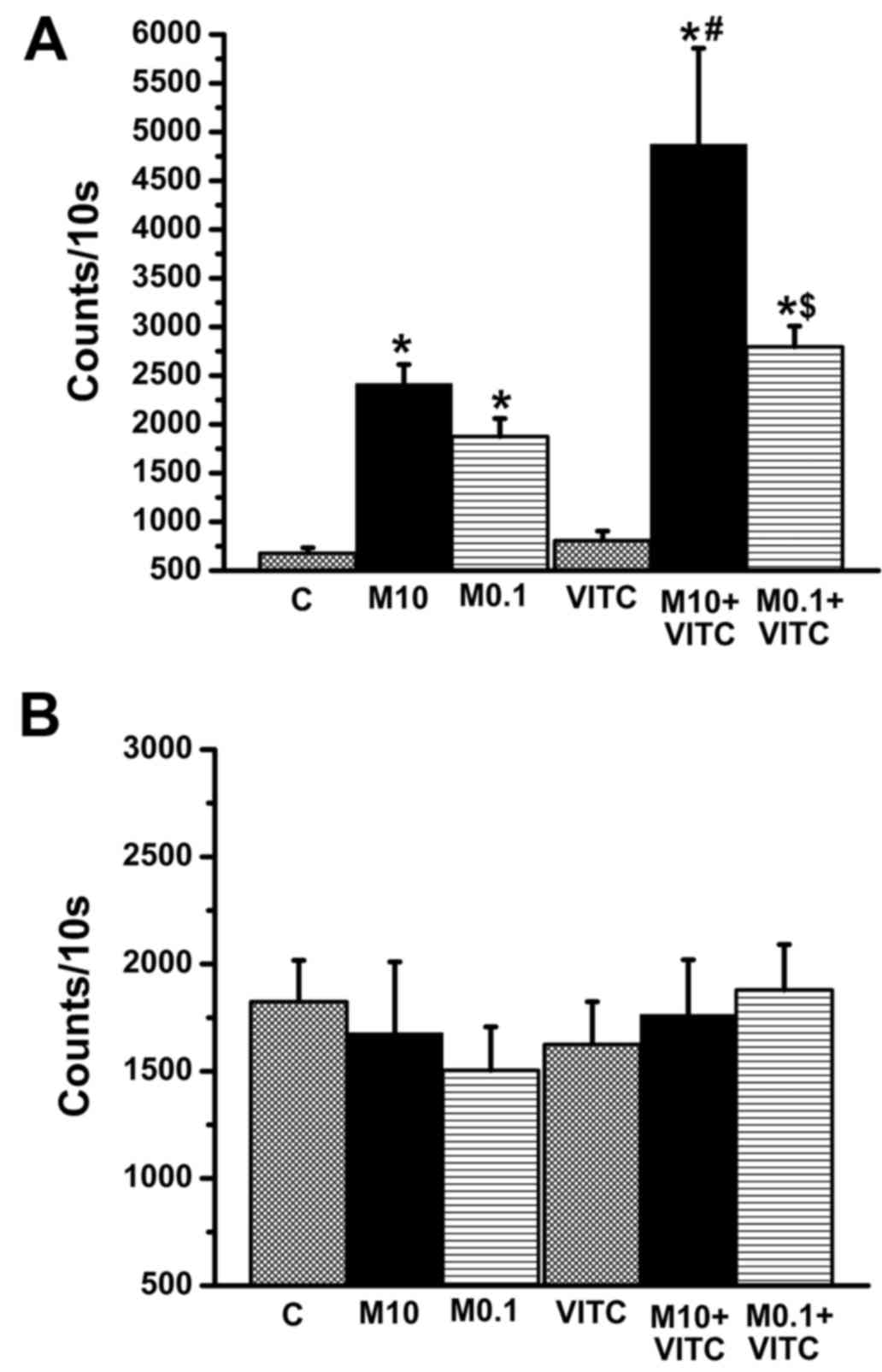
Vitamin C enhances MTX-induced
intracellular H2O2 accumulation
MTX can increase reactive oxygen species (ROS)
accumulation in cells with ensuing cytotoxicity (15,43).
In contrast, vitamin C is an anti-oxidant against ROS increase
(26,27). Both H2O2 and
O2− are major ROS species in cells.
Intracellular H2O2 and
O2− were compared among the control group,
MTX-treated group, vitamin C-treated group, and MTX plus vitamin
C-treated group (Fig. 3A).
Intracellular H2O2 levels were increased in
0.1 and 10 µM MTX groups, in accordance with a previous study
(15). Surprisingly, vitamin C did
not reduce H2O2 levels in MTX-treated groups.
Compared to MTX-treated groups, H2O2 levels
were increased significantly in the vitamin C plus MTX-treated
group. In contrast, O2− levels did not differ
among treatment groups (Fig. 3B).
Our data suggest that increased intracellular
H2O2 may contribute to the decrease in cell
viability induced by MTX plus vitamin co-treatment.
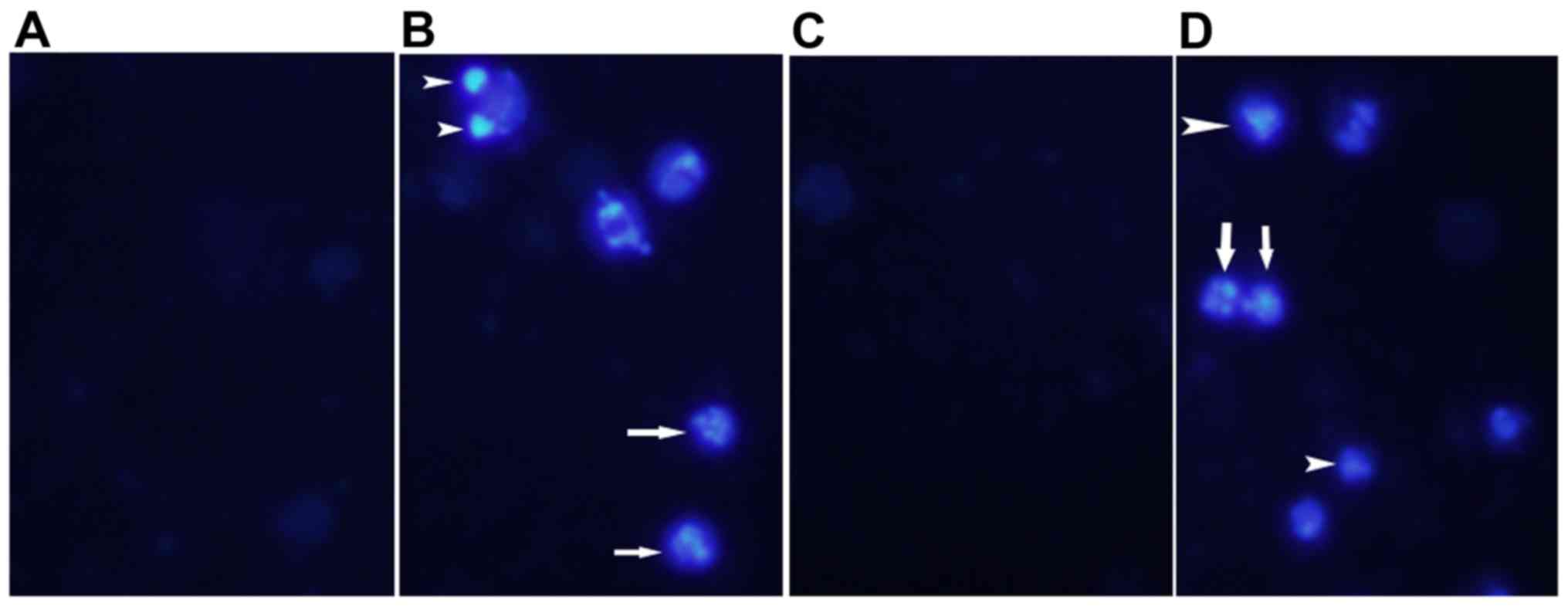
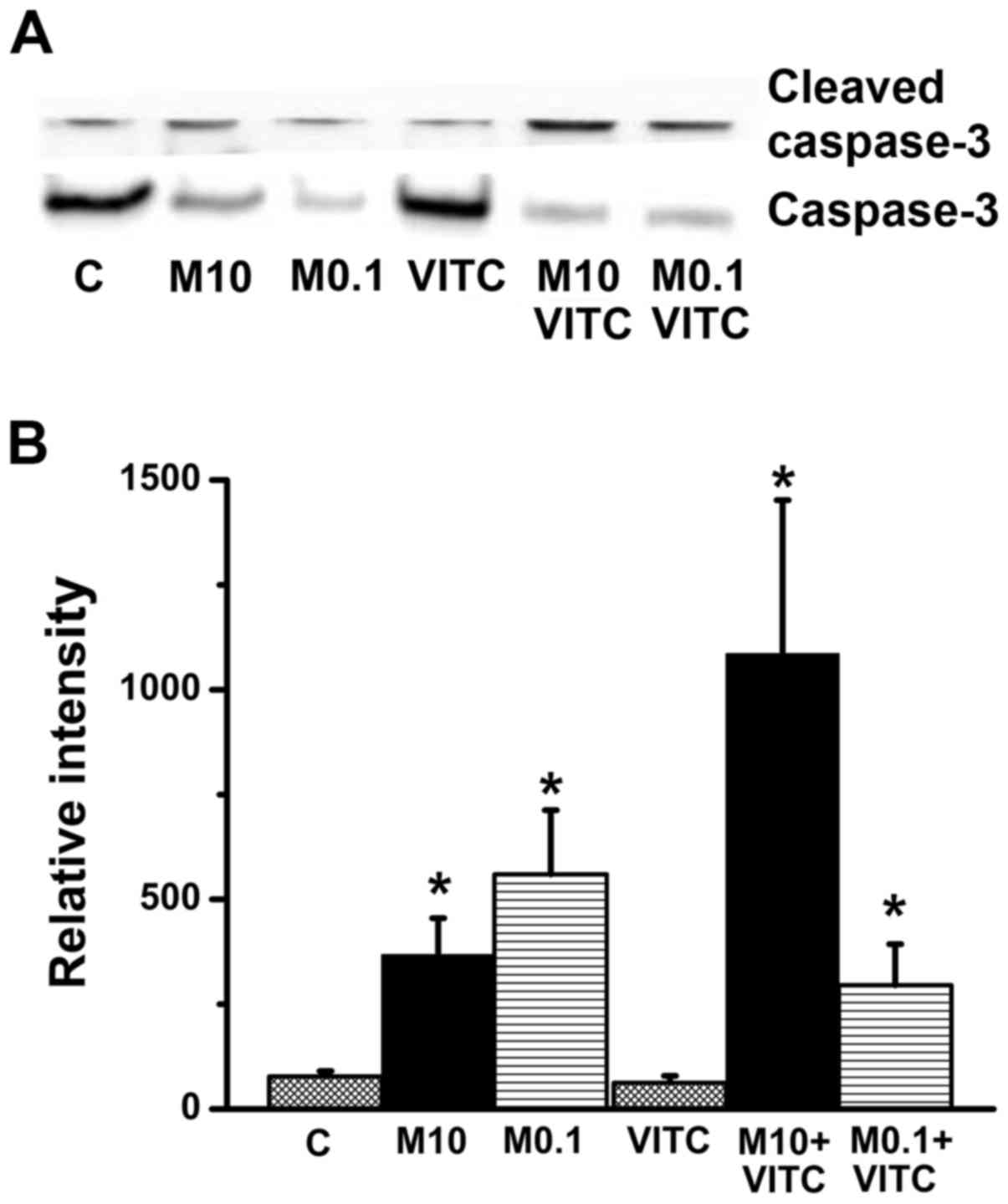
MTX-treatment and combined MTX plus
vitamin C treatment induce apoptosis and caspase-3 activation
Induction of apoptosis by these treatments was
assessed by nuclear staining. As shown in Fig. 4, nuclear condensation and DNA
fragmentation were observed in MTX-treated and MTX plus vitamin
C-treated groups. Apoptosis can be initiated by caspase-dependent
and caspase-independent pathways (44,45).
Therefore, caspase-3 activation was examined by western blotting.
As shown in Fig. 5, compared to the
control group, the ratio of cleaved (activated) caspase-3 to native
caspase-3 was increased significantly in both the MTX-treated and
MTX plus vitamin C-treated group. PARP is a downstream substrate of
caspase-3, thus, cleaved PARP is a sign of caspase-3 activation.
Indeed, cleaved PARP level was also increased in both MTX-treated
and MTX plus vitamin C-treated groups. Taken together, these
results indicate that combined MTX/vitamin C induces
caspase-3-dependent apoptosis in TNBC cells.
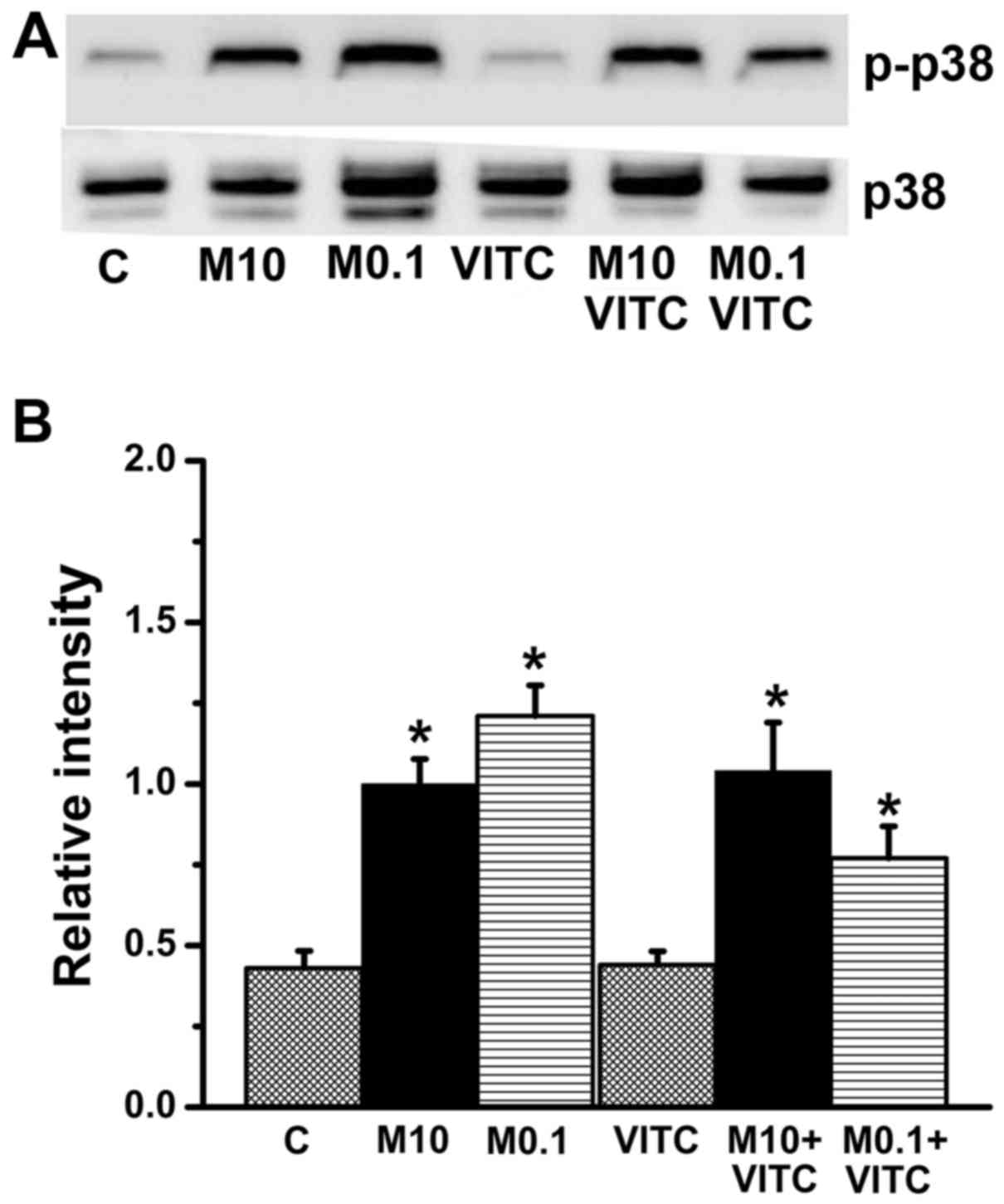
p38 phosphorylation in MTX-treated and
MTX plus vitamin C-treated cells
The MAPK family kinases ERK, JNK and p38 are
involved in cell death, cell differentiation, and cell
proliferation (46–48). In the present study, expression
levels of EKR, JNK, p38 and their phosphorylated (activated) forms
(p-ERK, p-JNK and p-p38) were estimated by western blotting. The
ratio of p-p38 to p38 was significantly increased in both
MTX-treated and MTX plus vitamin C-treated groups (Fig. 7), while ERK and JNK expression
levels did not differ significantly among groups (data not show).
Thus, MTX/vitamin C-induced cytotoxicity of TNBC cells is
associated with p38 activation.
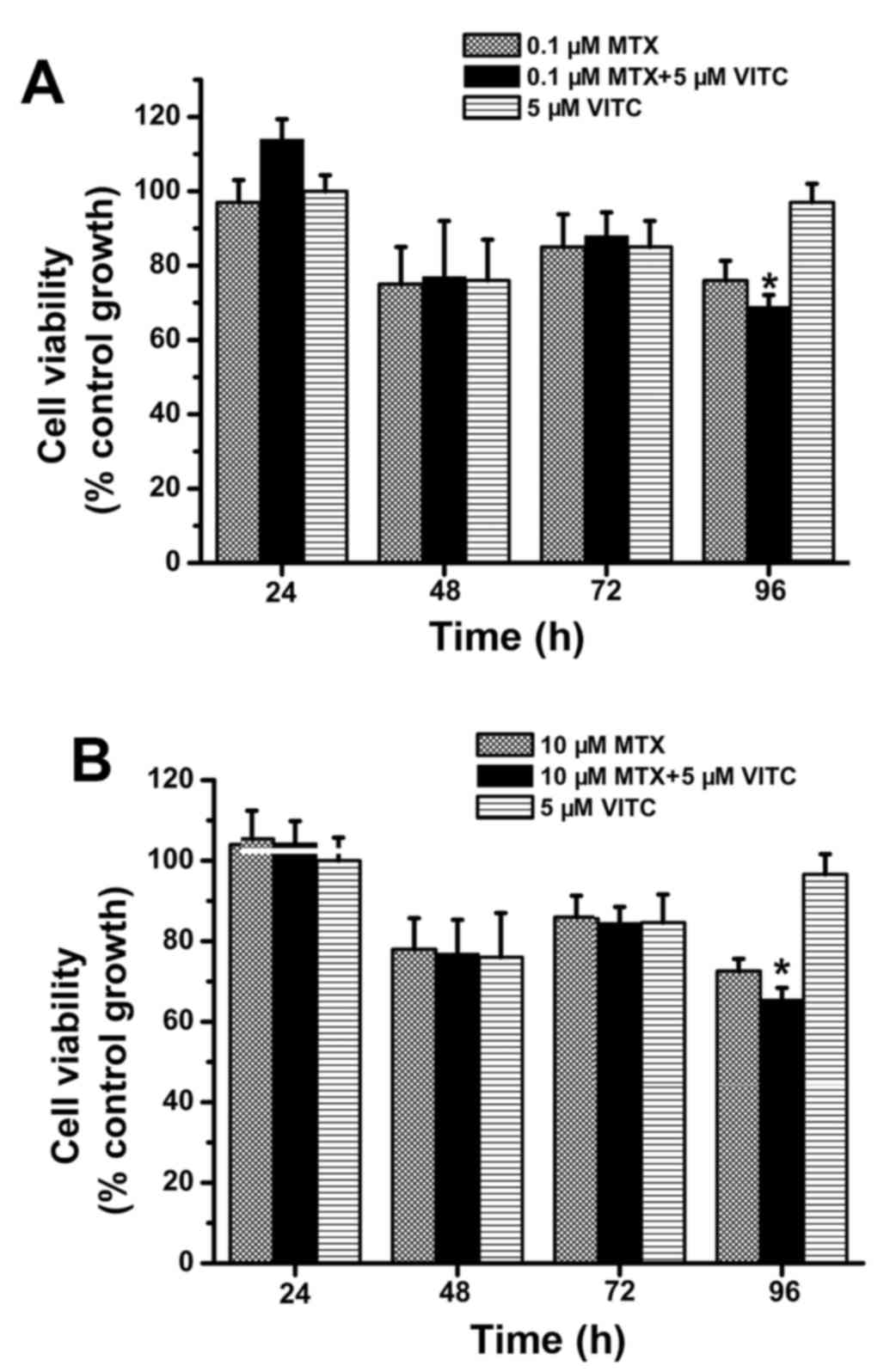
Combined treatment with vitamin C and
MTX inhibits growth of tarceva-sensitive, but not tarceva-resistant
TNBC cells
As shown in Fig. 2,
combined treatment with vitamin C and MTX effectively inhibited
MDA-MB-231 cell growth. However, it is unclear whether MTX plus
vitamin C is useful against other TNBC cells, thus, we examined the
cell viability of TNBC cell lines MDA-MB-468 and MDA-MB-231 TR
during MTX, vitamin C and combined treatment. Compared to the 10 µM
MTX-treated group, the combined treatment group exhibited lower
viability at 48, 72 and 96 h (Fig.
8B). Importantly, cell viability was <50% at 72 and 96 h in
the 10 µM MTX plus vitamin C-treated group (Fig. 8B). Alternatively, cell viability did
not differ among the control, 0.1 µM MTX-treated, and 0.1 µM MTX
plus vitamin C-treated groups (Fig.
8A). These data indicate that 1 µM MTX plus vitamin C can
effectively inhibit MDA-MB-468 proliferation/survival compared to
MTX treatment. However, in similar assays of tarceva-resistant
MDA-MB-231 TR cells, the MTX plus vitamin C-treated group exhibited
significantly lower cell viability only at 96 h and was >60% for
all other groups (Fig. 9). That is,
neither MTX alone nor MTX plus vitamin C inhibited the growth of
tarceva-resistant TNBC cells as effectively as tarceva-sensitive
TNBC cells. Taken together, these findings (Figs. 2, 8
and 9) suggest that MTX plus
vitamin C treatment can inhibit the growth of tarceva-responsive,
but not tarceva-resistant TNBC cells.
Discussion
MTX alone is not useful for breast cancer treatment,
but a recent study found that high-dose vitamin C (30 µM to 4 mM)
enhanced the anticancer activities of MTX on breast cancer cells,
including MCF-7 cells (ER-positive breast cancer) and MDA-MB-231
cells (30). That study also found
G2/M elongation and PI3K pathway activation associated with vitamin
C/MTX-induced cytotoxicity. The present study found that MTX had
anti-proliferative/cytotoxic effects on TNBC cells only when
combined with low-dose vitamin C (5 µM). The present study further
suggests that increased intracellular H2O2
levels and activation of caspase-3 and p38 pathways are involved in
vitamin C/MTX-induced cytotoxicity. In addition, combined treatment
with low-dose vitamin C and MTX inhibited cell growth not only of
MDA-MB-231 cells but also of MDA-MB-468 cells. Therefore, this
study suggests that vitamin C plus MTX treatment may be effective
for clinical suppression of TNBC cell growth.
Although a previous study (30) and the present study (Figs. 2 and 8) demonstrated that vitamin C plus MTX
effectively inhibits TNBC cell growth, vitamin C plus MTX treatment
did not effectively inhibit tarceva-resistant TNBC cells (Fig. 9). Tarceva (erlotinib) is an EGFR
tyrosine kinase inhibitor (49,50)
and has been applied for clinical treatment of lung and breast
cancers (51–54). These findings suggest that EGFR
signaling may be involved in vitamin C/MTX-induced
cytotoxicity.
Apoptosis can be induced via caspase-dependent and
caspase-independent pathways (44,45).
Vitamin C treatment can induce apoptosis of breast cancer cells and
lung cancer cells via the caspase-independent pathway (36,55,56).
On the other hand, vitamin C treatment can induce apoptosis of
melanoma cells and hepatoma cells via the caspase-dependent pathway
(15,57). In the present study, vitamin C plus
MTX treatment activated caspase-3 in TNBC cells. Thus, whether
apoptosis occurs via caspase-dependent or caspase-independent
pathways may depend on the specific cancer cell type.
At low doses, vitamin C has anti-oxidant activities
(26,27), and many studies have demonstrated
that vitamin C supplements can decrease oxidative stress (15,58,59).
However, high-dose (millimolar) vitamin C treatment can increase
oxidative stress (60–62), and previous studies showed that
high-dose vitamin C can enhance intracellular
H2O2, resulting in cancer cell death
(37,63,64).
Surprisingly, another study reported that low-dose (micromolar)
vitamin C attenuated H2O2 levels but enhanced
the anticancer activities of MTX on hepatoma cells (15). In addition, vitamin C/MTX only
enhances H2O2 levels but not
O2− levels in TNBC cells. These results
indicated vitamin C/MTX does not inhibit the function of superoxide
dismutase, while vitamin C/MTX may influence GSH levels or activity
of glutathione peroxidase.
As shown in Fig. 9,
the cell viability was not significantly different before 72 h
between MTX alone group and MTX plus vitamin C group. The cell
viability was significant different only at 96 h between MTX alone
group and MTX plus vitamin C group. The results indicated that
vitamin C is not efficient in enhancing MTX-induced cytotoxicity in
tarceva-resistant TNBC cells. Previously, many studies showed that
tarceva-resistant cells have EGFR gene mutations or additional
bypass signaling pathways to activate downstream of EGFR (65,66).
These mutation genes and bypass pathways are important factors for
cell proliferation with EGFR inhibitor treatment. These factors may
cause the vitamin C/MTX inefficiency in inducing cytotoxicity in
tarceva-resistant cells. However, many bypass signals are related
to tarceva-resistant cells, such as SOS1, NF-κB and Fas receptor,
signals of which influence vitamin C/MTX-induced cytotoxicity,
however, this remains to be studied in the future.
Collectively, we found that combined treatment with
low-dose vitamin C and MTX enhanced intracellular
H2O2 accumulation and suppressed TNBC cell
growth. Therefore, we suggest that both vitamin C dose and cell
type may influence cellular H2O2 levels
during treatment.
Acknowledgements
The present study was supported by funding from the
Ministry of Science and Technology, Taiwan (MOST103
2320-B-039-052-MY3; MOST104-2321-B-039-005), the Ministry of Health
and Welfare, Taiwan (MOHW104-TDU-B-212-124-002) and the Taipei Tzu
ChiHospital, Taiwan (TCRD-TPE-104-06; TCRD-TPE-104-34;
TCRD-TPE-105-20; TCRD-TPE-105-02).
References
|
1
|
Yu YL, Chou RH, Liang JH, Chang WJ, Su KJ,
Tseng YJ, Huang WC, Wang SC and Hung MC: Targeting the EGFR/PCNA
signaling suppresses tumor growth of triple-negative breast cancer
cells with cell-penetrating PCNA peptides. PLoS One. 8:e613622013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Metzger-Filho O, Tutt A, de Azambuja E,
Saini KS, Viale G, Loi S, Bradbury I, Bliss JM, Azim HA Jr, et al:
Dissecting the heterogeneity of triple-negative breast cancer. J
Clin Oncol. 30:1879–1787. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ojo D, Wei F, Liu Y, Wang E, Zhang H, Lin
X, Wong N, Bane A and Tang D: Factors promoting tamoxifen
resistance in breast cancer via stimulating breast cancer stem cell
expansion. Curr Med Chem. 22:2360–2374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Spring L, Bardia A and Modi S: Targeting
the cyclin D-cyclin-dependent kinase (CDK) 4/6-retinoblastoma
pathway with selective CDK 4/6 inhibitors in hormone
receptor-positive breast cancer: Rationale, current status, and
future directions. Discov Med. 21:65–74. 2016.PubMed/NCBI
|
|
5
|
Lumachi F, Chiara GB, Foltran L and Basso
SM: Proteomics as a guide for personalized adjuvant chemotherapy in
patients with early breast cancer. Cancer Genomics Proteomics.
12:385–390. 2015.PubMed/NCBI
|
|
6
|
Williams CB, Soloff AC, Ethier SP and Yeh
ES: Perspectives on epidermal growth factor receptor regulation in
triple-negative breast cancer: Ligand-mediated mechanisms of
receptor regulation and potential for clinical targeting. Adv
Cancer Res. 127:253–281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hudis CA and Gianni L: Triple-negative
breast cancer: An unmet medical need. Oncologist. 16:(Suppl 1).
1–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perou CM, Sørlie T, Eisen MB, van de Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schofield RC, Ramanathan LV, Murata K,
Fleisher M, Pessin MS and Carlow DC: Development of an assay for
methotrexate and its metabolites 7-hydroxy methotrexate and DAMPA
in serum by LC-MS/MS. Methods Mol Biol. 1383:213–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hara A, Taguchi A, Aoki H, Hatano Y, Niwa
M, Yamada Y and Kunisada T: Folate antagonist, methotrexate induces
neuronal differentiation of human embryonic stem cells transplanted
into nude mouse retina. Neurosci Lett. 477:138–143. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pavy S, Constantin A, Pham T, Gossec L,
Maillefert JF, Cantagrel A, Combe B, Flipo RM, Goupille P, Le Loët
X, et al: Methotrexate therapy for rheumatoid arthritis: clinical
practice guidelines based on published evidence and expert opinion.
Joint Bone Spine. 73:388–395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Whittle SL and Hughes RA: Folate
supplementation and methotrexate treatment in rheumatoid arthritis:
A review. Rheumatology (Oxford). 43:267–271. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho KM, Kim YJ, Kim SH, Kim JW, Lee JO,
Han JH, Lee KW, Kim JH, Kim CY, Bang SM, et al: Salvage treatment
with intracerebrospinal fluid thiotepa in patients with
leptomeningeal metastasis after failure of methotrexate-based
treatment. Anticancer Res. 35:5631–5638. 2015.PubMed/NCBI
|
|
14
|
van der Plas E, Nieman BJ, Butcher DT,
Hitzler JK, Weksberg R, Ito S and Schachar R: Neurocognitive late
effects of chemotherapy in survivors of acute lymphoblastic
leukemia: Focus on methotrexate. J Can Acad Child Adolesc
Psychiatry. 24:25–32. 2015.PubMed/NCBI
|
|
15
|
Yiang GT, Chou PL, Hung YT, Chen JN, Chang
WJ, Yu YL and Wei CW: Vitamin C enhances anticancer activity in
methotrexate-treated Hep3B hepatocellular carcinoma cells. Oncol
Rep. 32:1057–1063. 2014.PubMed/NCBI
|
|
16
|
Shirao K, Boku N, Yamada Y, Yamaguchi K,
Doi T, Goto M, Nasu J, Denda T, Hamamoto Y, Takashima A, et al:
Gastrointestinal Oncology Study Group of the Japan Clinical
Oncology Group: Randomized Phase III study of 5-fluorouracil
continuous infusion vs. sequential methotrexate and 5-fluorouracil
therapy in far advanced gastric cancer with peritoneal metastasis
(JCOG0106). Jpn J Clin Oncol. 43:972–980. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takemura Y and Jackman AL: Folate-based
thymidylate synthase inhibitors in cancer chemotherapy. Anticancer
Drugs. 8:3–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fukuda T, Tanabe M, Kobayashi K, Fukada I,
Takahashi S, Iwase T and Ito Y: Combination chemotherapy with
mitomycin C and methotrexate is active against metastatic
HER2-negative breast cancer even after treatment with
anthracycline, taxane, capecitabine, and vinorelbine. Springerplus.
4:3762015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leone JP, Leone J, Vallejo CT, Pérez JE,
Romero AO, Machiavelli MR, Romero Acuña L, Domínguez ME, Langui M,
Fasce HM, et al: Sixteen years follow-up results of a randomized
phase II trial of neoadjuvant fluorouracil, doxorubicin, and
cyclophosphamide (FAC) compared with cyclophosphamide,
methotrexate, and 5-fluorouracil (CMF) in stage III breast cancer:
GOCS experience. Breast Cancer Res Treat. 143:313–323. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu CE, Chen SC, Lin YC, Lo YF, Hsueh S and
Chang HK: Identification of patients with node-negative,
triple-negative breast cancer who benefit from adjuvant
cyclophosphamide, methotrexate, and 5-fluorouracil chemotherapy.
Anticancer Res. 34:1301–1306. 2014.PubMed/NCBI
|
|
21
|
Shea B, Swinden MV, Ghogomu ET, Ortiz Z,
Katchamart W, Rader T, Bombardier C, Wells GA and Tugwell P: Folic
acid and folinic acid for reducing side effects in patients
receiving methotrexate for rheumatoid arthritis. J Rheumatol.
41:1049–1060. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee HJ, Hong SK, Seo JK, Lee D and Sung
HS: A case of cutaneous side effect of methotrexate mimicking
Behçets disease. Ann Dermatol. 23:412–414. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Colomer Gallardo A, Martínez Rodríguez R,
Castillo Pacheco C, González Satue C and Ibarz Servio L:
Dermatological side effects of intravesical mitomycin C: Delayed
hypersensitivity. Arch Esp Urol. 69:89–91. 2016.PubMed/NCBI
|
|
24
|
Elazzazy S, Mohamed AE and Gulied A:
Cyclophosphamide-induced symptomatic hyponatremia, a rare but
severe side effect: A case report. Onco Targets Ther. 7:1641–1645.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun W, Yan C, Jia S and Hu J: Correlation
analysis of peripheral DPYD gene polymorphism with 5-fluorouracil
susceptibility and side effects in colon cancer patients. Int J
Clin Exp Med. 7:5857–5861. 2014.PubMed/NCBI
|
|
26
|
Rietjens IM, Boersma MG, Haan L,
Spenkelink B, Awad HM, Cnubben NH, van Zanden JJ, Woude H, Alink GM
and Koeman JH: The pro-oxidant chemistry of the natural
antioxidants vitamin C, vitamin E, carotenoids and flavonoids.
Environ Toxicol Pharmacol. 11:321–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mason SA, Gatta PA Della, Snow RJ, Russell
AP and Wadley GD: Ascorbic acid supplementation improves skeletal
muscle oxidative stress and insulin sensitivity in people with type
2 diabetes: Findings of a randomized controlled study. Free Radic
Biol Med. 93:227–238. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee WJ: The prospects of vitamin C in
cancer therapy. Immune Netw. 9:147–152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nagappan A, Park KI, Park HS, Kim JA, Hong
GE, Kang SR, Lee DH, Kim EH, Lee WS, Won CK, et al: Vitamin C
induces apoptosis in AGS cells by down-regulation of 14-3-3σ via a
mitochondrial dependent pathway. Food Chem. 135:1920–1928. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guerriero E, Sorice A, Capone F,
Napolitano V, Colonna G, Storti G, Castello G and Costantini S:
Vitamin C effect on mitoxantrone-induced cytotoxicity in human
breast cancer cell lines. PLoS One. 9:e1152872014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vetvicka V and Vetvickova J: Combination
of glucan, resveratrol and vitamin C demonstrates strong anti-tumor
potential. Anticancer Res. 32:81–87. 2012.PubMed/NCBI
|
|
32
|
Nagao T, Warnakulasuriya S, Nakamura T,
Kato S, Yamamoto K, Fukano H, Suzuki K, Shimozato K and Hashimoto
S: Treatment of oral leukoplakia with a low-dose of beta-carotene
and vitamin C supplements: A randomized controlled trial. Int J
Cancer. 136:1708–1717. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tokarski S, Rutkowski M, Godala M, Mejer A
and Kowalski J: The impact of ascorbic acid on the concentrations
of antioxidative vitamins in the plasma of patients with non-small
cell lung cancer undergoing first-line chemotherapy. Pol Merkur
Lekarski. 35:136–140. 2013.(In Polish). PubMed/NCBI
|
|
34
|
Nechuta S, Lu W, Chen Z, Zheng Y, Gu K,
Cai H, Zheng W and Shu XO: Vitamin supplement use during breast
cancer treatment and survival: A prospective cohort study. Cancer
Epidemiol Biomarkers Prev. 20:262–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Subramani T, Yeap SK, Ho WY, Ho CL, Omar
AR, Aziz SA, Rahman NM and Alitheen NB: Vitamin C suppresses cell
death in MCF-7 human breast cancer cells induced by tamoxifen. J
Cell Mol Med. 18:305–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hong SW, Jin DH, Hahm ES, Yim SH, Lim JS,
Kim KI, Yang Y, Lee SS, Kang JS, Lee WJ, et al: Ascorbate (vitamin
C) induces cell death through the apoptosis-inducing factor in
human breast cancer cells. Oncol Rep. 18:811–815. 2007.PubMed/NCBI
|
|
37
|
Uetaki M, Tabata S, Nakasuka F, Soga T and
Tomita M: Metabolomic alterations in human cancer cells by vitamin
C-induced oxidative stress. Sci Rep. 5:138962015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van der Reest J and Gottlieb E:
Anti-cancer effects of vitamin C revisited. Cell Res. 26:269–270.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mense SM, Singh B, Remotti F, Liu X and
Bhat HK: Vitamin C and alpha-naphthoflavone prevent
estrogen-induced mammary tumors and decrease oxidative stress in
female ACI rats. Carcinogenesis. 30:1202–1208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yiang GT, Yu YL, Lin KT, Chen JN, Chang WJ
and Wei CW: Acetaminophen induces JNK/p38 signaling and activates
the caspase-9-3-dependent cell death pathway in human mesenchymal
stem cells. Int J Mol Med. 36:485–492. 2015.PubMed/NCBI
|
|
41
|
Yu YL, Yiang GT, Chou PL, Tseng HH, Wu TK,
Hung YT, Lin PS, Lin SY, Liu HC, Chang WJ, et al: Dual role of
acetaminophen in promoting hepatoma cell apoptosis and kidney
fibroblast proliferation. Mol Med Rep. 9:2077–2084. 2014.PubMed/NCBI
|
|
42
|
Lin BR, Yu CJ, Chen WC, Lee HS, Chang HM,
Lee YC, Chien CT and Chen CF: Green tea extract supplement reduces
D-galactosamine-induced acute liver injury by inhibition of
apoptotic and proinflammatory signaling. J Biomed Sci. 16:352009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kolli VK, Abraham P, Isaac B and
Selvakumar D: Neutrophil infiltration and oxidative stress may play
a critical role in methotrexate-induced renal damage. Chemotherapy.
55:83–90. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ohgidani M, Komizu Y, Goto K and Ueoka R:
Residual powders from Shochu distillation remnants induce apoptosis
in human hepatoma cells via the caspase-independent pathway. J
Biosci Bioeng. 114:104–109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu VW and Ho WS: Tetrandrine inhibits
hepatocellular carcinoma cell growth through the caspase pathway
and G2/M phase. Oncol Rep. 29:2205–2210. 2013.PubMed/NCBI
|
|
46
|
Wu Y, van der Schaft DW, Baaijens FP and
Oomens CW: Cell death induced by mechanical compression on
engineered muscle results from a gradual physiological mechanism. J
Biomech. 49:1071–1077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen J, Li Y, Hao H, Li C, Du Y, Hu Y, Li
J, Liang Z, Li C, Liu J, et al: Mesenchymal stem cell conditioned
medium promotes proliferation and migration of alveolar epithelial
cells under Septic conditions in vitro via the JNK-P38 signaling
pathway. Cell Physiol Biochem. 37:1830–1846. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee K, Chung YH, Ahn H, Kim H, Rho J and
Jeong D: Selective regulation of MAPK signaling mediates
RANKL-dependent osteoclast differentiation. Int J Biol Sci.
12:235–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tan CS, Gilligan D and Pacey S: Treatment
approaches for EGFR-inhibitor-resistant patients with
non-small-cell lung cancer. Lancet Oncol. 16:e447–e459. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sato F, Kubota Y, Natsuizaka M, Maehara O,
Hatanaka Y, Marukawa K, Terashita K, Suda G, Ohnishi S, Shimizu Y,
et al: EGFR inhibitors prevent induction of cancer stem-like cells
in esophageal squamous cell carcinoma by suppressing
epithelial-mesenchymal transition. Cancer Biol Ther. 16:933–940.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li X, Qin N, Wang J, Yang X, Zhang X, Lv
J, Wu Y, Zhang H, Nong J and Zhang Q: Clinical observation of
icotinib hydrochloride for advanced non-small cell lung cancer
patients with EGFR status identified. Zhongguo Fei Ai Za Zhi.
18:734–739. 2015.(In Chinese). PubMed/NCBI
|
|
52
|
Osarogiagbon RU, Cappuzzo F, Ciuleanu T,
Leon L and Klughammer B: Erlotinib therapy after initial platinum
doublet therapy in patients with EGFR wild type non-small cell lung
cancer: Results of a combined patient-level analysis of the NCIC
CTG BR.21 and SATURN trials. Transl Lung Cancer Res. 4:465–474.
2015.PubMed/NCBI
|
|
53
|
Montagna E, Cancello G, Bagnardi V,
Pastrello D, Dellapasqua S, Perri G, Viale G, Veronesi P, Luini A,
Intra M, et al: Metronomic chemotherapy combined with bevacizumab
and erlotinib in patients with metastatic HER2-negative breast
cancer: Clinical and biological activity. Clin Breast Cancer.
12:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tolcher AW, LoRusso P, Arzt J, Busman TA,
Lian G, Rudersdorf NS, Vanderwal CA, Kirschbrown W, Holen KD and
Rosen LS: Safety, efficacy, and pharmacokinetics of navitoclax
(ABT-263) in combination with erlotinib in patients with advanced
solid tumors. Cancer Chemother Pharmacol. 76:1025–1032. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Evans MK, Tovmasyan A, Batinic-Haberle I
and Devi GR: Mn porphyrin in combination with ascorbate acts as a
pro-oxidant and mediates caspase-independent cancer cell death.
Free Radic Biol Med. 68:302–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ohtani S, Iwamaru A, Deng W, Ueda K, Wu G,
Jayachandran G, Kondo S, Atkinson EN, Minna JD, Roth JA, et al:
Tumor suppressor 101F6 and ascorbate synergistically and
selectively inhibit non-small cell lung cancer growth by
caspase-independent apoptosis and autophagy. Cancer Res.
67:6293–6303. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin SY, Lai WW, Chou CC, Kuo HM, Li TM,
Chung JG and Yang JH: Sodium ascorbate inhibits growth via the
induction of cell cycle arrest and apoptosis in human malignant
melanoma A375.S2 cells. Melanoma Res. 16:509–519. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nagao N, Nakayama T, Etoh T, Saiki I and
Miwa N: Tumor invasion is inhibited by phosphorylated ascorbate via
enrichment of intracellular vitamin C and decreasing of oxidative
stress. J Cancer Res Clin Oncol. 126:511–518. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Liu JW, Nagao N, Kageyama K and Miwa N:
Antimetastatic and anti-invasive ability of phospho-ascorbyl
palmitate through intracellular ascorbate enrichment and the
resultant antioxidant action. Oncol Res. 11:479–487.
1999.PubMed/NCBI
|
|
60
|
Pires AS, Marques CR, Encarnação JC,
Abrantes AM, Mamede AC, Laranjo M, Gonçalves AC, Sarmento-Ribeiro
AB and Botelho MF: Ascorbic acid and colon cancer: An oxidative
stimulus to cell death depending on cell profile. Eur J Cell Biol.
95:208–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Beck R, Pedrosa RC, Dejeans N, Glorieux C,
Levêque P, Gallez B, Taper H, Eeckhoudt S, Knoops L, Calderon PB,
et al: Ascorbate/menadione-induced oxidative stress kills cancer
cells that express normal or mutated forms of the oncogenic protein
Bcr-Abl. An in vitro and in vivo mechanistic study. Invest New
Drugs. 29:891–900. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Venturelli S, Sinnberg TW, Berger A, Noor
S, Levesque MP, Böcker A, Niessner H, Lauer UM, Bitzer M, Garbe C,
et al: Epigenetic impacts of ascorbate on human metastatic melanoma
cells. Front Oncol. 4:2272014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ma Y, Chapman J, Levine M, Polireddy K,
Drisko J and Chen Q: High-dose parenteral ascorbate enhanced
chemosensitivity of ovarian cancer and reduced toxicity of
chemotherapy. Sci Transl Med. 6:222ra182014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cieslak JA and Cullen JJ: Treatment of
pancreatic cancer with pharmacological ascorbate. Curr Pharm
Biotechnol. 16:759–770. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Presutti D, Santini S, Cardinali B, Papoff
G, Lalli C, Samperna S, Fustaino V, Giannini G and Ruberti G: MET
gene amplification and MET receptor activation are not sufficient
to predict efficacy of combined MET and EGFR inhibitors in EGFR
TKI-resistant NSCLC cells. PLoS One. 10:e01433332015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
De S, Dermawan JK and Stark GR: EGF
receptor uses SOS1 to drive constitutive activation of NF-κB in
cancer cells. Proc Natl Acad Sci USA. 111:11721–11726. 2014.
View Article : Google Scholar : PubMed/NCBI
|