Introduction
Lung cancer is one of the most malignant cancer
types all over the world (1), with
a low 5-year survival rate of 16.6% (2). Non-small cell lung cancer (NSCLC) is
the predominant form of lung cancer and accounts for the majority
of cancer-related deaths in the world (3). Conventional therapeutic strategies of
chemotherapy following surgery revealed limited effect for advanced
NSCLC patients (4). A better
understanding of the molecular mechanisms underlying NSCLC
resistance and developing personalized therapeutic strategies is
urgently needed to improve NSCLC prognosis.
Recently, an improved understanding of NSCLC
pathogenesis has led to the development of multiple kinase
inhibitors, such as gefitinib, one of known tyrosine kinase
inhibitors (TKIs). Gefitinib is an orally active, selective and
reversible TKI, which blocks ATP from binding to the EGFR-TK
activation (5). EGFR kinase domain
mutations, including T790M in exon 20 and L858R in exon 21 of EGFR,
represent the first molecular targeting markers for TKI treatment
such as gefitinib. Treatment of NSCLC with gefitinib has been found
in many clinical studies, and the output is complex (6,7).
Moreover, gefitinib may be effective at initial treatments, however
resistance may increase substantially after a period of exposure,
ending up with cancer progression after 6–15 months of therapy
(8). Thus, revealing the mechanism
of gefitinib resistance and discovering reliable biological targets
that play important roles in gefitinib resistance warranted
investigation.
Long non-coding RNAs (lncRNAs) are a major group of
ncRNAs that contain more than 200 nucleotides (9). During recent years, thousands of
studies have revealed that lncRNAs may serve as critical biological
regulators in the functions of cellular and molecular signaling
pathways. lncRNA H19 is located on chromosome 11 in humans and is a
maternally-expressed imprinted gene that plays a vital role in
mammalian development (10). Recent
studies revealed that H19 is overexpressed in several malignancies
and may serve as an oncogene via promotion of cell proliferation
and chemoresistance (11–13). However, most of the studies have
only revealed the function of H19 within cells (nuclear or
cytoplasm). The existing pattern of extracellular H19 is not well
known.
Exosomes are membrane-derived vesicles and have a
size range of 20–200 nm when released into body fluids such as
blood, urine, and malignant ascites. These vesicles contain DNAs,
protein fragment, coding or non-coding RNAs which are secreted from
their parental cell cytoplasm and can be enrolled into recipient
cells (14). Recently, some studies
have indicated that the exosomes from chemo-sensitive/resistant
cells could markedly influence chemo-response of receipt cells
through the transfer of specific genes, such as lncRNAs (15). However, this conclusion warrants
more persuasive support.
In this study, we hypothesized that extracellular
H19 promoted gefitinib resistance through incorporation into
exosomes. To validate this hypothesis, we built gefitinib-resistant
NSCLC cell lines and identified the expression of H19 in both
gefitinib-resistant cells and parental sensitive cells. By
performing a series of in vitro assays, we investigated the
functional relevance of exosomal H19 in gefitinib resistance of
NSCLC cells.
Materials and methods
Cell culture
The human NSCLC cell lines HCC827 and HCC4006, which
harbor EGFR activating mutations (16,17),
were purchased from the Chinese Academy of Sciences (Shanghai,
China). Both cell lines were cultured in RPMI-1640 medium
(BioWhittaker®; Lonza Group, Ltd., Basel, Switzerland)
supplemented with 10 mM HEPES, 1 mM L-glutamine, 100 U/ml
penicillin/streptomycin (BioWhittaker®; Lonza Group) and
heat inactivated 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and grown at 37°C in a 5% CO2
atmosphere. Gefitinib (Iressa; AstraZeneca, Macclesfield, UK) was
dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at a concentration of 10 mM and stored at
−20°C. Gefitinib-resistant HCC827R and HCC4006R cells were
established by initially culturing with 1 µM gefitinib in DMEM plus
10% FBS for 6 weeks. Subsequently, a 2-µM concentration of
gefitinib was used to treat the surviving cells for 8 weeks and 5
µM for another 8 weeks. Eventually, the gefitinib-resistant NSCLC
cell lines were successfully established by culturing the cells in
10 µM gefitinib.
Exosome isolation, labeling and RNA
extraction
Exosomes were extracted from culture medium using
ExoQuick precipitation kit (System Biosciences, Mountain View, CA,
USA) according to manufacturer's instructions. Briefly, the culture
medium was thawed on ice and centrifuged at 3,000 × g for 15 min to
remove cells and cell debris. Next, 250 µl of the supernatant was
mixed with 63 µl of ExoQuick precipitation kit and then incubated
for 40 min at 5°C after brief shaking and mixing, followed by
centrifugation at 1,500 × g for 30 min. Then, the supernatant was
removed by careful aspiration, followed by another 5 min of
centrifugation to remove the residual liquid. The
exosome-containing pellet was subsequently re-suspended in 250 µl
phosphate-buffered saline (PBS). The final pellets, containing
exosomes, were collected for characterization and RNA isolations.
Size distribution of exosomes was analyzed by Zetasizer (Malvern
Panalytical Ltd., Malvern, UK). Purified exosomes were labeled with
PKH26 Red Fluorescent Cell Linker Kit for General Cell Membrane
Labeling (Sigma-Aldrich; Merck KGaA) as per the manufacturer's
protocol.
RNA extraction
Extraction of RNA from exosomes was performed using
the commercial miRNeasy Serum/Plasma kit (Qiagen Sciences Inc.,
Gaithersburg, MD, USA), and RNA extraction from cell fraction was
performed using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. RNA
elution steps were carried out at 12,000 × g for 15 sec, and the
extracted RNA was dissolved in RNase-free ultra-pure water.
Transmission electron microscopy
(TEM)
We used 50 µl PBS to suspend the exosomes pellets
and then put one drop of this suspension on the parafilm. A copper
mesh coated with carbon was then used to drift on the drop for 5
min at 25°C. Then, the grid was removed, and the excess liquid was
drained by touching the grid edge against a piece of clean filter
paper. The grid was then placed onto a drop of 2% phosphotungstic
acid with pH 7.0 for approximately 5 sec, and the excess liquid was
drained off. The grid was allowed to dry for several minutes and
then examined using a JEM-1200 EX microscope (JEOL Ltd., Akishima,
Japan) at 80-kiloelectron volts.
Reverse transcription-quantitative PCR
(RT-qPCR)
The cDNA was synthesized from 200 ng extracted total
RNA using the PrimeScript RT reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China) and amplified by RT-qPCR with a SYBR Green Kit
(Takara Biotechnology Co.) on an ABI PRISM 7500 Sequence Detection
System (Life Technologies; Thermo Fisher Scientific, Inc.) with the
housekeeping gene GAPDH as an internal control by using the ∆∆Cq
method (18). The primer sequences
are presented in Table I.
 | Table I.Information of the RT-qPCR primer
sequences and siRNA sequences. |
Table I.
Information of the RT-qPCR primer
sequences and siRNA sequences.
| RT-qPCR primer
name | Primer sequence
(5′-3′) |
|---|
| H19 (forward) |
ATCGGTGCCTCAGCGTTCGG |
| H19 (reverse) |
CTGTCCTCGCCGTCACACCG |
| GAPDH
(forward) |
GCACCGTCAAGGCTGAGAAC |
| GAPDH
(reverse) |
ATGGTGGTGAAGACGCCAGT |
|
| siRNA
name | siRNA sequence
(5′-3′) |
|
| si-H19 #1 |
CCACTCCACCTCAAACTCTTACCTT |
| si-H19 #2 |
GGGTCATTAAGGGACAGAGTTCAAG |
| si-H19 #3 |
CAGGTGGACTCACAATTCCAAATAT |
| si-hnRNPA2B1 |
AATTGATGGGAGAGTAGTTGA |
| si-NC | Cat. no. 12935-110
(Invitrogen; Thermo Fisher Scientific, Inc.) |
Cell transfection
The small interfering RNA against H19 (si-H19) and
hnRNPA2B1 (si-hnRNPA2B1) were synthesized and prepared by Shanghai
GenePharma Co., Ltd. (Shanghai, China). Negative control siRNA was
purchased from Invitrogen; Thermo Fisher Scientific, Inc.
(Shanghai, China; cat. no. 12935-110). All the vectors were labeled
with green fluorescence protein (GFP). Briefly, a total of
1.2×107 cells were plated in 15-cm culture dishes for 24
h and then transfected with the vectors described above using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) for
24 h. The cells were then subjected to RNA/protein extraction and
further functional assays. The sequences of small interfering RNAs
are presented in Table I.
TUNEL assay
Gefitinib-induced nuclear apoptosis was evaluated by
performing TUNEL analysis. In brief, cells were treated with
si-H19#3 or negative control for 24 h and fixed by using 4%
formaldehyde. Cells were fixed and stained with TUNEL kit according
to the manufacturer's instructions (TUNEL Bright-Red Apoptosis
Detection kit; cat. no. A113; Vazyme Biotech Co., Ltd., Nanjing,
China). TUNEL-positive cells were counted under fluorescence
microscopy (DMI4000B; Leica Microsystems GmbH, Wetzlar,
Germany).
RNA immunoprecipitation (RIP)
NSCLC cells were rinsed with cold PBS and fixed by
1% formaldehyde for 10 min. After centrifugation at 1,500 × g for 5
min at 4°C, cell pellets were collected and re-suspended in the
NP-40 lysis buffer. For RIP assay, the supernatant was incubated
overnight with beads conjugated with anti-hnRNPA2B1 antibody (1:50;
cat. no. ab31645; Abcam, Cambridge UK) or negative control mouse
IgG (cat. no. 12-371; EMD Millipore, Burlington, MA, USA). The
beads were then rinsed with cold NT2 buffer and cultured with
proteinase K at 10 mg/ml (Sigma-Aldrich; Merck KGaA).
Cell viability assay
Alterations in cell viability following transfection
or gefitinib treatment was assayed using CCK-8 kit (Dojindo
Molecular Technologies, Inc., Rockville, MD, USA). In brief, cells
were seeded into a 96-well plate in triplicate and then treated
with si-H19 or (and) gefitinib for different periods of time. The
cell cultures were then treated with CCK-8 reagent and further
cultured for 2 h. The optical density at 450 nm was measured using
a spectrophotometer (Thermo Fisher Scientific, Inc.). The
percentage of the control samples of each cell line was calculated
thereafter.
Nanoparticle tracking analysis
(NTA)
Briefly, ~0.3 ml supernatant was loaded into the
sample chamber of an LM10 Nano-sight unit (NanoSight Ltd.,
Amesbury, UK) and three videos of either 30 or 60 sec were recorded
for each sample. Data analysis was performed with both NTA 2.1
software (NanoSight). The diffusion coefficient and
sphere-equivalent hydrodynamic radius were then determined using
the Stokes-Einstein equation, and results were displayed as a
particle size distribution.
Western blotting and antibodies
Cell lysates were prepared with RIPA buffer
containing protease inhibitors (Sigma-Aldrich; Merck KGaA). Protein
concentrations were assessed with the BCA Protein Assay kit
according to the manufacturer's instructions (Beyotime Institute of
Biotechnology, Shanghai, China). Equal amounts of protein (25 µg)
were separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene fluoride
membranes (EMD Millipore). Then, the membrane was blocked with 5%
(5 g/100 ml) non-fat dry milk in Tri-buffered saline plus Tween
(TBS-T) buffer for 2 h at room temperature. The membranes were
incubated overnight at 4°C with a 1:1,000 solution of antibodies:
Anti-TSG101 (cat. no. ab125011), anti-hnRNPA2B1 (cat. no. ab31645),
anti-CD63 (cat. no. ab134045) and anti-β-actin (cat. no. ab8226;
all from Abcam). The horseradish peroxidase-conjugated (HRP)
anti-rabbit antibody (1:5,000; cat. no. 7074; Cell Signaling
Technology, Inc., Danvers, MA, USA) was used as a secondary
antibody for immunostaining for 1 h at room temperature. The
proteins were visualized using a detection system of enhanced
chemiluminescence (ECL) by using Immobilon Western Chemiluminescent
HRP Substrate (EMD Millipore).
Statistical analysis
Mann-Whitney U test was used for the comparison of
datasets containing two groups. The Kruskal-Wallis test followed by
post-hoc test with Bonferroni's was used for evaluating the
difference among multiple groups. The survival curves of NSCLC
cells were estimated via the Kaplan-Meier method, and the
difference in survival rate was analyzed using the log-rank
testing. Statistical analysis was performed using Prism 4 (GraphPad
Software Inc., San Diego, CA, USA) and a P-value threshold of
<0.05 was considered to indicate a statistically significant
difference.
Results
H19 expression is increased in
gefitinib-resistant NSCLC cells
To investigate the underlying regulatory mechanism
of gefitinib resistance, two gefitinib-resistant sub-lines derived
from HCC827 and HCC4006 cell lines were constructed (HCC827R and
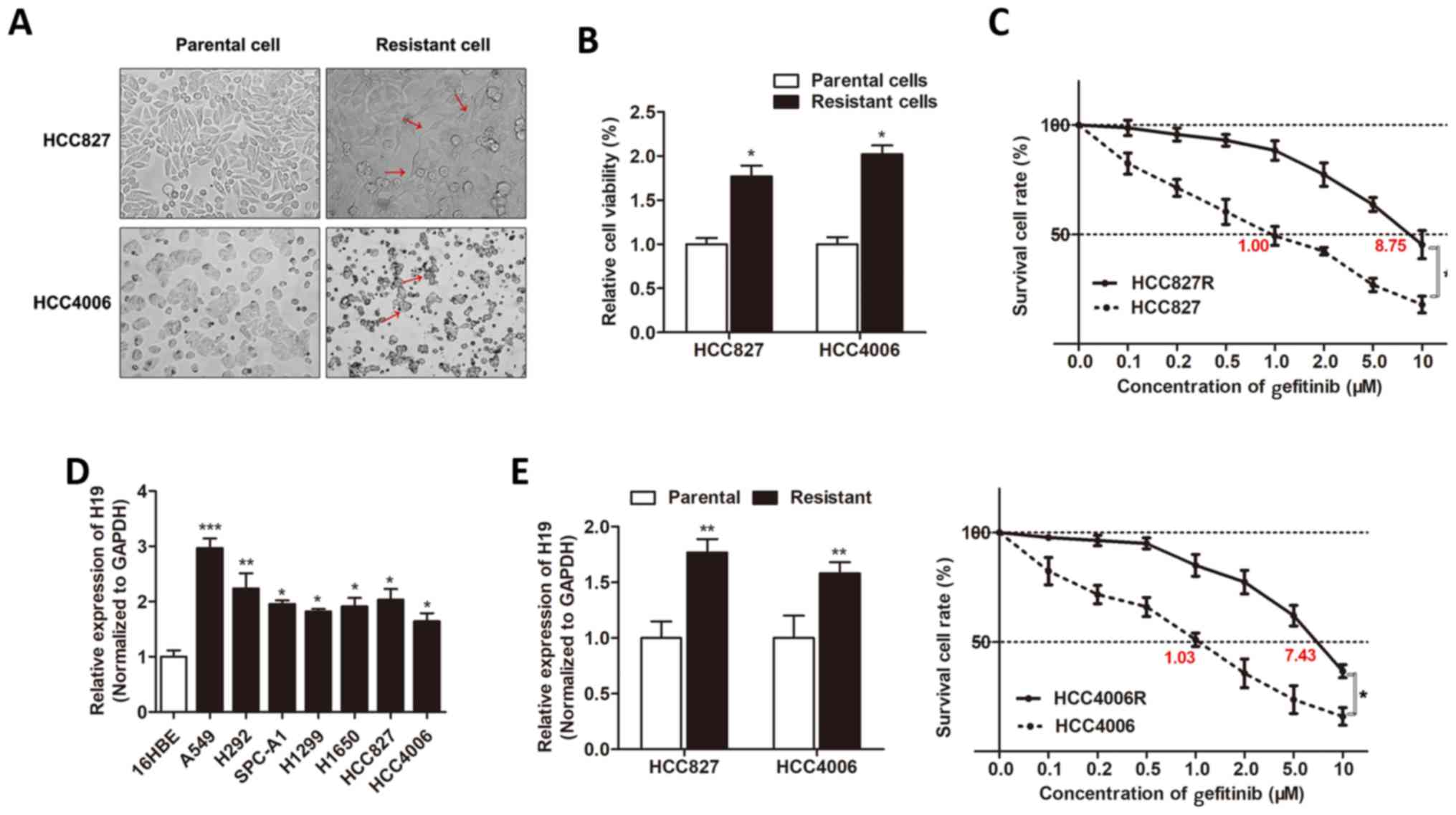
HCC4006R, respectively). We found that the built
gefitinib-resistant cells exhibited characteristic changes,
including loss of cell polarity, increased intercellular
separation, and increased formation of pseudopodia (Fig. 1A). CCK-8 assay revealed that the
cell viability of HCC827R and HCC4006R cells was significantly
increased when compared to respective parental cells under the
treatment of gefitinib for 48 h (Fig.
1B). A dose-effect curve was built, and we identified that the
IC50 value of gefitinib for HCC827R cells was 8.75 µM,
whereas HCC827 was 1.00 µM, meaning that the ability of resistance
to gefitinib was 8.75 times higher for the HCC827R cells than that
of the HCC827 cells. Similarly, the HCC4006R cell exhibited a 7.21
times higher resistance to gefitinib than that of the HCC4006 cells
(7.43/1.03; Fig. 1C). Then, we
determined the expression level of H19 in NSCLC cell lines by using
RT-qPCR, and we identified that H19 was upregulated in most NSCLC
cells in contrast to normal epithelial cells, 16HBE (Fig. 1D). In addition, H19 was upregulated
in gefitinib-resistant cell lines when compared to the respective
parental cells (Fig. 1E). This
indicated that H19 may be important for gefitinib resistance of
NSCLC cells.
Knockdown of H19 resensitized
gefitinib resistance in NSCLC cells
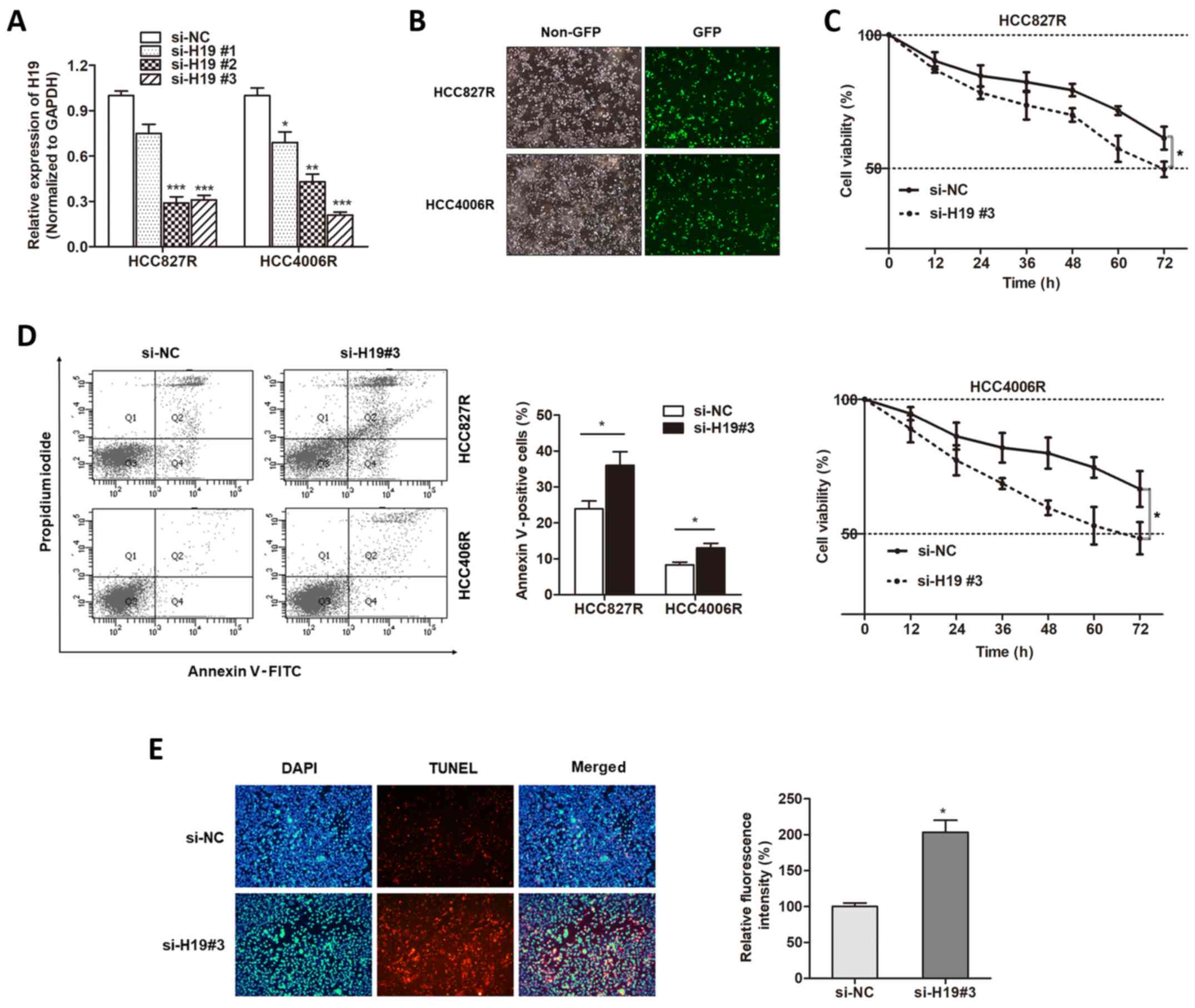
With the verification of aberrant expression of H19
in gefitinib-resistant cells, we sought to determine whether H19
was involved in gefitinib resistance. Three small interfering RNAs
against H19 were generated, and we found that H19 expression was
mostly silenced in the HCC827R and HCC4006R cells when incubated
with si-H19#3 (Fig. 2A), which was
used for the following experiments. We transfected
gefitinib-resistant cells with si-H19#3 (Fig. 2B). Compared with the response of the
control group, silencing of H19 promoted gefitinib-induced cell
cytotoxicity (Fig. 2C). FACS
apoptosis assay revealed that gefitinib exposure caused an
increased proportion of apoptotic cells in H19-knockdown cells in
contrast to si-NC-transfected cells (Fig. 2D). Then, we used TUNEL assay to
detect whether H19 influenced the nuclear apoptosis induced by
gefitinib. We found that knockdown of H19 increased the
gefitinib-induced nuclear apoptosis of HCC827R cells (Fig. 2E). Therefore, we demonstrated that
H19 was essential for gefitinib resistance in NSCLC.
H19 is secreted through incorporation
into exosomes
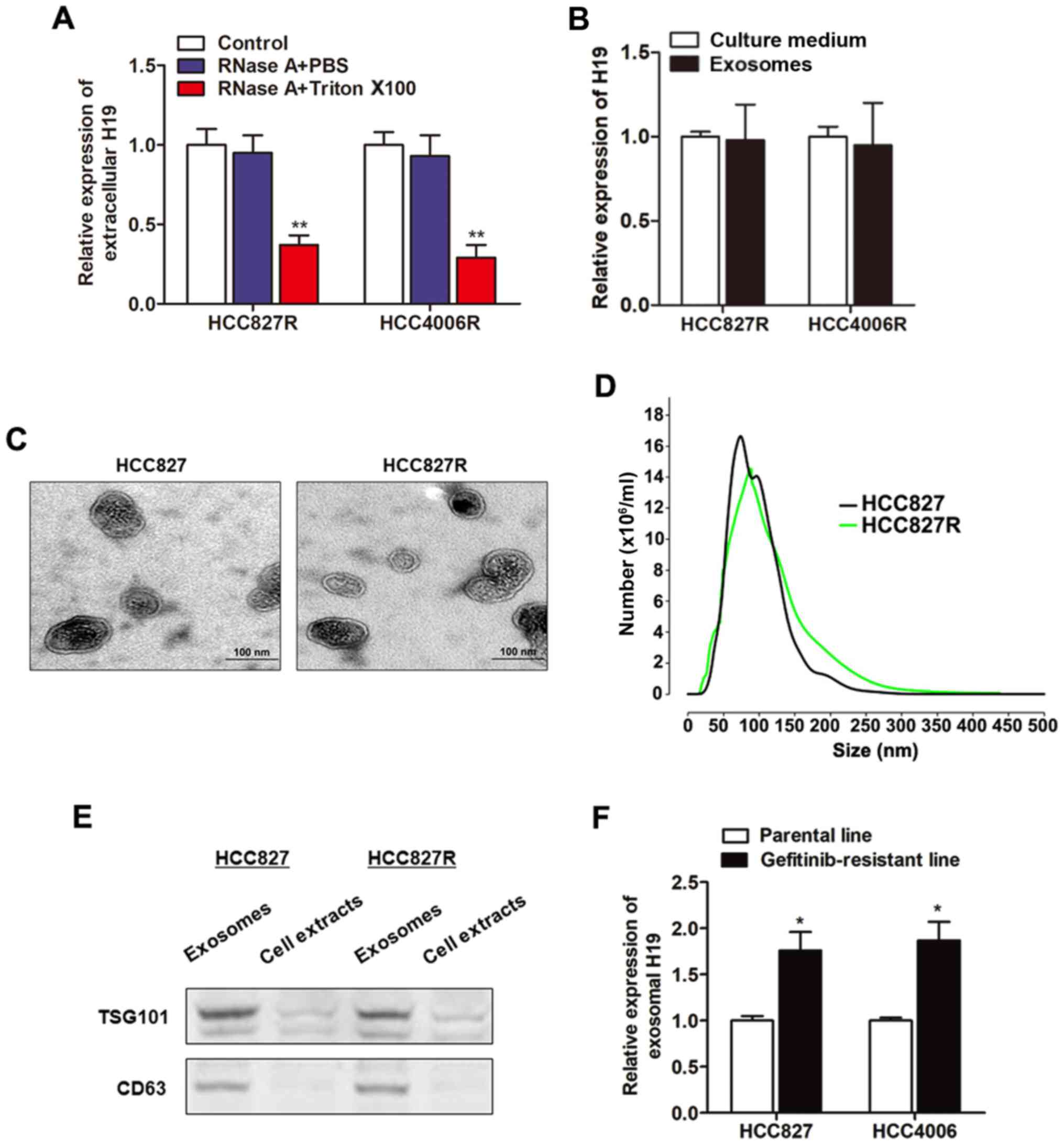
To investigate how H19 regulates gefitinib
resistance, we firstly localized the expression of H19 in NSCLC
cells. By using the online lncRNA location prediction software
lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/), we
identified that intracellular H19 was located in exosomes (Table II). To verify whether H19 is
secreted through packaging into exosomes, we detected the
expression level change of extracellular H19 after treatment with
RNase. As revealed in Fig. 3A, H19
in culture medium was little influenced by the treatment of RNase
alone but significantly decreased when treated with RNase and
Triton X-100 simultaneously, suggesting that extracellular H19 was
protected by the membrane instead of being directly secreted. H19
expression levels in exosomes were almost equal to that in whole
culture medium, indicating that extracellular H19 was contained in
exosomes (Fig. 3B). We then
purified and extracted exosomes from culture medium, and the
representative micrograph captured by Transmission Electron
Microscopy (TEM) was revealed in Fig.
3C. A similar morphology, size, and number were identified
between HCC827R and HCC827 cells by NTA analysis (Fig. 3D). Western blot assays further
confirmed their identity by enriched exosome proteins, such as
TSG101 and CD63 (Fig. 3E). Then, we
determined whether H19 was incorporated into exosomes by isolation
of exosomes with the ExoQuick purification kit followed by qPCR. As
anticipated, H19 was detectable in exosomes, and the expression
level was significantly higher in gefitinib-resistant cells than in
the respective parental cells (Fig.
3F), indicating that extracellular H19 was secreted through
incorporation into exosomes in NSCLC cells.
| Table II.Presentation of score of H19 at
different subcellular locations by lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/). |
Table II.
Presentation of score of H19 at
different subcellular locations by lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/).
| Subcellular
locations | Score |
|---|
| Cytoplasm |
0.0912586130123 |
| Nucleus |
0.0608739221682 |
| Ribosome |
0.0128328125515 |
| Cytosol | 0.511872590452 |
| Exosome | 0.323162061816 |
hnRNPA2B1 mediates the packaging of
H19 into exosomes
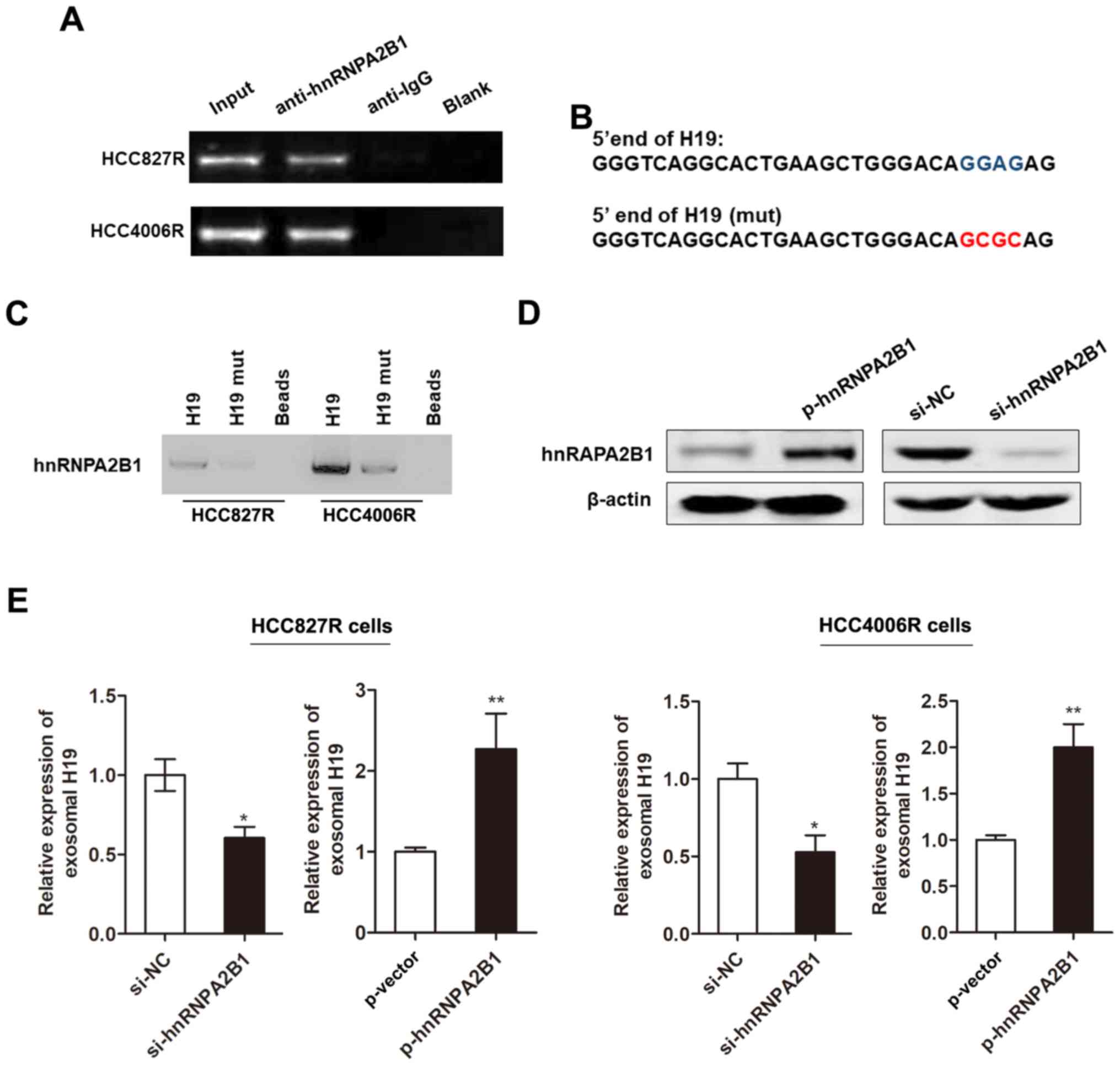
Subsequently, we investigated whether the packaging
of H19 into exosomes was mediated by specific regulator. Previous
literature reported that heterogeneous nuclear ribonucleoprotein
A2B1 (hnRNPA2B1), an RNA-binding protein, could control RNA loading
into exosomes by binding to the specific motif (GGAG) (19), which is found at the 5′ end region
of H19. Herein, we used RIP assay to verify the association between
H19 and hnRNPA2B1. As anticipated, a substantial enrichment was
identified between H19 and hnRNPA2B1 (Fig. 4A). Moreover, RNA pull-down analysis
highlighted that hnRNPA2B1-binding ability was impaired when the
‘GGAG’ sequence is mutated (Fig. 4B and
C). We then generated hnRNPA2B1 silencing or overexpressing
vector (Fig. 4D). We found that the
enhanced expression of hnRNPA2B1 promoted the level of exosomal H19
whereas knockdown of hnRNPA2B1 decreased its expression in HCC827R
cells (Fig. 4E). Our results
indicated that hnRNPA2B1 mediated the packaging of H19 into
exosomes.
Exosome-mediated transfer of H19
disseminates gefitinib resistance
To examine whether H19 regulates gefitinib
resistance through the delivery of exosomes, we demonstrated the
H19-contained exosomes can be taken up by recipient cells by two
steps. First, we examined whether secreted exosomes can be taken up
by recipient cells by labeling isolated exosomes with PKH26 dye
from HCC827R cells. The labeled exosomes were then added and
incubated with HCC827 and HCC4006 cells for 24 h. As revealed in
Fig. 5A, the recipient cells
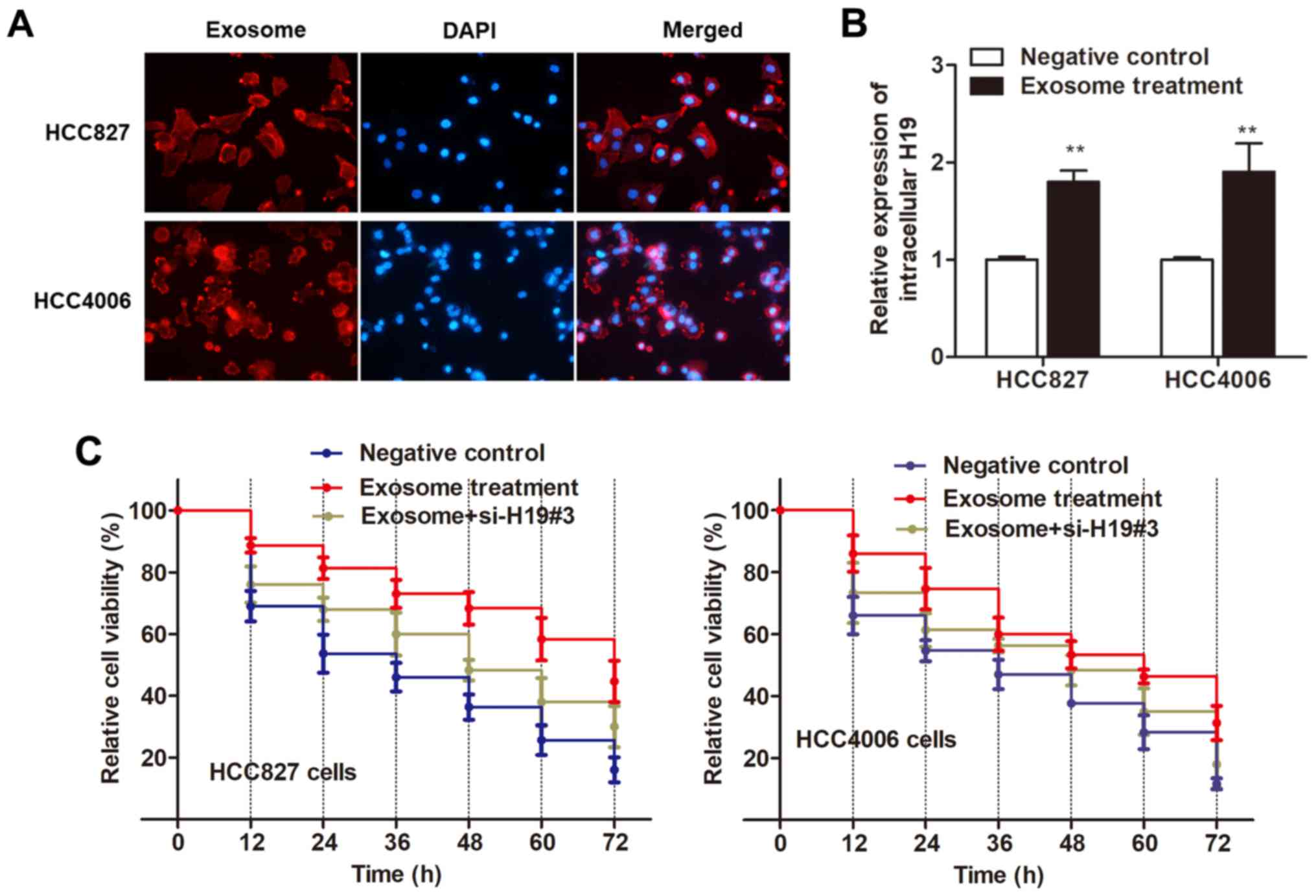
exhibited a red signal under a confocal microscope. Second, we
examined whether these exosomes could deliver H19 to recipient
cells similar to the intercellular transfer of other non-coding RNA
as previously reported (20,21).
As anticipated, RT-qPCR revealed an increased expression of H19 in
both recipient cells incubated with exosomes (Fig. 5B). Thus, we ascertained that
H19-contained exosomes can be taken up by recipient cells.
Next, we determined whether HCC827 and HCC4006 cells
with elevated H19 levels displayed an increased resistance to
gefitinib. As revealed in Fig. 5C,
both recipient cell lines exhibited a promoted cell viability after
treatment of exosomes as compared with the control cells. However,
this effect was reversed by the transfection of si-H19#3 (Fig. 5C), indicating that it is exosomal
H19 that induced gefitinib resistance.
Discussion
Extensive efforts in the past have contributed to
the understanding of both molecular and cellular mechanisms of
action of chemoresistance, one of the major causes for the failure
of treatment with advanced cancer types. However, little progress
has been made ever since (22).
Thus, novel molecular signatures appear to hold great promise in
tumor characterization and could be used as potential prognostic
markers and treatment targets. To identify potential molecular
therapeutic markers for gefitinib treatment, we built
gefitinib-resistant cell lines, and investigated the functional
association between gefitinib resistance and lncRNA H19. Our
original data revealed that H19 expression was increased in
gefitinib-resistant cells, and extracellular H19 promoted gefitinib
resistance of NSCLC cells by packaging into exosomes.
EGFR is critical in proliferation and survival
pathways, and activating mutations are often observed in NSCLC
(23). EGFR mutations occur more
frequently in Asian patients compared with Caucasian patients
(24). Gefitinib (Iressa;
AstraZeneca) is an orally administered, small-molecule EGFR-TK
inhibitor that blocks signal transduction pathways implicated in
proliferation and survival of cancer cells. Large phase III or IV
clinical trials in patients with locally advanced or metastatic
NSCLC revealed that gefitinib as first- or subsequent-line
treatment significantly prolonged progression-free survival (PFS)
and improved objective response rates (25). However, acquired resistance, defined
as progression after initial benefit, to targeted therapies
inevitably occurs (26). Therefore,
breakthroughs are needed in the understanding and treatment of
acquired gefitinib resistance in NSCLC, especially for patients
with EGFR mutant and ALK rearrangement-positive sites.
The roles of lncRNAs in cancer progression have long
been researched and H19 was widely accepted as an oncogene in
cancers, including NSCLC (27).
However, its role in gefitinib resistance is not well known. In
this study, we found that H19 was upregulated in
gefitinib-resistant cells and was essential for gefitinib
resistance as evidenced by the result that knockdown of H19
resensitized cells to gefitinib treatment. Next, we investigated
how H19 exerted its oncogenic function, herein we focused on
exosomes. Exosomes are nano-sized vesicles secreted upon the fusion
of vesicular-like properties with plasma membranes in large amount
of cell types (28). They have been
well identified as a way of information exchange between different
type of cells, through the transfer of constituents, such as
lncRNAs (29). As expected, we
demonstrated that H19 participated in gefitinib resistance through
incorporation into exosomes by using two-steps approaches.
Exosomes contain genes and proteins, reflecting the
features of cancer cells, which provides us with the development of
highly sensitive diagnostic strategies for monitoring the
therapeutic response conditions of cancer in a rapid and
non-invasive manner (30). We then
determined whether the ectopic expression of exosomal H19 mediated
gefitinib resistance in NSCLC cells. As expected, treatment with
exosomes extracted from culture medium of resistant cells potently
reduced the gefitinib-induced cell apoptosis, indicating that H19
promoted gefitinib resistance via packaging into exosomes. Our
study was consistent with the study by Conigliaro et al.
They demonstrated that CD90+ liver cancer cells
modulated endothelial cell phenotype through the release of
exosomes containing H19 lncRNA (12). Another study performed by Li et
al revealed that cholangiocyte-derived exosomal H19 promoted
cholestatic liver injury in mouse and humans (31), indicating that H19 in exosomes may
be important for the progression of multiple diseases. Furthermore,
the exosomal secretion of RNAs was reported as highly selective and
different between cancer and normal cells (32), so the identification of cellular
molecules responsible for specific RNA secretion may propose unique
strategies to block cell-specific RNA secretion. In the present
study, we identified the RNA-binding protein, hnRNPA2B1, which is
known to transport RNA to exosomes probably dependent on its
ability to interact with cytoskeletal components (33). We found that hnRNPA2B1 interacted
with H19 and was responsible for the packaging of H19 into
exosomes.
We must admit that our results warrant further
support by the data from clinical trials, and in future our
translational study will be enhanced for the clinical benefit of
NSCLC patients. In addition, gefitinib resistance has been widely
studied and other involved pathways have been indicated, such as
the PI3K/Akt pathway (34,35) and the NF-κB/STAT3 pathway (36). However, whether these pathways were
linked and co-regulated is still not well known. Thus, a
comprehensive understanding of gefitinib resistance is needed to
identify the underlying associations between the pathway we
revealed and other chemo-resistance pathways. Finally, it is still
unknown whether the expression of H19 is responsible for the
resistance of gefitinib in NSCLC by regulating genetic mutation. In
the future, we will extend our study to reveal the underlying
regulatory mechanisms.
In conclusion, the present study demonstrated that
upregulated H19 promoted gefitinib resistance in NSCLC cells
through incorporation into exosomes. Therefore, H19 in exosomes
could be a promising therapeutic target for NSCLC patients.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Science Foundation of China (87394733).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL, WG and WL acquired the data and created a draft
of the manuscript; YL, WG and BC prepared the experimental
materials and performed the in vitro assays; BC, LC and JG
interpreted the data, performed the statistical analysis and
analyzed the results; YL and WL revised and approved the final
version of the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ettinger DS, Akerley W, Bepler G, Blum MG,
Chang A, Cheney RT, Chirieac LR, D'Amico TA, Demmy TL, Ganti AK, et
al: Non-small cell lung cancer. J Natl Compr Canc Netw. 8:740–801.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pastorino U: Lung cancer screening. Br J
Cancer. 102:1681–1686. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gettinger S and Lynch T: A decade of
advances in treatment for advanced non-small cell lung cancer. Clin
Chest Med. 32:839–851. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Javle M, Pande A, Iyer R, Yang G, LeVea C,
Wilding G, Black J, Nava H and Nwogu C: Pilot study of gefitinib,
oxaliplatin, and radiotherapy for esophageal adenocarcinoma: Tissue
effect predicts clinical response. Am J Clin Oncol. 31:329–334.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Masago K, Fujita S, Irisa K, Kim YH,
Ichikawa M, Mio T and Mishima M: Good clinical response to
gefitinib in a non-small cell lung cancer patient harboring a rare
somatic epidermal growth factor gene point mutation; codon 768 AGC
> ATC in exon 20 (S768I). Jpn J Clin Oncol. 40:1105–1109. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ferry DR, Anderson M, Beddard K, Tomlinson
S, Atherfold P, Obszynska J, Harrison R and Jankowski J: A phase II
study of gefitinib monotherapy in advanced esophageal
adenocarcinoma: Evidence of gene expression, cellular, and clinical
response. Clin Cancer Res. 13:5869–5875. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jackman DM, Holmes AJ, Lindeman N, Wen PY,
Kesari S, Borras AM, Bailey C, de Jong F, Jänne PA and Johnson BE:
Response and resistance in a non-small-cell lung cancer patient
with an epidermal growth factor receptor mutation and
leptomeningeal metastases treated with high-dose gefitinib. J Clin
Oncol. 24:4517–4520. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang S, Qin C, Cao G, Xin W, Feng C and
Zhang W: Systematic analysis of long noncoding RNAs in the
senescence-accelerated mouse prone 8 brain using RNA sequencing.
Mol Ther Nucleic Acids. 5:e3432016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Keniry A, Oxley D, Monnier P, Kyba M,
Dandolo L, Smits G and Reik W: The H19 lincRNA is a developmental
reservoir of miR-675 that suppresses growth and Igf1r. Nat Cell
Biol. 14:659–665. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ding D, Li C, Zhao T, Li D, Yang L and
Zhang B: LncRNA H19/miR-29b-3p/PGRN axis promoted
epithelial-mesenchymal transition of colorectal cancer cells by
acting on Wnt signaling. Mol Cells. 41:423–435. 2018.PubMed/NCBI
|
|
12
|
Conigliaro A, Costa V, Lo Dico A, Saieva
L, Buccheri S, Dieli F, Manno M, Raccosta S, Mancone C, Tripodi M,
et al: CD90+ liver cancer cells modulate endothelial cell phenotype
through the release of exosomes containing H19 lncRNA. Mol Cancer.
14:1552015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun H, Wang G, Peng Y, Zeng Y, Zhu QN, Li
TL, Cai JQ, Zhou HH and Zhu YS: H19 lncRNA mediates
17β-estradiol-induced cell proliferation in MCF-7 breast cancer
cells. Oncol Rep. 33:3045–3052. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Théry C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pefanis E, Wang J, Rothschild G, Lim J,
Kazadi D, Sun J, Federation A, Chao J, Elliott O, Liu ZP, et al:
RNA exosome-regulated long non-coding RNA transcription controls
super-enhancer activity. Cell. 161:774–789. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fustaino V, Presutti D, Colombo T,
Cardinali B, Papoff G, Brandi R, Bertolazzi P, Felici G and Ruberti
G: Characterization of epithelial-mesenchymal transition
intermediate/hybrid phenotypes associated to resistance to EGFR
inhibitors in non-small cell lung cancer cell lines. Oncotarget.
8:103340–103363. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Presutti D, Santini S, Cardinali B, Papoff
G, Lalli C, Samperna S, Fustaino V, Giannini G and Ruberti G: MET
gene amplification and met receptor activation are not sufficient
to predict efficacy of combined MET and EGFR inhibitors in EGFR
TKI-resistant NSCLC cells. PLoS One. 10:e01433332015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu SY, Huang X and Cheong KL: Recent
advances in marine algae polysaccharides: Isolation, structure, and
activities. Mar Drugs. 15:pii: E3882017. View Article : Google Scholar
|
|
19
|
Villarroya-Beltri C, Gutiérrez-Vázquez C,
Sánchez-Cabo F, Pérez-Hernández D, Vázquez J, Martin-Cofreces N,
Martinez-Herrera DJ, Pascual-Montano A, Mittelbrunn M and
Sánchez-Madrid F: Sumoylated hnRNPA2B1 controls the sorting of
miRNAs into exosomes through binding to specific motifs. Nat
Commun. 4:29802013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xin H, Li Y, Buller B, Katakowski M, Zhang
Y, Wang X, Shang X, Zhang ZG and Chopp M: Exosome-mediated transfer
of miR-133b from multipotent mesenchymal stromal cells to neural
cells contributes to neurite outgrowth. Stem Cells. 30:1556–1564.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valencia K, Luis-Ravelo D, Bovy N, Antón
I, Martínez-Canarias S, Zandueta C, Ormazábal C, Struman I, Tabruyn
S, Rebmann V, et al: miRNA cargo within exosome-like vesicle
transfer influences metastatic bone colonization. Mol Oncol.
8:689–703. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang Y, Soroush F, Tong Z, Kiani MF and
Wang B: Targeted multidrug delivery system to overcome
chemoresistance in breast cancer. Int J Nanomedicine. 12:671–681.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Melosky B: Review of EGFR TKIs in
metastatic NSCLC, including ongoing trials. Front Oncol. 4:2442014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rosell R, Moran T, Queralt C, Porta R,
Cardenal F, Camps C, Majem M, Lopez-Vivanco G, Isla D, Provencio M,
et al: Screening for epidermal growth factor receptor mutations in
lung cancer. N Engl J Med. 361:958–967. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dhillon S: Gefitinib: A review of its use
in adults with advanced non-small cell lung cancer. Target Oncol.
10:153–170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kani K, Garri C, Tiemann K, Malihi PD,
Punj V, Nguyen AL, Lee J, Hughes LD, Alvarez RM, Wood DM, et al:
JUN-mediated downregulation of EGFR signaling is associated with
resistance to gefitinib in EGFR-mutant nsclc cell lines. Mol Cancer
Ther. 16:1645–1657. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang Z, Lei W, Hu HB, Zhang H and Zhu Y:
H19 promotes non-small-cell lung cancer (NSCLC) development through
STAT3 signaling via sponging miR-17. J Cell Physiol. 233:6768–6776.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Denzer K, Kleijmeer MJ, Heijnen HF,
Stoorvogel W and Geuze HJ: Exosome: From internal vesicle of the
multivesicular body to intercellular signaling device. J Cell Sci.
113:3365–3374. 2000.PubMed/NCBI
|
|
29
|
Kucharzewska P and Belting M: Emerging
roles of extracellular vesicles in the adaptive response of tumour
cells to microenvironmental stress. J Extracell Vesicles. 2:2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kourembanas S: Exosomes: Vehicles of
intercellular signaling, biomarkers, and vectors of cell therapy.
Annu Rev Physiol. 77:13–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X, Liu R, Huang Z, Gurley EC, Wang X,
Wang J, He H, Yang H, Lai G, Zhang L, et al: Cholangiocyte-derived
exosomal long noncoding RNA H19 promotes cholestatic liver injury
in mouse and humans. Hepatology. 68:599–615. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pigati L, Yaddanapudi SC, Iyengar R, Kim
DJ, Hearn SA, Danforth D, Hastings ML and Duelli DM: Selective
release of microRNA species from normal and malignant mammary
epithelial cells. PLoS One. 5:e135152010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alarcón CR, Goodarzi H, Lee H, Liu X,
Tavazoie S and Tavazoie SF: HNRNPA2B1 is a mediator of
m(6)A-dependent nuclear RNA processing events. Cell. 162:1299–1308.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang J, Qin G, Luo M, Chen J, Zhang Q, Li
L, Pan L and Qin S: Reciprocal positive regulation between Cx26 and
PI3K/Akt pathway confers acquired gefitinib resistance in NSCLC
cells via GJIC-independent induction of EMT. Cell Death Dis.
6:e18292015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jeannot V, Busser B, Brambilla E, Wislez
M, Robin B, Cadranel J, Coll JL and Hurbin A: The PI3K/AKT pathway
promotes gefitinib resistance in mutant KRAS lung adenocarcinoma by
a deacetylase-dependent mechanism. Int J Cancer. 134:2560–2571.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chiu CF, Chang YW, Kuo KT, Shen YS, Liu
CY, Yu YH, Cheng CC, Lee KY, Chen FC, Hsu MK, et al: NF-κB-driven
suppression of FOXO3a contributes to EGFR mutation-independent
gefitinib resistance. Proc Natl Acad Sci USA. 113:E2526–E2535.
2016. View Article : Google Scholar : PubMed/NCBI
|