Introduction
Chronic wounds are open wounds of the skin taking
longer than 8 weeks to heal. An impaired healing process is often
associated with chronic wounds (1).
As an ongoing challenge in advanced wound care, chronic wounds can
lead to severe outcomes, including infection, amputation and even
mortality (2). Evidence has emerged
that adipose tissue-derived stem cells (ADSCs) hold great potential
in the stem cell-based therapy of chronic wounds (3,4). ADSCs,
which are also referred to as adipose mesenchymal stromal cells or
adipose tissue-derived stromal cells, are adult stem cells that are
derived from adipose tissue stroma, possess considerable
self-renewal ability and can differentiate into various types of
functional cells. ADSCs show a greater ability to reconstitute
hematopoiesis and repair tissue than bone marrow stromal stem cells
(5,6). However, a significant barrier to the
successful use of mesenchymal stem cells (MSCs) in any potential
cell therapy has been post-implantation cell survival in cutaneous
chronic wound models (7). When cells
are transplanted into damaged cutaneous tissue where they
experience a harsh wound environment, including hypoxia,
inflammation, oxidative stress or other adverse conditions, this
inevitably leads to the poor survival of seed cells following
implantation, which markedly disturbs the therapeutic effects of
MSCs (8). Various strategies have
been used to improve cell survival following implantation (9,10).
However, simple and efficient interventions are required to
strengthen the resistance of cells to apoptosis and increase the
therapeutic effect of ADSCs.
Stromal cell-derived factor-1 (SDF-1) is an
important chemokine that plays a central role in tissue repair and
angiogenesis. SDF-1 exerts its biological function by binding to
two G-protein-coupled receptors: CXC chemokine receptor (CXCR)4 and
CXCR7 (11). SDF-1 functions in the
mobilization and recruitment of stem and progenitor cells,
including hematopoietic stem cells, MSCs and other CXCR4-expressing
cells (11,12). Increasing evidence suggests that the
interaction of SDF-1 with CXCR4 and CXCR7 is critical in
maintaining the proliferation and survival of MSCs in response to
stress (13,14). Studies have confirmed that SDF-1 can
enhance the proliferation, migration, secretion and anti-apoptotic
capacity of MSCs and improve cell transplant survival (15–17).
However, very little is known about the effect of SDF-1 on ADSC
apoptosis and survival. The current authors have previously shown
that the interactions of SDF-1 with CXCR4 and CXCR7 are important
in the paracrine actions, proliferation and migration of ADSCs
(18). In the present study, the
function of SDF-1 as an apoptosis inhibitor in vitro and the
survival rate and therapeutic effect of ADSCs administered to the
chronic wounds of diabetic nude mice were investigated. To the best
of our knowledge, this is the first study to investigate whether
SDF-1 improves the ability of ADSCs to repair chronic wounds.
Materials and methods
ADSC culture
Human ADSCs were isolated from subcutaneous adipose
tissue samples obtained from the liposuction aspirates of patients
undergoing cosmetic liposuction as previously described (19). Three healthy female patients with
abdominal fat accumulation who underwent liposuction surgery at the
Affiliated Hospital of Xuzhou Medical College (Xuzhou, China) were
selected. This study was performed using a protocol approved by the
Institutional Review Board of the Affiliated Hospital of Xuzhou
Medical College and all examinations were performed after obtaining
written informed consent from the patients. ADSCs were maintained
in L-Dulbecco's modified Eagle's medium (DMEM; Invitrogen Life
Technologies, Carlsbad, CA, USA) containing 10% fetal bovine serum
(FBS; Invitrogen Life Technologies). Cells were cultured in a 37°C
humidified incubator with 95% air and 5% CO2. Prior to
experimental use, it was confirmed that the ADSCs possessed the
ability to differentiate into osteoblasts and keratinocytes.
Hypoxic and serum-free conditions
ADSCs from the same passage were obtained from six
flasks and randomly divided into three groups. These were the
control group (L-DMEM containing 10% FBS, normoxic conditions),
apoptosis model group (L-DMEM containing 1% FBS, hypoxic
conditions) and SDF-1 treatment group (L-DMEM containing 1% FBS and
0.5 mg/l SDF-1). In the SDF-1 treatment group, ADSCs were initially
cultured with L-DMEM containing 10% FBS and 0.5 mg/l SDF-1 (R&D
Systems, Minneapolis, MN, USA) in normoxic conditions for 1 h. The
cells were then rinsed twice with PBS and exposed to hypoxic and
serum-free conditions with 0.5 mg/l SDF-1 for 6 h. The apoptosis
model and SDF-1 treatment group ADSCs were put into a hypoxic
incubator (Forma Series II 3110 Water-Jacketed CO2
Incubator; Thermo Fisher Scientific, Waltham, MA, USA) and
maintained in 1% O2 to culture the cells under hypoxic
conditions.
Western blot analysis
Following the treatments, total protein from the
cells was extracted using radio-immunoprecipitation assay lysis
buffer (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) containing
60 µg/ml phenylmethylsulfonyl fluoride. Protein concentrations were
assessed using a bicinchoninic acid protein assay kit (Wuhan Boster
Biological Technology, Ltd., Wuhan, China). Proteins were separated
on SDS-polyacrylamide gel, then transferred to a nitrocellulose
membrane and incubated overnight at 4°C with the following
antibodies: Rabbit monoclonal anti-procaspase 3 (1:1,000; cat. no.
9665), anti-cleaved caspase 3 (1:1,000; cat. no. 9664), anti-poly
ADP ribose polymerase (PARP; 1:1,000; cat. no. 9532) and
anti-cleaved PARP (1:1,000; cat. no. 5625) and mouse monoclonal
anti-β-actin (1:5,000; cat. no. 3700) (Cell Signaling Technology,
Inc., Beverly, MA, USA). Blots were washed four times with TBS
containing 0.1% Tween-20 (TBST), and then incubated with
horseradish peroxidase (HRP)-linked horse anti-mouse and
anti-rabbit secondary antibodies (1:200; cat. no. 7076 and 7074S
respectively; Cell Signaling Technology, Inc.) at room temperature
for 1 h. Antibody binding was detected using a SuperSignal West
Pico Chemiluminescent Substrate kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA) according to the manufacturer's
instructions.
Measurement of apoptosis by Annexin V
analysis
Following treatment, ADSCs were collected and dual
staining with Annexin V-fluorescein isothiocyanate (FITC) and
propidium iodide (PI) was performed according to the manufacturer's
instructions (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China). In
brief, the collected cells were briefly washed with ice-cold
phosphate-buffered saline (PBS) twice and resuspended in 200 µl 1X
binding buffer (Beyotime Institute of Biotechnology, Haimen, China)
containing 5 µl Annexin V-FITC for 15 min followed by 300 µl 1X
binding buffer containing 5 µl PI for 5 min at room temperature in
the dark. Following incubation, the cells were analyzed using a
FACStar flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
An inverted fluorescence microscope (Olympus Corporation, Tokyo,
Japan) was used to observe the cells in the early and late stages
of apoptosis.
Dermal chronic wound model
The diabetic chronic skin wound model was
established according to a previously described procedure with
certain improvements (20). A total
of 8 female mice were used that were housed with a constant
humidity of 50–60% and a temperature of 22–24°C with a 12h/12h
light/dark cycle and access to food and water ad
libitum.
Diabetes was induced in nude mice (Shanghai SLAC
Laboratory Animal Co., Shanghai, China) by the intraperitoneal
injection of streptozotocin (STZ; Sigma-Aldrich, St. Louis, MO,
USA), dissolved in sodium citrate buffer (0.01 M; pH 4.5). Control
mice were treated with DMEM only. Blood glucose levels >250
mg/dl in two consecutive measurements separated by an interval of
48 h were considered to indicate that the animals were diabetic.
After anesthetizing the animals, a full thickness skin defect of
1-cm diameter was made on the back. The wound was covered with a
sterile dressing, and 10 days later the granulation tissue was
debrided, to re-establish a 1-cm-diameter wound. Following each
surgery, 4,000 units penicillin sodium was injected intramuscularly
to prevent infection.
ADSC survival assay in vivo
ADSCs were marked with 5-bromo-2′-deoxyuridine
(5-BrdU; Sigma-Aldrich) prior to injection. Cells suspended in
serum-free DMEM were treated with BrdU solution at a final
concentration of 10 mmol/l at 37°C for 48 h. 1×106 ADSCs
that were pretreated with 0.5mg/l SDF-1 or untreated were injected
around the wound bed. The animals were sacrificed 7 days after
treatment, and wounds were harvested with the surrounding tissue.
The specimens were fixed with 4% paraformaldehyde for 24 h and
embedded in paraffin for histological sectioning. FITC
immunofluorescence staining was performed according to the
manufacturer's instructions (Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China); primary mouse monoclonal
anti-BrdU antibodies (1:1,000; cat. no. sc-51514; Santa Cruz
Biotechnology, Inc.) and FITC-labeled goat anti-mouse secondary
antibodies (1:1,000; cat. no. sc-2010; Santa Cruz Biotechnology,
Inc.) were added in sequence. Immunofluorescence microscopy was
used, and the green fluorescence indicated 5-BrdU-labeled ADSC
nuclei. Counting was conducted in 10 immunofluorescent images per
group to determine the survival rate.
Cutaneous wound repair analysis
The animals were sacrificed by cervical dislocation
10 days after treatment. The wound area was analyzed by tracing the
wound margin and calculating the area using ImageJ software
(version 1.38; National Institutes of Health, Bethesda, MD, USA).
The percentage of wound closure was calculated as follows: Wound
closure index (%) = (original wound area - unhealed wound
area)/original wound area) × 100. Tissue specimens were embedded in
paraffin for sectioning. The 4-mm sections were stained with
hematoxylin and eosin. Using a light microscope, the
re-epithelialization length of each specimen was measured from the
marginal skin to the end of the neoepithelium on each side of the
cross-section.
Statistical analysis
Data are expressed as mean ± standard deviation and
were statistically analyzed using a Student's t-test performed with
SPSS software, version 11.5 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
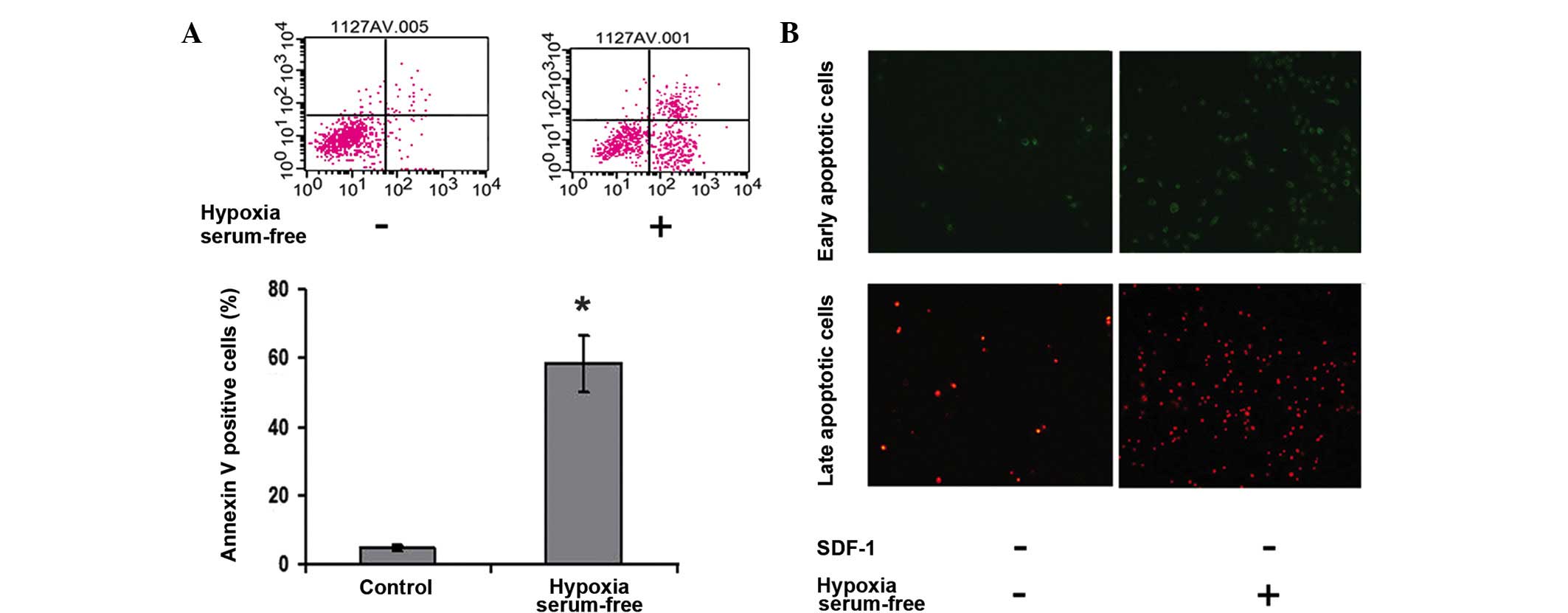
Hypoxia and serum depletion induce
apoptosis in ADSCs
To determine the effect of hypoxia and serum
depletion on cell apoptosis, ADSCs were exposed to hypoxic and
serum-free conditions for 6 h. Flow cytometry was then performed
using cell binding of the fluorescent dye Annexin V-FITC, which
marks the early stage membrane change in apoptosis, and PI staining
of the nucleus, which labels late apoptotic events and necrosis. As
shown in Fig. 1, hypoxia and serum
depletion evidently induced the number of apoptotic cells
(P<0.05).
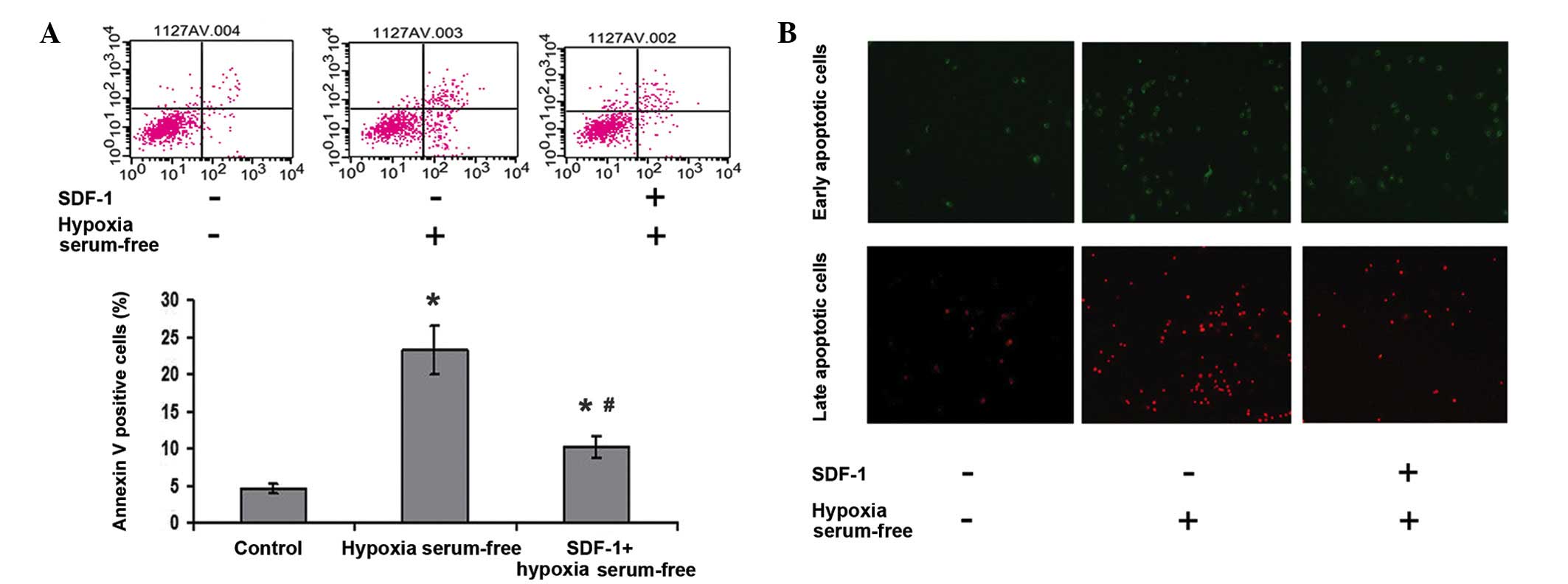
SDF-1 inhibits the hypoxia and serum
depletion-induced apoptosis of ASDCs
To explore the effect of SDF-1 on the apoptosis of
ADSCs induced by hypoxia and serum depletion, ADSCs were treated
with SDF-1 prior to induction, then collected and analyzed by flow
cytometry. As shown in Fig. 2, the
number of apoptotic cells was increased in the hypoxia and serum
depletion (apoptosis model) group compared with that in the control
group (P<0.05); however, in the SDF-1 treatment group the
numbers of early and late apoptotic cells were clearly decreased
compared with those in the apoptosis model group. This suggests
that SDF-1 suppresses the apoptosis of ADSCs induced by hypoxia and
serum depletion.
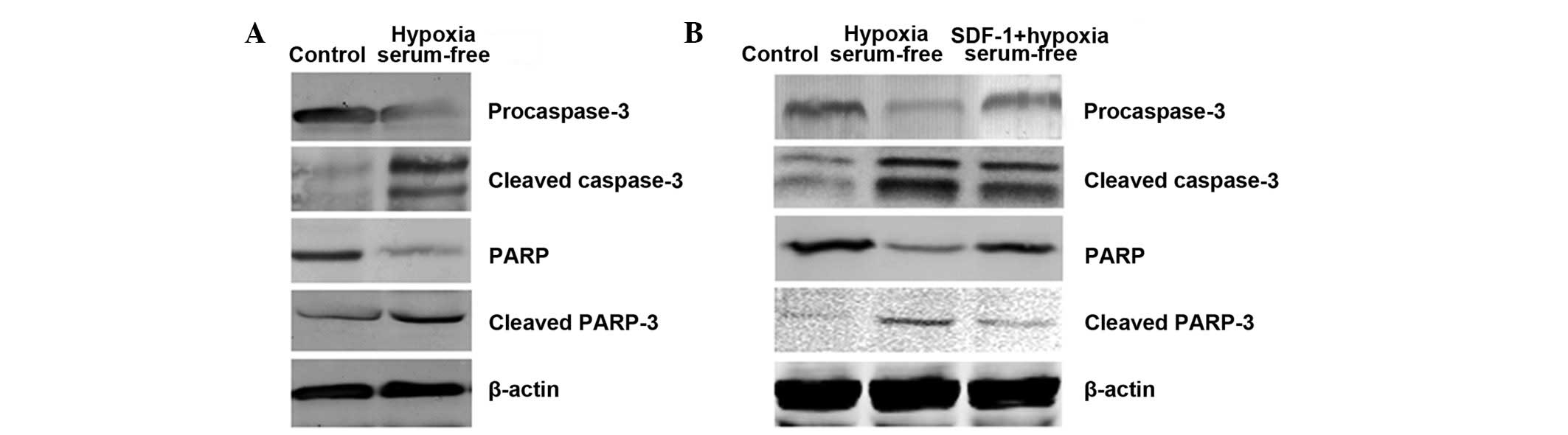
Caspase-3 and PARP activation are
involved in the inhibitory effect of SDF-1 on the hypoxia and serum
depletion-induced apoptosis of ASDCs
To further determine whether hypoxia and serum
depletion-induced apoptosis is associated with activation of the
caspase signaling pathway in ADSCs, caspase-3 and PARP were
detected after induction. As shown in Fig. 3A, compared with that in the control
group, the procaspase-3 level decreased following induction. By
contrast, the cleaved caspase-3 level increased compared with that
in the control group. Similar changes were observed for PARP.
However, when pretreated with SDF-1, procaspase-3 and PARP protein
levels partly recovered and cleaved caspase-3 and PARP protein
levels decreased compared with those in the apoptosis model group.
This shows that SDF-1 protects ADSCs from apoptosis under hypoxic
and serum-free conditions by activating the caspase signaling
pathway.
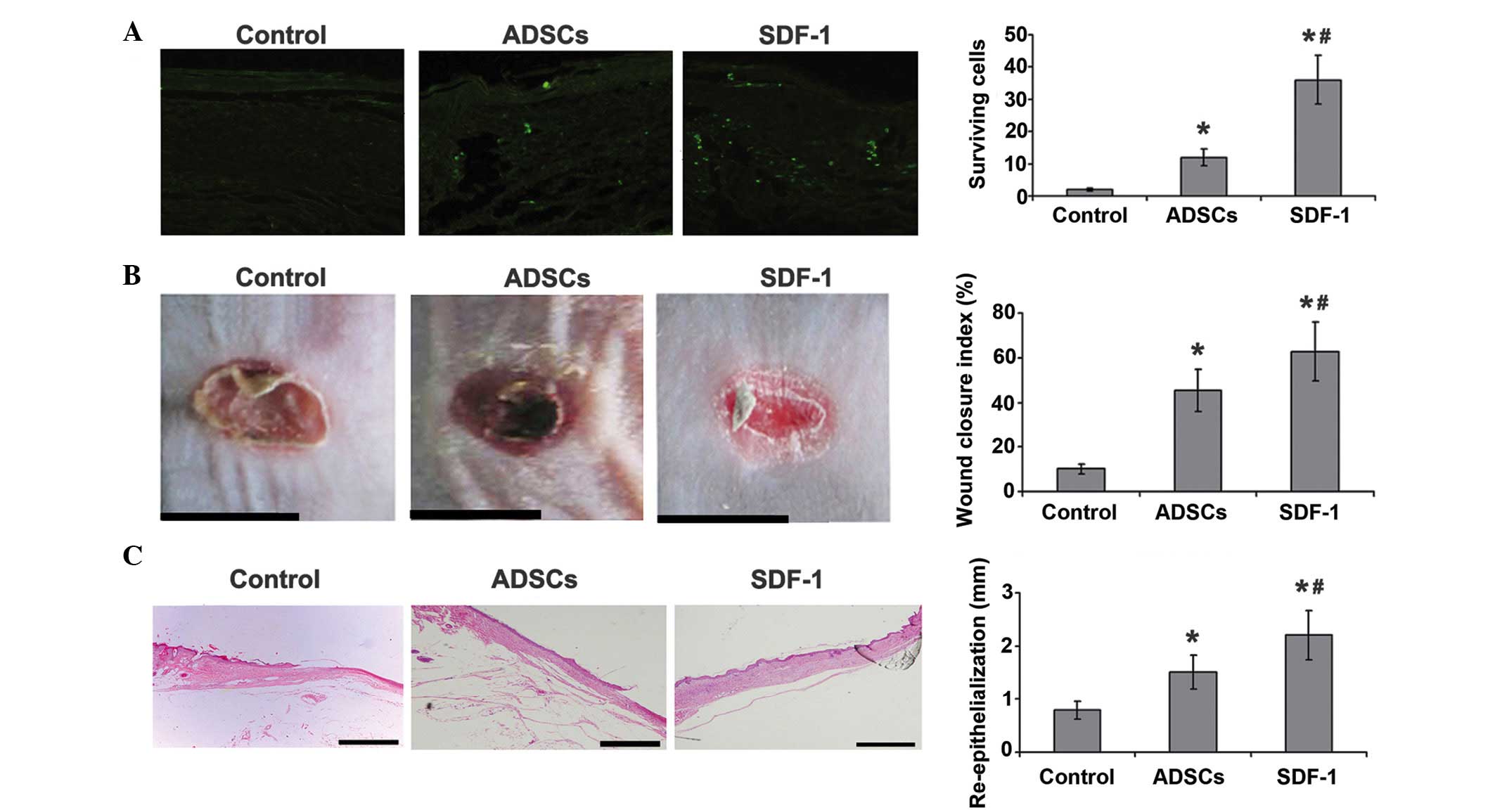
SDF-1 promotes ADSC survival in
vivo
To investigate whether SDF-1 inhibits ASDC apoptosis
in vivo, ADSCs marked with 5-Brd were injected into the
chronic wounds of diabetic mice. It was found that only a few cells
survived in the wound bed 7 days after cell transplantation. SDF-1
pretreatment exhibited a protective effect on ADSC survival. As
shown in Fig. 4A, the number of
surviving cells was significantly higher in the SDF-1 treatment
group than in the apoptosis model group, suggesting that SDF-1
suppresses ADSC apoptosis in vivo.
SDF-1-pretreated ADSCs improve chronic
wound healing in vivo
To further determine whether SDF-1 is able to
promote the wound healing ability of ASDCs in diabetic mice, the
healing of wounds was evaluated 10 days after treatment. The wound
area was significantly smaller in the two groups treated with ADSCs
than in the control group, and the wound closure index for
SDF-1-pretreated ADSCs was significantly better than that in the
other two groups (Fig. 4B).
Histological results for hematoxylin and eosin staining of the
wounds indicate that SDF-1-pretreated ADSCs can promote chronic
wound repair in diabetic mice, as is shown by increased dermal
collagen levels, fibroblast numbers and capillary density. The
re-epithelization length in the SDF-1-pretreated ADSC group was
significantly greater than that in the ADSC group and the control
(Fig. 4C).
Discussion
In physiological cutaneous wound healing, MSCs are
mobilized from host sources and localize to the site of injury,
persisting to improve angiogenesis and support tissue repair and
immunomodulation in cutaneous wounds (21). Despite somewhat challenging
conditions in the wound bed, these host MSCs are able to perform
normal functions. However, in the case of cell therapy where MSCs
are administered to the wound bed, due to a lack of oxygen and
nutrients and the effects of free radicals and inflammation,
>99% of the administered cells die following implantation
(7,14,22).
Poor post-implant cell numbers often lead to the absence of more
definitive results. Therefore, a significant barrier to the
effective use of ADSCs in cell therapy has been the insufficient
post-implant cell survival. As the benefits of ADSC therapy for
cutaneous wound healing are dependent on cell survival in the wound
bed, strategies to improve survival following implantation, such as
hypoxic pretreatment (11) or
genetic modification (10,19) have received some attention. The
present study identified for the first time, to the best of our
knowledge, that SDF-1 pretreatment protects ADSCs from apoptosis in
a hypoxic and serum-free environment. Furthermore, in vivo
experiments established that the post-implant cell survival and
chronic wound healing ability of ADSCs were increased following
pretreatment with SDF-1 in a diabetic mouse model of chronic wound
healing.
A number of studies have confirmed that SDF-1
enhances the proliferation, migration and secretion capacity of
BMSCs and cardiac stem cells so as to improve the cell therapeutic
effect in vivo (16,17,23). For
cell apoptosis, Yin et al found that SDF-1 inhibits hypoxia
and serum deprivation-induced apoptosis in MSCs through
phosphoinositide 3-kinase/Akt and extracellular signal-regulated
kinase 1/2 signaling pathways (24).
Herberg et al demonstrated that SDF-1 mediates MSC survival
through the enhancement of autophagy (14). Therefore, it was hypothesized that
SDF-1 pretreatment may promote the activity and survival of ADSCs
in an oxygen and nutrient-deficiency environment. In the present
study, harsh ischemic microenvironment stress was mimicked by the
use of serum-free and hypoxic conditions and these were found to
induce marked apoptosis of ADSCs within 6 h. Moreover, it was found
that the apoptosis induced in hypoxic and serum-free conditions was
ameliorated by SDF-1. Furthermore, the caspase-3 and PARP signaling
pathway may be involved in the inhibition of cell apoptosis by
SDF-1.
SDF-1 is a chemokine considered to play an important
role in CXCR4+ and CXCR7+ progenitor cell
mobilization and homing for ischemic or damaged tissues. It is
secreted by endothelial and reticular cells and upregulated in
acute cutaneous wounds for several days following trauma (25). SDF-1 upregulation in acute wounds
takes part in the initial localization, retention and support of
CXCR4+ stem cells, thus playing a critical role in
physiological cutaneous wound healing (26). However, SDF-1 is expressed at low
levels in chronic wounds, and may be hydrolytically inactivated by
matrix metalloproteinases, leading to an SDF-1/CXCR4, CXCR7 axis
disorder, hindering the migration and homing of cells and resulting
in chronic healing or an absence of healing (27,28).
Therefore, in the cell therapy of chronic wounds, the wound
environment becomes increasingly harsh, with excessive inflammation
and an environment that is not conducive to angiogenesis (29,30). The
deficiency in the normal physiological process of cell mobilization
and activation that SDF-1 induces leads to poor ADSC survival and
homing. Therefore, it was hypothesized in the present study that
pretreatment with SDF-1 may simulate the physiological process of
cell mobilization to a certain extent and promote the therapeutic
effect of ADSCs in cutaneous chronic wounds. Thus, ADSCs, either
untreated or pretreated with 0.5 mg/l SDF-1, were administered by
injection in a diabetic chronic wound model. It was found that the
surviving cell number was significantly higher, and the wound
closure index and length of re-epithelization were improved by
SDF-1 pretreatment of the ADSCs, indicating a significant promoting
effect of SDF-1 on the survival and therapeutic effect of ADSCs
in vivo.
In conclusion, the present study demonstrated that
the chemokine SDF-1 is important for ADSC survival and the
prevention of apoptosis. Pretreating ADSCs with SDF-1 prior to
local administration or intravenous transplantation may
significantly enhance cell survival and the therapeutic effect of
ADSCs in cutaneous chronic wounds.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81571901 and
81501671).
References
|
1.
|
Montfrans CV, Stok M and Geerkens M:
Biology of chronic wounds and new treatment strategies. Phlebology.
29(Suppl): 165–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Charbonneau L, Perrenoud B, Gallant S,
Lehn I and Champier V: Chronic wound management in hospital care
units - a literature review. Rech Soins Infirm. 96:58–68. 2009.(In
French). View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Fumimoto Y, Matsuyama A, Komoda H, et al:
Creation of a rich subcutaneous vascular network with implanted
adipose tissue-derived stromal cells and adipose tissue enhances
subcutaneous grafting of islets in diabetic mice. Tissue Eng Part C
Methods. 15:437–444. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Ohmura Y, Tanemura M, Kawaguchi N, et al:
Combined transplantation of pancreatic islets and adipose
tissue-derived stem cells enhances the survival and insulin
function of islet grafts in diabetic mice. Transplantation.
90:1366–1373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Zhu Y, Liu T, Song K, Fan X, Ma X and Cui
Z: Adipose-derived stem cell: A better stem cell than BMSC. Cell
Biochem Funct. 26:664–675. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Han C, Zhang L, Song L, Liu Y, Zou W, Piao
H and Liu J: Human adipose-derived mesenchymal stem cells: A better
cell source for nervous system regeneration. Chin Med J (Engl).
127:329–337. 2014.PubMed/NCBI
|
|
7.
|
Nuschke A: Activity of mesenchymal stem
cells in therapies for chronic skin wound healing. Organogenesis.
10:29–37. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Das R, Jahr H, van Osch GJ and Farrell E:
The role of hypoxia in bone marrow-derived mesenchymal stem cells:
Considerations for regenerative medicine approaches. Tissue Eng
Part B Rev. 16:159–168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Liu H, Xue W, Ge G, et al: Hypoxic
preconditioning advances CXCR4 and CXCR7 expression by activating
HIF-1α in MSCs. Biochem Biophys Res Commun. 401:509–515. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Shevchenko EK, Makarevich PI, Tsokolaeva
ZI, et al: Transplantation of modified human adipose derived
stromal cells expressing VEGF165 results in more efficient
angiogenic response in ischemic skeletal muscle. J Transl Med.
11:1382013. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
de Ruiz Almodovar C, Luttun A and
Carmeliet P: An SDF-1 trap for myeloid cells stimulates
angiogenesis. Cell. 124:18–21. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Togel FE and Westenfelder C: Role of SDF-1
as a regulatory chemokine in renal regeneration after acute kidney
injury. Kidney Int Suppl. 1:87–89. 2011. View Article : Google Scholar
|
|
13.
|
Nakamura Y, Ishikawa H, Kawai K, Tabata Y
and Suzuki S: Enhanced wound healing by topical administration of
mesenchymal stem cells transfected with stromal cell-derived
factor-1. Biomaterials. 34:9393–9400. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Herberg S, Shi X, Johnson MH, Hamrick MW,
Isales CM and Hill WD: Stromal cell-derived factor-1β mediates cell
survival through enhancing autophagy in bone marrow-derived
mesenchymal stem cells. PLoS One. 8:e582072013. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Liu H, Liu S, Li Y, et al: The role of
SDF-1-CXCR4/CXCR7 axis in the therapeutic effects of
hypoxia-preconditioned mesenchymal stem cells for renal
ischemia/reperfusion injury. PLoS One. 7:e346082012. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Ghadge SK, Mühlstedt S, Ozcelik C and
Bader M: SDF-1α as a therapeutic stem cell homing factor in
myocardial infarction. Pharmacol Ther. 129:97–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Wang K, Zhao X, Kuang C, et al:
Overexpression of SDF-1α enhanced migration and engraftment of
cardiac stem cells and reduced infarcted size via CXCR4/PI3K
pathway. PLoS One. 7:e439222012. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Li Q, Zhang A, Tao C, Li X and Jin P: The
role of SDF-1-CXCR4/CXCR7 axis in biological behaviors of adipose
tissue-derived mesenchymal stem cells in vitro. Biochem Biophys Res
Commun. 441:675–680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Li Q, Li PH, Hou DJ, et al: EGF enhances
ADSCs secretion via ERK and JNK pathways. Cell Biochem Biophys.
69:189–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Martínez-Santamaría L, Conti CJ, Llames S,
et al: The regenerative potential of fibroblasts in a new
diabetes-induced delayed humanised wound healing model. Exp
Dermatol. 22:195–201. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Li H and Fu X: Mechanisms of action of
mesenchymal stem cells in cutaneous wound repair and regeneration.
Cell Tissue Res. 348:371–377. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Gojo S, Gojo N, Takeda Y, et al: In vivo
cardiovasculogenesis by direct injection of isolated adult
mesenchymal stem cells. Exp Cell Res. 288:51–59. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Liu X, Zuo D, Fan H, et al:
Over-expression of CXCR4 on mesenchymal stem cells protect against
experimental colitis via immunomodulatory functions in impaired
tissue. J Mol Histol. 45:181–193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Yin Q, Jin P, Liu X, et al: SDF-1α
inhibits hypoxia and serum deprivation-induced apoptosis in
mesenchymal stem cells through PI3K/Akt and ERK1/2 signaling
pathways. Mol Biol Rep. 38:9–16. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Zong ZW, Cheng TM, Su YP, et al: Crucial
role of SDF-1/CXCR4 interaction in the recruitment of transplanted
dermal multipotent cells to sublethally irradiated bone marrow. J
Radiat Res (Tokyo). 47:287–293. 2006. View
Article : Google Scholar
|
|
26.
|
Askari AT, Unzek S, Popovic ZB, et al:
Effect of stromal-cell-derived factor 1 on stem-cell homing and
tissue regeneration in ischaemic cardiomyopathy. Lancet.
362:697–703. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Brem H and Tomic-Canic M: Cellular and
molecular basis of wound healing in diabetes. J Clin Invest.
117:1219–1222. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Litwiniuk M, Bikowska B, Niderla-Bielińska
J, et al: Potential role of metalloproteinase inhibitors from
radiation-sterilized amnion dressings in the healing of venous leg
ulcers. Mol Med Rep. 6:723–728. 2012.PubMed/NCBI
|
|
29.
|
Khanna S, Biswas S, Shang Y, et al:
Macrophage dysfunction impairs resolution of inflammation in the
wounds of diabetic mice. PLoS ONE. 5:e95392010. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Krisp C, Jacobsen F, McKay MJ, Molloy MP,
Steinstraesser L and Wolters DA: Proteome analysis reveals
antiangiogenic environments in chronic wounds of diabetes mellitus
type 2 patients. Proteomics. 13:2670–2681. 2013. View Article : Google Scholar : PubMed/NCBI
|