Introduction
Sepsis is one of the leading causes of mortality and
morbidity in children and adults (1). A hostile gut environment can always be
found in patients with sepsis. Intestinal epithelium dysfunction
and microbiota imbalance induced by critical illnesses results in
increased translocation of bacteria to the blood, which contributes
to the formation and aggravation of sepsis (2). Probiotics, the living microorganisms
colonizing in the intestine, bring benefits to the host through
rebalancing microbiota, modulating intestinal barrier function
(3), restraining tumorigenesis
(4) and enhancing the immune system
(5). Currently, a growing number of
articles have indicated the potential of probiotics in numerous
diseases, including diarrhea (6),
allergic diseases (7), inflammatory
bowel disease (3) and sepsis
(2). However, the majority of these
articles focus on evaluating the prevention and treatment effects
of probiotics on diseases. Only a small number of studies attempt
to explain the precise molecular mechanism underlying the
beneficial effects of these microorganisms, including the
regulation of gene expression (5).
Thus, there is a growing need for exploring the exact mechanisms
underlying the protective effects of probiotics in different
diseases. The present study sought to delineate the potential
mechanism of pre-administered probiotic therapy in experimental
sepsis using a C57BL6 mouse peritonitis model.
Metabolomics, one of the ‘-omics’ technologies, can
provide quantitative measures to distinguish global changes in the
metabolic profiles of individuals responding to pathophysiological
stimuli or genetic modification (8).
In the development of sepsis, metabolic disorder has been regarded
as a key feature. Therefore, metabolomics will provide an effective
way to identify sepsis, and assess prognosis and insight into the
mechanisms of sepsis. In the present study, metabolic differences
between serum samples from cecal ligation and puncture
(CLP)-induced septic mice, septic mice pretreated with
Lactobacillus rhamnosus GG (LGG) and sham mice were analyzed
using ultra-performance liquid chromatography coupled with
quadrupole time-of-flight mass spectrometry (UPLC-QTOF-MS).
Materials and methods
Ethics statement
All procedures for animal care and use were approved
by the Animal Care Ethics Committee of the First Affiliated
Hospital, Zhejiang University (Zhejiang, China). A total of 42 four
week-old male C57BL6 mice (weight, 11 g) were purchased from
Zhejiang University and housed in specific pathogen-free animal
facilities under a standard 12-h-light/12-h-dark cycle at room
temperature with relative humidity between 50 and 60% and ad
libtum access to food and water. Standard mouse diet and water
were administered throughout the study.
Reagents and materials
The probiotic capsule contains 280 mg inulin powder
and 1010 colony forming unit (CFU) LGG (Culturelle;
ConAgra Foods, Omaha, NE, USA). High performance liquid
chromatography-grade acetonitrile, leucine-enkephalin and formic
acid were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Deionized water was obtained from an ultrapure water machine (Merck
Millipore, Bedford, MA, USA). All standard samples were purchased
from Sigma-Aldrich.
Probiotic administration and septic
peritonitis model
Four week-old male C57BL6 mice were orally gavaged
with 200 µl/day LGG (2×109 CFU/ml) or normal saline
(control) 4 weeks prior to CLP. To establish the murine septic
peritonitis model, the mice were anesthetized with an
intraperitoneal injection of 4% chloral hydrate (10 ml/kg). A 1-cm
incision was made in the middle of the abdomen and the cecum was
then exposed carefully through the incision. In order to induce
mid-grade sepsis, the cecum was ligated at the middle of the bottom
and distal pole, and was punctured from the mesenteric towards the
antimesenteric direction using a 23-gauge needle. A droplet of
faeces was extruded through the holes, and the cecum was relocated
into the abdominal cavity. Finally, the fascia and skin incision
were closed in various layers. The mice in the sham group underwent
the same procedure but without the cecum ligation and puncture. In
all of the mice, 1 ml pre-warmed normal saline was injected
subcutaneously following CLP surgery (9).
Survival studies
Survival studies were conducted in order to
determine whether LGG pretreatment decreases the mortality
resulting from CLP-induced sepsis. Mice pretreated with LGG (n=8)
or normal saline (n=8) for 4 weeks were subjected to CLP. Sham mice
(n=5) underwent sham laparotomy. Following the operation, all mice
had free access to water and food and were closely observed for a 7
day survival rate.
Histological study
Ileum tissues were collected from mice (n=21) 24 h
after CLP operation and fixed in 10% formalin for 24 h. The fixed
ileum tissues were embedded in paraffin and sectioned at 4 µm. The
sections were stained with hematoxylin and eosin for histological
observation in order to examine the pathology of intestinal injury
induced by CLP.
Blood sample collection and
preparation
Blood samples were obtained at 24 h from the orbital
vein of the mice following CLP. In all of the mice 1 ml of blood
was introduced in the EP tube without any anticoagulant substances.
The blood collected was kept at 4°C for 3 h prior to centrifugation
(3,000 × g, 5 min, 4°C), then carefully absorbed and injected into
a clean EP tube and then stored at −80°C.
Prior to the metabolomics analysis, the serum sample
was thawed at room temperature. In order to precipitate the protein
in the serum sample, 200 µl sample was added into 600 µl ice-cold
acetonitrile. Next, the mixture was vortexed and centrifuged at
12,000 × g for 15 min and 4°C. Finally, the supernatant was moved
into a glass bottle and stored at 4°C for UPLC-MS analysis. To
evaluate reproducibility and stability of the UPLC-MS system, 10 µl
of each sample was added into a bottle to generate a pooled quality
control sample, which was inserted every five samples throughout
the run. The stable retention times and tight overlap of the peaks
reveal high repeatability and stability of the analytical
system.
UPLC-QTOF-MS analysis
The sample analysis was performed using an acquity
ultra-performance liquid chromatography system (Waters Corporation,
Milford, MA, USA) with a conditioned autosampler at 4°C. A 5 µl
sample was injected into a Waters BEH C8 column (2.1×100 mm, inner
diameter × length, 1.7 µm particle size) and kept at 50°C with a
thermostat. Mobile phases, delivered at 300 µl/min, were formed of
0.1% formic acid and 99.9% water (A), or 0.1% formic acid and 99.9%
acetonitrile (B). The gradient elution program was as follows: 97%
A in the initial 0–7 min followed by 20% A in 8 min and
consequently 2% A in 16 min. The conditions were kept constant for
5 min and then changed to 100% B in 50 sec and maintained for 3
min. Next the compound was returned to 97% A in 25 min and kept for
5 min.
Mass spectrometry (Waters Q-TOF Premier) detection
was performed in the positive electrospray ionization mode. The
apparatus was previously calibrated using sodium formate. A lock
mass of leucine enkephalin was used for an accurate mass
determination setting at m/z 556.2771 in the positive
ion mode. The concentration of leucine enkephalin was 0.5 ng/µl.
The detection parameters were optimized as follows: Capillary
voltage of 3 kV and cone voltage of 40 V. The scan time was 0.3 sec
covering the 50 to 1,000 Dalton mass range. The source temperature
was set to 120°C and the desolvation gas temperature was 450°C.
Nitrogen was used as the nebulizer gas at a flow rate of 600
l/h.
Data analysis
The UPLC-MS data collected in positive ion mode were
obtained and preprocessed using the Masslynx software version 4.1
(Waters Corporation). This application was used for peak alignment
to obtain a list containing the m/z, retention time and
intensities for all peaks detected. The preprocessed data were
exported and analyzed using the SIMCA-P+12.0 (MKS Umetrics AB,
Umea, Sweden). All the data were normalized and Pareto scaled prior
to a supervised partial least-squares-latent structure discriminate
analysis (PLS-DA). PLS-DA was used to highlight the difference
between groups and obtain metabolites that contributed to the
classification. Potential biomarkers were identified according to
the ‘variable of importance in the project’ (VIP) values and
S-plots. To further identify the potential biomarkers, PubChem
compound (http://www.ncbi.nlm.nih.gov/) and KEGG (http://www.genome.jp/kegg/) databases were searched in
order to match the selected ion spectrum with the message of
metabolites obtained from databases. MS analysis was conducted to
validate the potential biomarkers.
Statistical analysis
The homogeneity of variances was verified using
Bartlett's test. One-way analysis of variance (with post-hoc
Bonferroni) was then performed to compare the spectral variables
among different serum samples. Survival studies were conducted via
the log-rank test. P≤0.05 was used to indicate a statistically
significant difference. Statistical analysis was conducted using
GraphPad Prism version 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA).
Results
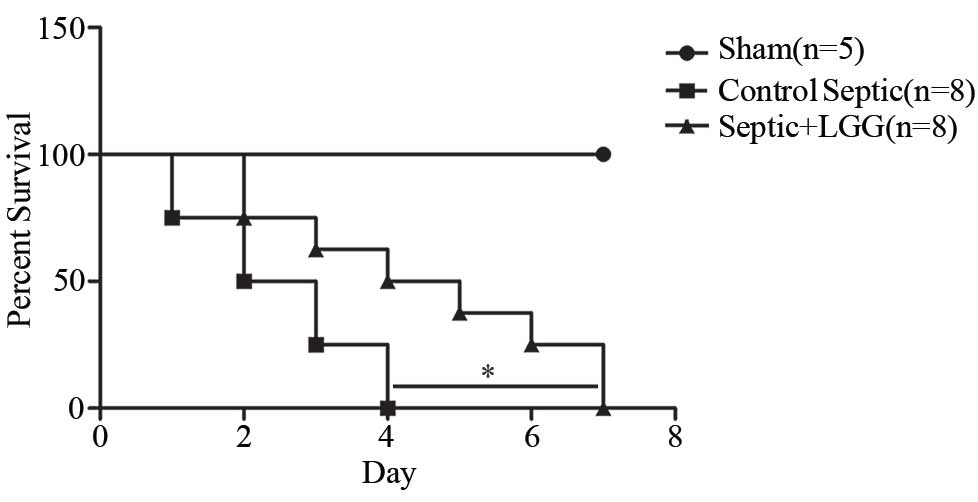
LGG reduces mortality in septic
peritonitis
To determine whether LGG pretreatment will decrease
mortality in experimental peritonitis-induced sepsis, 8 LGG
pretreated mice and 8 control mice were subjected to CLP, and were
observed for a 7 day survival rate (Fig.
1). Septic mice pretreated with LGG had a significantly
improved 7 day survival rate compared with the control septic mice
(P=0.029). Furthermore, 5 sham mice survived.
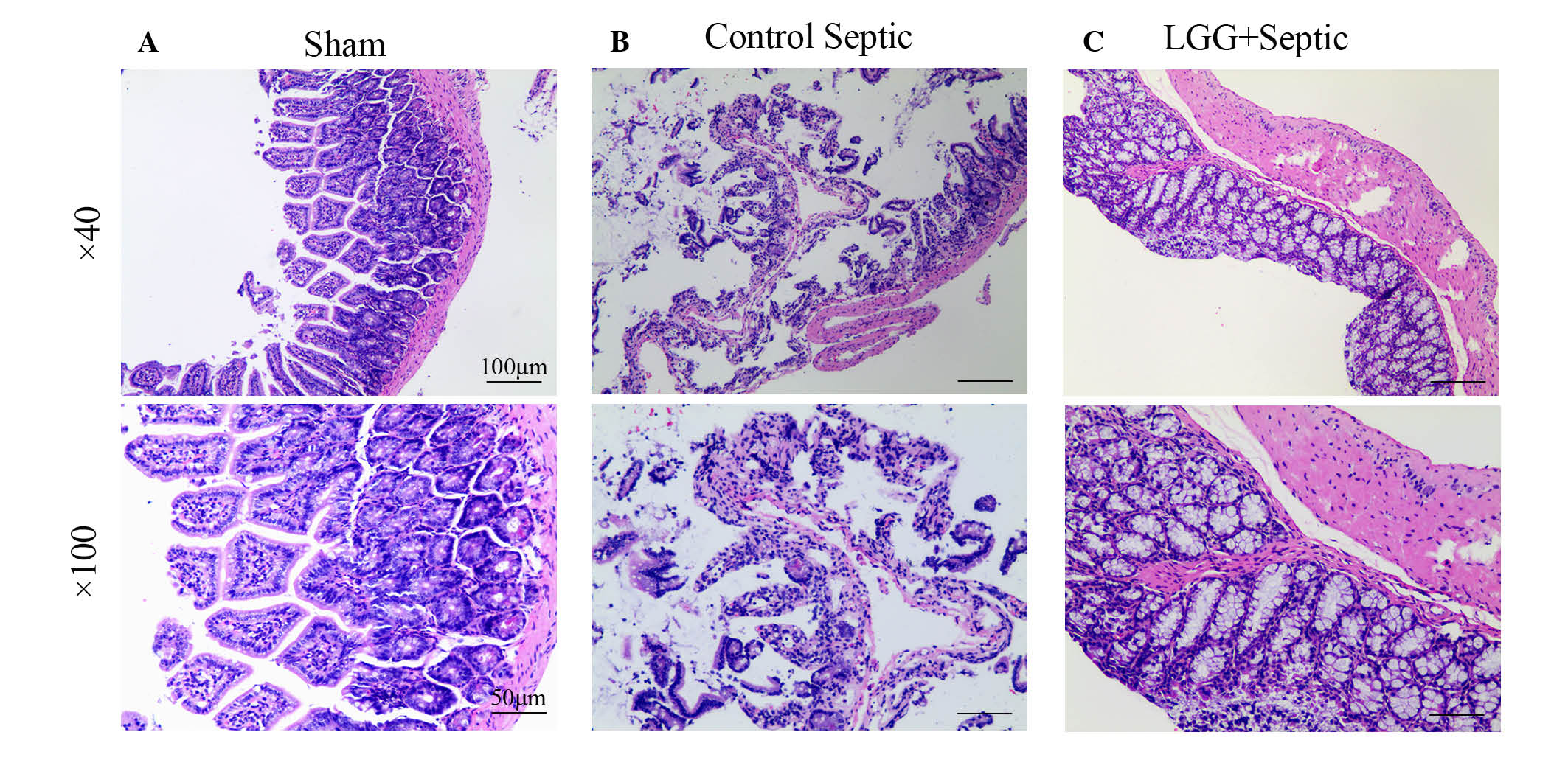
Alleviation of injury in intestinal
mucosa occurred following treatment with LGG
Ileum tissues in sham mice were histologically
normal in all layers (Fig. 2A). In
control septic mice, the lamina propria was almost entirely
digested and disintegrated accompanied with exfoliating villi that
made the epithelial structure difficult to identify (Fig. 2B). In LGG pretreated septic mice,
although the lamina propria was exposed to exfoliating villi the
epithelial appearance was regular with a large number of crypts
(Fig. 2C). Ileum mucosal damage can
evidently be observed in septic mice. Furthermore, the damage in
septic mice pretreated with LGG was less compared with the control
septic mice. In addition, marked differences were observed in the
mucosal thickness, villi number and length amongst the LGG
pretreated septic, control septic and sham mice.
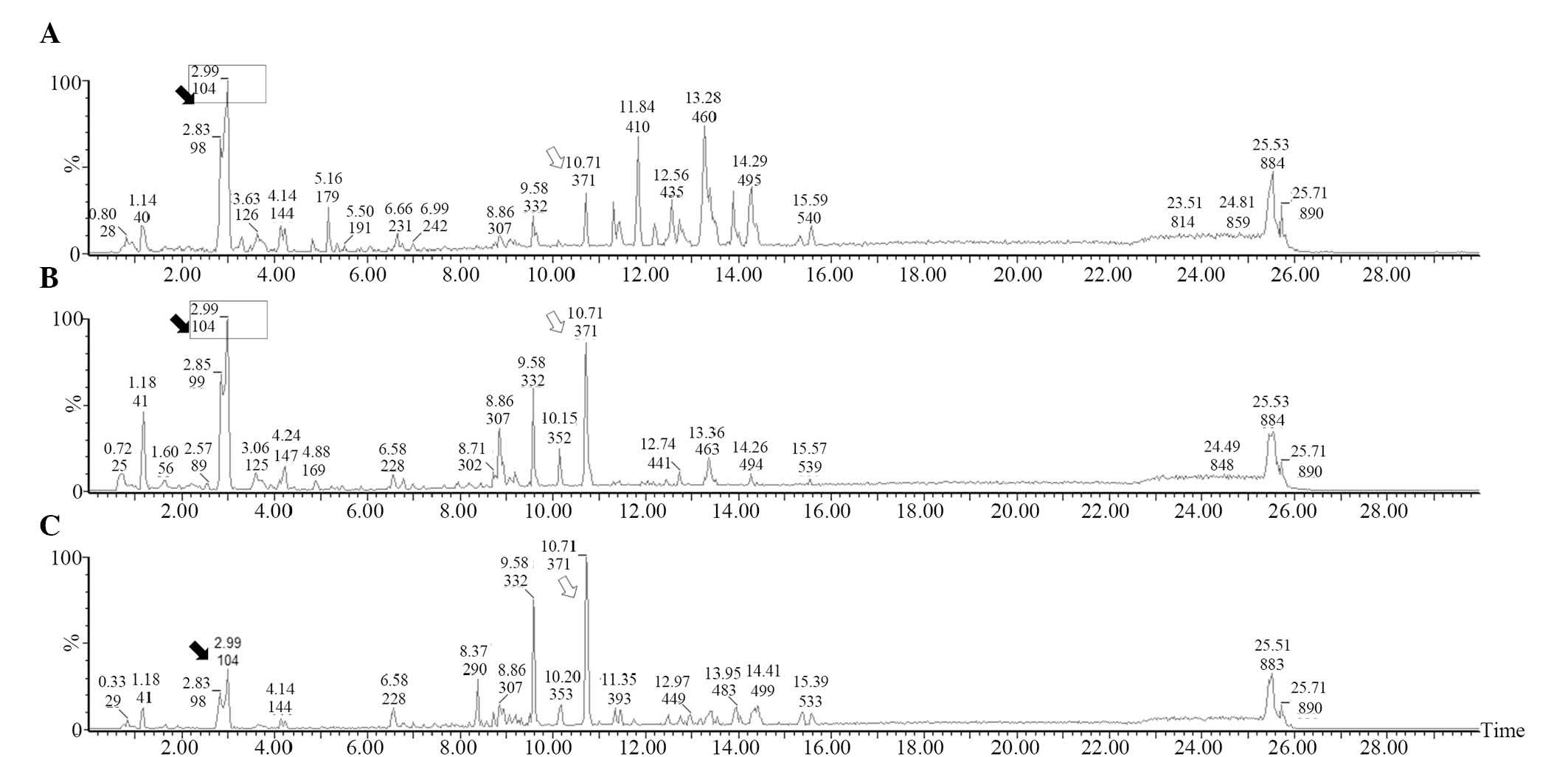
Results of the UPLC-MS analysis
Low-molecular-weight metabolites in the serum of the
sham, septic mice pretreated with LGG and control septic mice are
presented in base peak intensity chromatograms (Fig. 3). Compared with the sham mice, it was
evident that the levels of a number of metabolites increased
(hollow arrow) in the septic mice while others decreased (black
arrow). However, compared with the control septic mice, the peak
patterns in septic mice pretreated with LGG were more similar to
those in sham mice (box), indicating that early therapy with LGG
can normalize the metabolic profile in septic mice.
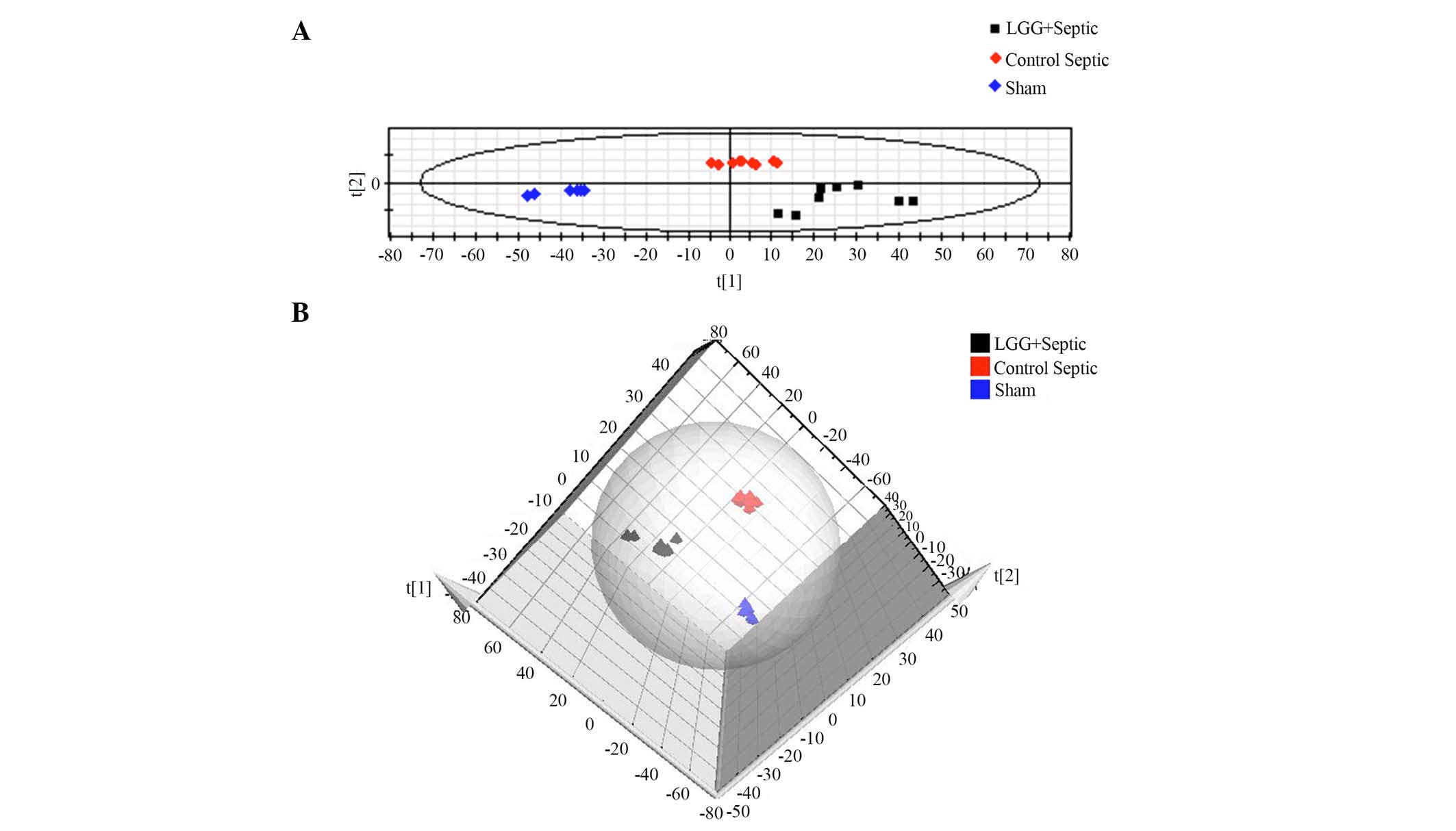
PLS-DA analysis of UPLC-MS data
In order to demonstrate the difference of the
metabolic profile amongst the three groups, a supervised PLS-DA
method was used to analyze the multivariate data. As shown in
Fig. 4, where each point represents
an independent sample, there was a distinguished classification
among all three groups. In-group similarity was observed in each
group, and the three distinct clusters represented the LGG
pretreated septic mice, control septic mice and sham mice that were
clearly demonstrated on the PLS-DA scores plot indicating metabolic
differences among the three groups. In addition, the distance
between LGG pretreated septic mice and sham mice was closer
compared with the distance between control septic and sham mice,
indicating a shift towards a normalized metabolic profile in the
septic mice pretreated with LGG.
Candidate biomarker
identification
According to the results of the VIP values (the top
19 VIPs) and the S-plot, 19 metabolites were selected as candidate
biomarkers (Table I). Besides the
S-plot, different metabolites that significantly contribute to the
separation of the three groups were identified among LGG pretreated
septic, control septic and sham mice. The PubChem compound and KEGG
databases were searched to compare the MS data using chemical
standards, and the results were used to help us identify potential
biomarkers. The biomarkers identified are summarized in Table I. The levels of two groups of
biomarkers-lysophosphatidylcholines (LPCs) and phosphatidylcholines
(PCs) were different between the septic and the sham mice. Amongst
them, PCs were increased and LPCs that contain polyunsaturated
fatty acids were decreased in septic mice, whereas saturated fatty
acid LPCs demonstrated no significant difference between the septic
and sham mice. The difference between LGG pretreated septic mice
and sham mice were less compared with the control septic and sham
mice, although 19 discriminating metabolites revealed an unclear
distinction between LGG pretreated septic and control septic mice
(P>0.05).
 | Table I.Serum biomarkers associated with
sepsis. |
Table I.
Serum biomarkers associated with
sepsis.
|
| Trend |
|---|
|
|
|
|---|
| No. | Rt | m/z | VIP | Adduction | Identify results | Group 1–2 | Group 1–3 | Group 2–3 |
|---|
| 1 | 11.88 | 991.6767 | 8.61692 | [2M+H]+ | LPC16:0 | ns | ns | ns |
| 2 | 11.9 | 496.3404 | 8.48299 | [M+H]+ | LPC16:0 | ns | ns | ns |
| 3 | 19.05 | 760.5878 | 6.86831 | [M+H]+ | 16:0/18:1-PC | ns | ↑↑↑ | ↑↑↑ |
| 4 | 19.05 | 782.5693 | 5.97245 | [M+Na]+ | 16:0/18:1-PC | ns | ↑↑↑ | ↑↑↑ |
| 5 | 18.31 | 780.5543 | 5.01658 | [M+Na]+ | 16:0/18:2-PC | ns | ↑↑↑ | ↑↑↑ |
| 6 | 18.18 | 478.3293 | 4.98452 | [M+H]+ | PC neutral loss
fragment | ns | ↓↓ | ↓↓↓ |
| 7 | 17.92 | 804.5538 | 7.03144 | [M+Na]+ | 18:2/18:2-PC | ns | ↓↓ | ↓ |
| 8 | 18.11 | 806.5718 | 4.87843 | [M+H]+ | 22:6/16:0-PC | ns | ↑↑↑ | ↑↑↑ |
| 9 | 18.31 | 758.5716 | 4.68084 | [M+H]+ | 16:0/18:2-PC
LPC22:6 | ↑ | ↑↑↑ | ns |
| 10 | 11.44 | 568.3413 | 4.63146 | [M+H]+ | 22:6/18:0-PC
LPC18:1 | ns | ↓↓↓ | ↓ |
| 11 | 19.23 | 834.6033 | 4.60648 | [M+H]+ | 18:0/18:2-PC | ns | ↑↑↑ | ↑↑ |
| 12 | 11.43 | 520.3405 | 4.55215 | [M+H]+ | 22:6/16:0-PC | ns | ↓↓↓ | ↓↓↓ |
| 13 | 19.49 | 786.6028 | 4.52772 | [M+H]+ | 18:0/18:2-PC
LPC18:0 | ns | ↑↑↑ | ↑ |
| 14 | 18.11 | 828.5551 | 4.29195 | [M+Na]+ | LPC16:0 | ns | ↑↑↑ | ↑↑↑ |
| 15 | 19.49 | 808.5854 | 4.16349 | [M+Na]+ | LPC18:1 | ns | ↑↑↑ | ↑ |
| 16 | 13.31 | 546.354 | 3.95349 | [M+Na]+ | LPC16:0 isomer | ns | ns | ns |
| 17 | 11.54 | 518.3221 | 3.88142 | [M+Na]+ | LPC16:0 | ns | ns | ns |
| 18 | 12.29 | 544.339 | 3.7194 | [M+Na]+ | LPC18:1 | ns | ↓↓↓ | ↓↓ |
| 19 | 11.9 | 518.3223 | 3.59502 | [M+Na]+ | LPC16:0 isomer | ns | ns | ns |
Discussion
Growing evidence indicates that human physiology and
metabolism are regulated by intestinal microbiota (10). Consisting of different species of
bacteria, the intestinal microbiota is important in maintaining
integrity of the epithelial barrier and participating in mucosal
immunity (11). However, bacteremia
may result from an imbalanced intestinal microbiota of a patient
when critical illness occurs (12).
Probiotics can provide health benefits and protect the body from
diseases that may attribute to their ability to rebalance the
microbiota, and as a consequence affect metabolism (10). However, a series of clinical trials
have demonstrated that probiotic administration does not reduce
mortality rates in patients with a critical illness effectively
(13–16). One possible reason for this is that
probiotic administration is not provided as early as possible. In
the present study on septic mice, probiotics were administered
orally prior to infection to prevent the progress of sepsis. The
data demonstrated a significant reduction in mortality rate and an
enhancement of ileum epithelial integrity in probiotic pretreated
septic mice compared with control septic mice, indicating that
early administration of probiotics has a protective effect against
infection.
In the present study, 1,714 peaks were detected in
the serum samples using UPLC-MC. According to PLS-DA, three
distinct clusters represented the three different groups observed.
Each group was in its own cluster with in-group similarity
observed, separated and apart from the others. Candidate biomarkers
of septic mice pretreated with probiotics were selected according
to VIP values and the S-plot. Metabolites that contributed
significantly to the discrimination were identified, namely PCs and
LPCs. The majority of the PCs were increased in septic mice, while
the majority of the LPCs were markedly decreased. Furthermore, LPCs
that contain polyunsaturated fatty acids [e.g. LPC (18:1) and LPC
(22:6)] were decreased, whereas saturated fatty acid LPCs [e.g. LPC
(16:0) and LPC (18:0)] revealed no statistical difference in septic
mice.
PCs is the most abundant phospholipid class found in
intestinal mucus (17). They are
important in establishing a hydrophobic surface of mucus, which
protects the intestinal epithelial cells against commensal bacteria
from the intestinal lumen (18). In
a patient with ulcerative colitis (UC) whose PC level in the mucus
had significantly decreased due to inflammation, added PC elevated
the concentration of PC in the mucus. As a result, inflammation
improved and even resolved in patients with UC (19). The data indicate the important role
of the mucus PC in barrier function. During the process of
inflammation, a series of pro-inflammatory mediators, including the
cytokine tumor necrosis factor (TNF) are released by intestinal
epithelial cells (20). TNF results
in increasing intestinal permeability (21), inhibition of nutrient absorption
(22), and alteration of cell events
(23). Exogenous addition of PC
attenuated the level of serum cytokines (TNF-α and interleukin-10),
and markedly inhibited the processes induced by cytokines such as
TNF (17). In the present study, the
PC concentration that was significantly elevated in septic mice
compared with sham mice indicates an anti-inflammatory effect of
PC. However, the level of PC (16:0/18:2-PC) in LGG pretreated
septic mice were lower compared with the control septic mice,
suggesting that LGG pretreatment may reduce the inflammatory
response.
The majority of LPCs are derived from PCs, and are
formed through different mechanisms (24). A number of them are catalyzed by the
glycoprotein lecithin cholesterol acyltransferase (25), whereas others are synthesized from PC
hydrolysis via the secretory phospholipase A2 family (26). Although LPCs are typically regarded
as pro-inflammatory mediators, they were increased in inflammatory
diseases (27,28). A previous series of studies have
demonstrated a protective effect of LPCs against diseases,
including type 2 diabetes (29),
cancer (30) and inflammatory
diseases (31).
In the results of the present study, LPCs that
contain polyunsaturated fatty acids were decreased in septic mice,
whereas saturated fatty acid LPCs demonstrated no significant
difference between septic and sham mice. Furthermore, unsaturated
LPCs combine with the albumin in blood plasma, causing
polyunsaturated fatty acids to move into organs where fatty acids
are rich, such as in the brain. Therefore, unsaturated LPCs
primarily associate with albumin instead of lipoproteins (32). Furthermore, a number of studies
indicated that the rate of albumin degradation and albumin loss in
tissue increased in patients with critical illnesses, such as
sepsis (33–35). In addition, the concentration of
albumin would return to normal values (32) as soon as the infection was
controlled. Compared with control septic mice, the level of
unsaturated LPCs obtained from the septic mice with LGG
pretreatment in the present study was much closer to those in sham
mice, indicating that LGG may reverse the progress of sepsis by
elevating albumin in the blood plasma. In addition, one of the
possible explanations for a non-significant difference existing in
saturated LPCs between septic and sham mice may be the different
role that different LPC subtypes serve during sepsis.
In conclusion, the potential impact of live
probiotic therapy on the improvement of the survival rate in
experimental septic mice was demonstrated in the present study.
This improvement was associated with reduced ileum mucosal damage,
increased gut barrier integrity and altered global serum metabolic
profiles.
References
|
1
|
Bouza C, López-Cuadrado T, Saz-Parkinson Z
and Amate-Blanco JM: Epidemiology and recent trends of severe
sepsis in Spain: A nationwide population-based analysis
(2006-2011). BMC Infect Dis. 14:38632014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khailova L, Frank DN, Dominguez JA and
Wischmeyer PE: Probiotic administration reduces mortality and
improves intestinal epithelial homeostasis in experimental sepsis.
Anesthesiology. 119:166–177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sokol H: Probiotics and Antibiotics in
IBD. Dig Dis. 32(Suppl 1): S10–S17. 2014. View Article : Google Scholar
|
|
4
|
Wan Y, Xin Y, Zhang C, Wu D, Ding D, Tang
L, Owusu L, Bai J and Li W: Fermentation supernatants of
Lactobacillus delbrueckii inhibit growth of human colon cancer
cells and induce apoptosis through a caspase 3-dependent pathway.
Oncol Lett. 7:1738–1742. 2014.PubMed/NCBI
|
|
5
|
Plaza-Diaz J, Gomez-Llorente C, Fontana L
and Gil A: Modulation of immunity and inflammatory gene expression
in the gut, in inflammatory diseases of the gut and in the liver by
probiotics. World J Gastroenterol. 20:15632–15649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dietrich CG, Kottmann T and Alavi M:
Commercially available probiotic drinks containing Lactobacillus
casei DN-114001 reduce antibiotic-associated diarrhea. World J
Gastroenterol. 20:15837–15844. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu CT, Chen PJ, Lee YT, Ko JL and Lue KH:
Effects of immunomodulatory supplementation with Lactobacillus
rhamnosus on airway inflammation in a mouse asthma model. J
Microbiol Immunol Infect. Nov 11–2014.(Epub ahead of print).
View Article : Google Scholar
|
|
8
|
Griffin JL and Bollard ME: Metabonomics:
Its potential as a tool in toxicology for safety assessment and
data integration. Curr Drug Metab. 5:389–398. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lahti L, Salonen A, Kekkonen RA, Salojärvi
J, Jalanka-Tuovinen J, Palva A, Orešič M and de Vos WM:
Associations between the human intestinal microbiota, Lactobacillus
rhamnosus GG and serum lipids indicated by integrated analysis of
high-throughput profiling data. Peer J. 1:e322013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan F and Polk DB: Probiotics: Progress
toward novel therapies for intestinal diseases. Curr Opin
Gastroenterol. 26:95–101. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Quigley EM: Gut Bacteria in health and
disease. Gastroenterol Hepatol (N Y). 9:560–569. 2013.PubMed/NCBI
|
|
13
|
Barraud D, Blard C, Hein F, Marçon O,
Cravoisy A, Nace L, Alla F, Bollaert PE and Gibot S: Probiotics in
the critically ill patient: A double blind, randomized,
placebo-controlled trial. Intensive Care Med. 36:1540–1547. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Honeycutt TC, El Khashab M, Wardrop RM
III, McNeal-Trice K, Honeycutt AL, Christy CG, Mistry K, Harris BD,
Meliones JN and Kocis KC: Probiotic administration and the
incidence of nosocomial infection in pediatric intensive care: A
randomized placebo-controlled trial. Pediatr Crit Care Med.
8:452–458. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gou S, Yang Z, Liu T, Wu H and Wang C: Use
of probiotics in the treatment of severe acute pancreatitis: A
systematic review and meta-analysis of randomized controlled
trials. Crit Care. 18:R572014. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barraud D, Bollaert PE and Gibot S: Impact
of the administration of probiotics on mortality in critically ill
adult patients: A meta-analysis of randomized controlled trials.
Chest. 143:646–655. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Treede I, Braun A, Sparla R, Kühnel M,
Giese T, Turner JR, Anes E, Kulaksiz H, Füllekrug J, Stremmel W, et
al: Anti-inflammatory effects of phosphatidylcholine. J Biol Chem.
282:27155–27164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stremmel W, Merle U, Zahn A, Autschbach F,
Hinz U and Ehehalt R: Retarded release phosphatidylcholine benefits
patients with chronic active ulcerative colitis. Gut. 54:966–971.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stremmel W: Mucosal protection by
phosphatidylcholine as new therapeutic concept in ulcerative
colitis. Z Gastroenterol. 51:384–389. 2013.(In German). PubMed/NCBI
|
|
20
|
Zhang H, Kovacs-Nolan J, Kodera T, Eto Y
and Mine Y: γ-Glutamyl cysteine and γ-glutamyl valine inhibit TNF-α
signaling in intestinal epithelial cells and reduce inflammation in
a mouse model of colitis via allosteric activation of the
calcium-sensing receptor. Biochim Biophys Acta. 1852:792–804. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsieh CY, Osaka T, Moriyama E, Date Y,
Kikuchi J and Tsuneda S: Strengthening of the intestinal epithelial
tight junction by Bifidobacterium bifidum. Physiol Rep. 3:pii.
e123272015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barrenetxe J, Sánchez O, Barber A, Gascón
S, Rodríguez-Yoldi MJ and Lostao MP: TNFα regulates sugar
transporters in the human intestinal epithelial cell line Caco-2.
Cytokine. 64:181–187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Goretsky T, Dirisina R, Sinh P, Mittal N,
Managlia E, Williams DB, Posca D, Ryu H, Katzman RB and Barrett TA:
P53 mediates TNF-induced epithelial cell apoptosis in IBD. Am J
Pathol. 181:1306–1315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao X, Qu H, Ai CZ, Cao YF, Huang T, Chen
JX, Zeng J, Sun XY, Hong M, Gonzalez FJ, et al: Regulation profile
of phosphatidylcholines (PCs) and lysophosphatidylcholines (LPCs)
components towards UDP-glucuronosyltransferases (UGTs) isoforms.
Xenobiotica. 45:197–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu M, Subramanian VS and Subbaiah PV:
Modulation of the positional specificity of lecithin-cholesterol
acyltransferase by the acyl group composition of its
phosphatidylcholine substrate: Role of the sn-1-acyl group.
Biochemistry. 37:13626–13633. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miklishanskaia SV, Liakishev AA and
Kukharchuk VV: Clinical role of lipoprotein-associated
phospholipase A2. Kardiologiia. 53:59–70. 2013.(In Russian).
PubMed/NCBI
|
|
27
|
Domeij H, Hua X, Su J, Bäcklund A, Yan Z,
Frostegård AG, Haeggström JZ, Modéer T and Frostegård J: Annexin A5
inhibits atherogenic and pro-inflammatory effects of
lysophosphatidylcholine. Prostaglandins Other Lipid Mediat.
106:72–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kajander K, Myllyluoma E, Kyrönpalo S,
Rasmussen M, Sipponen P, Mattila I, Seppänen-Laakso T, Vapaatalo H,
Oresic M and Korpela R: Elevated pro-inflammatory and lipotoxic
mucosal lipids characterise irritable bowel syndrome. World J
Gastroenterol. 15:6068–6074. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang-Sattler R, Yu Z, Herder C, Messias
AC, Floegel A, He Y, Heim K, Campillos M, Holzapfel C, Thorand B,
et al: Novel biomarkers for pre-diabetes identified by
metabolomics. Mol Syst Biol. 8:6152012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jantscheff P, Schlesinger M, Fritzsche J,
Taylor LA, Graeser R, Kirfel G, Fürst DO, Massing U and Bendas G:
Lysophosphatidylcholine pretreatment reduces VLA-4 and
P-Selectin-mediated b16.f10 melanoma cell adhesion in vitro and
inhibits metastasis-like lung invasion in vivo. Mol Cancer Ther.
10:186–197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smani Y, Domínguez-Herrera J,
Ibáñez-Martínez J and Pachón J: Therapeutic efficacy of
lysophosphatidylcholine in severe infections caused by
Acinetobacter baumannii. Antimicrob Agent Chemother.
59:3920–3924. 2015. View Article : Google Scholar
|
|
32
|
Croset M, Brossard N, Polette A and
Lagarde M: Characterization of plasma unsaturated
lysophosphatidylcholines in human and rat. Biochem J. 345:61–67.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou B, Ren J, Han G, Chen YAJ, Gu G, Chen
J, Wang G and Li J: Dynamics of albumin synthetic response to
intra-abdominal abscess in patients with gastrointestinal fistula.
Surg Infect (Larchmt). 15:111–117. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ruot B, Papet I, Bechereau F, Denis P,
Buffiere C, Gimonet J, Glomot F, Elyousfi M, Breuille D and Obled
C: Increased albumin plasma efflux contributes to hypoalbuminemia
only during early phase of sepsis in rats. Am J Physiol Regul
Integr Comp Physiol. 284:R707–R713. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wiedermann CJ, Wiedermann W and Joannidis
M: Hypoalbuminemia and acute kidney injury: A meta-analysis of
observational clinical studies. Intensive Care Med. 36:1657–1665.
2010. View Article : Google Scholar : PubMed/NCBI
|