Introduction
Ulcerative colitis (UC), which is a subtype of
inflammatory bowel disease (IBD) (1), is a chronic and debilitating condition
that results in serious intestinal injuries. UC typically occurs as
a result of inflammatory dysfunction (2). Patients with UC often exhibit
intestinal barrier dysfunction, as well as microbiota and bacterial
dysbiosis (3).
Although UC is common, its etiology remains poorly
understood (4). In a previous study,
epithelial barrier impairment was demonstrated to be associated
with low-grade inflammation and dysbiosis as potential causative
factors, and are associated with the severity of UC (2). Furthermore, in patients with UC, an
increase in gut permeability has previously been associated with
the altered expression levels or distribution of tight junction
proteins, including occludin and zonula occludens-1 (ZO-1)
(5). Therefore, increased intestinal
permeability and the occurrence of dysbiosis may be the cause of
UC-symptoms (6). The evidence that
gut microbiota may have a role in the pathophysiology of UC
provides a rationale for probiotic use, which has exhibited
beneficial effects (7). However, the
therapeutic mechanism of action for the effect of probiotics in UC
has yet to be elucidated.
Probiotics are defined as live organisms that exert
a health benefit on the host through diverse mechanisms.
Bacillus subtilis is a type of probiotic tolerated by humans
and animals (8). B. subtilis
is hypothesized to affect the composition or function of the
commensal, bacterial and host epithelia. Furthermore, it also
influences immunological responses and restricts bacterial and
lipopolysaccharide (LPS) translocation, and decreases visceral
sensitivity (8). In recent clinical
trials, probiotics have been widely used to treat disorders of the
intestine (7). As enhancement of the
intestinal barrier has been associated with the repair of mucosal
injuries, the role of B. subtilis treatment in maintaining
gut barrier integrity was investigated in the present study, due to
its potential usage in the alleviation of UC mucosal injuries.
Changes in the gut microbiota have been associated
with IBD, including alterations in the relative abundance of
bacteria that are both beneficial and detrimental to gut health,
and a decrease in the diversity of the microbiota (9,10).
Although previous reports have demonstrated the protective effect
of B. subtilis in gut protection (11,12), the
impact of B. subtilis administration on gut microbiota
alteration remains unknown. The present study aimed to elucidate
the role of B. subtilis in the restoration of mucosa,
determine its effective dose, and provide preclinical data for
B. subtilis usage in UC therapy.
Materials and methods
Modeling of colorectal colitis in mice
and treatment
Male C57 mice (body weight, 23±1 g; 6 weeks old)
were obtained from the Animal Center, Nanjing Drum Tower Hospital
(Nanjing, China) and the in vivo experiment was performed in
the same facility. Mice were maintained under controlled conditions
(25°C, 55% humidity, 12 h light/dark cycle) and fed standard
laboratory food. Mice were administered 3% (wt/vol) dextrose
sulfate sodium (DSS) (molecular weight, 35,000–44,000; MP
Biomedicals, Inc., Aurora, OH, USA) via drinking water for seven
days. Additionally, mice were treated daily with different reagents
via gavage (catheter diameter, 1.2 mm), including normal saline
(NS; n=8) or B. subtilis (R179; Beijing Hanmi Pharm Co.,
Ltd., Beijing, China) at a high (1×109 CFU/mouse/day;
n=8) or low (1×108 CFU/mouse/day; n=8) dosage until the
end of the study. On day eight, mice were weighed and then
sacrificed via ether exposure (200 mg/l; Shanghai National Medicine
Group, Shanghai, China) in an airtight container in a biosafety
cabinet. Colons were harvested, measured and fixed in 4% formalin
for subsequent histological examination. Animal experiments were
approved by the Ethics Committee of Medical Research, Huashan
Hospital of Fudan University (Shanghai, China).
Assessment of colitis
Following the initiation of DSS treatment, daily
changes in body weight and clinical signs of colitis, such as
rectal bleeding, diarrhea and piloerection, were examined. The
disease activity index consisted of scoring for rectal bleeding
(0–4), as previously reported (13).
Hemoccult SENSA (Beckman Coulter, Inc., Brea, CA, USA) was used to
examine rectal bleeding.
Periodic acid-Schiff/alcian blue
staining
Alcian blue staining was performed according to a
previous report (14). Tissue
sections (6 µm thick) were immersed in 100% ethanol for 10 min,
rinsed in water for 10 min, immersed in 3% acetic acid for 2 min
and subsequently stained in 1% alcian blue 8GX in 3% acetic acid
(pH 2.5) for 2.5 h. To remove non-specific staining, 3% acetic acid
and water was used to rinse the sections for 10 min. Slides were
subsequently oxidized in 1% periodic acid in water at room
temperature for 10 min, washed in water for 5 min, immersed in
Schiff's reagent for 10 min, rinsed in water for 5 min and three
times in 0.5% sodium metabisulphite prior to a final wash in water.
To reveal O-acetylated oligosaccharides, sections were treated with
0.1 M KOH for 30 min and 1 mM periodic acid prior to the Schiff
reagent.
Immunofluorescence
Frozen tissue sections (6 µm thick) were
immunostained with 1:100 primary antibodies against ZO-1
(clonality, H-300; cat. no. sc-10804) and claudin (clonality, D-4;
cat. no. sc-137121; species: mouse, rat, human, equine, canine,
bovine, porcine) (both Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA). Images were analyzed using a BIOREVO immunofluorescence
microscope (Keyence Corp., Osaka, Japan). Each result was obtained
from at least three separate experiments. Six mice per group were
prepared for each experiment.
Measurement of intestinal
permeability
Intestinal permeability was determined according to
a previously described method (15).
DSS-treated mice with high/low-dose B. subtilis or control
saline (n=5; 4 days) were fasted for 12 h prior to oral gavage of
disaccharide permeability probes [100 mg/ml lactulose and 50 mg/ml
mannitol (both Sigma-Aldrich; Merck Millipore, Darmstradt, Germany)
dissolved in 2 ml water] and urine was collected 12 h later. Urine
volume was measured and the concentrations of lactulose and
mannitol were determined by high-performance liquid chromatography
with an NH2 column (Bischoff Chromatohraphy, Leonberg, Germany) and
acetonitrile (70%) based elution. The ratio of the amount of probe
in urine to the amount administered as lactulose or mannitol
recovery rate was calculated accurately. Intestinal permeability
was evaluated as a ratio of lactulose recovery rate to mannitol
recovery rate.
Measurement of serum cytokines and
endotoxin
On the 4th day following DSS treatment, mice were
anesthetized with ether (200 mg/l; Shanghai National Medicine
Group) in an airtight container within a biosafety cabinet and
blood was collected from the retrobulbar venous plexus using
pyrogen-free heparinized syringes. Cytokine [interleukin (IL)-10,
IL-12 p70, IL-17A, and IL-23] levels were analyzed by ELISA
according to the manufacturer's protocol (R&D Systems, Inc.,
Minneapolis, MN, USA). Plasma endotoxin was measured using a
Limulus amebocyte lysate pyrogen test kit (Xiamen Houshiji, Ltd.,
Xiamen, China; cat. no. KC48).
Short-chain fatty acid (SCFA)
assay
Fresh mice fecal samples were collected from the
cages, weighed and stored at −80°C. Fecal samples were mixed with
distilled water and centrifuged (2,500 × g). The supernatant was
removed, filtered and mixed with ether and sulfuric acid. Following
centrifugation (2,500 × g), the ether layer was collected and
measured in an Agilent 6890N gas chromatograph machine (Agilent
Technologies, Inc., Santa Clara, CA, USA) to determine the total
SCFA concentrations.
Microbiological analysis of mice fecal
samples
Microbiota composition was assessed by 454
pyrosequencing (GS FLX TI technology, Genoscreen, Lille, France)
targeting the V3-V4 region of the bacterial 16S rRNA gene (V3,
forward 5′-TACGGRAGGCAGCAG-3′ and V4 reverse
5′-GGACTACCAGGGTATCTAAT-3′). Sequences were binned for a minimal
sequence length of 300 pb, a minimal base quality threshold of 30
cycles and a maximum homopolymer length of 6 cycles. Resulting
sequences were assigned to different taxonomic levels, from phylum
to genus using the Ribosomal Database Project (16). Sequences were further clustered into
operational taxonomic units (OTUs) or phylotypes at 97% of identity
using the Quantitative Insights into Microbial Ecology pipeline and
CD-HIT (17,18). OTUs were assigned to their closest
taxonomic neighbors and relative bacterial species using Seqmatch
(Michigan State University, East Lansing, MI, USA) and Blastall
(National Centre for Biotechnology Information, Bethesda, MD,
USA).
Statistical analysis
Data were expressed as the mean ± standard error of
the mean. Differences were analyzed using Student's t-test,
Chi-square test, or one-way analysis of variance with Tukey's
post-hic test for multiple group comparison. P<0.05 was
considered to indicate a statistically significant difference.
Results
B. subtilis alleviates DSS-induced
lethality and intestinal injuries in mice
To examine the role of B. subtilis in the
amelioration of UC in vivo, mice were initially exposed to a
lethal dose of 4% DSS (20 ml/d), a pharmacological agent used to
induce UC that also causes severe secondary symptoms. Mice were
subsequently treated orally with either a high- or low-dose of
B. subtilis preparation, or phosphate-buffered saline for
control at eight days post-administration. The results indicated
that the high dose of B. subtilis solution protected mice
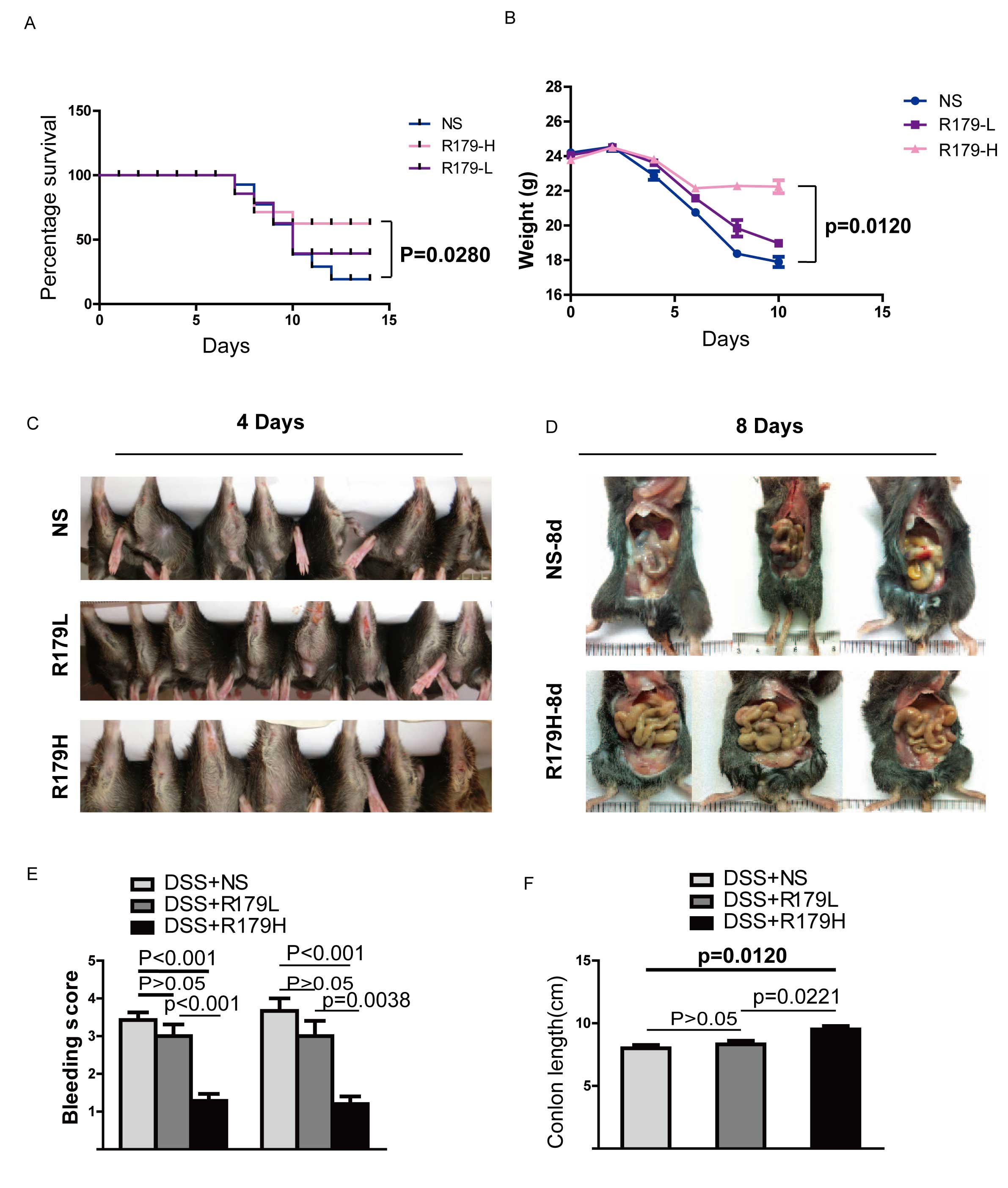
from the lethal effect of DSS-induced UC (Fig. 1A). At the end of the study, ~50%
survival of probiotic-treated mice was observed. In contrast, only
~1/3 of mice who received normal saline survived for the same
period as the high-dose probiotic-treated group and this difference
was significant (P=0.0280; Fig. 1A).
In addition, a high-dose of B. subtilis administration
significantly ameliorated weight loss compared with the normal
saline group (P=0.0120; Fig. 1B). As
DSS can promote intestinal damage, such as hematochezia and
intestinal bleeding, the anuses and intestinal tracts of the three
groups were examined. For mice in the high-dose B.
subtilis-treated group, the anus and intestinal tract exhibited
reduced bleeding and anabrosis than the control and low dose groups
(Fig. 1C and D). Mice in the control
and low-dose groups suffered more severe colon necrosis and shorter
colons compared with the high-dose B. subtilis-treated group
(Fig. 1E and F). These results
indicate that high-dose B. subtilis administration
alleviates DDS-induced colon damage and that B. subtilis
induces a dose-dependent effect.
B. subtilis protects against
DSS-induced intestinal mucosal damage in mice
DSS induces UC by causing serious intestinal mucosal
damage (19). In accordance with the
findings mentioned, low or high doses of B. subtilis may
attenuate the symptoms of DSS-induced UC. In light of this, it was
hypothesized that B. subtilis may produce its effect by
protecting the intestinal mucosa from damage and by reinforcing its
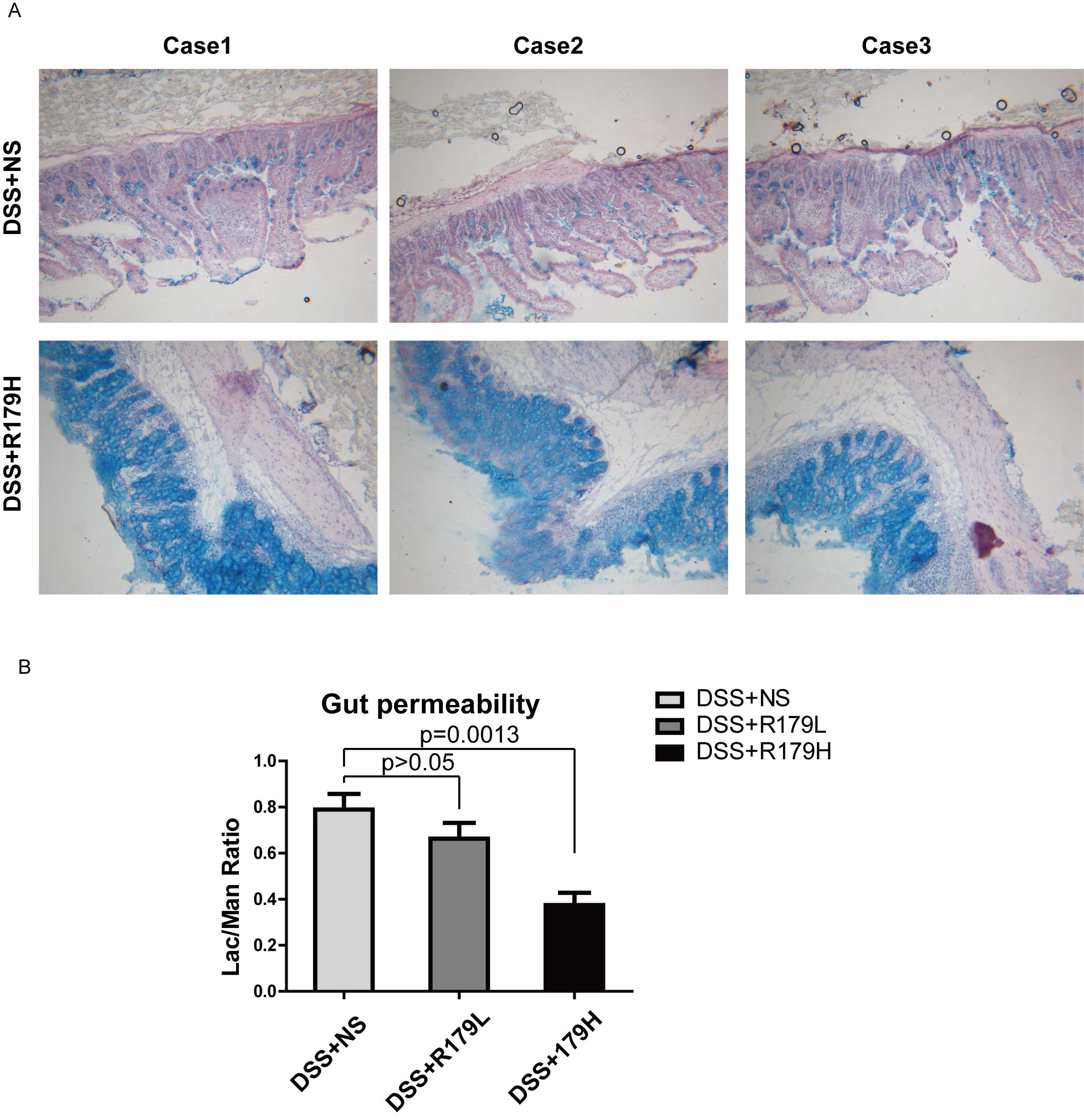
repair. To test this hypothesis, alcian blue staining was conducted
to determine how the intestinal mucosa reacted to the
administration of B. subtilis following DSS treatment. As
shown in Fig. 2A, mice treated with
high-dose B. subtilis exhibited increased expression levels
of mucins compared with the control group, which indicated repair
of the colon mucosa. These results suggest that the high dose of
the B. subtilis probiotic promoted the restoration of
intestinal mucosa.
To further test this hypothesis, intestinal
permeability was measured, as DSS-induced intestinal mucosa damage
may lead to an increase in intestinal permeability. The results of
the present study demonstrated that intestinal permeability was
recovered in the B. subtilis-treated group (Fig. 2B). This further supports the
hypothesis that treatment with B. subtilis may protect
intestinal mucosa from DSS-induced damage and attenuate the
inflammatory reaction.
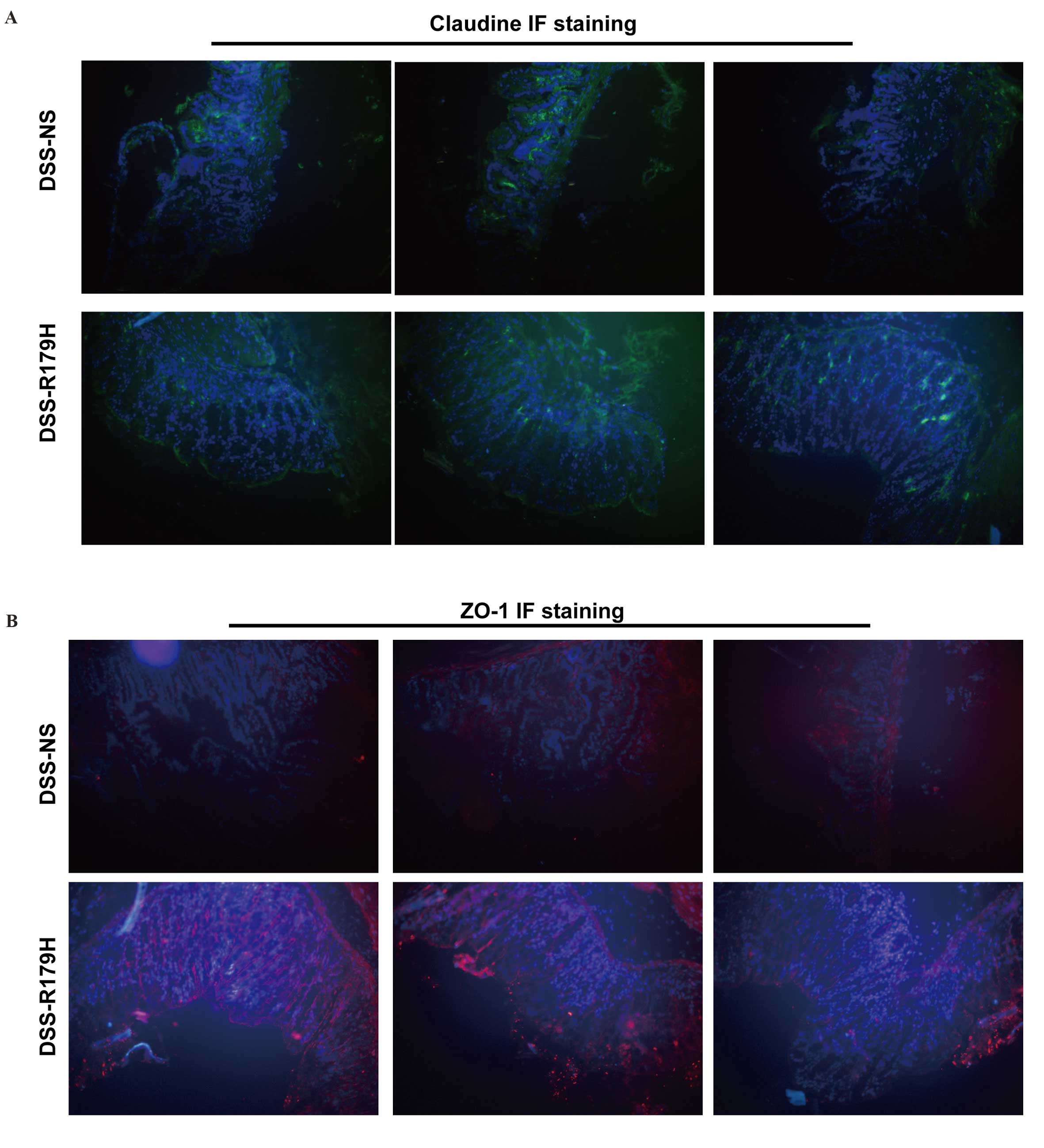
B. subtilis helps to restore tight
junctions
The tight junction complex of the intestinal mucosa
is considered to be the first ‘firewall’ for gut immunity (20). ZO-1 and claudins are two large
families of the tight junction complex. To further explore whether
B. subtilis was able to repair DSS-induced damage to the
tight junctions, the intestines of mice treated with or without
B. subtilis were harvested and probed with ZO-1 and
claudins. The samples were then observed under a confocal laser
scanning microscope, and the results demonstrated that the two
tight junction-associated markers increased following high-dose
B. subtilis treatment (Fig. 3A
and B). This indicated that B. subtilis was involved in
the repair process of DSS-induced mucosal damage and restored the
mucosal tight junction complex.
B. subtilis administration alleviates
systemic inflammation upon DSS treatment
Restoration of intestinal permeability may be
related to the relief of the inflammation reaction (21). Therefore, the effect of B.
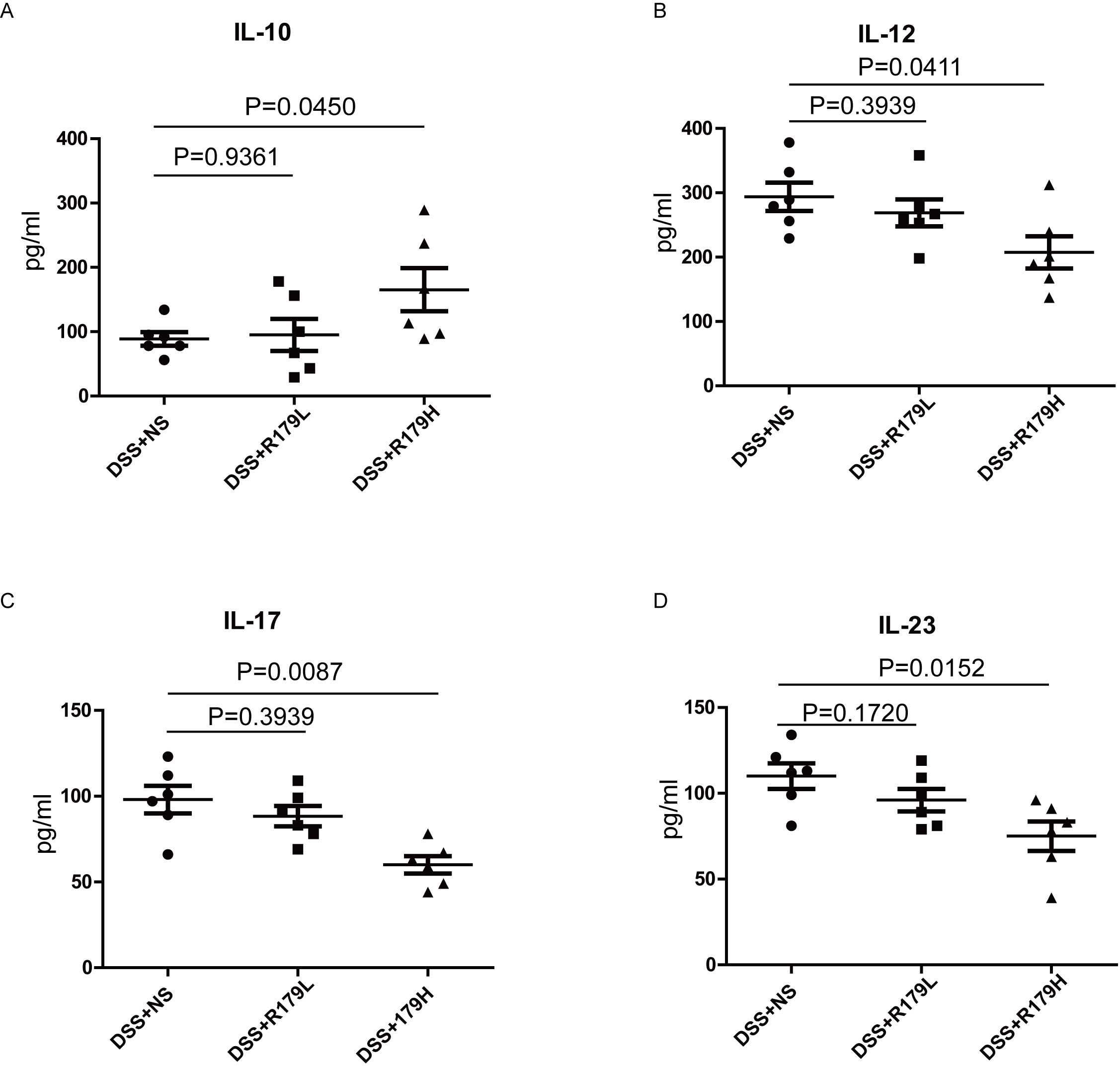
subtilis administration on gut inflammation was explored using
ELISA. An increase in plasma cytokines has previously been
reported, including IL-12, IL-17 and IL-23, whereas IL-10 decreased
in IBD (22). In the present study,
the mean plasma levels of IL-12, IL-17 and IL-23 in the high-dose
B. subtilis group were significantly reduced (P=0.0411,
0.0087 and 0.0152, respectively) and the mean IL-10 plasma levels
were significantly increased (P=0.0450) (Fig. 4) compared with the NS group. However,
low-dose B. subtilis treatment produced a less-marked
effect.
B. subtilis administration balances
anti-and pro-inflammatory factors in the gut of mice
Gut microbiota is a primary source of LPS endotoxin,
which is a damage-associated pathogen that promotes gut
inflammatory reactions and systemic inflammation (23,24).
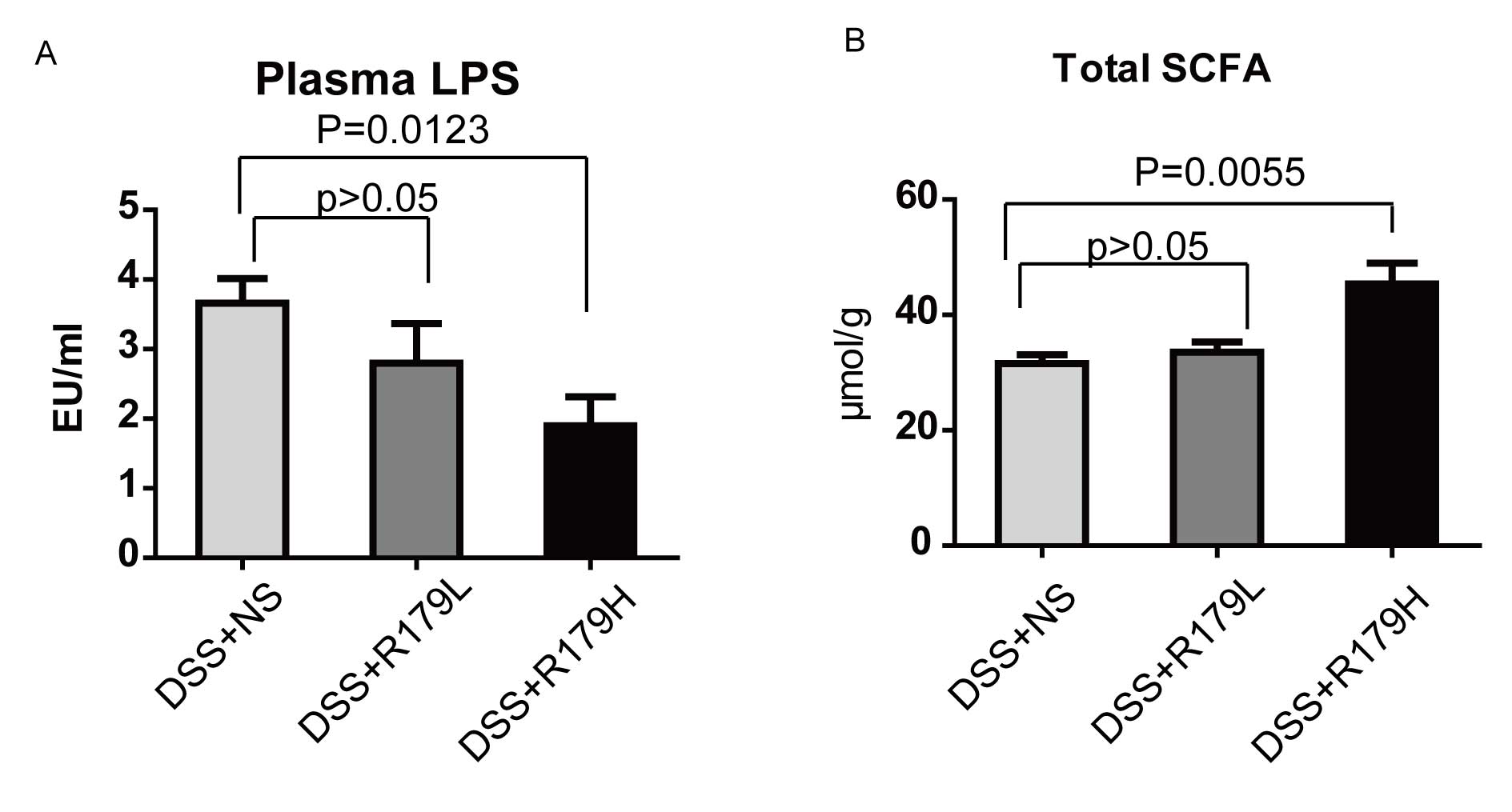
Plasma LPS of mice was tested and a difference in LPS was detected
between the high-and low-dose B. subtilis-treated groups and
the control group. These results indicated a significant decrease
of LPS concentration in the probiotic group with the administration
of high-dose B. subtilis compared with the control group
(P<0.001; Fig. 5A).
SCFAs have anti-inflammatory functions via
interaction with G protein-coupled receptor 43 and have been
demonstrated to induce pro-inflammatory cytokines in various models
of colitis (25,26). The concentration of total SCFAs was
significantly higher in the high-dose B. subtilis group
compared with the control group (P=0.0055; Fig. 5B), suggesting that high-dose B.
subtilis was beneficial in maintaining SCFA content, which in
turn reduced gut inflammation.
B. subtilis administration ameliorates
DSS-induced dysbiosis in the gut of mice
There are 100 trillion microorganisms housed in the
human body. These microorganisms are maintained as commensals on
the gut mucosa, and are associated with metabolism, including
maintaining the internal environment and regulating the immune
system (27). Consequently, 16s-rDNA
sequencing analysis was performed in the present study to examine
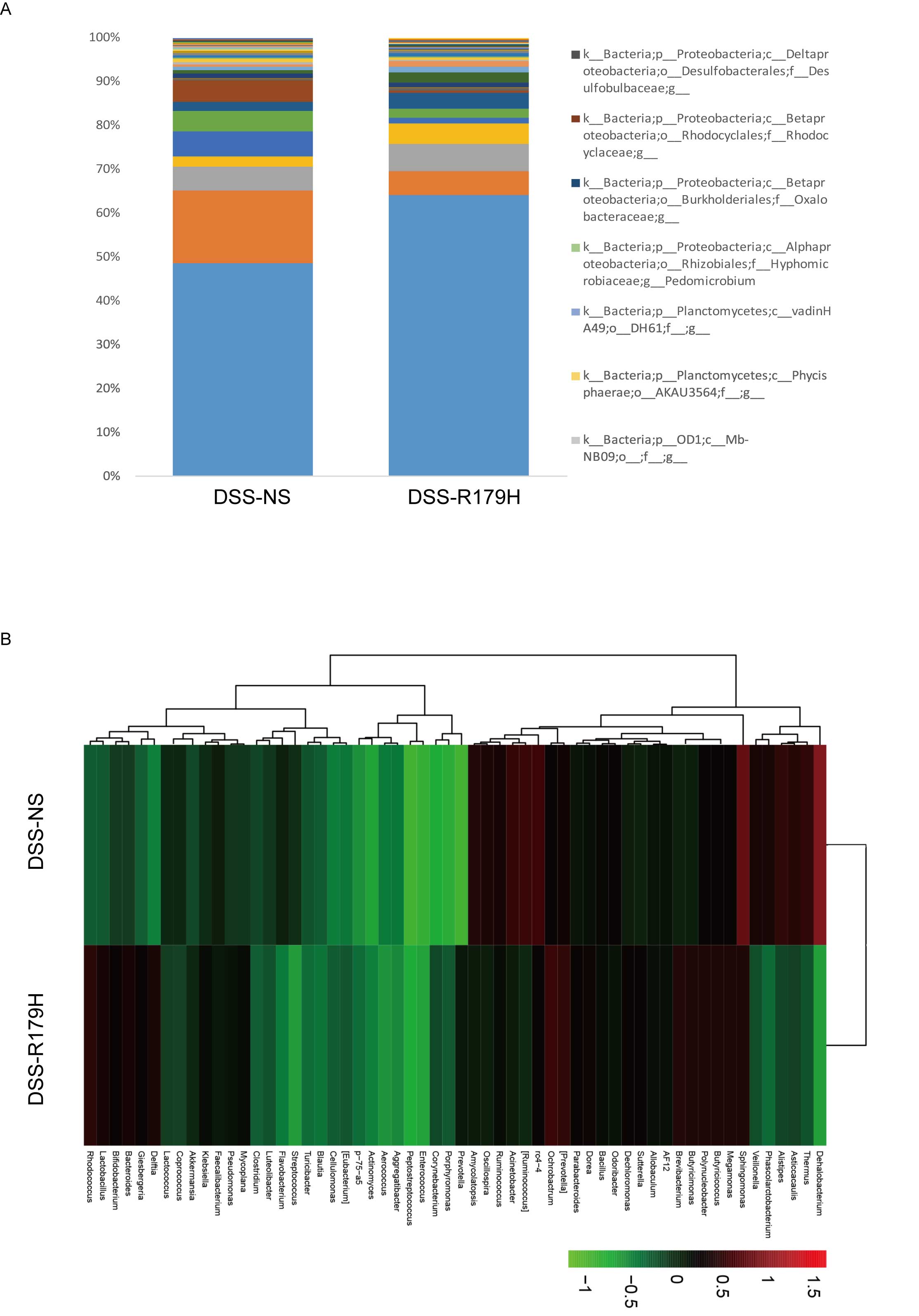
changes in the microbiota. As demonstrated in Fig. 6, it was observed that the gut
microbiota was markedly altered in the high-dose B.
subtilis-treated group compared with the control group.
Specifically, a reduction of Acinetobacter sp.,
Ruminococcus sp., Clostridium spp. and
Veillonella sp. was detected upon high-dose B.
subtilis treatment, whereas levels of Bifidobacterium
sp., Lactobacillus sp., and Butyricicoccus sp. were
increased. Acinetobacter spp., Ruminococcus spp.
Clostridium sp. and Veillonella sp. have been shown
to be overrepresented in IBD patients, whereas
Bifidobacterium spp., Lactobacillus spp., and
Butyricicoccus sp. are decreased (28–32).
These results demonstrated the role of B. subtilis in the
amelioration of DSS-induced dysbiosis in the gut of model mice.
Discussion
In the present study, DSS was used to induce UC in a
mouse model, and treatment with B. subtilis was revealed to
markedly decrease the mortality of DSS-treated mice and protect the
intestine from further damage. In addition, B. subtilis
treatment decreased the damage caused by DSS, which supports the
hypothesis that B. subtilis is able to repair epithelial
cell injury in intestinal inflammation via immunomodulation
(9). Elevated levels of IL-12, IL-17
and IL-23 have previously been found in the epithelial mucosal
barrier of subjects with IBD, whereas IL-10 is known to have a
protective role in alleviating gut inflammation (9,33,34).
Additionally, the present study detected elevated levels of IL-10
and decreased levels of IL-12, IL-17 and IL-23 in the high-dose,
but not low-dose, B. subtilis-treatment groups. Therefore,
the present study provides evidence that B. subtilis
regulates gut immune balance in a dose-dependent manner.
A reduction in the number of SCFA-producing bacteria
can result in a degree of focal metabolic stress and vulnerability
to inflammatory disease (35). Using
gas chromatography, it was determined that B. subtilis
administration increased the levels of SCFAs. In addition, a
significant increase of the Butyricicoccus spp., which
contributes to butyrate generation (36), was detected upon B. subtilis
treatment in the present study. Previous studies have shown that
the IBD phenotype was associated with lower levels of the
clostridial cluster IV genus Butyricicoccus (36,37).
These findings indicate that B. subtilis may be beneficial
for the survival and expansion of Butyricicoccus spp. under
the conditions of gut damage.
A balance of healthy gut commensal bacteria is
required for the suppression of pathogenic infections (38), with increasing evidence suggesting
that the restoration of normal commensals via transplant is more
effective at fighting Clostridium sp. infection than
antibiotics (39). Commercially
available probiotics, including Lactobacillus and
Bifidobacterium spp., are used to attenuate inflammatory
activity and prevent relapses in UC (40). In the present study, it was
determined that beneficial Bifidobacterium,
Lactobacillus, and Butyricicoccus spp. increased in
the high-dose B. subtilis-treated groups, compared with the
control group. Species known to promote gut damage, such as
Acinetobacter sp., Ruminococcus sp.,
Clostridium spp. and Veillonella sp, were found to be
decreased following B. subtilis treatment in the present
study. These results indicate the potential role of B.
subtilis administration in restoring a healthy balance of
beneficial and harmful bacteria in the gut.
In conclusion, dose-dependent B. subtilis
administration was demonstrated to aid intestinal mucosa recovery
from DSS-induced damage and protect the intestinal mucosa by
balancing beneficial and harmful bacterium and their respective,
associated anti- and pro-inflammatory agents. The present study
elucidated the mechanisms of B. subtilis action and provided
preclinical data for B. subtilis use in UC therapy.
Acknowledgements
This study was supported by grants from the Basic
Research Program of Beijing Renze Foundation (grant no.
ZX-IBD-13002). The authors would like to thank Dong-Ping Hu,
Dan-Dan Huang, Shan-Hua Tang, Lin-Na Guo and Dan Cao, Technicians
from the International Cooperation Laboratory on Signal
Transduction (Eastern Hepatobiliary Surgery Hospital, Second
Military Medical University, Shanghai, China), for their technical
assistance.
References
|
1
|
Mosli MH, Feagan BG, Sandborn WJ, D'Haens
G, Behling C, Kaplan K, Driman DK, Shackelton LM, Baker KA,
Macdonald JK, et al: Histologic evaluation of ulcerative colitis: A
systematic review of disease activity indices. Inflamm Bowel Dis.
20:564–575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rosenberg L, Nanda KS, Zenlea T, Gifford
A, Lawlor GO, Falchuk KR, Wolf JL, Cheifetz AS, Goldsmith JD and
Moss AC: Histologic markers of inflammation in patients with
ulcerative colitis in clinical remission. Clin Gastroenterol
Hepatol. 11:991–996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Manichanh C, Borruel N, Casellas F and
Guarner F: The gut microbiota in IBD. Nat Rev Gastroenterol
Hepatol. 9:599–608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Conrad K, Roggenbuck D and Laass MW:
Diagnosis and classification of ulcerative colitis. Autoimmun Rev.
13:463–466. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suzuki T: Regulation of intestinal
epithelial permeability by tight junctions. Cell Mol Life Sci.
70:631–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Buning C, Geissler N, Prager M, Sturm A,
Baumgart DC, Büttner J, Bühner S, Haas V and Lochs H: Increased
small intestinal permeability in ulcerative colitis: Rather genetic
than environmental and a risk factor for extensive disease? Inflamm
Bowel Dis. 18:1932–1939. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shen J, Zuo ZX and Mao AP: Effect of
probiotics on inducing remission and maintaining therapy in
ulcerative colitis, Crohn's disease, and pouchitis: Meta-analysis
of randomized controlled trials. Inflamm Bowel Dis. 20:21–35. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aly SM, Ahmed Y Abdel-Galil, Abdel-Aziz
Ghareeb A and Mohamed MF: Studies on Bacillus subtilis and
Lactobacillus acidophilus, as potential probiotics, on the immune
response and resistance of Tilapia nilotica (Oreochromis niloticus)
to challenge infections. Fish Shellfish Immunol. 25:128–136. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sarra M, Pallone F, Macdonald TT and
Monteleone G: IL-23/IL-17 axis in IBD. Inflamm Bowel Dis.
16:1808–1813. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nell S, Suerbaum S and Josenhans C: The
impact of the microbiota on the pathogenesis of IBD: Lessons from
mouse infection models. Nat Rev Microbiol. 8:564–577. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Okamoto K, Fujiya M, Nata T, Ueno N, Inaba
Y, Ishikawa C, Ito T, Moriichi K, Tanabe H, Mizukami Y, et al:
Competence and sporulation factor derived from Bacillus subtilis
improves epithelial cell injury in intestinal inflammation via
immunomodulation and cytoprotection. Int J Colorectal Dis.
27:1039–1046. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Selvam R, Maheswari P, Kavitha P,
Ravichandran M, Sas B and Ramchand CN: Effect of Bacillus subtilis
PB6, a natural probiotic on colon mucosal inflammation and plasma
cytokines levels in inflammatory bowel disease. Indian J Biochem
Biophys. 46:79–85. 2009.PubMed/NCBI
|
|
13
|
Axelsson LG, Landström E, Goldschmidt TJ,
Grönberg A and Bylund-Fellenius AC: Dextran sulfate sodium (DSS)
induced experimental colitis in immunodeficient mice: Effects in
CD4(+)-cell depleted, athymic and NK-cell depleted SCID mice.
Inflamm Res. 45:181–191. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lindén SK, Florin TH and McGuckin MA:
Mucin dynamics in intestinal bacterial infection. PLoS One.
3:e39522008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Olson TS, Reuter BK, Scott KG, Morris MA,
Wang XM, Hancock LN, Burcin TL, Cohn SM, Ernst PB, Cominelli F, et
al: The primary defect in experimental ileitis originates from a
nonhematopoietic source. J Exp Med. 203:541–552. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cole JR, Wang Q, Cardenas E, Fish J, Chai
B, Farris RJ, Kulam-Syed-Mohideen AS, McGarrell DM, Marsh T,
Garrity GM and Tiedje JM: The ribosomal database project: Improved
alignments and new tools for rRNA analysis. Nucleic Acids Res.
37:(Database Issue). D141–D145. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li W and Godzik A: Cd-hit: A fast program
for clustering and comparing large sets of protein or nucleotide
sequences. Bioinformatics. 22:1658–1659. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Caporaso JG, Kuczynski J, Stombaugh J,
Bittinger K, Bushman FD, Costello EK, Fierer N, Peñna AG, Goodrich
JK, Gordon JI, et al: QIIME allows analysis of high-throughput
community sequencing data. Nat Methods. 7:335–336. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chassaing B, Aitken JD, Malleshappa M and
Vijay-Kumar M: Dextran sulfate sodium (DSS)-induced colitis in
mice. Curr Protoc Immunol. 104:Unit 15-25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Goto Y, Kurashima Y and Kiyono H: Roles of
the gut mucosal immune system in symbiosis and immunity. Rinsho
Ketsueki. 56:2205–2212. 2015.(In Japanese). PubMed/NCBI
|
|
21
|
Genser L, Poitou C, Brot-Laroche É,
Rousset M, Vaillant JC, Clément K, Thenet S and Leturque A:
Alteration of intestinal permeability: The missing link between gut
microbiota modifications and inflammation in obesity? Med Sci
(Paris). 32:461–469. 2016.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujino S, Andoh A, Bamba S, Ogawa A, Hata
K, Araki Y, Bamba T and Fujiyama Y: Increased expression of
interleukin 17 in inflammatory bowel disease. Gut. 52:65–70. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Calvano SE, Xiao W, Richards DR, Felciano
RM, Baker HV, Cho RJ, Chen RO, Brownstein BH, Cobb JP, Tschoeke SK,
et al: A network-based analysis of systemic inflammation in humans.
Nature. 437:1032–1037. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cetin S, Ford HR, Sysko LR, Agarwal C,
Wang J, Neal MD, Baty C, Apodaca G and Hackam DJ: Endotoxin
inhibits intestinal epithelial restitution through activation of
Rho-GTPase and increased focal adhesions. J Biol Chem.
279:24592–24600. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maslowski KM, Vieira AT, Ng A, Kranich J,
Sierro F, Yu D, Schilter HC, Rolph MS, Mackay F, Artis D, et al:
Regulation of inflammatory responses by gut microbiota and
chemoattractant receptor GPR43. Nature. 461:1282–1286. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mirmonsef P, Zariffard MR, Gilbert D,
Makinde H, Landay AL and Spear GT: Short-chain fatty acids induce
pro-inflammatory cytokine production alone and in combination with
toll-like receptor ligands. Am J Reprod Immunol. 67:391–400. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Magri G and Cerutti A: A touch of youth in
gut microbiota development. Immunity. 45:12–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bergogne-Bérézin E and Towner KJ:
Acinetobacter spp. as nosocomial pathogens: Microbiological,
clinical, and epidemiological features. Clin Microbiol Rev.
9:148–165. 1996.
|
|
29
|
Sokol H, Leducq V, Aschard H, Pham HP,
Jegou S, Landman C, Cohen D, Liguori G, Bourrier A, Nion-Larmurier
I, et al: Fungal microbiota dysbiosis in IBD. Gut.
pii:gutjnl-2015-310746. 2016.
|
|
30
|
Nones K, Knoch B, Dommels YE, Paturi G,
Butts C, McNabb WC and Roy NC: Multidrug resistance gene deficient
(mdr1a-/-) mice have an altered caecal microbiota that precedes the
onset of intestinal inflammation. J Appl Microbiol. 107:557–566.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kummen M, Holm K, Anmarkrud JA, Nygård S,
Vesterhus M, Høivik ML, Trøseid M, Marschall HU, Schrumpf E, Moum
B, et al: The gut microbial profile in patients with primary
sclerosing cholangitis is distinct from patients with ulcerative
colitis without biliary disease and healthy controls. Gut.
pii:gutjnl-2015-310500. 2016.
|
|
32
|
Gevers D, Kugathasan S, Denson LA,
Vázquez-Baeza Y, van Treuren W, Ren B, Schwager E, Knights D, Song
SJ, Yassour M, et al: The treatment-naive microbiome in new-onset
Crohn's disease. Cell Host Microbe. 15:382–392. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schreiber S, Heinig T, Thiele HG and
Raedler A: Immunoregulatory role of interleukin 10 in patients with
inflammatory bowel disease. Gastroenterology. 108:1434–1444. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Steinbach EC and Plevy SE: The role of
macrophages and dendritic cells in the initiation of inflammation
in IBD. Inflamm Bowel Dis. 20:166–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schoultz I, Söderholm JD and McKay DM: Is
metabolic stress a common denominator in inflammatory bowel
disease? Inflamm Bowel Dis. 17:2008–2018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eeckhaut V, Machiels K, Perrier C, Romero
C, Maes S, Flahou B, Steppe M, Haesebrouck F, Sas B, Ducatelle R,
et al: Butyricicoccus pullicaecorum in inflammatory bowel disease.
Gut. 62:1745–1752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Frank DN, Robertson CE, Hamm CM, Kpadeh Z,
Zhang T, Chen H, Zhu W, Sartor RB, Boedeker EC, Harpaz N, et al:
Disease phenotype and genotype are associated with shifts in
intestinal-associated microbiota in inflammatory bowel diseases.
Inflamm Bowel Dis. 17:179–184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kamada N, Kim YG, Sham HP, Vallance BA,
Puente JL, Martens EC and Núñez G: Regulated virulence controls the
ability of a pathogen to compete with the gut microbiota. Science.
336:1325–1329. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
van Nood E, Vrieze A, Nieuwdorp M, Fuentes
S, Zoetendal EG, de Vos WM, Visser CE, Kuijper EJ, Bartelsman JF,
Tijssen JG, et al: Duodenal infusion of donor feces for recurrent
Clostridium difficile. N Engl J Med. 368:407–415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Meijer BJ and Dieleman LA: Probiotics in
the treatment of human inflammatory bowel diseases: Update 2011. J
Clin Gastroenterol. 45(Suppl): S139–S144. 2011. View Article : Google Scholar : PubMed/NCBI
|